

ABSTRACT
Strategies aimed at reducing cerebral accumulation of the amyloid-β (Aβ) peptides have therapeutic potential in Alzheimer’s disease (AD). Aβ immunization has proven to be effective at promoting Aβ clearance in animal models but adverse effects have hampered its clinical evaluation. The first anti-Aβ immunization clinical trial, which assessed a full-length Aβ1-42 vaccine, increased the risk of encephalitis most likely because of autoimmune pro-inflammatory T helper 1 (Th1) response against all forms of Aβ. Immunization against less abundant but potentially more pathologically relevant Aβ products, such as N-terminally-truncated pyroglutamate-3 Aβ (AβpE3), could provide efficacy and improve tolerability in Aβ immunotherapy. Here, we describe a selective vaccine against AβpE3 using the diphtheria toxin mutant CRM197 as carrier protein for epitope presentation. CRM197 is currently used in licensed vaccines and has demonstrated excellent immunogenicity and safety in humans. In mice, our AβpE3:CRM197 vaccine triggered the production of specific anti-AβpE3 antibodies that did not cross-react with Aβ1-42, non-cyclized AβE3, or N-terminally-truncated pyroglutamate-11 Aβ (AβpE11). AβpE3:CRM197 antiserum strongly labeled AβpE3 in insoluble protein extracts and decorated cortical amyloid plaques in human AD brains. Anti-AβpE3 antibodies were almost exclusively of the IgG1 isotype, suggesting an anti-inflammatory Th2 response bias to the AβpE3:CRM197 vaccine. To the best of our knowledge, this study shows for the first time that CRM197 has potential as a safe and suitable vaccine carrier for active and selective immunization against specific protein sequence modifications or conformations, such as AβpE3.
INTRODUCTION
Anti-amyloid-β (Aβ) immunotherapy is under intense investigation in Alzheimer’s disease (AD) (Lemere 2013; Wisniewski & Goñi 2015). Aβ is the core component of the amyloid plaques, a hallmark of the AD brain, and mutations in its precursor APP or in presenilins—the catalytic components of the Aβ-producing enzyme γ-secretase—cause familial AD (Checler 1995; Marambaud & Robakis 2005; Selkoe 2001). Thus, strategies aimed at preventing or lowering Aβ cerebral accumulation might interfere with AD pathogenesis (Citron 2010). Aβ immunization has proven to be very effective at promoting Aβ clearance, at least in animal models. Preclinical and clinical studies, however, have been hampered by unforeseen side effects. The first clinical trial AN-1792, which evaluated an Aβ1-42/QS21 vaccine, was halted in phase II when about 6% of the patients developed meningoencephalitis (Holmes et al. 2008). Although the exact mechanism that led to acute brain inflammation in this clinical trial remains unclear, it is believed that encephalitis arose from an autoimmune reaction triggered by a vaccine directed against the abundant self-protein Aβ coupled to the strong adjuvant QS21, thus favoring pro-inflammatory T helper 1 (Th1) immune responses (Tabira 2010). In this context, second generation anti-Aβ vaccines were designed to prevent T-cell responses during anti-Aβ immunization. These vaccines tested, for instance, N-terminal epitopes within the Aβ sequence and adjuvants that minimize T-cell engagement and favor B-cell responses (Wisniewski & Goñi 2015). Another approach is passive immunization, which has the advantage to bypass T-cell engagement and allow a better control of monoclonal antibody (mAb) dosage and epitope targeting. However, recent phase III AD trials of two anti-Aβ mAb, solanezumab and bapineuzumab, failed to slow cognitive or functional decline in patients with mild-to-moderate AD (Hampel et al. 2015). The main argument put forward to explain the lack of efficacy of these passive immunization approaches was that treatment might have started too late to reverse or delay the disease process (Karran & Hardy 2014). It is also possible that passive immunization might not deliver enough mAb to promote plaque clearance. Passive immunization also raised concerned because of the practical and financial sustainability of injecting and monitoring mAb injections on a regular basis for several years (Golde 2014).
In the amyloidogenic pathway, APP is sequentially endoproteolyzed by the proteases β-secretase/BACE1 and the presenilin/γ-secretase complex to produce various Aβ peptides, including the most abundant isoforms Aβ1-40 and Aβ1-42 (De Strooper et al. 2010). In addition to these major Aβ isoforms, N-terminally truncated Aβ products have been identified in the AD brain, including peptides starting with pyroglutamate residues at the position 3 (AβpE3) and 11 (AβpE11) (Mori et al. 1992; Portelius et al. 2010; Saido et al. 1995). N-terminal truncation was proposed to be mediated, at least in part, by aminopeptidase A (Sevalle et al. 2009) and cyclization of N-terminally-exposed glutamates is catalyzed by the enzyme glutaminyl cyclase (Schilling et al. 2008). Pyroglutamate Aβ is a promising target because appears to play a key role in Aβ oligomerization, seeding, and stabilization (Dammers et al. 2015; He & Barrow 1999; Nussbaum et al. 2012; Piccini et al. 2005; Schilling et al. 2006; Schlenzig et al. 2009). Furthermore, pyroglutamate Aβ has specific neurotoxic properties in cell cultures and leads to cerebral neuronal loss and synaptic function impairments in mice (Alexandru et al. 2011; Gunn et al. 2016; Russo et al. 2002; Schlenzig et al. 2012).
In this context, recent studies have proposed that immunization against less abundant but potentially more amyloidogenic and more neurotoxic isoforms of Aβ, such as AβpE3, could improve tolerability and efficacy in Aβ immunotherapy (Bayer & Wirths 2014; Cynis et al. 2016; Frost et al. 2015). These studies, however, are so far limited to passive immunization using antibodies previously screened in vitro for their specificity to AβpE3 and non-cross-reactivity to full length Aβ. Here, we describe a novel AβpE3 vaccine using the non-toxic mutant of diphtheria toxin CRM197 (cross-reacting material 197) as carrier protein for epitope presentation. CRM197 has been extensively used in licensed vaccines directed against capsular polysaccharides of several bacterial pathogens and has demonstrated excellent efficacy and tolerability in humans (Bröker et al. 2011; Shinefield 2010). Because recent data have shown that CRM197 is also suitable for conjugation to, and presentation of, peptides (Caro-Aguilar et al. 2013), we speculated that CRM197 could be conjugated to minimal peptide epitopes to facilitate the generation of conformation/modification-specific antibodies directed against pyroglutamate Aβ. We show that our vaccines in mice—composed of AβpE3-8 or AβpE11-16 peptides covalently conjugated to CRM197—triggered the production of fully specific antibodies directed against AβpE3 and AβpE11, respectively. Anti-AβpE3 antibodies stained brains from AD patients by western blotting (WB) and immunohistochemistry (IHC), and were almost exclusively of the IgG1 isotype, indicating the engagement of a Th2 response.
MATERIALS AND METHODS
Preparation of the AβpE3-8:CRM197 and AβpE11-16:CRM197 conjugate vaccines
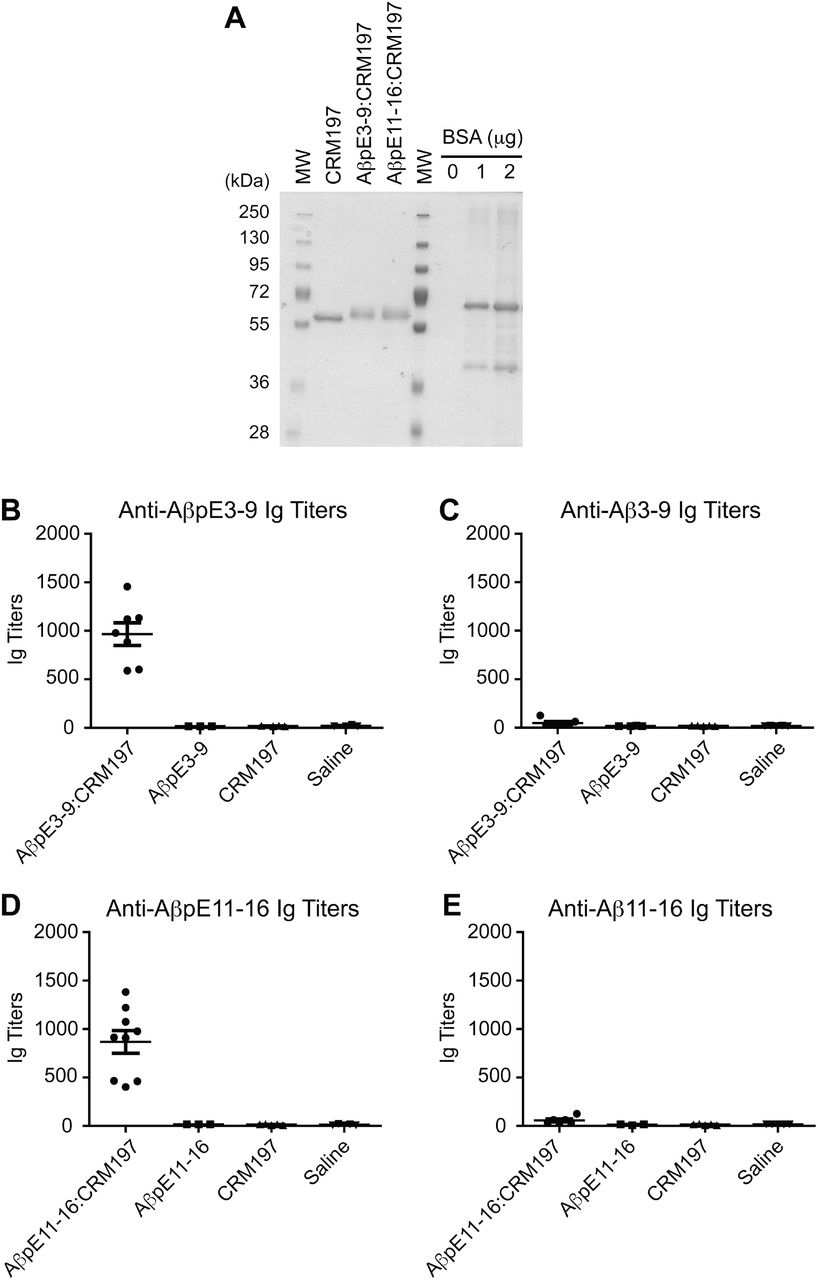
N-terminally acetylated and C-terminally amidated AβpE3-8 and AβpE11-16 peptides containing a C-terminal cysteine residue preceded by a two-glycine-bridge (pEFRHDSGGC and pEVHHQKGGC, respectively; GenScript) were solubilized in phosphate buffered saline (PBS) containing 2 mM EDTA (PBS/EDTA) to obtain 4 mg/mL solutions. CRM197 (List Biological Labs) was reconstituted with PBS/EDTA to obtain a 2 mg/mL solution. To irreversibly crosslink the peptides to the carrier protein, CRM197 was first activated with a 20-fold molar excess of succinimidyl-4-(N-maleimidomethyl)cyclohexane-1-carboxylate (SMCC) crosslinker (Pierce, at 1.5 mg/mL in DMSO). After a 30 min incubation at room temperature (RT), the SMCC/CRM197 mixture was desalted on a resin column (Zeba Desalt Spin Column, Pierce). Activated CRM197 was then combined with AβpE3-8 and AβpE11-16 peptide solutions and incubated for 30 min at RT. A ratio of 1:10 (CRM197 to peptides) was empirically chosen. Successful conjugation was confirmed by SDS-PAGE and coomassie staining (GelCode, Pierce, see Fig. 2A).
Immunization and sample collection
Animal experiments were performed according to procedures approved by the Feinstein Institute for Medical Research Institutional Animal Care and Use Committee. C57BL/6J male mice (~3-month-old) were injected subcutaneously with 10 μg of AβpE3-8:CRM197 and AβpE11-16:CRM197 conjugate vaccines or with 10 μg of free AβpE3-8 and AβpE11-16 peptides, used as negative controls. A prime injection (Day 0) was followed by a boost injection at Day 14 of the same amount of vaccines or free peptides. Blood samples were taken at Day 0 and Day 30.
ELISA for measurement of antibody responses
Anti-AβpE3-8, anti-AβpE11-16, and anti-CRM197 antibody levels in mouse serum were determined by enzyme-linked immunosorbent assay (ELISA). 96-well plates (Maxisorp, Nunc) were coated with 100 μL of 2 μg/mL AβpE3-8, AβpE11-16, or CRM197 in carbonate buffer (0.05 M, pH 9.6) and incubated overnight at 4oC. The following morning, plates were blocked for 1 h at RT with 5% skim milk in TBS containing 0.05% Tween 20 (TBST). After washing with TBST, serial dilutions of individual mouse serum samples (diluted in PBST containing 1% skim milk) were prepared and 100 μL/well of mouse serum was incubated for 2 h at RT. After 5 more washes, 100 μL/well horseradish peroxidase (HRP)-conjugated goat anti-mouse immunoglobins (Igs) secondary antibody (Southern Biotech, diluted 1:500 in PBST containing 1% skim milk) was incubated for 1 h at RT. TMB substrate was added after another wash and the reaction was allowed to develop for 30 min at RT. The optical density was measured at 405 nm using a TECAN GENios Pro plate reader. Antibody responses were expressed as titers. Antibody titers were determined via a linear fit for optical density values of 9 dilutions, and expressed in arbitrary units by calculating the reciprocal dilution that gave 50% of the maximum absorbance response. For measurement of Ig isotype specificity (IgG1, IgG2b, IgG2c, IgG3, IgA, and IgM), an ELISA protocol similar to that described above was followed. Briefly, ELISA plates were coated as described above. Reference mouse IgG1, IgG2b, IgG2c, IgG3, IgA and IgM antibodies were diluted in TBST containing 1% skim milk. To detect specific binding, HRP-conjugated anti-mouse IgG1, IgG2b, IgG2c, IgG3, IgA and IgM antibodies (Southern Biotech) were used at a dilution of 1:500, TMB was then added as described above. The presence of antigen-specific isotype-specific antibodies was detected and measured as described above. Antibody levels were expressed in micrograms per milliliter.
Statistical analysis
For comparison of results within experimental groups, a Student’s t test was performed. For all multigroup comparisons, one-way analysis of variance (ANOVA) and post-hoc Bonferoni test for multiple comparisons were used. Statistical significance was defined as p < 0.05.
Human cases and brain protein extraction
Cases were obtained from the Albert Einstein College of Medicine human brain bank, Bronx, NY (see Table). Brain samples (see Table) were processed as previously described (Vingtdeux et al. 2011). Briefly, human brains were sequentially extracted to obtain a soluble SDS fraction and an insoluble formic acid (FA) fraction. Samples were first homogenized and sonicated in Tris-buffered saline (TBS) containing 2% SDS and 1x Complete protease inhibitor mixture (Roche Applied Science) and centrifuged at 100,000 x g for 1 h at 4°C. The supernatant was removed and the resulting pellet was then extracted with 70% formic acid in water. FA extracts were dried under vacuum in a speed vacuum and thus dissolved in DMSO.
Western blot (WB)
Protein extracts from SDS and FA fractions obtained from human brain samples were analyzed by SDS-PAGE using the indicated antibodies. Samples were electrophoresed on 10% Tris-HCl gels (phospho-tau and actin analysis) or on 16.5% Tris-Tricine gels (Aβ isoforms, Biorad) and transferred onto nitrocellulose membranes. Aβ was detected as previously described (Vingtdeux et al. 2015). Briefly, membranes were microwaved for 5 min in PBS. Membranes were then blocked in 5% fat-free milk in TBS, and incubated with primary antibodies overnight at 4°C. A standard ECL detection procedure was then used.
Immunohistochemistry (IHC)
Five-μm-thick sections of formalin-fixed paraffin-embedded brain tissue were immunostained with 6E10 mAb (Covance, 1:1000 dilution) or AβpE3:CRM197 antiserum (1:2 dilution). Sections were deparaffinized by immersion in xylene and hydration through graded ethanol solutions. Denaturation for antigen recovery was performed by incubation of the slides in 70% formic acid for 30 min at RT. Denaturation was stopped by incubation in 100 mM Tris-HCl, pH 7.4 for 5 min. After washing once in TBS containing 0.05% Triton-X100 (TBSTx), endogenous peroxidase activity was inhibited by incubation in 5% hydrogen peroxide in TBSTx for 30 min at RT. After washing twice in TBSTx for 5 min and once in water, sections were blocked in 10% fat-free milk (6E10) or 5% normal goat serum (AβpE3:CRM197 antiserum) in TBSTx containing 1 mg/mL BSA and 1 mM NaF for 1 h at RT. Sections were then incubated in the presence of primary antibodies diluted in 10% fat-free milk (6E10) or 5% normal goat serum (AβpE3:CRM197 antiserum) in TBSTx containing 1 mg/ml BSA and 1 mM NaF overnight at 4oC in a humidified chamber. After washing, the sections were incubated with biotin-coupled anti-mouse IgG1 secondary antibodies (1:1,000 dilution in TBSTx with 20% Superblock, Pierce) before incubation with streptavidin-HRP (1:1,000 dilution in TBSTx with 20% Superblock, Southern Biotech) and visualization with diaminobenzidine tetrahydrochloride.
RESULTS
Immunogenicity of CRM197 in mice
We first verified that unconjugated CRM197 without adjuvant was immunogenic in C57BL/6J mice. Based on previously published dose-ranging immunogenicity analyses of different CRM197-based vaccines (Caro-Aguilar et al. 2013; Rondini et al. 2011), we choose to inject 10 μg of CRM197. A prime injection of unconjugated CRM197 followed by a boost injection at Day 15 of the same amount of carrier protein elicited robust immunogenicity against CRM197 at Day 30 (Fig. 1A). Saline control injections as expected did not elicit an anti-CRM197 antibody response (Fig. 1A). Ig isotype determination revealed that anti-CRM197 antibodies were exclusively of the IgG1 isotype and reached levels of ~35 μg/mL at Day 30 post-prime injection (Fig. 1B).

Specific anti-CRM197 Ig titers (A) and Ig isotype concentration (IgG1, IgG2a, IgG2b, IgG3, IgA, IgM) (B) quantified by ELISA in sera obtained from C57BL/6J mice immunized (CRM197) or not (Saline) with CRM197. Sera were collected 30 days after prime injection. Data are mean ± SEM from 6 mice per group, each dot represent individual mouse serum.
Immunogenicity of the AβpE3:CRM197 and AβpE11:CRM197 conjugate vaccines
A short peptide sequence of 6 residues starting at pE3 (pE3-8) or pE11 (pE11-16) linked in C-terminal to a 3-residue spacer (see Methods) was chosen to facilitate the production of conformation/modification-specific antibodies directed against Aβ. A 6-residue peptide epitope has also the advantage to be shorter than conventional T-cell epitopes and thus might prevent unwanted T-cell activation and detrimental pro-inflammatory responses (Rammensee 1995). Molecular weight analysis by protein staining after SDS-PAGE revealed that pE3-8 and pE11-16 peptides could be covalently conjugated to CRM197 (Fig. 2A). The generated AβpE3:CRM197 and AβpE11:CRM197 conjugates (10 μg) elicited robust antibody responses against AβpE3-8 and AβpE11-16 peptides, respectively, when assessed by ELISA. Total serum Ig titers were in average of 1,000 at Day 30 after one prime injection and one boost injection at Day 15 (Figs. 2B and 2D). Importantly, the vaccines did not generate antibodies directed against the corresponding non-cyclized Aβ3-8 and Aβ11-16 peptides (Figs. 2C and 2E), showing that immunogenicity was specific to the pE modification. No anti-AβpE3- or anti-AβpE11-specific antibodies were detected in control mice injected with unconjugated CRM197 or saline (Figs. 2B and 2D). Injection of free AβpE3-8 and AβpE11-16 peptides (unconjugated to CRM197) did not elicit immunogenic responses against these peptides (Figs. 2B and 2D).

Coomassie blue staining of CRM197 alone or conjugated to AβpE3-9 (AβpE3-9:CRM197) or AβpE11-16 (AβpE11-16:CRM197) (A). Anti-AβpE3-9 (B), anti-Aβ3-9 (C), anti-AβpE11-16 (D), and anti-Aβ11-16 (E) Ig titers in sera obtained from mice immunized with AβpE3-9 or AβpE11-16 conjugated (AβpE3-9:CRM197, AβpE11-16:CRM197) or not (AβpE3-9, AβpE11-16) with CRM197. Data represents mean ± SEM from 3-9 mice per group. MW, molecular weight makers.
As observed for the immunogenic responses to unconjugated CRM197 (Fig. 1), the anti-AβpE3 and anti-AβpE11 antibodies were exclusively of the IgG1 isotype (Figs. 3A and 3B). At Day 30 post-prime injection, AβpE3:CRM197 and AβpE11:CRM197 vaccines elicited the production of ~1.5 μg/mL of serum specific IgG1.

Anti-AβpE3-9 (A) or anti-AβpE11-16 Ig isotype (IgG1, IgG2a, IgG2b, IgG3, IgA, IgM) concentrations in sera obtained from mice immunized with AβpE3-9 or AβpE11-16 conjugated (AβpE3-9:CRM197, AβpE11-16:CRM197) or not (AbpE3-9, AbpE11-16) with CRM197 (B). Data represents mean ± SEM from 3-7 mice per group.
The AβpE3:CRM197 vaccine produces antibodies that react with amyloid deposits in AD brains
Antiserum produced after immunization with the AβpE3:CRM197 vaccine labeled synthetic AβpE3-42 peptide by WB (Fig. 4A). Importantly, AβpE3:CRM197 antiserum did not cross-react with synthetic AβpE11-42 or synthetic Aβ1-42 (Fig. 4A), showing that the antibodies produced did not recognize non-specifically the pE modification nor they interacted with an internal epitope containing residues 3-8 of Aβ1-42. AβpE3:CRM197 antiserum is thus fully specific to 241 AβpE3. Strikingly, AβpE3:CRM197 antiserum was also able to label AβpE3 in insoluble/aggregated brain protein preparations (FA fractions, see Methods) obtained from 6 independent well-characterized AD patients (see Table). Almost no immunoreactivity was observed in the corresponding soluble preparations (SDS fractions) (Fig. 4B), indicating that the anti-AβpE3 antibodies produced by the AβpE3:CRM197 vaccine recognize human amyloid 246 material in the AD brain, and that AβpE3 is mostly aggregated. Of note, AβpE3 immunoreactivity was also observed in normal control brains that contained large amounts of aggregated total Aβ (also detected with 6E10 mAb in the insoluble fractions) (Fig. 4B). In fact, a strong parallel was observed between the levels of aggregated total Aβ and aggregated AβpE3 in brains from both normal and AD individuals (Fig. 4B), suggesting that AβpE3 is a marker for amyloid deposition but is not specific to AD.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
WB analysis of recombinant Aβ1-42, AβpE3-42, and AβpE11-42 using 6E10, 4G8, or AβpE3-9:CRM197 antiserum showing the specificity of the AβpE3-9:CRM197 antiserum towards AβpE3-42 (A). WB analysis of human brain homogenates obtained from normal or AD cases (Braak stage V-VI, see Table) using PHF1, actin, and 6E10 antibodies, or AβpE3-9:CRM197 antiserum (B). IHC of serial brain sections from an AD case stained with 6E10 antibody or AβpE3-9:CRM197 antiserum (C).
In addition, AβpE3:CRM197 antiserum decorated amyloid plaques in human AD brain slices by IHC. Most of the anti-AβpE3 immunoreactivity was observed on plaques present in the cortex (entorhinal and parahippocampal cortices, Fig. 4C). Although amyloid plaque deposition was very pronounced in the AD case analyzed, only a few plaques were labeled with the AβpE3:CRM197 antiserum in the hippocampal formation (the dentate gyrus, CA1, and subiculum, Fig.4C). Interestingly, in adjacent sections of the parahippocampal cortex, identical plaques stained for both total Aβ (6E10 mAb) and AβpE3 (Fig. 4C, arrows). This co-staining revealed that anti-AβpE3 antiserum preferentially decorated highly dense plaques. Of note, anti-IgG1 secondary antibodies were used in combination with AβpE3:CRM197 antiserum for WB and IHC further showing that the immunoreactive anti-AβpE3 antibodies are of the IgG1 isotype.
DISCUSSION
In this study, we show that minimal epitopes can be designed to generate vaccines that are fully specific to the pyroglutamate modifications of Aβ. Indeed, antiserum obtained after immunization with the AβpE3:CRM197 conjugate vaccine, demonstrated excellent immunoreactivity against synthetic and AD brain-derived AβpE3, with no detectable cross-reactivity with Aβ1-42, non-cyclized AβE3, or AβpE11. Further analysis using AD brain samples revealed that AβpE3:CRM197 antiserum mainly decorated highly aggregated amyloid material, supporting the notion that AβpE3 is associated with the dense core of the senile plaques (Sullivan et al. 2011).
CRM197 is routinely used as conjugate vaccine in licensed vaccines directed against bacterial capsular polysaccharides (Bröker et al. 2011; Shinefield 2010). The use of CRM197 for peptide epitope presentation has not been approved yet for human use, but pre-clinical evidence has already suggested that it has therapeutic potential (Caro-Aguilar et al. 2013). In addition, an experimental vaccine against Aβ using CRM197 is currently being evaluated in a Phase II trial (ACC-001, Elan/J&J/Wyeth). In the ACC-001 vaccine, CRM197 is conjugated to the N-terminal residues 1-6 of Aβ and thus is aimed at targeting all Aβ isoforms containing this N-end of Aβ (Winblad et al. 2014). Our study strengthens the notion that CRM197 has strong potential for peptide presentation. This work further demonstrates that CRM197 might particularly be useful for the generation of conformation/modification-specific anti-peptide vaccines.
Both active and passive immunization directed against Aβ have their respective pros and cons (see Introduction) and there is no consensus yet as to whether one approach has a stronger therapeutic potential for AD. Both approaches—targeting different Aβ epitopes—are therefore actively investigated at the pre-clinical and clinical levels. Our approach was aimed at increasing the anti-Aβ immunotherapy toolkit by proposing an active immunization strategy specifically targeting N-terminally-truncated pyroglutamate Aβ. Further studies will be required to validate the utility of such a vaccine in AD mouse models of amyloid deposition. It should be noted, however, that several studies using passive immunization specifically targeting AβpE3 have already been conducted in different mouse amyloid models. These studies provided solid and concordant evidence that anti-AβpE3 antibodies have an overall plaque lowering effect and can improve the associated cognitive/behavioral deficits (Demattos et al. 2012; Frost et al. 2015; Wirths et al. 2010). Thus, it will be important to determine whether selective and active anti-AβpE3 immunization could also reduce the pathology in these models.
In this context, new immunization formulations for the AβpE3:CRM197 vaccine will have to be tested to boost antibody production. Indeed, although the AβpE3:CRM197 antiserum was able to react with both synthetic AβpE3 and AD-brain derived amyloid material, its antibody levels in the serum around 1.5 μg/mL are likely to be too low to allow clearance of AβpE3 and amyloid in mouse AD models. Thus, future experiments will also have to determine whether immunogenicity of the AβpE3:CRM197 vaccine can be improved by repeated injections and/or addition of adjuvants. Interestingly, recent data demonstrated that combined addition of the adjuvants aluminum hydroxide and CpG can increase more than 100-fold the antibody titer of a nicotine:CRM197 conjugate vaccine (McCluskie et al. 2015) and QS-21 elicited consistently higher anti-Aβ IgG titers in a phase IIa trial for the Aβ1-6:CRM197 ACC-001 vaccine (Pasquier et al. 2016).
In conclusion, we propose that conjugation of peptides AβpE3-8 and AβpE11-16 to CRM197 can generate fully specific vaccines directed against AβpE3 and AβpE11, respectively. The main strength of this immunization strategy is to combine the advantages of a vaccine with the high specificity of targeting particularly amyloidogenic and neurotoxic sub-species of Aβ, while sparing the more abundant and maybe more physiologically relevant full-length Aβ40 and Aβ42.
AD, Alzheimer’s disease; Mid-Temp Cortex, mid-temporal cortex; Hp, hippocampal region. Cases were obtained from the Albert Einstein College of Medicine human brain bank, Bronx, NY.
REFERENCES




