

Abstract
The convoluted architecture of dendritic arbors poses a challenge to understanding age-dependent alterations and regeneration following injury. Here, we show that induction of cellular fusogens can remodel and facilitate regeneration of dendrites in polymodal PVD neurons of aging Caenorhabditis elegans. Using whole-animal live imaging, we find that the PVD dendritic trees, composed of repetitive "menorah" units, show age-dependent hyperbranching, disorganization, and loss of self-avoidance. These processes, while independent of canonical lifespan-regulating pathways, can be partially rescued by ectopic expression of the fusogen EFF-1. Furthermore, the decreased capacity of old animals to repair laser-induced severed dendrites via auto-fusion can be restored by reducing DAF-2 (Insulin/IGF-1Receptor) function or by ectopic expression of the EFF-1 paralog AFF-1. Our findings demonstrate that fusogens are sufficient to maintain the dendritic arbor structure and increase its regeneration potential in aging animals. These antiaging strategies can be potentially applied to other organisms to protect them from neurodegeneration.
Introduction
Aging is the primary risk for neuronal diseases and general cognitive decline in humans 1, yet, our understanding of the process of neuronal aging at the molecular and cell biological levels is still limited. In particular, very few studies have investigated the fate of complex dendritic arbors during aging, and their regenerative capacity following injury. C. elegans is a powerful system to study the genetics of neuronal aging and regeneration 2–4 Signaling via the DAF-2 Insulin/IGF-1 receptor is the most prominent and conserved pathway that controls aging and longevity of C. elegans, flies, and mammals 2. Normally, when the DAF-2/IGF-1 receptor is activated it induces a conserved PI3K/AKT kinase cascade, which in turn inhibits the DAF-16/FOXO transcription factor from entering into the nucleus. Reduction of daf-2 function doubles life span, and long-lived worms are considered to stay healthy for longer 2,5. In these animals, DAF-16/FOXO affects transcription of different genes that encode heat-shock proteins, antimicrobial proteins, antioxidants, and other molecules, which leads ultimately to extended lifespan 6,7 Recently, aging-associated axonal morphological alterations and decline in regenerative capacity were described in C. elegans. daf-2 mutations delayed these age-related morphological changes and improved regeneration of aged severed axons in a daf-16-dependent manner 8–11. Moreover, in the case of GABA motor neurons, as adult animals age there is a reduction in axon growth, retraction and regrowth in response to injury. Surprisingly, the decline in regeneration is controlled by daf-16 in a cell-autonomously fashion and independently of lifespan 11.
Different invertebrates, including nematodes and crustaceans, use membrane fusion as an alternative mechanism for repair of injured axons 12–15. Cell fusion events were also observed in the brains of mammals both spontaneously and as a result of injury such as stroke 16–18, but the role of these events has remained unclear. In C. elegans, axonal regeneration via auto-fusion is mediated by the fusogen EFF-1 12,13,15. EFF-1 is the first bona fide eukaryotic developmental cell-cell fusion protein. It is expressed in different cell types including neurons, and mediates fusion between cells by a homotypic mechanism 19–21.
We have been studying the role of fusion proteins in the PVD neuron, a polymodal nociceptor that senses harsh touch to the body, cold temperature, and proprioception 22–25. The PVD neuron exhibits an elaborate and invariant network of dendrites, which is composed of a repetitive unit that is termed "menorah" (Figure 1A) 23. Menorahs arise and are maintained through several intrinsic and extrinsic genetic pathways 23–33.

(A-D) PVD in wild-type (wt; A and C) and daf-2 mutants (B and D) at L4 stage and 9 days of adulthood (9d). Upper panels, magnified boxed areas with “menorah” colored according to the scheme (A). c, cell body. Scale bars, 50 μm and 20 μm in magnified images. Anterior is left and ventral is down in all the figures. Filled arrows, self-avoidance defects. Empty arrows, disorganized menorahs. (E-I) Quantitation of phenotypes shown in images. Percentages are for 100 μm of length around cell body. Error bars, ± s.e.m. p values from t tests: *p<0.01, **p<0.001. Number of animals analyzed: daf-2 n ≥ 8 and wild-types n ≥ 5.
The PVD develops in a stereotypic fashion from the L2 larval stage to the young adult 23. The fusion protein EFF-1 mediates dendrite retraction and auto-fusion of excess branches by a novel cell-autonomous pruning mechanism 23. Thus, this fusogen maintains the menorah structure by trimming excess branching during normal developmental arborization 23. AFF-1, a paralog of EFF-1, is a second C. elegans fusogen displaying a more restricted tissue distribution pattern 34; AFF-1 was not found to be involved in PVD remodeling during development 35. AFF-1 and EFF-1 fusion proteins also auto-fuse epithelial and myoepithelial cells to form tubes and reshape glial cells 36–38. Moreover, it has recently been demonstrated that in vertebrates auto-fusion takes place in the development of the vascular endothelium, where it leads to pruning of excess blood vessels 39 in a process that remarkably resembles EFF-1-mediated PVD pruning 23. It is unknown whether PVD structure and function is affected by aging and by the activity of fusion proteins.
Here, we use the PVD polymodal neuron as a model to study how aging affects the morphology, function, and regeneration of dendrites following injury. Our study demonstrates the age-related progressive morphological alterations of intricate dendritic arbors in a living organism. We found that the fusogen EFF-1, when expressed in the PVD neuron, simplifies and therefore rejuvenates aged dendritic trees. In contrast, insulin/IGF-1 receptor mutations (daf-2) fail to inhibit the progressive aging of dendrites and do not prevent the decline in response to harsh touch during aging. We also discovered that PVD aging is characterized by a decline in regenerative potential of dendrites following experimental laser dendrotomy. Furthermore, the regeneration of transected dendritic trees can be restored in old animals by DAF-2 insulin/IGF-1 receptor mutations, and can be differentially reestablished by ectopic AFF-1 fusogen expression. Thus, fusogens ectopically expressed in the PVD and mutations in DAF-2/IGF-1R, differentially rejuvenate some aspects of dendritic architecture and regeneration potential in aging C. elegans.
Results
Progressive dendritic remodeling during aging is independent from the Insulin/IGF-1 pathway
To determine how aging affects the complex arborized dendritic structure, we analyzed the C. elegans PVD dendritic branching patterns from the fourth larval stage (L4) to ten-day-old adults. We found that during aging PVD’s dendrites and menorah structure undergo disorganization and hyperbranching, which is particularly evident in regions closer to the cell body (Figure 1A and 1C). Remarkably, we found that these age-dependent morphological changes of the PVD dendritic pattern were not affected in long-lived animals carrying a mutation in daf-2 (Figure 1A-1H, and Figure 1-figure supplement 1). Adjacent menorahs normally avoid each other 40, and we found that as the animal ages menorahs lose their selfavoidance properties (Figure 1C, 1D, and 1I). Consistently, this age-dependent dendritic pattern did not improve in daf-2 mutant animals; young daf-2 animals at the L4 stage exhibited significantly more daf-16-dependent self-avoidance deficiencies in comparison to wild-type (Figure 2). Thus, taken together our results reveal that aging causes a daf-2-independent increase in disorganized branching and loss of menorah self-avoidance.
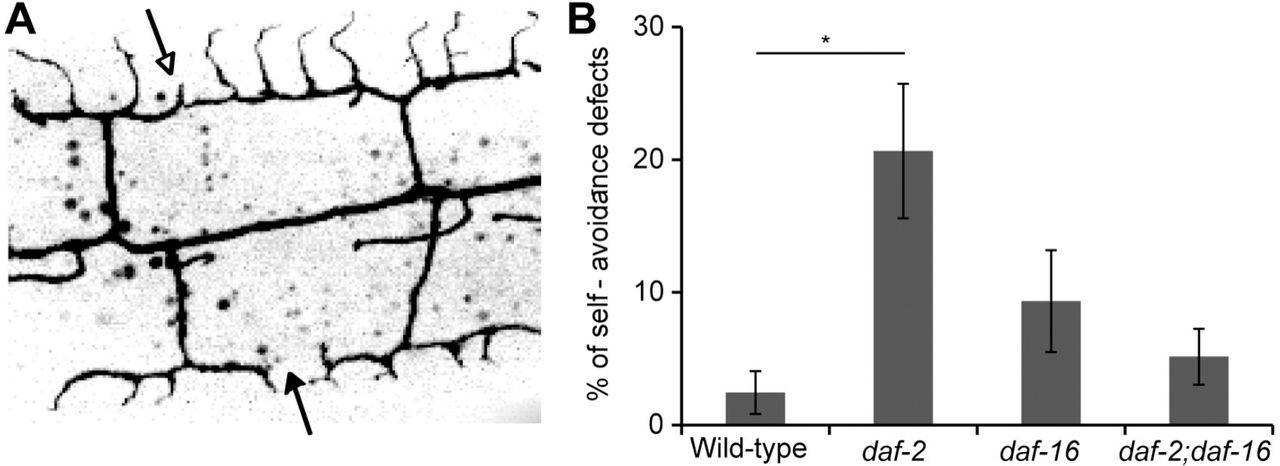
(A) Image showing four wild-type menorahs labeled with GFP, two of them do not overlap (filled arrow) and two show defective self-avoidance (empty arrow).
(B)Percentage of defects in self-avoidance in 100 μm of length around PVD cell body at L4 stage in different genotypes, n ≥ 7. Defective self-avoidance increased in daf-2 mutants in a daf-16-dependent manner. Error bars are ± s.e.m. p value from t test: * p<0.05.
To further understand the nature of the age-dependent dendritic tree remodeling, we followed individual animals over time and analyzed time-lapse movies. We found that 5-day adults still exhibited some plasticity of the dendritic tree, with dynamic growth and retraction events (Figure 3A] and Movie S1); however, both growth and retraction were approximately two times slower (Figure 3B) compared to younger L4 animals. To shed more light on the link between structure and function of dendrites during aging, we performed a functionality test. PVD responds to harsh stimulations, and we used a classic harsh touch assay in mec-4 mutant animals to specifically test PVD activity without the background light touch response mediated by the six light-touch mechanosensory neurons 23,41. First, we found that although mec-4 animals have shorter life spans 10, their dendrites looked similar to wild-type between the ages L4-5d (Figure 4), which further demonstrates that the morphological alterations we see are lifespan-independent. Second, we found that the PVD functionality decreased with aging, with 5d adults presenting a reduced harsh touch response in comparison to 1d adults, in both mec-4 as well as mec-4;daf-2 double mutants (Figure 4F). The specific components in the PVD circuit affected in 5d adults have not been identified and it is possible that multiple components of the sensorimotor circuit contribute to the age-dependent decline in PVD activities. Thus, our results reveal that the morphological and behavioral hallmarks of aging in PVD dendritic arbors are independent from the canonical IGF-1 pathway that affects lifespan.

(A) Time-lapse confocal projections of 5 days of adulthood (5d) wild-type animal. Boxed areas are enlarged in the lower panel and reveal dynamic growth (arrows) and retraction (arrow heads) of branches. Scale bars, 10 μm.
(B)Growth and retraction rates in μm/minute of branches at L4 stage and 5d as measured from time-lapse movies. Number of branches analyzed ≥ 18 from 4 L4 animals and 2 5d animals. Error bars are ± s.e.m. p values from t tests: *p<0.01, **p<0.001. See also Movie S1.

(A-D)PVD’s cell-body region of wt and light-touch insensitive mec-4 mutants at the L4 stage and at 5 days of adulthood (5d). Scale bars are 20 μm.
(E)Total number of branches counted in 100 μm of length around cell-body in wt and mec-4 mutants at L4 and 5d (no significant differences between wt and mec-4). Error bars are ± s.e.m.
(F)Percentage of animals responding to harsh touch by escaping away from the stimulus. At the age of 5d, the response declined in mec-4 mutants and in daf-2;mec-4 double mutants. p values from Χ2 test as compared between 1d to other ages (3d or 5d) for each genotype: * p <0.05. Number of animals: mec-3 ≥ 50; mec-4 ≥ 70; daf-2;mec-4 ≥ 40; wt ≥ 26.
Age-dependent dendrites remodeling can be modulated by EFF-1
The fusogen EFF-1 is essential to fuse some injured axons in C. elegans 12,15, and is involved in PVD’s dendrite pruning in a cell-autonomous and dosage-dependent manner during development and in young adults 23. When EFF-1 is overexpressed in the PVD, a strong gradient of arborization is seen in L4s and young adults, with almost complete lack of branches in areas that are distal from the cell body. However, in areas around the cell body the PVD menorahs appear similar to those in the wild-type 23 (Figure 5A and 5B). Since EFF-1 retracts and simplifies dendritic arbors in young animals 23 and is expressed in the PVD throughout adulthood (Figure 5-figure supplement 1), we hypothesized that EFF-1 overexpression will be able to retract dendrites in aged adults. Indeed we found that EFF-1 overexpression in the PVDs simplified the hyperbranching around the cell body at 9-10 days adults (Figure 5C-5E); in particular, the quaternary branch order was decreased (Figure 5F). A trend of reduction in fifth and higher order branches also appeared in aged animals overexpressing EFF-1 (Figure 5G). Thus, overexpression of the fusogen EFF-1 in the PVD neuron is sufficient to simplify aged menorahs.

(A-D)Inverted fluorescence images of PVD neurons.
(A and C)Represent wild-type neurons from L4 and 9 days of adulthood (9d).
(B and D)Represent EFF-1 overexpression under PVD specific promoter (PVDp) at L4 and 9d. In each panel one menorah (boxed) is enlarged and colored (see Figure 1A). Scale bars, 20 μm and 10 μm in the enlarged images.
(E-G)Graphs showing number of branches in 100 μm of length around cell body. Error bars, ± s.e.m. p values from t tests: * p<0.05, ** p<0.001, *** p<0.0001 Number of animals: n ≥ 4. PVDp::EFF-1 line 1 and 2 correspond to worms carrying the extrachromosomal arrays hyEx392 and hyEx23, respectively.
Age-dependent decline in dendrite regeneration is dependent on Insulin/IGF-1
Mammalian axons regenerate better in younger individuals than in adults 42. Similarly, axonal regenerative ability declines drastically as nematodes advance through development and age 11,43. Our knowledge is still poor on the regenerative capacity of the dendrite, and how aging affects this process of neuronal repair. To study dendritic regeneration we severed the primary dendrites of PVD neuron in aging adults. Typically, the PVDs show robust regeneration at L4 larval stage, consisting in dendrite sprouting from the proximal fragment still attached to the cell body and reconnection via fusion with the separated distal dendrite fragment (Figure 6A, Movie S2). To directly measure reconnection by auto-fusion, we used the photoconvertible reporter Kaede (Figure 6-figure supplement 1 and Movie S3). If fusion fails to occur, the detached distal part eventually degenerates. We found that 2-3 day-old wild-type adults respond more slowly to laser dendrotomy in comparison to L4 and young adults (~70% of the young animals presented regeneration, whereas at the age of 2-3d neither regeneration nor degeneration occurred within 3-6 hours (Figure 6E and Movie S4). At the age of 5 days, the ability to regenerate by dendrite auto-fusion was almost completely lost (Figure 6C and 6F). Remarkably, long-lived daf-2 mutants showed similar regeneration to wild-type at L4 stage (Figure 6B and 6F), whereas at older age (5d) daf-2 mutants had a much higher regenerative ability than wild-types (70% successful regeneration in daf-2 versus 12.5% in wild-type) (Figure 6D and 6F). Similarly to axonal response to injury 11, we found that DAF-2 inhibits regeneration of aged dendrites through inhibition of DAF-16, as daf-16 mutants and daf-2;daf-16 double mutants showed regenerative decline during aging similar to wild-type (Figure 6F). In conclusion, our results reveal that dendrite regeneration following transection declines with aging, a phenotype that is dependent on DAF-2/IGF-1R and its target DAF-16/FOXO.

(A-D) PVD neurons immediately after cut and 24 h later in wild-type and daf-2 mutants at L4 and 5 days of adulthood (5d), as indicated. Schematic illustrations below negative images. Red lightings, injury sites. Blue arrows, fusion sites (successful regeneration). Scale bars, 20 μm.
(E) Response to injury of wild-types at L4-young adult (YA) and 2-3d within a short time (36 h) after injury. Number of animals: n=7 (L4-YA) and n=4 (2-3d).
(F) Percentage of successfully regenerating animals within 24-48 h post injury. p values from Fisher’s exact tests; * p<0.05, ** p<0.01, *** p<0.001. Number of animals: n ≥ 8. See also Figure 6-figure supplement 1 and Movies S2-S4.
AFF-1 mediates and restores dendrite regeneration in aged animals
A second fusogen in C. elegans is AFF-1 (Anchor cell Fusion Failure 1), a transmembrane protein related to EFF-1 that executes several fusion events during development 34 aff-1 mutants have no evident morphological phenotypes in the PVD neuron and expression has not been detected in the PVD (Figure 5-figure supplement 1); moreover, when AFF-1 is ectopically expressed in the PVD in an eff-1 mutant background it does not retract excess branching, demonstrating that it is unable to rescue the pruning defects of eff-1 (Figure 7-figure supplement 1). However, AFF-1 is required for dendrite fusion in response to injury in L4s and young adults 35. Adults carrying loss-of-function mutations in aff-1 have severe egg laying defects, shorter life spans and excretory system defects that prevented us from studying regeneration in aging aff-1 mutant animals.
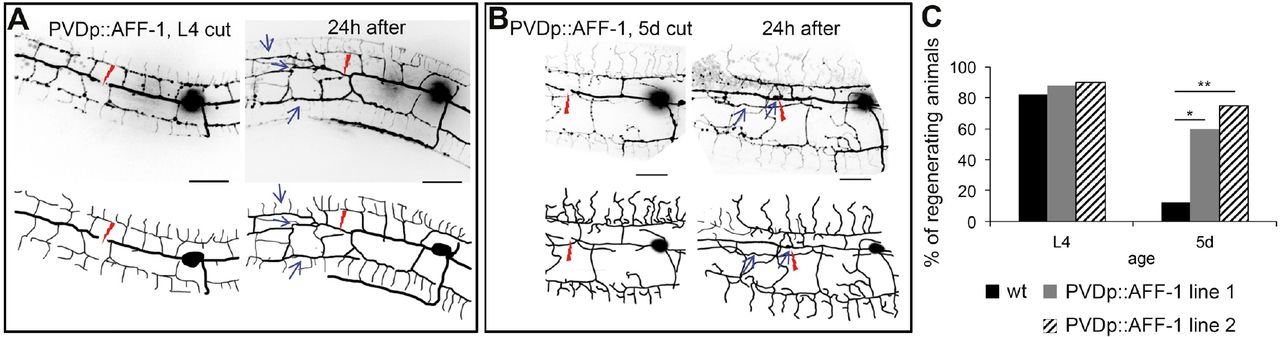
We next tested whether AFF-1 can restore the regenerative ability of dendrites in older animals by overexpressing AFF-1 specifically in the PVD (PVDp::AFF-1; Figure 7-figure supplement 2). We found that menorahs of young animals expressing PVDp::AFF-1 appeared morphologically wild-type and responded similarly to dendrotomy (Figure 7A, Movie S5). However, when 5-day old PVDp::AFF-1 animals were dendrotomized, the percentage of regenerating worms by dendrite fusion was significantly higher compared to wild-type animals (60-80% and 13%, respectively; Figure 7B and 7C, Movie S6).

(A-B)Inverted fluorescence images of PVD neurons immediately after cut and 24 hours (h) later in animals overexpressing AFF-1 in the PVD (PVDp::AFF-1), at L4 and 5 days of adulthood (5d), as indicated on each image (schematic drawings below each image). Red lightning marks the injury site and blue arrows point at fusion sites (menorah-menorah fusion). Scale bars, 20 μm.
(C)Percentage of successfully regenerating animals within 24-48h after injury. p value from Fisher’s exact test: * p<0.05; ** p<0.001. Number of animals: n ≥ 10. See also Figure 7-figure supplement 1 and Movies S5-S6. PVDp::AFF-1 line 1 and 2 correspond to worms carrying the extrachromosomal arrays hyEx390 and hyEx391, respectively.
Thus, AFF-1-specific overexpression in the PVD, as well as systemic daf-2 reduction of function, enables dendrite regeneration in older animals (Figures 6F and 7C).
Differential rejuvenation of fusion potential of transected dendrites by daf-2(−) and AFF-1(+)
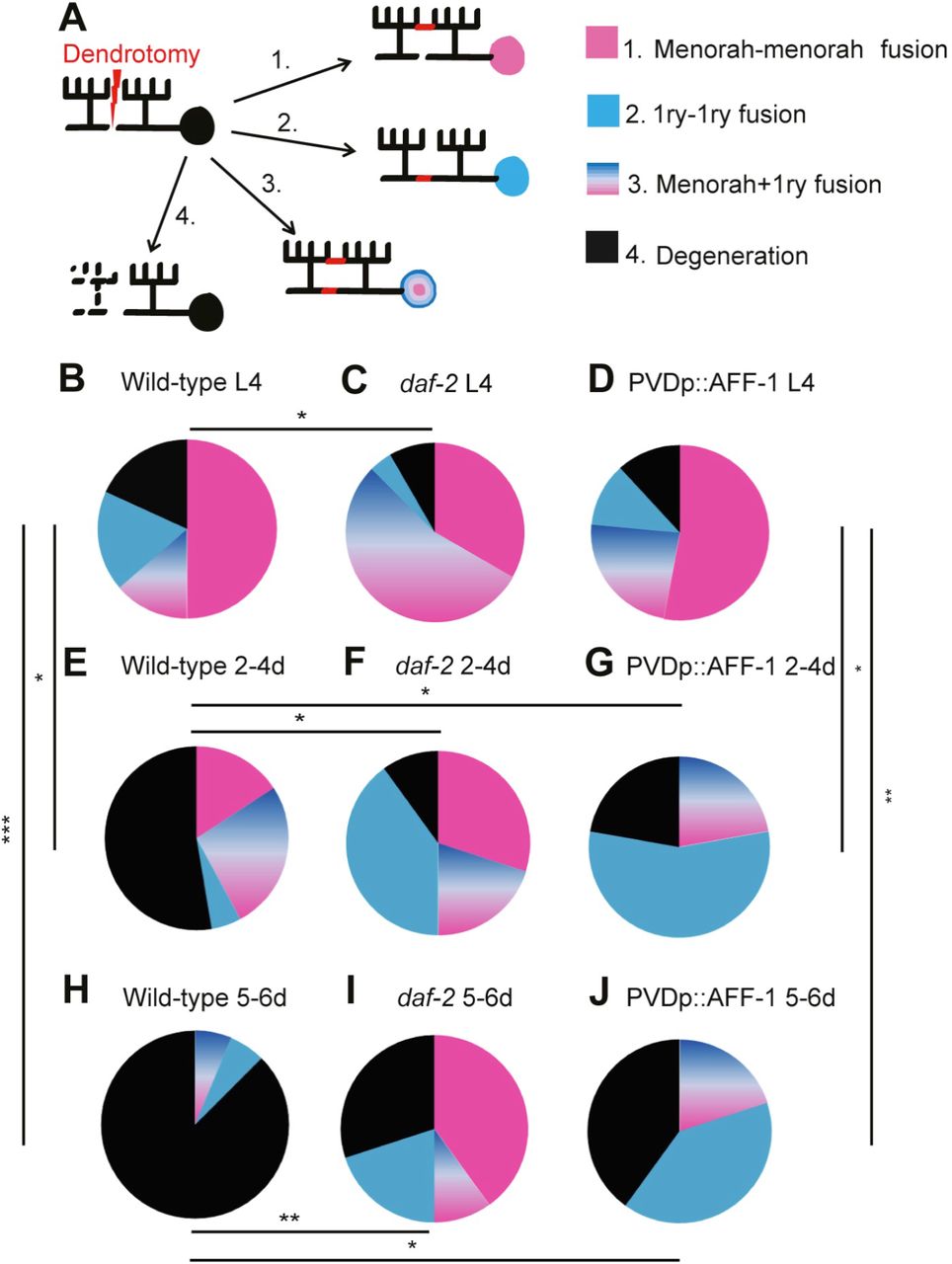
We wished to further compare the regenerative modalities of AFF-1 overexpression and daf-2 mutation following dendritic laser-induced severing. Dendritic fusion following injury can occur in three possible ways: 1) tertiary branch fusions that bypass the injury site ("menorah-menorah" fusion); 2) fusion of the proximal primary (1ry) branch to the detached distal 1ry; 3) both menorah-menorah fusion and 1ry-1ry fusion (Figure 8A). We found that in wild-type and PVDp::AFF-1 animals at the L4 stage, the most prevalent mechanism of repair is menorah-menorah fusion, whereas in daf-2 mutants menorah-menorah together with 1ry-1ry fusion (outcome 3) increased compared to wild-type (Figure 8B-8D). In wild-type, we found degeneration in 50% (2-4d) and 80% (5-6d) of the dendrotomized animals (Figure 8E and 8H). Adult daf-2 mutant animals (5-6d) presented a response to injury that resembled that observed in L4 wild-type animals, with 70% of the animals showing regeneration, mainly via menorah-menorah fusions (Figure 8F and 8I). In contrast, in PVDp::AFF-1 animals the response to injury followed a different mechanism, with more regeneration via enhanced 1ry-1ry fusion (Figure 8G and 8J). However, the AFF-1 effect was unrelated to longevity as PVDp::AFF-1 animals had normal lifespans (Figure 8-figure supplement 1). In summary, rejuvenation of the dendritic arbors uncovered here involves three distinct activities: i) EFF-1 pruning of old hyperbranched dendrites, ii) negative regulation of dendrite regeneration by DAF-2/ IGF-1R, and iii) AFF-1-induced auto-fusion of old transected primary dendrites (Figure 9).

(A)Cartoon representing four possible outcomes of dendrotomy. Dendrite auto-fusion indicates successful regeneration (outcomes 1-3), if fusion does not occur degeneration takes place (outcome 4).
(B-J)Percentages of wild-type worms (wt), daf-2 mutants and AFF-1 overexpressing animals (PVDp::AFF-1) at different ages, divided into the four different types of response to injury as described in (A). Genotype and age are listed above each plot. p values from Fisher’s exact tests: * p<0.05, ** pμ0.01, *** p<0.001. Additional significant differences were found between: (C and F), (C and I) and between (B and J) (pμ0.05). See also Figure 8-figure supplement 1.

Morphological agingof PVD menorahs is a progressive and dynamic process that results in loss of self-avoidance, disorganization and hyperbranching. When the fusogen EFF-1 is ectopically expressed in the PVD neuron it has an antiaging activity that involves pruning of hyperbranched dendritic trees.
Aged menorahs lose their regenerative potentialx because following laser-induced dendrotomy old neurons usually fail to auto-fuse broken dendrites and undergo degeneration. DAF-2/ IGF-1R negatively regulates the regeneration process. When the fusogen AFF-1 is ectopically expressed in the PVD neuron it has an antiaging activity that promotes autofusion of old transected primary dendrites.
Discussion
Age-related deterioration of dendritic trees is insulin/IGF1-independent
Precise dendritic arborization is critical for proper functioning of neuronal networks, while defective dendritic development and maintenance lead to neuropathologies 44,45. There is evidence from mammalian CNS showing increase in number and length of terminal dendritic arbors during aging, and reduced arborization in senile dementia 46. However, it remains unclear whether there is a direct link between altered dendritic structure and reduced function in mammals. Axons of aging neurons also show altered morphologies in diverse species. In C. elegans these axonal alterations are not affected by organismal longevity 11,42,43,47. Here we also found that dendritic alterations during aging are not affected by chronological age; however, unlike the PVD dendrites, in axons the daf-2 mutation was found to delay the morphological alterations of aged animals 8–10. Nevertheless, the daf-2 effect in age-related axonal branching was reported to be uncoupled of its role in extending the lifespan of the worms 8,9 It thus appears that DAF-2 is specifically involved in cell-autonomous pathways that maintain axonal, but not dendritic morphology. While progressive age-related hyperbranching of the PVD arbors was independent of insulin/IGF-1 pathway, overexpression of EFF-1 in the PVD was sufficient to partially rescue this phenotype in old animals (Figure 5). EFF-1 appears to act cell-autonomously to simplify the dendritic trees via its pruning activity mediated by branch retraction 23 (Figures 5 and 9).
AFF-1 restores regeneration of old broken dendrites via plasma membrane fusion
We further found that dendrite regeneration following PVD dendrotomy decreases with age. This decline was "rescued" in daf-2 mutants (Figure 6). Axonal regeneration is known to decline with age in different organisms 11,42,43,47–49. In GABA neurons of C. elegans the decline in axonal regeneration is also delayed in daf-2 mutants 11. However, these neurons do not engage in self-fusion during regeneration, whereas in PVD dendrites the main outcome of regeneration is auto-fusion. Recently, an important role of auto-fusion during axonal regeneration was demonstrated in the PLM sensory neurons of C. elegans, a process mediated by the fusogen EFF-1 12,15. In the PVD EFF-1 is not necessary for dendrite reconnection following injury 35. In the PVD dendrites, this role is fulfilled by the EFF-1 paralog AFF-1. Significantly, when AFF-1 is ectopically expressed, this fusogen plays a role in maintenance of regenerative potential during aging (Figures 7 and 9). These findings highlight fusion mechanisms as potential target for pharmacological intervention in neuropathologies that result from both injury and aging.
Author contributions
V.K., M.O.S. and B.P. designed the experiments. V.K. performed most experiments. M.O.S. developed the system to study neuronal degeneration and regeneration following dendrotomy using the PVD neurons. M.O.S. generated the PVDp::AFF-1 and PVDp::EFF-1 transgenics and M.O.S. and V.K. tested the effects of EFF-1 and AFF-1 over-expression on regenerating animals. B.P. supervised this work. V.K. and B.P. wrote the paper with input from M.O.S.
The authors declare no conflict of interest.
Acknowledgments
We thank D. Cassel, M. Hilliard, and A. Sapir for critically reading this manuscript. M. Heiman and C. Yip for providing CHB392, C. Smith and D. Miller for NC1841, and T. Gattegno for BP709. We also want to thank R. Kishony and all members of the Podbilewicz laboratory for discussion. This work was supported by Israel Science Foundation 443/12 and ERC ELEGANSFUSION 268843 grants to BP. Some strains were provided by the CGC, which is funded by NIH Office of Research Infrastructure Programs (P40 OD010440).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}