

Abstract
Nitric oxide performs a wide variety of versatile functions in the immune system. The precise role of nitric oxide in pathogenesis of acute inflammation and sepsis is still controversial. A direct physical interaction between inducible nitric oxide synthase (iNOS) and Nitric oxide synthase interacting protein (NOSIP) was observed that regulates Nitric Oxide production by human monocytes. A novel mutation in nitric oxide synthase interacting protein (NOSIP) determines NO levels and influence adverse prognosis and mortality in human sepsis. Differences in production of Nitric oxide by murine and human macrophages could be attributed to differential expression of NOSIP by the two species. The study reveals NOSIP as an important regulator of inflammation by virtue of its ability to influence nitric oxide production which inhibits IL-lβ levels in both mice and in humans and opens up novel avenues for therapeutic strategies against acute inflammation.
Introduction
Nitric oxide (NO) is a well-known modulator of a wide variety of immune functions1. After its discovery, the initial function of NO was shown as a molecule with anti-microbial2,3 and tumoricidal4,5 functions. Later NO was reported to regulate other facets of immune responses like inhibition of T and B lymphocyte proliferation6–10 and leukocyte trafficking11; modulation of cytokine, chemokine and growth factor production12–16; and T helper cell subtype differentiation17–19 etc., The role of NO in regulating hyperinflammtion has been a subject of contemporary interest since it appears to play a duel function of being inflammtotory as well as antiinflammatory in different contexts. Many reports in literature describe links between NO and sepsis. Demonstration of elevated levels of NO by-products in sepsis20 and observations on NOS inhibitors alleviating the hemodynamic manifestations of septic shock21 led to the general belief that NO was detrimental to host during sepsis. This resulted in undertaking a randomized controlled trial that involved treatment of sepsis patients with a nonspecific NOS inhibitor22. However, the trial had to be truncated due to observed higher mortality in the drug treated group over placebo trated control group. On the other hand few reports do suggest a protective role for NO during inflammation and sepsis23,24. Nitric oxide synthase (NOS), the enzyme that produces NO in mammalian cells using L-Arginine as its substrate, has a multitude of interacting partners that regulate its function, localization and trafficking25,26. Of the three NOS isoforms, NOS1 (nNOS) and NOS3 (eNOS) binding proteins have been fairly well characterized25. One such protein is nitric oxide synthase interacting protein (NOSIP), that has been reported to interact with both nNOS27 and eNOS28 and regulate their function. Surprisingly however the role of NOSIP and its interaction with NOS2 (iNOS) and its ability to regulate function of iNOS has not been investigated so far and this study attempts to fill this lacuna through a combination of biochemical and imaging techniques. Using wild type and iNOS deficient mice models of acute inflammation and human sepsis, we validate a protective role for nitric oxide during inflammation and sepsis, an observation reported earlier 23,24. More critically we demonstrate for the first time a novel mutation in a regulatory region of human NOSIP gene and its influence on NOSIP synthesis and association with enhanced mortality in human sepsis. Finally, We provide evidence to suggest that species specific differences in nitric oxide production between murine macrophages and human monocytes could be due to differential expression of NOSIP that determine differential production of NO by the two species.
Materials and methods
Reagents and kits
Gram-negative bacterial lipopolysaccharide (LPS, E.coli O55:B5), Ficoll Histopaque, Blood Genomic DNA extraction kit, Duolink Proximity Ligation Assay (PLA) kit, Nitrite/Nitrate estimation kit, Nω-Nitro-L-arginine methyl ester hydrochloride (L-NAME), Bovine Serum Albumin (BSA), Phorbol 12-myristate 13-acetate (PMA), Paraformaldehyde (PFA) and Triton X-100 was obtained from Sigma chemicals. Dulbecco’s Modified Eagle’s Medium (DMEM), Foetal Bovine Serum (FBS), and Phosphate Buffered Saline (PBS) were from PAN Biotech GmbH. Acetic acid, citric acid, and dextrose were all from Fisher Scientific. Human 27-plex cytokine and Mouse 23-plex cytokine analysis kits were procured from Bio-Rad Laboratories. FACS lysing solution, Lyse/Fix buffer, Permeabilization buffer IV and FACSComp beads were bought from BD Biosciences. Brefeldin A was obtained from eBiosciences. Taq DNA polymerase for PCR was from Thermo Scientific. 4’,6-Diamidino-2-Phenylindole, Dihydrochloride (DAPI) and ProLong® Gold Antifade were purchased from Molecular Probes.
Antibodies
Anti-goat IgG Alexa Fluor 488 and anti-rabbit IgG Alexa Fluor 647 antibodies were purchased from Molecular Probes. Rabbit polyclonal antibody to iNOS was obtained from Abeam and goat polyclonal antibody to NOSIP was purchased from SantaCruz Biotechnolgies.
Human sepsis patients
The study was approved by ethics committees of Institute of Life Sciences and S.C.B. Medical College, and signed informed consent was obtained from all participants. 139 sepsis patients who were admitted to medical intensive care units at the Department of Medicine, S.C.B. Medical College and Hospital (Cuttack, India) were recruited for the study. For sepsis, patients were eligible for inclusion only if they had systemic inflammatory response syndrome and had a source of infection, proven or suspected. The acute physiological and chronic health evaluation II (APACHE II) scoring system was used to categorize the patients. Definitions of sepsis, severe sepsis, septic shock, and multi-organ dysfunction syndrome (MODS) were in accordance with published criteria32,33. The following categories of patients were excluded from the study: patients with diabetes mellitus, hypertension, nephrotic syndrome, chronic kidney disease (sonographic feature of CKD and/or GFR<30 ml/min), patients with cardiac failure and immunocompromised individuals. Blood was collected in vials containing 15% (v/v) Acetate Citrate Dextrose (ACD). This was followed by isolation of plasma by centrifugation at 2000 rpm for 10 min. Plasma was stored in single-use aliquots at -80°C.
Mouse model of endotoxemia
8-10 weeks old male C57BL/6 mice were used for the study. All animal experiments were approved by the institutional animal ethics committee of Institute of Life sciences, Bhubaneswar. To simulate two groups based on lethality in human sepsis cases, a 5mg/kg nonlethal dose and 35mg/kg lethal dose of gram negative bacterial lipopolysaccharide (E.coli O55:B5, Sigma) was injeced intraperitoneally. The animals were sacrificed at 2, 4, 8 and 16 hours post injection to mimic early and late stages of endotoxemia. Blood was collected in vials containing ACD as anticoagulant (15% v/v). collected blood was centrifuged at 2000 rpm for 10 minutes for isolation of plasma. Isolated plasma was stored in 100 μl single-use aliquots at -80°C.
Assessment of nitric oxide deficiency on LPS - mediated inflammation
8-10 weeks old male BALB/C mice were injected with 100 mg/kg L-NAME once every 24 hours for inhibition of nitric oxide synthesis. To examine effect of nitric oxide synthase inhibition on LPS – mediated pathology, untreated or L-NAME treated mice were injected with 2 mg/kg LPS and mortality was monitored for 4 days. For comparing LPS toxicity between wild – type and inos knockout mice, male animals between 8 – 10 weeks age were injected with LPS at 15 mg/kg and either mortality was scored for 5 days or animals were sacrificed at 2, 6 and 12 hours to estimate cytokines in plasma.
Ex vivo stimulation of human and mouse peripheral blood
Whole blood was withdrawn from apparently healthy human donors by venepuncture or from normal healthy mouse by cardiac puncture in acid citrate dextrose (ACD) anticoagulant at 15% v/v. Human donors were recruited from institute students. To test for cytokine expression, fifty microlitre whole blood was left untreated or stimulated with LPS at lμg/ml for 2 hours in 37°C water bath along with brefeldin A (eBiosciences) at 1:1000 dilution. Post stimulation, cells were stained with fluorochrome conjugated antibodies and analysed on a flow cytometer.
Measurement of cytokines in plasma
Human plasma was analysed using the human 27-plex cytokine panel (Bio-Rad) according to manufacturer’s instructions and contained the following targets: IL-lβ, IL-lra, IL-2, IL-4, IL-5, IL-6, IL-7, IL-8, IL-9, IL-10, IL-12(p70), IL-13, IL-15, IL-17, Basic FGF, Eotaxin, G-CSF, GM-CSF, IFN-γ, IP-10, MCP-1, MIP-lα, MIP-lβ, PDGF, RANTES, TNF-α, and VEGF. Mouse plasma was analysed using the mouse 23-plex cytokine panel (Bio-Rad) as specified by the manufacturer and contained the following targets: IL-lα, IL-lβ, IL-2, IL-3, IL-4, IL-5, IL-6, IL-9, IL-10, IL-12(p40), IL-12(p70), IL-13, IL-17, Eotaxin, G-CSF, GM-CSF, IFN-y, KC, MCP-1, MIP-lα, MIP-lβ, RANTES, and TNF-α. All samples were read on a Bioplex 200 system (Bio-Rad). Concentrations of unknown samples were interpolated from a 8-point standard curve fitted with a five-parameter logistic regression model.
Measurement of plasma nitrate and nitrite
Levels of nitrate and nitrite in plasma obtained from human and mice were estimated by a Griess colorimetric assay kit (Sigma chemicals) according to manufacturer’s instructions. Briefly, nitrate in plasma was first converted into nitrite by nitrate reductase. Total nitrite was then estimated by a colorimetric reaction involving Griess reagent A (a solution of sulphanilamide in phosphoric acid) and Griess reagent B (Napthylethylenediamine in phosphoric acid). Absorbance was read at 540 nm. Unknown concentrations were estimated from a standard curve generated with known concentrations of sodium nitrite.
Assessment of intracellular localization of iNOS and NOSIP
Mouse and human whole blood was fixed, lysed and permeabilized followed by staining with anti-iNOS and anti-NOSIP antibodies. Followed by washing, the cells were analyzed on a AMNIS ImagestreamX for intracellular expression and colocalization. Alternately, the cells were fixed on a slide and colocalization was scored on a Leica Laser Scanning Confocal Microscope. For studying interaction between iNOS and NOSIP, in situ Proximity Ligation Assay was used using a commercially available kit (Sigma Duolink) according to manufacturer’s instructions. In case of the human monocytic cell line THP-1, cells were grown in a coverslip-bottomed culture dish in complete IMDM media containing 10% fetal bovine serum. Cells were made to differentiate into macrophages by treatment with 10 nM Phorbol myristate acetate (PMA) for 48 hours. Following adherence, cells were washed with PBS and fixed with 2% paraformaldehyde (PFA). After rinsing with PBS, cells were permeabilized with 0.1% Triton X-100 following which they were incubated in a blocking buffer containing 2% bovine serum albumin (BSA) and 2% fetal bovine serum (FBS) in PBS to minimize nonspecific binding of antibodies. After blocking, cells were first incubated with primary antibody followed by fluorochrome conjugated secondary antibodies. Finally, cells were washed, counterstained with DAPI and image was acquired in a Leica SP5 confocal microscope.
Cloning, Expression and Purification of iNOS and NOSIP proteins
The human iNOS oxygenase domain containing 424 nucleotide sequence and NOSIP gene containing 301 nucleotide sequences were synthesized by GenScript and cloned in pET22b expression vector. The recombinant pET22b: iNOS and pET22b: NOSIP plasmid was used to transform competent E. coli BL21 (DE3) cells. The protein was over expressed using 0.2 mM IPTG at 250C. The harvested cells were resuspended and sonicated in 20 mM Tris-HCI, pH 7.5, 200 mM NaCI, 0.05% Triton X-100 and protease inhibitor (Sigma). The expressed proteins were purified by nickel nitrilotriacetic acid column using 250 mM imidazole.
Physical interaction by Surface Plasmon Resonance
Interaction of purified NOSIP with iNOS protein was monitored using Bio-Rad XPR 36 surface Plasmon resonance biosensor instrument. About 3 μg/ml of iNOS was immobilized on GLC chip by amine coupling method as suggested by manufacturer’s instructions. NOSIP was injected at concentration of 500nM with running buffer composed of PBST and 0.005 % Tween-20 at a flow rate of 50 μl/min. Molecular interaction was carried out at 20 °C. Further kinetic parameter were determined, after fitting the association and dissociation curves to a 1:1 (Langmuir)-binding model.
An activated channel without immobilized ligand was used to evaluate nonspecific binding. The response curves were also recorded on control surfaces. Results were calculated after subtraction of the control values using the ProteOn Manager software.
NOSIP knockdown and functional study
THP-1 cells were grown in complete RPMI medium with 10% FBS. NOSIP shRNA plasmids were obtained from Sigma (MISSION shRNA, cat. No. NM_015953). A cocktail of three different shRNA plasmids was used. Nucleofection was carried out in a Amaxa Nucleofector II machine according to manufacturer’s protocols with slight modifications. Briefly, 5 μg of NOSIP shRNA plasmids or control TRC2 plasmid was mixed with 100 μl Cell Line Nuceleofector Solution V (Lonza) and added to one million cells. For nucleofection, preset program V - 001 was used. Post nucleofection, cells were allowed to grow in complete RPMI media for 48 hours followed by which cells were washed and stimulated with 1 μg/ml LPS for an additional 24 hours. After stimulation, total nitrate + nitrite was estimated in the culture supernatant by Griess assay.
Identification of NOSIP gene polymorphism
Genomic DNA from whole human blood was isolated using a commercially available kit (Sigma) according to manufacturer’s protocol. The NOSIP gene is present on human chromosome number 19 and spans seven exons. A 503bp segment spanning exon 2 was amplified and tested for novel polymorphism in a cohort of 49 sepsis cases. Primer sequences for amplification are: Forward - 5’ TCCCCATATTCCCACCAGTTTC 3’; and Reverse - 5’ GCCGATGCTAGCTACCACTTGA 3’. PCR was performed for 20 μl reactions containing 2 μl genomic DNA, IX PCR buffer containing MgCh2 (Sigma), 250 μM dNTP (Sigma), 10 pM of each of forward and reverse primers and 1U of Taq DNA polymerase (Thermo Scientific). PCR cycling conditions were as follows: an initial denaturation step at 94°C for 2 minutes; followed by 35 cycles of 94°C for 15 seconds, annealing at 56°C for 30 seconds and extension at 72°C for 30 seconds; and a final extension at 72°C for 5 minutes. Amplification was checked by running PCR products on a 2% agarose gel. The amplified PCR products were then sequenced on a Genetic Analyzer 3500 DNA sequencing platform (Applied Biosystems).
RNA extraction and qRT-PCR
Total RNA was extracted from mouse or human whole blood using RNA blood mini kit (Qiagen). Isolated RNA was converted to complementary DNA (cDNA) using RT2 first strand cDNA synthesis kit (Qiagen). The cDNA thus obtained was subjected to quantitative real-time PCR analysis with RT2 SYBR® Green qPCR Mastermix (Qiagen) in 20 μl reaction volumes on a LightCycler 480 thermal cycler (Roche).
qRT-PCR array
Human whole blood was stimulated for 4 hours with LPS at lμg/ml. Following stimulation, total RNA was extracted using a RNA blood mini kit (Qiagen). Total RNA to cDNA conversion was done using RT2 first strand cDNA synthesis kit (Qiagen). Prepared cDNA was subjected to a customised qRT – PCR array (SABiosciences) using RT2 SYBR® Green qPCR Mastermix (Qiagen). Fold change over untreated control was calculated using the 2-xΔΔCt method. The obtained fold change values were used for pathway enrichment analysis (Ingenuity Systems).
Statistical analysis
For comparison between two groups, either an unpaired t-test or the nonparametric Man – Whitney U-test was conducted. One – way analysis of variance (ANOVA) followed by Bonferroni’s multiple comparison test was carried out to compare means between three or more groups. Difference in survival among different groups was estimated by constructing Kaplan – Meier survival curves followed by assessment of significant difference using the log – rank test. Fisher’s exact test was used for comparison of genotype, allele frequencies and to test association of combined genotype distribution among various clinical categories. For all statistical comparisons, a p-value <0.05 was considered statistically significant.
Results
NOSIP interacts with iNOS and modulates nitric oxide synthesis
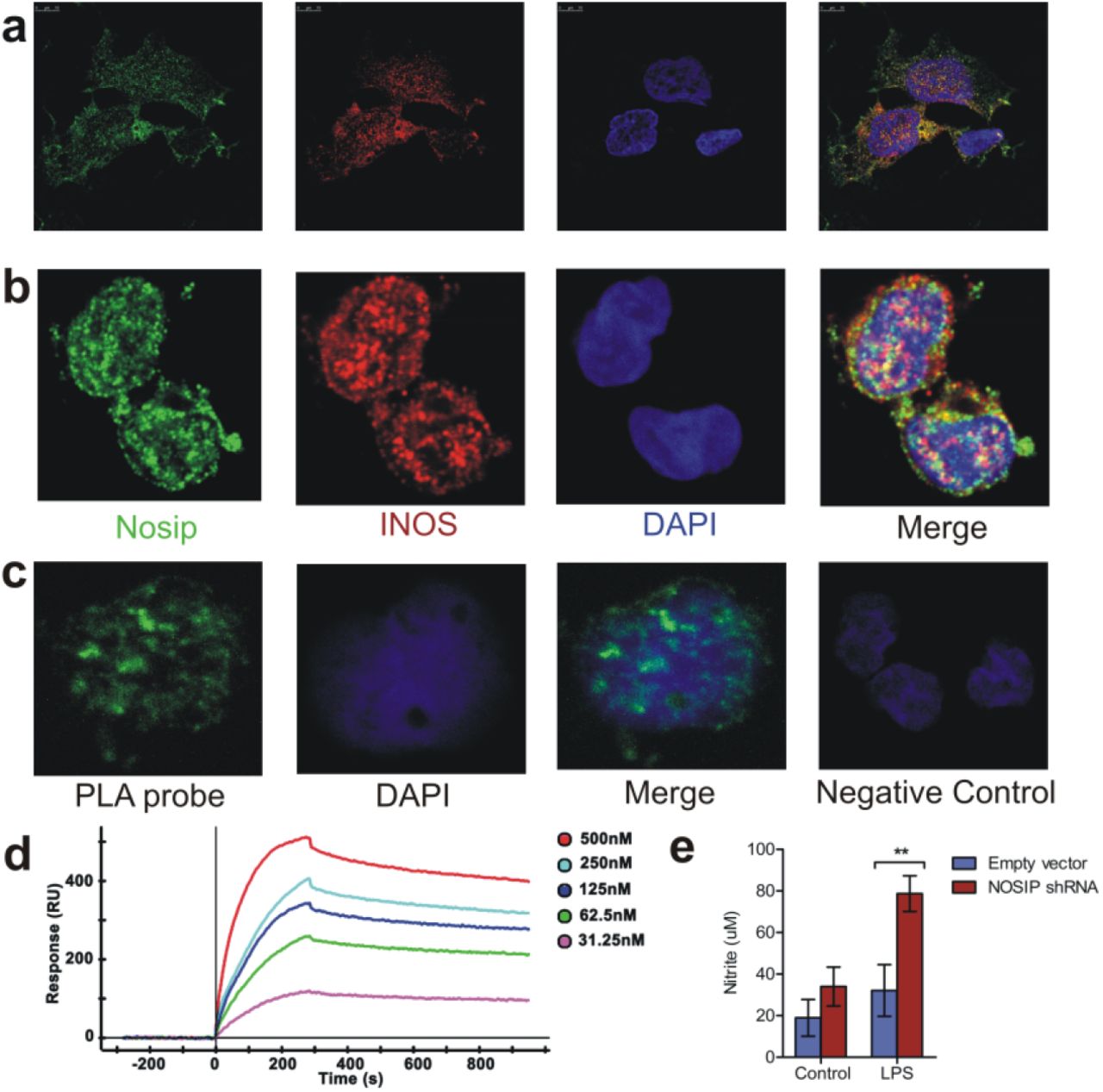
Since binding of NOSIP to eNOS (endothelial NOS, NOS3) and nNOS (neuronal NOS, NOS1) and its inhibition of their function has been reported earleir, the first objective of the current study was to investigate whether NOSIP interacts with iNOS. Co–localization of iNOS and NOSIP in the human monocytic cell line THP-1 and primary human monocytes indicated possible interaction between the two proteins (Figure 1a and b). To confirm the interaction between iNOS and NOSIP proximity ligation assay (PLA) was employed in primary human monocytes – bright fluorescent spots were observed only when antibodies to both NOSIP and iNOS were used and not when either of the two antibodies was used separately, indicating a direct physical interaction between iNOS and NOSIP in primary human monocytes (Figure 1c). Confirmation of interactions between the two molecules in a cell free system was demonstrated by performing surface plasmon resonance (SPR) studies with recombinant iNOS and NOSIP - a KD value of 7.11X10-9 M confirmed significant interation between the two proteins (Figure 1d). The next objective was to test consequence of this interaction on nitric oxide synthesis. To this end, targeted knockdown of NOSIP was performed in THP-1 cell line. Stimulation of THP-1 cell lines with LPS for 24 hours revealed significantly higher levels of nitrite in culture supernatants of cells transfected with NOSIP shRNA as compared to cells transfected with null vector (Figure 1e), suggesting that NOSIP inhibits nitric oxide production likely thro its interaction with iNOS. Taken together, these findings demonstrate that NOSIP interacts with iNOS and modulates synthesis of nitric oxide synthesis.

Cellular localization of iNOS and NOSIP in monocytic cell – line THP – 1 (a) and primary human monocytes (b) showing co - localization of the two proteins, (c) In situ proximity ligation assay demonstrating diresct physical interaction between iNOS and NOSIP in primary human monocytes. Each speckle corresponds to one interacting pair. Negative control cells with only one primary antibody shows no background staining, (d) Dose dependent binding of NOSIP recombinant protein (analyte) to immobilize iNOS solid phase (ligand) as measured by surface plasmon resonance. Recombinant NOSIP protein was injected at concentrations of 500nM, 250nM, 125nM, 62.5nM to 31.25nM over ligand. The KD = 7.11 e-09 M was obtained in Kinetic – Langmuir analysis, (e) THP-1 cells were nucleofected with empty plasmid or NOSIP shRNA plasmid by Amaxa nucleofector II using manufacturer’s kit. After 48 hours, the cells were washed and incubated with or without LPS at lμg/ml for a further 24 hours. Total nitrate+nitrite in culture supernatant was measured by a commercial Griess assay kit. Statistical significance was assessed by two - way ANOVA following Bonferroni’s post test (** p<0.01). The graph represents data from 5 separate plates performed on 2 different days.
Mutation in NOSiP gene leads to increased protein expression and is associated with increased mortality in patients with sepsis
Since NOSIP regulates NO production we searched for possible polymorphism in NOSIP gene that may have functional consequences in inflammatory responses in humans. Multiple sequence alignment of NOSIP protein sequence revealed remarkable conservation between different species ranging from zebrafish to humans (Supplementary Figure 1). Five different sets of primers each amplifying different regions of nosip gene were designed and the amplified products were sequenced. Sequence matching of 59 DNA samples isolated from healthy volunteers with that of the database revealed presence of a four nucleotide deletion (TCTC) downstream of exon two in about 71% (42/59) of the subjects (Table 1). Next we addressed the issue whether the polymorphism influences NOSIP expression – intracellular NOSIP levels between ‘insertion’ and ‘deletion’ individuals was scored in circulating leucocytes of healthy human volunteers. Imaging cytometry analysis revealed three distinct cell populations based on expression of NOSIP and side scatter signal (Figure 2b). Comparison of intracellular NOSIP expression in all three populations revelaed increased expression in individuals with deletion polymorphism as compared to those with Insertion (Figure 2 e-g). Significance of NOSIP mutation on status and expression of 84 genes involved in host inflammatory and other responses were studied. Whole blood of normal human volunteers were stimulated with LPS for 4 hours and gene expression was compared between ins or del subjects by a custom designed qRT-PCR array (Fig 3a) – very broadly, expression of many of the cytokine and chemokine genes in subjects with del/del genotype were higher in comparison to those with Ins/Ins genotype (Cluster 4, Figure 3a). In order to gain insights into global differences in signalling pathways as a consequence of differential NOSIP expression, a pathway enrichment analysis was performed. The analysis revealed increased enrichment of pathways associated with inflammatory response, viz. Toll-like receptor signalling, NF-kB signalling, p38 MAP kinase signalling and HMGB1 signalling in individuals with del allele (Figure 3b). A similar enrichment analysis conducted for disease associated networks revealed higher enrichment of sepsis, septic shock network in Del individuals. Higher representation of networks associated with liver and kidney damage, apoptosis of liver and kidney cells, and apoptosis of macrophages, phagocytes and antigen presenting cells were also observed suggesting an increased risk of sepsis associated pathology in individuals with increased expression of NOSIP (Figure 3c). Visual comparison of septic shock associated networks between Ins/Ins (Figure 3d) and Del/Del (Figure 3e) individuals confirmed this. Therefore, an analysis of NOSIP polymorphic genotype distribution was performed in a cohort of patients with sepsis. Although there was no significant difference in distribution of NOSIP mutation between controls and sepsis patients (data not shown), the results revealed significantly higher mortality among sepsis patients displaying homozygous deletion polymorphism. Significantly high odds ratio (5/8 deaths, P-value = 0.0159, OR = 10.56, 95% C.l. = 1.611 to 69.15) indicated that patients with homozygous deletion mutation are highly prone to mortality in the cohort of sepsis patients studied (Table 2).
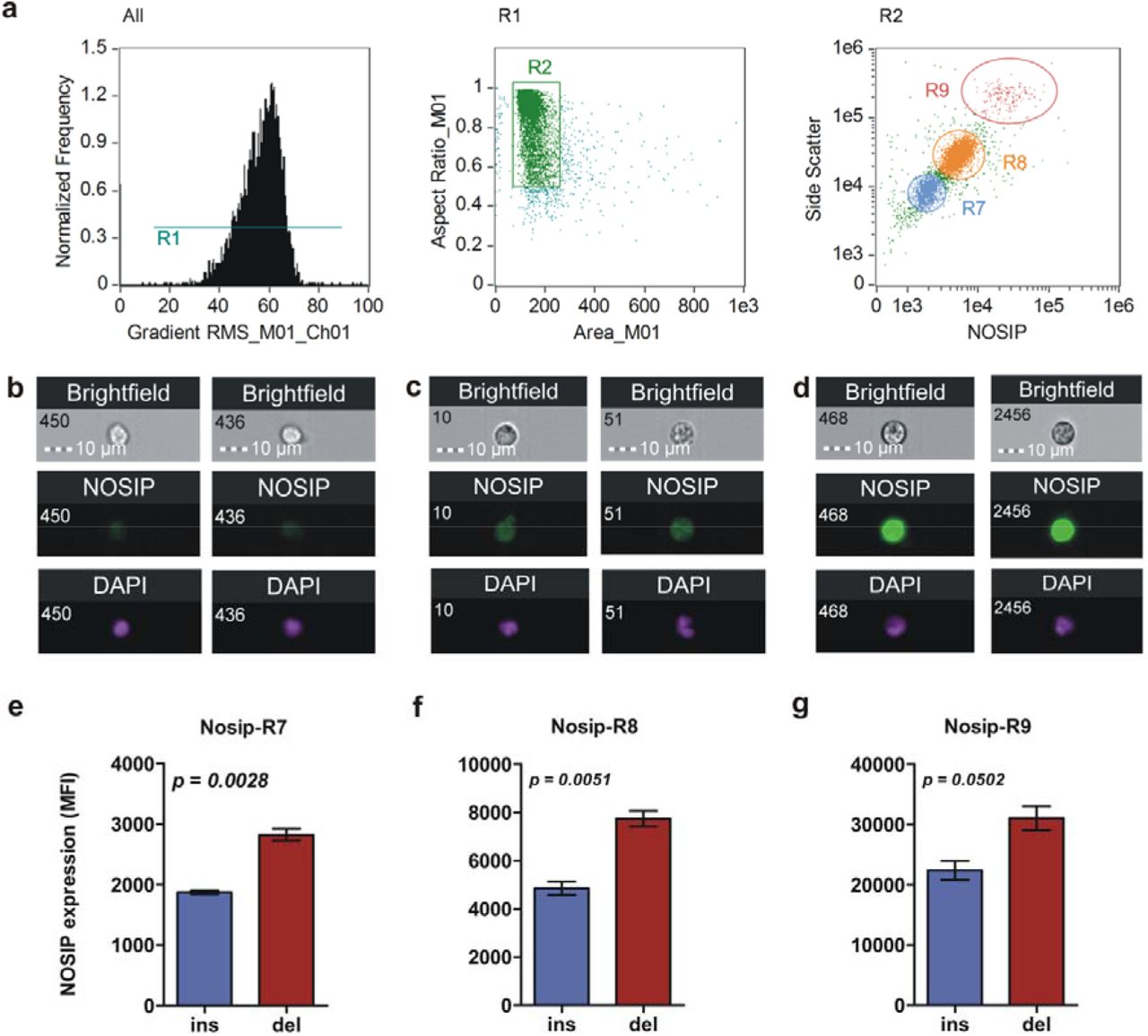
(a) Imaging cytometry analysis of intracellular NOSIP expression. In the first plot, the histogram depicts selective removal of ‘out – of – focus’ cells. In the next plot, only single cells are selected for further analysis (R2). Finally, they were discriminated on a bivariate scatter plot based on differences in side scatter signal and intracellular NOSIP expression (regions R7, R8 and R9). (b – d) Representative imgaes showing cell and nuclear morphology along with intracellular NOSIP expression of populations R7 (b), R8 (c) and R9 (d). (e – g) Graphs showing significantly elevated levels of NOSIP protein as a consequence of presence of deletion allele as assessed by unpaired t – test (n = 4 for insand n = 3 for del category).

Whole blood was collected from two healthy individuals each of ins/ins and del/del genotypes and stimulated with LPS at 1 μg/ml for 4 hours. After stimulation, total RNA was extracted, cDNA synthesized and subjected to a custom designed RT2 q-PCR array. Raw Cp values were exported in Qiagen’s online tool and fold increase was calculated after normalizing with β-actin as housekeeping gene, (a) Clustering heatmap depicting differential expression of 85 immune response genes between ins/ins and del/del individuals, (b) Differences in pathway enrichment between ins/ins and del/del individuals as assessed by Ingenuity Pathway Analysis, (c) Differential enrichment of disease - associated networks between ins/ins and del/del individuals, (d and e) Representative networks associated with septic shock response between ins/ins (d) and del/del (e) individuals.
Genotype and allele distribution of NOSIP (TCTC ins/del) polymorphism in healthy individuals
Genotype and allele distribution of NOSIP (TCTC ins/del) polymorphism and its association with mortality in sepsis
In summary, above data provides evidence for existance of a novel polymorphism in NOSIP gene that leads to higher intracellular expression of NOSIP, which in turn results in increased risk for sepsis associated mortality.
Plasma nitrite levels are inversely associated with inflammatory cytokines in human sepsis and experimental murine endotoxemia
The observation that increased NOSIP leads to decreased NO synthesis may be harmful to host during acute inflammation led to us to investigate the role played by NO during acute inflammatory disorders. In order to gain insight into the role of nitric oxide in acute inflammation, a multivariate correlation analysis of plasma cytokines and nitrite levels was conducted in human sepsis patients.
A significant negative association was observed between plasma nitrite and some of the cytokines/chemokines (Table 3) suggesting a possible protective role for increased plasma nitrate/nitrite levels in human sepsis. To test this hypothesis experimentally, male C5BI/6 mice were injected with two different doses of LPS and plasma cytokines and nitrite levels were estimated at 2, 4, 8 and 16 hours post injection. Plasma nitrite levels negatively correlated with inflammatory mediators in mouse model of experimental endotoxemia also (Table 3). Taken together these observations indicate a protective role for nitric oxide in pathogenesis of endotoxemia/ sepsis.
Inverse association of plasma nitrite with inflammatory factors in a mouse model of endotoxemia and human sepsis
Deficiency in nitric oxide synthesis increases susceptibility to acute inflammation by increased IL-lβ production
To investigate the effect of nitric oxide deficiency on LPS induced inflammation, male BALB/C mice were treated with 100 mg/kg of Nω-Nitro-L-arginine methyl ester (L-NAME) hydrochloride once every 24 hours to block nitric oxide synthase activity. L-NAME is a structural analogue of L-Arginine that competitively inhibits all three isoforms of nitric oxide synthase. Untreated and L-NAME treated mice were then injected with 2 mg/kg of LPS and mortality was assessed for 96 hours. Figure 4a shows comparison in LPS toxicity between untreated and L-NAME treated animals clearly demonstrating that inhibition of nitric oxide synthesis is detrimental to host in a mouse model of endotoxemia. This was further tested by studying LPS toxicity in mice deficient in Nos2 gene. Comparison of lethal dose of LPS between wild type C57BL/6 mice and Nos2 null revealed that 15mg/kg of LPS did not result in mortality of wild type mice, while about 84% of the animals in the Nos2-/- group died at the same dose of LPS (Figure 4b). This confirmed the hypothesis that absence of nitric oxide inducing enzyme and consequently nitric oxide results in increased susceptibility to endotoxemia. To test for possible mechanisms of the observed phenomenon, wild type and Nos2-/- mice were injected with LPS at a dose of 15mg/kg and the animals were sacrificed at 2, 6 and 12 hours post administration and plasma cytokines were quantified. The results shown in Figure 4c reveals significantly higher levels of interleukin-lβ (IL-lβ) in mice lacking the Nos2 gene. However, plasma levels of TNF-α were comparable in both (Figure 4c), suggesting that IL-lβ could be playing a more central role in poor disease outcome in endotoxemia/sepsis and that NO is a key molecule regulating NLRP3 mediated inflammasome pathway. Very similar observations on the role of IL-lβ in endotoxemia has been reported by earlier42. Interestingly, increased production of interleukin-10 (IL-10) was also observed in the Nos2 null mice (Figure 4c), which we interpret as a compensatory mechanism of the host triggered in the absence of nitric oxide leading to over-production of IL-10.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
(a) Treatment of BALB/C mice with a nonselective NOS inhibitor renders the animals more susceptible to LPS – mediated endotoxic shock in comparison to untreated mice. L-NAME was injected at 100 mg/kg once every 24 hours. LPS was injected at 2 mg/kg body weight, (b) iNOS-/- C57BL/6 mice and their wild-type littermates were injected with 15 mg/kg LPS and mortality was scored for 120 hours, (c) The animals (n = 4 in each group) were weighed and LPS was injected at 15 mg/kg intraperitoneally. For assessment of cytokine production, animals were sacrificed at the indicated time points and isolated plasma was subjected to a multiplex cytokine assay. Significance levels were assessed by two-way ANOVA with Bonferroni’s multiple comparison test. (*p<0.05, **p<0.01) (d – e) Mouse (n = 4) and human (n = 5) whole blood were left untreated or treated with 1 μg/ml LPS for 2 hours along with Brefeldin A. Following stimulation, cells were fixed, permeabilized, washed and stained with flurochrome-conjugated antibodies for analysis on a flow cytometer. Only monocytes were analysed. Alternately, mouse (n = 5) and human (n = 8) whole blood was left untreated or treated with 1 μg/ml LPS for 4 hours and IL-lβ mRNA expression was scored by qRT – PCR. (f) Intracellular protein and mRNA expression of iNOS in human and mouse circulating monocytes showing no difference between the two species as assessed by unpaired t – test, (g) Intracellular protein and mRNA expression of NOSIP in human and mouse circulating monocytes showing higher expression in humans as assessed by unpaired t – test. For both f and g, n = 6 for both mouse and human (intracellular protein); n = 4 (mouse) and n = 5 (humans) for mRNA expression, which is depicted as fold change upon LPS stimulation over untreated control. *** p<0.0001.
Taken together, the above findings demonstrate that nitric oxide protects against experimental endotoxemia by decreasing IL-lβ levels.
NOSIP levels determine variable NO production by murine and human mononuclear cells
Following the observations that NOSIP regulates NO production we reasoned that it could be contributing to differences observed between species in producing NO – mouse macropages are known to be aggressive producers of NO while human monocytes/macrophages do not release significant levels of NO when stimulated with TLR agonists. To test whether NOSIP levels contribute to differences observed in NO production, human and mouse whole blood were stimulated with LPS and intracellular IL-lβ and TNF-α were measured by flow cytometry – the assumption here was that lower nitric oxide production in humans would lead to increased IL-lβ synthesis since nitric oxide is know to regulate IL-lβ. Figure 4d shows that this is indeed the scenario. Human peripheral blood monocytes produced significantly higher IL-lβ as compared to monocytes of mice upon stimulation with LPS; TNF-α levels however were comparable between the two species (Figure 4e). A comparison of intracellular iNOS and NOSIP expression between circulating monocytes of human and mouse revealed comparable levels of iNOS (Figure 4f) but significantly higher levels of NOSIP in human monocytes (Figure 4g), suggesting that low NO production by human monocytes could be due to increased inhibition of iNOS activity by NOSIP. A search of existing literature indicates an association between plasma nitrite and susceptibility to acute inflammation mediated by LPS. Data on susceptibility to LPS were either obtained from the present study (C57BL/6 mice), or from published literature. An inverse relationship between plasma nitrite levels and lethal dose to LPS was observed (Table 4), suggesting that higher a species’ ability for nitrite production, higher is it’s ability to resist pathology mediated by endotoxin. These observations offer credence to the view that differential synthesis of nitric oxide (as a consequence of differences in NOSIP expression) may be regulating sensitivity to LPS mediated inflammation in endotoxemia/sepsis.
Association between nitrite production and susceptibility to LPS
Discussion
Nitric oxide (NO) is synthesised by many cell types during host response to pathogens and injury35. The principal enzyme responsible for nitric oxide production in immune cells during inflammation is inducible nitric oxide synthase (iNOS or NOS2). Role of nitric oxide in endotoxemia/sepsis has been very contractory in reported literature. Earlier studies suggested a detrimental role for nitroic oxide and as a consequence inhibitors of iNOS were tested for treatment of endotoxemia/sepsis. Although pre-clinical trials offered promising results a human trial conducted with iNOS inhibitor, aminoguanidine had to be truncated since prognosis was better in patients treated with placebo22. Viewed from the observations of the current study which offers evidence for an anti-inflammatory role for Nitric Oxide, failure of clinical trial with iNOS inhibitor is not surprising. A recent study by further adds credence to protective role of Nitric oxide in sepsis. Therapeutic potency of transfer of ex vivo expanded fibroblastic reticular cells (FRCs) into mice with endotoxemia or cecal-ligation- and-puncture induced sepsis, was found to be dependent on iNOS activity expressed by FRCs36. In a similar sepsis model, iNOS-dependent upregulation of cGMP and subsequent activation of TACE was found to protect against organ injury37. Other studies in animal models have also documented possible protective role of NO in endotoxemia38.
The current manuscript presents several novel findings on modulation of iNOS activity and nitric oxide biology in the context of host response by Nitric oxide synthase interacting protein (NOSIP). Several years ago NOSIP was discovered to interact with eNOS and inhibit its function by sequestering it in Golgi complex28. Subsequently, interaction of NOSIP with nNOS was also reported27. Curiously however, till date the role of NOSIP and its with iNOS and regulation of nitric oxide production by immune cells has not been investigated – the present study fills this lacuna. Physical interaction between NOSIP with iNOS were demonstrable in immune cells as well as in cell free systems using recombinant proteins. More significantly, NOSIP emerged as a key molecule regulating iNOS function and release of nitric oxide. More critically a four nucleotide deletion upstream of first exon of NOSiP gene was found to be associated with increased intracellular expression of NOSIP and higher risk of mortality due to sepsis. This observation has opened up an entirely new area of therapeutic strategy against sepsis. Structural biology of iNOS-NOSIP interaction can be expected to lead to discovery of small molecules for blocking their interactions which could modulate nitrci oxide levels. More significantly such small molecules could also be useful in regulating nitric oxide production by nNOS and eNOS in neuronal and endothelial cells respectively.
An inverse association of plasma nitrite with inflammatory cytokines was observed in human sepsis and mouse model of endotoxemia, suggesting a reciprocal regulation of nitric oxide and inflammatory mediators. Species-specific differences in nitric oxide synthesis have been well documented in literature. A major issue of debate in literature has been differences in NO production among species, most notably between human and mouse29,30. It has been reported earlier that human monocytes and macrophages produce significantly lower amounts of NO than their mouse counterparts31. Whether such differences translate into differences in susceptibility to acute inflammation remains unknown is one of the key questions addressed in the present study. The results show a clear positive association between nitrite oxide production and lethal dose of LPS, suggesting that a given species’ ability to tolerate inflammatory insult is, at least in part, dictated by its ability to produce nitric oxide. This conclusion may have important bearing on a study published earlier that demonstrated that proteins in serum rather than intrinsic cellular differences may play a role in regulating variations in resilience to microbe-associated molecular patterns between species34. It apparent that identification of such proteins may open up new areas of targeted therapy. However, given the large number of proteins in serum, this is an arduous task. The observations made in the current study could help in narrowing down the search by initially probing enzymes involved in nitric oxide biosynthesis.
Another important finding of the present investigation was demonstration of increased IL-lβ production in mice lacking a functional iNOS gene (during inflammatory activation), suggesting that protective effect of nitric oxide in endotoxemia is mediated through inhibition of IL-lβ synthesis - more so since TNF-α levels remained comparable in wild type and iNOS deficient mice. This is particularly interesting especially in the light of an earlier study that demonstrated no difference in LPS toxicity between wild type and TNF-α knockout mouse39. On the other hand, mice deficient in IL-lβ converting enzyme were relatively resistant in comparison to wild type mice when challenged intraperitoneally with endotoxin40. These observations suggest a central role played by IL-lβ (inflammasome pathway) in pathogenesis in endotoxemia/sepsis. Similar to nitric oxide, species specific dichotomy in terms of IL-lβ production was observed between human and mouse circulating immune cells that negatively correlated with nitric oxide production, further demonstrating IL-lβ as a central pathogenic hub in endotoxemia.
Despite advances in both understanding of basic biology as well as better critical care support, mortality due to sepsis remains unacceptably high. As of now, no effective therapy exists to combat sepsis. Given the protective role played by nitric oxide in sepsis, it is tempting to suggest administration of nitric oxide donors for clinical management of sepsis. Indeed, several studies have attempted NO supplementation in sepsis, and a systematic review and meta-analysis of such studies showed that this line of therapy could be promising41. However, because of the extremely short half-life of nitric oxide, NO donors require administration on a regular basis, thus potentially raising the cost of treatment. In this context, the identification of NOSIP as a key regulator of NO synthesis immediately opens up exciting possibilities for developing targeted therapy. Designing inhibitors of NOSIP could lead to increased NO production which, in turn, could control hyperinflammation observed in sepsis. However, owing to the novelty of NOSIP, extreme caution should be exercised before designing such therapies as the overall function of NOSIP in other cellular processes need to be evaluated first, particularly their effect on eNOS and nNOS mediated biological activities. From an evolutionary perspective, it would be interesting to investigate basis of such discrepancies in NOSIP expression between species, particularly since increased NOSIP appears to render a host more susceptible to acute inflammation associated with endotoxemia.
Acknowledgements
The authors thank Dr. Dileep Vasudevan, Institute of Life Sciences for his assistance in cloning and purification of iNOS and NOSIP proteins and Dr. Narottam Acharya, Institute of Life Sciences for his assistance in interaction studies by Surface Plasmon Resonance. The Institute of Life Sciences is funded by grants from Department of Biotechnology, Govt, of India. R.M. is a recipient of senior research fellowship from Indian Council of Medical Research.
References




