

Abstract
During colonization of the human nasopharynx, multiple strains of the Gram-positive pathogen Streptococcus pneumoniae coexist and compete with each other using secreted antimicrobial peptides called bacteriocins. The major class of pneumococcal bacteriocins is encoded by the blp operon, whose transcription is controlled by the secretion and detection of a polymorphic family of quorum sensing (QS) peptides. We examined interactions between the blp QS signal (BlpC) and receptor (BlpH) across 4,096 pneumococcal genomes. Imperfect genomic concordance between nine QS signal peptides and five phylogenetically-related QS receptor groups suggested extensive signal crosstalk (where cells produce signals that non-clonal cells detect) and eavesdropping (where cells respond to signals that they do not produce). To test this, we quantified the response of reporter strains containing one of six different blp QS receptors to cognate and non-cognate synthetic peptide signals. As predicted, we found evidence for eavesdropping in four of these receptors and for crosstalk in five of six tested signals. These in vitro results were confirmed during interactions between adjacent pneumococcal colonies, providing evidence that crosstalk and eavesdropping occur at endogenous, ecologically-relevant, levels of signal secretion. Using a spatially explicit stochastic model, we show that eavesdropping genotypes gain evolutionary advantages during inter-strain competition, even when their affinity to non-cognate signals is as low as 10% of the affinity to their cognate signal. Our results highlight the importance of social interactions in mediating intraspecific competition among bacteria and clarify that diverse competitive interactions can be mediated by polymorphism in QS systems.
Significance Statement Quorum sensing (QS), where bacteria secrete and respond to chemical signals to coordinate population-wide behaviors, has revealed that bacteria are highly social. Here, we use bioinformatics, experiments, and simulations to investigate how diversity in QS signals and receptors can modify social interactions controlled by the QS system regulating antimicrobial peptide secretion in Streptococcus pneumoniae. We experimentally confirmed that single receptors can detect multiple signals (eavesdropping) and single signals activate multiple receptors (cross-talk), while simulations revealed that eavesdropping can be evolutionarily beneficial even when the affinity for non-cognate signals is very weak. Our results highlight the importance of eavesdropping and crosstalk as drivers of the outcome of competitive interactions mediated by bacterial quorum sensing.
Introduction
Quorum sensing (QS) is a mechanism of intercellular communication that allows bacterial populations to coordinately regulate gene expression in response to changes in population density. QS is controlled by the secretion and detection of diffusible signaling molecules that, at threshold concentrations, lead to increased signal secretion as well as the induction of coupled downstream pathways (1, 2). By this process, QS ensures that pathways for metabolically costly products are only induced when bacterial populations would benefit from their production, i.e. when they are at high concentrations (2, 3). Signaling in QS systems is coordinated by the fact that cells are simultaneously capable of sending and detecting a specific signal (2⇓–4), a characteristic that serves to increase the likelihood that QS functions as a private message among clonemates, thereby coordinating gene regulation between cells sharing evolutionary interests (5, 6). However, outside the laboratory, bacteria often reside in multispecies communities where secreted QS signals can be detected by any cell, not just their clonemates (3). Thus, although QS works as an effective means of gene regulation in the laboratory in single strain cultures, QS in nature may be less reliable because it is susceptible to signal eavesdropping (i.e. where a promiscuous QS receptor can detect a QS signal not produced by that genotype) and signal crosstalk (i.e. where a non-specific QS signal can activate QS receptors in genotypes that produce other QS signals) (Fig. 1A; 3, 7). Signal-blind bacteria that produce, but are incapable of responding to, QS signals can engage in signal crosstalk in order to manipulate the behavior of other cells, e.g. by inducing them to produce expensive public goods (8). However, crosstalk and eavesdropping can occur even if all cells within a population are otherwise phenotypically wild-type if (i) QS signals and receptors are polymorphic and (ii) signals can bind and activate more than one receptor variant. At present, the incidence and consequences of these deviations from canonical QS systems remain poorly understood. Here we examine these issues using the polymorphic QS system regulating bacteriocin production in the Gram-positive opportunistic pathogen Streptococcus pneumoniae, where QS is integral for mediating intraspecific competition.

A) Eavesdropping occurs when a QS receptor of a cell is activated by a QS signal that the cell does not produce, such as activation of the blue QS receptor by both the cognate blue square signal and non-cognate green triangle signal. Crosstalk occurs when a QS signal activates more than one receptor, such as the green triangle signal activating both the cognate green QS receptor and the non-cognate blue QS receptor. B) blp QS regulation. External BlpC signal binds to histidine kinase receptor BlpH. This activates response regulator BlpR through phosphorylation, which increases transcription of blpABC, blpT, and the blp bacteriocins (including blpK) and immunity genes. Pre-BlpC is processed and transported out of the cell by ABC transporters ComAB and BlpAB.
To initiate infection, S. pneumoniae must first successfully colonize the nasopharynx and then persist during subsequent colonization attempts from other strains. Commensal carriage of S. pneumoniae is ubiquitous, affecting up to 88% of children worldwide (9, 10), while between 5-52% of individuals are co-colonized with multiple strains (10⇓⇓–13). The interactions between different strains during colonization are thus widespread and dynamic, leading to rates of clonal turnover — where one strain displaces another — that occur on a timescale of days to months (14, 15). Among the key factors thought to mediate intraspecific competition among pneumococcal strains are small anti-microbial peptides with narrow target ranges called bacteriocins, many of which are regulated by QS. While some studies in this species have examined the dynamics of bacteriocin-mediated competition in vitro (16, 17) and in vivo (16) between strains either lacking or carrying a single QS type, there is limited understanding of the evolution and ecology of polymorphic QS systems that regulate bacteriocin production and immunity between competing strains. S. pneumoniae presents an ideal opportunity to study the evolution of QS systems in an easily manipulated, highly relevant study system.
The genome of S. pneumoniae encodes several bacteriocin families, the most diverse of which is the bacteriocin suite encoded by the blp (bacteriocin-like peptides) operon (16, 17). Our recent work revealed that the number of distinct combinations of bacteriocins and immunity genes can extend into the trillions, although phylogenetic and functional constraints reduce this number to several hundred realized combinations (18). As with other Gram-positive peptide signals, the QS signal peptide (BlpC) regulating the blp operon is constitutively produced at low levels, but is auto-induced at high levels once a threshold concentration has been reached (17). Secreted BlpC binds to the extracellular domain of the membrane-bound histidine kinase BlpH, and upon binding the kinase phosphorylates the response regulator BlpR (Fig. 1B; 14, 15), which initiates production of the blp bacteriocin genes and increases production of the BlpC signal. Additionally, blpC expression is enhanced by the induction of competence for genetic transformation, which is regulated by the paralogous com QS signaling system (21). Both ABC transporters BlpAB (22) and ComAB (21, 23) cleave the N-terminal, double-glycine leader sequence of BlpC before export of the mature peptide signal by the same transporters (Fig. 1B). Using QS to regulate their secretion ensures that Blp peptides are only produced when there is a sufficiently high cell number to allow the bacteriocins to reach effective concentrations.
Importantly, both the BlpC signal and its dedicated receptor, BlpH, are highly polymorphic. Our survey of 4,096 genomes identified 29 amino acid variants of the BlpC gene and 156 BlpH amino acid variants (18). What are the effects of this variation, and how does this diversity influence the competitive interactions between strains that are affected by blp bacteriocins? One extreme option is that each BlpC signal corresponds to a non-overlapping set of BlpH receptors to which it specifically and exclusively binds. By this explanation, strains respond only to their own signal in determining the threshold at which they induce the blp operon. Such exclusivity is found in the competence signaling system where the two dominant peptide signals, CSP1 and CSP2, only induce cells expressing the cognate receptor (24). Similarly, there is near absolute concordance between the signal and receptors carried by any single genome, suggesting that tight coupling of these loci is crucial for the activation of competence (25). An alternative possibility is that BlpC peptides cross-react via crosstalk or eavesdropping with different BlpH receptors with varying affinity levels, thereby leading to a scenario whereby strains interact and possibly cause competing strains to induce the production of either immunity or bacteriocins at densities that would be insufficient for activation by auto-induction. Previous experimental research supported restricted crosstalk in the BlpH receptor, with a trade-off between signal sensitivity and signal specificity (26). By this explanation, strains may benefit if they are forewarned of the threats from others, allowing them to induce their own bacteriocins or immunity. Alternatively, eavesdropping may be costly if strains with promiscuous receptors are induced to secrete bacteriocins at densities that are too low to provide sufficient benefits to offset the costs of their production.
To understand the incidence and consequences of crosstalk and eavesdropping in a QS system that contains functional diversity, we investigated the fidelity between BlpC signals and BlpH histidine kinases across thousands of genomes using a bioinformatics approach. These results then informed experiments that quantified the response of bacterial strains to cognate and non-cognate peptide signals across the major signaling classes. Finally, these results were then examined in light of a stochastic model that investigated the consequences of QS eavesdropping for bacteriocin regulation. Our results reveal the importance of QS signaling polymorphism on blp operon regulation and clarify its ecological effects on S. pneumoniae intraspecific interactions. More broadly, we explore crosstalk and eavesdropping in a QS system that contains functional diversity.
Results
Molecular diversity of blpH and blpC
We examined 4,096 S. pneumoniae genomes across six unbiased data sets (Maela, Massachusetts Asymptomatic, GenBank, Hermans, Georgia GenBank, and PMEN) alongside two additional data sets that represent intentionally biased samples of clonal sub-groups (Complex 3 and PMEN-1, 322 genomes combined). We identified blpC in 99.0%, blpH in 99.0%, and both blpC and blpH in 98.2% of the 4,418 genomes using a DNA reciprocal BLAST algorithm (18). We note that the few genomes apparently lacking a blp operon gene may still contain these genes, as the data sets contain incomplete draft genomes.
We observed extensive allelic variation within blpC, which contains 37 alleles at the nucleotide level, 29 protein variants, and 20 different mature BlpC signal peptides, including signal peptides lacking a double-glycine cleavage site. Nine of these peptide signal sequences were found in more than 0.5% of genomes (i.e., over 20 genomes; Table 1), and together these nine comprise ~98% of all signal variants. All signals under this 0.5% threshold were each confined to a single clade in the whole-genome phylogeny (Fig. S1A). Each unique BlpC signal was designated with a letter from the NATO phonetic alphabet. As expected for the genomes from intentionally biased samples, the PMEN-1 data set almost exclusively carried the Golf signal (93.8%; Table 1), while the Clonal Complex 3 data set almost exclusively carried the Kilo signal (97.6%; Table 1). The Bravo and Hotel signal peptides were exclusively found in strains collected as part of the Maela data set. Even though the Maela genomes composed the majority of genomes in our data, the relatively high proportion of Maela genomes with the Bravo and Hotel signals (3.8% and 8.2%, respectively) suggests that either natural selection or limited admixture prevented these signals from appearing in the other data sets. There was more variation in blpH (194 alleles at the nucleotide level across 156 amino acid variants) than in blpC, although this is a likely consequence of the fact that blpH is longer than blpC. Rarefaction curves of non-singleton protein variants of BlpC and BlpH show that the diversity of protein variants reached saturation after ~2,000 and ~3,000 sampled genomes, respectively (Fig. S1B).
Predicted blpC mature peptide signals and their frequencies in each data set
Indicators of molecular evolution showed that blpH and blpC are evolving rapidly (Table 2). blpH had the highest peaks of nucleotide diversity across the blp regulatory genes (Fig. S2A). Additionally, higher dN / dS ratios (i.e. a higher non-synonymous to synonymous mutation ratio) in the receptor versus the kinase region of this protein (Table 2) suggests stronger diversifying selection acting on peptide:receptor binding than on the intracellular kinase domain responsible for downstream signal transfer by phosphorylation. Inferred rates of recombination peaked within the second transmembrane domain of BlpH; however, specific peaks of either nucleotide diversity or recombination were not distinguished by predicted transmembrane regions (Fig. S2B). Recombination also locally peaked within blpC (Fig. S2B), which also had an increased dN / dS ratio when compared to estimates for housekeeping genes (Table 2).
Nucleotide diversity and dN/dS for selected genes
blpC/blpH intragenomic pairing is highly biased but not exclusive
Phylogenetic analysis of blpC revealed four well-supported clades (Fig. 2) containing the following signals: 1) Alpha, Bravo, and Kilo; 2) Golf and Hotel; 3) Charlie; and 4) Delta, Echo, and Foxtrot. The relationships between signaling groups within these major clades are uncertain, although there is evidence (0.75 < posterior probability < 0.95) that the Hotel, Bravo, and Delta signals are each monophyletic within their respective larger clades.

Taxa are colored by mature BlpC signal with the signal designation followed by the number of genomes containing the allele. Internal nodes show the posterior probabilities of clades; we collapsed clades with less than 0.75 posterior probability.
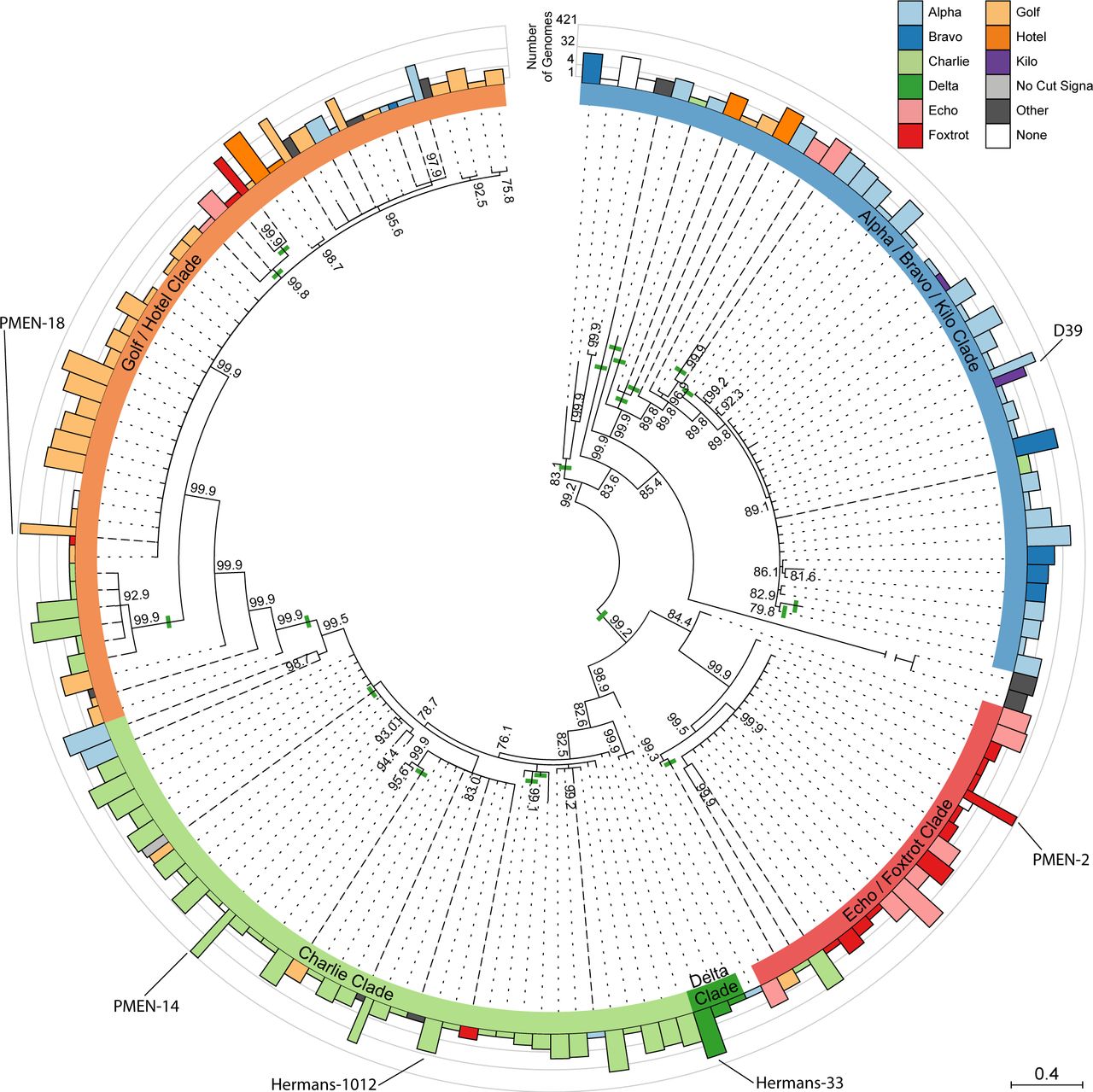
After accounting for recombination, phylogenetic analysis of blpH identified five paraphyletic clades that are broadly concordant with the divisions observed for BlpC signals (Fig. 3), although there are clear exceptions to this correspondence. Across the five clades, the classification of blpH alleles correlated with the mature BlpC signal in at least 75% of cases: (Alpha / Bravo / Kilo Clade: 88.5%; Echo / Foxtrot Clade: 94.6%; Delta Clade: 100%: Charlie Clade: 86.3%; Golf / Hotel Clade: 75.5%). Examples of a mismatched BlpC signal and blpH receptor occur at 27 distinct clades on the blpH tree. Evidence of extensive recombination affecting the blpH kinase, intergenic region, and blpC signal (Fig. S2) suggests that recombination has caused some of these mismatches; however, multiple recombination events in the same region obscure reconstructing these evolutionary events with confidence. Overall, from the 4,096 genomes in this analysis, 596 genomes (14.6%) show a lack of correspondence between signal and peptide, suggesting either that these strains are deficient in blp signaling or that these BlpH histidine kinase receptors can be cross-induced by non-cognate BlpC signals. Overall frequencies by signal and receptor class are summarized in Fig. 4A.

The outer ring shows the number of 4,096 genomes with each blpH allele, color-coded by their co-occurring BlpC signal and on a log scale. The inner ring denotes the blpH clade type, and recombination events within blpH are shown as solid green lines. Mismatches between blpH clade and BlpC signal are indicated by dashed lines. Internal nodes show the posterior probabilities of clades; we collapsed clades with less than 0.75 posterior probability.

A) Proportion of each BlpC signal within genomes containing each blpH clade. The phylograms are simplified versions of Fig. 1 and Fig. 2. B) The relative maximal expression levels of luc following addition of 1 ?g/ml of synthesized BlpC signal peptide. The maximum expression level for each reporter strain was set to 1. Raw data is found in Fig. S6. C) The minimum concentration of synthesized BlpC signal peptide required for luc induction in reporter strains with different BlpH. Example of raw data is provided in Fig. S6. The Bravo, Kilo, and Hotel signal peptides were not synthesized and are denoted with slashes.
Crosstalk and eavesdropping between BlpC signals and BlpH receptors
To examine the incidence of crosstalk and eavesdropping between signals and receptors experimentally, we measured the responsiveness of each of the major BlpH clades to synthetic peptides from each BlpC class. We transformed a S. pneumoniae D39 strain lacking the native blp regulatory genes (blpSRHC) with constructs expressing one of six different BlpH histidine kinases alleles: blpSRHD39 from the Alpha/Bravo/Kilo clade, blpSRHPMEN-2 from the Echo/Foxtrot clade, blpSRHHermans-33 from the Delta clade, blpSRHHermans-1012 and blpSRHPMEN-14 from the Charlie clade, and blpSRHPMEN-18 from the Golf/Hotel clade. The respective strains also contained a reporter cassette, in which a typical blp-promoter (PblpK or PblpT) was controlling expression of firefly luciferase (luc), GFP ((sf)gfp), and β-galactosidase (lacZ) (21). Deletion of the blpC signal gene and the native blpSRH genes from the D39 ancestor ensured that the reporter strains would only be induced in response to exogenously added signal via the introduced blpSRH systems. By exposing cells to a concentration gradient of exogenous peptide, we could estimate the peptide concentration that induced the maximum response as well the minimum concentration required to elicit a response. While the maximum response indicates the overall influence of a given peptide on each receptor, the minimal concentration required to induce a response provides an indication of the sensitivity of each receptor to every potential peptide partner. Figures 4A-B shows that five of six PblpK reporter strains were maximally induced by the BlpC signal carried by a significant majority of their wild type counterparts. However, we also see extensive evidence for crosstalk and eavesdropping between mismatched peptide:receptor pairs, demonstrating that some BlpH receptors are highly promiscuous while equally, several BlpC peptides can induce the blp operon in strains carrying non-complementary BlpH receptors. For example, blpSRHPMEN-2 (Echo / Foxtrot BlpH clade) could be induced by 4 out of 6 synthetic peptides, and the strain with blpSRHHermans-1012 (Charlie BlpH clade) was strongly induced by the Echo and Foxtrot signals at 65% and 71% expression of its cognate signal. While there is clear evidence for cross-induction, these responses tended to be less sensitive to non-cognate peptides, with a minimum concentration required for induction of between 2-500-fold greater than with the cognate signal (Fig. 4C). By contrast, the strain with blpSRHHermans-1012 (Charlie BlpH clade) was more sensitive to the non-cognate Echo and Foxtrot signals (1 ng/ml and 3.9 ng/ml) than to its complementary Charlie signal (7.8 ng/ml; Fig. 4C). The reporter strain carrying blpSRHHermans-33 did not respond to any of the BlpC peptides, not even its cognate Delta BlpC (Fig. 4B-C). Interestingly, blpSRHHermans-33, as well as all other strains with blpH alleles in the Delta clade, contains a frameshift in the blpR gene, encoding the response regulator, thus preventing expression of the full-length blpR. This probably renders the QS systems non-functional and therefore not responsive to added peptide. All results were mirrored with a different set of reporter strains that used the blpT promoter for the reporter cassette (Fig. S3A). We conclude from these results that crosstalk among quorum-dependent peptide BlpC signals is widespread and concentration dependent, with strains able to eavesdrop onto multiple signals using cross-responsive receptors. Furthermore, these results are highly concordant with the patterns of co-association observed in our bioinformatics survey of pneumococcal strains.
Cross-induction between colonies
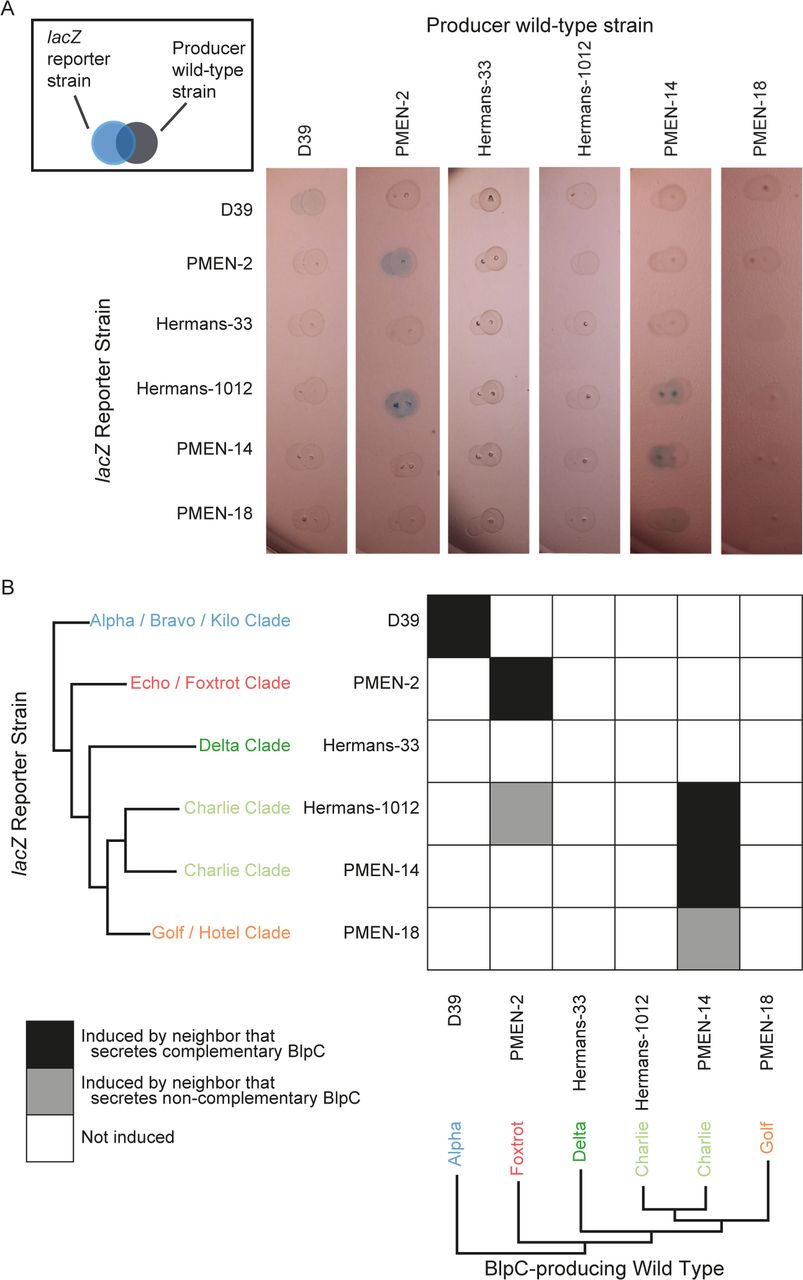
Pneumococci in the nasopharynx live in spatially structured colonies or biofilms. In order to determine if cross-induction could occur under these conditions, we examined interactions between neighboring colonies endogenously secreting either cognate or non-cognate signals (Fig. 5). In control assays, we first demonstrated that colonies were induced by exogenous addition of peptide to the plate surface; these results were concordant with those in Figure 4B in 14 of 15 combinations (Fig. S3B). Next, we measured expression of reporter strains when grown adjacent to wild-type colonies that secreted BlpC peptides at endogenous levels (Fig. 5A). We observed a response in the reporter strains as estimated by increased LacZ activity in 3 out of 6 strains, with 2 examples of induction by non-cognate BlpC signals. Interestingly, when the reporter strain expressing the BlpH from Hermans-1012 was grown adjacent to its wild type counterpart, there was no induction; instead this strain was induced by PMEN-14, which also produced the Charlie signal. The same strain was also induced by PMEN-2, which produced the Foxtrot signal (which induces Hermans-1012 at a lower concentration than with its cognate signal; Fig. 4C), and strain PMEN-18 (Golf/Hotel BlpH clade) was induced by PMEN-14, which produced the Charlie signal (Fig. 5). This may suggest that in addition to differences in the binding affinities of BlpC and BlpH, strains may also vary in the concentration of diffusible signal that they secrete, at least under these experimental conditions. Consistent with our in vitro assays with synthesized peptides, these results show that blp operon expression can be activated by crosstalk between neighboring competing colonies secreting peptides at wild-type concentrations.

A) The wild-type strains were spotted next to the reporter strains (see box), and induction of blp expression by the wild-type produced BlpC is shown as faint blue colonies. The experiment was repeated three times with the same result, and a representative photo of the plates is shown. B) Summary of the results from B. Squares in white indicate no induction of the reporter strain for colony pairs, while black and blue indicate induction by complementary and on-complementary BlpCs, respectively.
Evolutionary consequences of eavesdropping genotypes
Because the blp operon is auto-induced via a quorum dependent process, cross-induction can potentially influence other strains by lowering the population density required for auto-induction. To examine this issue, we developed a spatially explicit stochastic model to investigate conditions where genotypes with eavesdropping receptors may be favored over strains only able to respond to a single peptide signal. We further varied the signal affinity to eavesdropping receptors to determine how this altered the selective benefits of cross-induction. Simulations are initiated with cells of four genotypes randomly spaced upon a plane. The four genotypes each release their own QS signal at equal concentrations (Table S1). Cells bind these secreted signals in a concentration dependent manner, at which point they are induced to produce bacteriocins that kill susceptible neighbor cells at the cost of reduced growth for the producer (27). While two faithful-signaling genotypes are only able to respond to their own signals, the two other eavesdropping genotypes can respond to multiple signals. Our results shown in Fig. 6 lead to two important conclusions. First, we observe strong benefits to eavesdropping cells that depends on the degree of cross-sensitivity, or affinity, to non-cognate signals. Specifically, we found that higher affinity to non-cognate signal provides stronger ecological benefits. This results from earlier potential activation (Fig. S4) and secretion of bacteriocins in these cells, an effect that increases with greater affinity to non-cognate signals. Second, we find that the benefits to eavesdropping are strongly negative frequency-dependent, i.e. eavesdropping cells only gain benefits (in the form of earlier bacteriocin induction) when surrounded by faithful-signaling cells. When eavesdropping cells are rare, they benefit through maximum exposure to the alternative peptide, while after they increase they must rely solely on auto-induction. Because the benefits of eavesdropping are frequency dependent, these simple simulations thus suggest that promiscuous receptor mutants with increased affinity to non-cognate signals will be able to rapidly invade populations of cells that can only respond to a single signal. Interestingly, the simulations also clarify that the affinity to non-cognate signals can be extremely low — even at 10% of the affinity to cognate signals — to provide benefits (Fig. 6).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Average fitness of eavesdropping genotypes that produce bacteriocins in response to multiple signals in a spatially explicit, stochastic model. Simulations were started with five proportions of eavesdropping genotypes mixed with signal-faithful genotypes, as indicated on the x-axis. Absolute fitness values on the y-axis above 1.0 indicate that the genotype can increase in frequency in the population. Affinity to other genotypes’ signals are a percentage of affinity to a genotype’s own signal for eavesdropping genotypes. Error bars link the 25% and 75% quantiles for the final eavesdropping genotypes’ fitness across 100 simulations.
Discussion
Pneumococcal bacteriocins are believed to play a key role in mediating intraspecific competitive interactions. Here, we show that the QS system regulating blp bacteriocins is highly polymorphic, that there is widespread evidence that QS signals are cross-reactive (crosstalk), and that promiscuous receptors can detect and respond to non-cognate signals (eavesdropping). Assays between adjacent colonies revealed that both behaviors are manifest at endogenous concentrations of secreted peptides, while simulations revealed significant ecological benefits to strains that express promiscuous receptors. Together, these results suggest that social interactions influenced by QS signaling may have a strong influence on pneumococcal competition.
Previous surveys (19, 28) of BlpC and BlpH identified four BlpC signals: the Alpha, Charlie, Foxtrot, and Golf signals in our nomenclature, which together represent ~75% of the strains in our sample (Table 1). By expanding our survey to 4,096 strains, we identified several additional signal peptide families (Fig. 2): the Echo, Hotel, Delta, Bravo, and Kilo signals. Two of these (Bravo and Hotel) were found only in the Maela dataset, consistent with the idea of strong geographic structuring in this species; furthermore, because these signal variants appear in significant frequencies (3.8% and 8.1%, respectively, within the Maela data set), we infer that they are not actively selected against. The additional signals reported here suggest flexibility (and perhaps diversifying selection) in the mature signal peptide sequence, such as in the first two residues (which differ between the Alpha, Bravo, and Kilo signals, which share a co-occurring BlpH clade) and in signal residue 22, which differs between the Echo and Foxtrot signals despite the signals activating BlpH variants near identically in our experiments (Fig. 4B, Fig. 4C). Previous work suggested that differences in the electric charge of signal residue 14 is crucial for specificity (26); this residue is undoubtedly important because it differentiates the Alpha/Bravo/Kilo, Echo/Foxtrot, and Golf/Hotel signal groups. However, other signal residues are also likely to be important for BlpH binding, as signals that are identical at this site 14 (e.g. Golf/Hotel, and Alpha/Bravo/Kilo/Charlie) differentially bind/activate BlpH (Fig. 4). Although rarefaction analysis indicates that we have essentially saturated the diversity of signal and receptor types (Fig. S1B), more extensive sampling in other geographic locations, such as with the Global Pneumococcal Sequencing project (http://www.pneumogen.net/gps), is likely to uncover further rare variants.
The concordance between the phylogenies of blpC and blpH and the extensive co-occurrence in individual genomes suggest that these genes are co-evolving (Fig. 2, Fig. 3). At the same time, both blpC and blpH are changing rapidly, as indicated by their relatively high levels of nucleotide diversity (Fig. S2) and their high non-synonymous/synonymous substitution rates (Table 2), which is consistent with the idea that the genes are evolving non-neutrally. The co-variation between BlpC and BlpH also allows inferences on the key residues within each gene that mediate their binding. We used mutual information based upon this co-variation to identify residues in BlpH that are crucial for binding the QS signal, and these results can serve as a guide for experimental approaches to unravel the specificity of the BlpC/BlpH interaction (Fig. S5). Notably, our analysis of the BlpH receptor residues support previous findings that residues 17 and 119-124 are important for activation by BlpC signals (26), although additional residues co-vary with specific BlpC signals (Fig. S5).
While the correlation between blpH clade and co-occurring BlpC signal is high, in some clades the correlation drops to 75.5%, and BlpH / BlpC mismatches (Fig. 3) are widespread across the pneumococcal phylogeny; this can be compared to the exceptionally tight, > 99% correlation between the ComD QS receptor and CSP signal also in S. pneumoniae (25). There are at least two explanations for this difference. First, we do not have know if different BlpH variants are functionally distinct; all blpH alleles could, in principle, be most responsive to their co-occurring BlpC. This seems unlikely, given the high frequency (up to 36 signal:receptor pairs) of blpH clade / BlpC mismatches (Fig. 3). Second, weaker selection for a highly auto-inducing blp QS could explain the difference between the blp and com QS systems. After a recombination event that results in a sub-optimal BlpH/ BlpC pair for auto-induction, the BlpC signal may still be able to active the co-occurring BlpH variant through crosstalk, albeit at a higher concentration of BlpC (Fig. 4C). While auto-induction may be decreased, such a genotype would gain an eavesdropping receptor that can potentially detect signals of surrounding genotypes. For comparison, there is no eavesdropping between CSP pherotypes in the com QS system, and very rare signal/receptor mismatches (24, 25).
These signal/receptor mismatches can result in two outcomes for cell-to-cell communication. First, this can result in cells unable to detect the signal that they produce, rendering them unable to auto-induce. The lack of QS activation in strains producing the Delta signal (Hermans-33; Fig. 4) seemingly fits into this description; however, interestingly, this is not caused by signal / receptor mismatch because there is perfect concordance between the Delta signal and the Delta blpH clade, and no tested signal activated strains with Delta blpH. Instead, all 143 strains carrying the Delta signal have a frameshift in blpR, which suggests functional deterioration of the QS system in these strains, which has not yet led to deterioration of blpH and blpC. These Delta BlpC strains are not simply ‘cheater’ cells, as they continue to pay the cost of synthesizing BlpC. This suggests there may be weakened selection for functional blp QS.
The second outcome of signal/receptor mismatches for cell-to-cell communication is crosstalk and eavesdropping. We have ample evidence for crosstalk in the blp QS system, as all signal peptides except for the Alpha signal activated QS receptors in genotypes that produce other QS signals (Fig. 4B, Fig. 4C). Similarly, BlpH receptors (aside from the Alpha clade) were eavesdropping QS receptors able to detect more than one QS peptide signal (Fig. 4B, Fig. 4C). Each of the receptors we tested (except for the signal-blind BlpH Delta clade) was maximally induced with a single set of related signals and decreased to 0-71% with signals that the receptors were eavesdropping upon (Fig 4B). This suggests that there are no ‘generalist’ receptors that are able to listen to multiple signals with equal affinity. Previous research indicated that blpH alleles with more crosstalk were less sensitive to BlpC (26); however, reporter strain PMEN-2, Hermans-1012, and PMEN-14 were all sensitive to their complementary signal (≤1.0 ng/ml) but show extensive crosstalk (Fig. 4C). Crosstalk was seen in previous research, specifically induction from both the Charlie (signal 6A in (26)) and Foxtrot signal (signal 184 in (26)) in strains with a BlpH Charlie clade allele (6A.3 in (26)), and in induction from the Charlie and Golf (signal T4 in (26)) signals with the BlpH Golf clade (T4 in (26)). These two cases do require 2-7.8 times more of their own signal for minimum activation, compared to our BlpH Alpha clade and BlpH Foxtrot clade strains; this finding does supports a trade-off between signal sensitivity and specificity (26) but not to the exclusion of crosstalk. Overall, our experimental evidence supports extensive crosstalk and eavesdropping in the blp QS system.
What are the potential consequences of crosstalk and eavesdropping? The result of crosstalk could be to manipulate other, non-clonal, strains into inducing their QS system at lower densities, thereby causing competing strains to secrete bacteriocins and induced immunity proteins earlier. At present, it is unclear how such crosstalk would be beneficial to cells producing cross-reactive signals, unless premature production of bacteriocins or immunity introduces energetic or other costs to cells responding at sub-quorum densities. Similar benefits are thought to exist for other bacterial public goods (8, 29). By contrast, it is easier to envision the potential benefits of eavesdropping, which can both lead to earlier activation of bacteriocins (although this may have attendant costs) and earlier induction of cross-reactive immunity. Importantly, our simulations suggest that this could be beneficial even if the affinity of promiscuous receptors is only 10% of the affinity for their cognate signal (Fig. 6). This value falls within the range of responses we measured experimentally (Fig. 4C). This level of responsiveness is also sufficient to induce the blp operon among adjacent colonies secreting peptides at endogenous levels (Fig. 5).
Our results make clear that signaling interactions in vitro can lead to complex ecological outcomes that may strongly influence competitive interactions between strains. As yet, however, it is unclear how these interactions will play out in the complex within-host environment of the human nasopharynx. In addition, it remains unclear how these interactions directly influence bacteriocin-mediated killing and immunity. Clearly, the heterogeneous conditions in vivo differ markedly between liquid cultures or even agar surfaces. Diffusion is more limited, while population densities may be strongly constrained overall and spatially. These factors, among others, may alter the level and dispersion of signal peptides as well as the sensitivity of individual strains to these signals. It will remain an important aim for future work to elucidate the influence of these real-world factors. More generally, our results reinforce the importance of social interactions among bacteria for mediating competitive dynamics. Many ecologically relevant bacterial traits are regulated by QS, and many of these systems, especially in Gram-positive peptide signaling systems, are polymorphic. While some of these systems (e.g. pneumococcal competence regulated by the com QS system) have only few signal types and show no cross-reactivity, many others (e.g. Agr in S. aureus (30) and ComX in B. subtilis (31)) do or have yet to be measured. It remains to be investigated which, if any, of these polymorphic QS signals have ecological effects and which factors (such as co-colonization or extensive intraspecific competition) result in the evolution of crosstalk and eavesdropping.
Materials and Methods
Genomic data
We used S. pneumoniae genomes from eight publicly available sets, six of which contain strains that were randomly sampled from either cases of disease or asymptomatic carriage: 3,017 genomes from refugees in Maela, Thailand (32); 616 genomes from Massachusetts carriage strains (33); 295 genomes from GenBank, which include 121 genomes from Atlanta, Georgia, The United States (34); 142 genomes from Rotterdam, the Netherlands (Hermans data set) (18, 35); and 26 PMEN (Pneumococcal Molecular Epidemiology Network) genomes (18, 36). The PMEN-1 (37) and Clonal Complex 3 (38) data sets, containing 240 and 82 genomes, respectively, were a result of targeted sampling for specific clonal complexes of S. pneumoniae; as such, these strains were excluded from analyses that assumed random sampling. In each of these genomes, we located the blpC and blpH alleles using a DNA reciprocal BLAST search as previous described (18).
Phylogenetic Analysis
We examined if the rates of transition in changing BlpC signals were independent from the rates of transition between bacteriocin groups along the whole-genome phylogeny from (18). We did so by measuring the likelihood while estimating both independent and dependent transitions using BayesTraits (39) for each signal and bacteriocin group separately. We used a log ratio test to measure P-values from the likelihoods.
To infer the evolutionary history of blpC, we aligned the 37 alleles of blpC from all genomes, removed nucleotide sites caused by insertion mutations in single alleles, and reconstructed the phylogeny using Geneious 7.1.9 (40) and MrBayes 3.2.2 (40⇓–42) with an HKY+γ nucleotide substitution model as determined by jModelTest 2.1.7 (43).
Because blpH contained evidence of recombination, we modified our approach to infer its evolutionary history. We aligned the unique 161 full-length (≥ 1341 bp) blpH alleles using Geneious 7.1.9 (40). We then used Gubbins 1.4.2 (44) to detect and place recombination events onto a phylogenetic tree. To reconstruct the phylogeny while measuring the confidence for each clade, we used the filtered polymorphic sites from Gubbins, which replaced recombination fragments with N’s, as input for MrBayes 3.2.2 (40⇓–42) using a GTR+γ nucleotide substitution model as determined by jModelTest 2.1.7 (43). We defined blpH based on genetic association with BlpC; in cases of ambiguity, such as grouping the clade of six blpH variants with the Charlie clade located at position 9 o’clock on Figure 2 within the Charlie blpH clade instead of within the Golf/Hotel blpH clade, we used parsimony with regard to BlpC to assign blpH clades.
Sequence Analysis
We measured the nucleotide diversity and dN / dS ratio of blpH, blpC, as well as the seven housekeeping genes used for MLST analysis (aroE, ddlA, gdhA, glkA, lepB, recP, and xpt) using DNAsp 5.10.01 (45). We separated blpH after nucleotide 687 to measure these values for the receptor and kinase domains separately (26).
We extracted the region 1500 bp before blpT to 100bp after blpB from the 4,096 randomly sampled genomes from a previous alignment with R6_uid57859 (25), which included sequences with interrupted BlpA and BlpB genes (46). We measured nucleotide entropy (measured as Shannon entropy by site) and recombination breakpoints (measured from previously calculated data from GeneConv 1.81a (25, 47) for these sequences. Using only pairwise recombination breakpoints with unique start and stop positions prevented overestimating signs of recombination events caused by the common descent of strains.
Mutual information was calculated using a custom Python script. For each signal, we grouped signals into signal groups, such as Alpha:Bravo:Kilo or Golf:Hotel. We grouped amino acids as either acidic, aliphatic, aromatic, basic, cyclic, or hydroxyl residues. We then normalized mutual information to correct for the entropy of signals and of amino acids at each site.
Transmembrane domains in BlpH were predicted using TOPCONS 2.0 (48).
Bacterial strains and growth conditions
S. pneumoniae strains were grown as liquid cultures in C+Y medium (49) at 37°C and transformed as described previously (21). For selection, S. pneumoniae was plated on Columbia agar plates supplemented with 2% defibrinated sheep blood (Johnny Rottier, Kloosterzande, Netherlands) and 1 μg/ml tetracycline, 100 μg/ml spectinomycin or 0.25 μg/ml erythromycin, when appropriate. E. coli was grown in LB medium with shaking at 37°C or plated on LA containing 100 μg/ml ampicillin.
Strain construction
Strains and plasmids used in this study are listed in Table S2.
Constructs for expression of blpSRH from different strains in S. pneumoniae D39
The blpSRH genes, including the constitutive blpS-promoter (21), was amplified from the genome of S. pneumoniae strains D39, PMEN-18 and Hermans-1012 using primers blpS-F-ClaI-SphI and blpH-R-Hermans-1012-NotI-SpeI, from PMEN-2, using primers blpS-up-F-PMEN2-SphI and blpS-down-R-PMEN2-SpeI-NotI from PMEN14 with primers BlpS-PMEN14-F-SphI and BlpH-R-Hermans-1012-NotI-SpeI and from Hermans-33 with primers blpS-F-ClaI-SphI and blpH-Hermans33/35-R-NotI-SpeI. The PCR products were digested with SphI and NotI and ligated into the corresponding sites of plasmid pJWV25 (between the bgaA homology regions) and transformed into E. coli DH5α. The resulting plasmids were verified by PCR and sequencing. The plasmids were then transformed into S. pneumoniae. Correct integration of the PblpS-blpSRH constructs into the non-essential bgaA-locus was verified by PCR. Primers used for these constructs are listed in Table S3.
Deletion of blpSRHC
The native blp-regulatory genes (blpS, blpR, blpH, blpC) of S. pneumoniae D39 were deleted by replacement with an erythromycin-resistance cassette as described previously (21).
Reporter constructs
Two different blp promoters were used to monitor blp-expression; PblpK, controlling expression of the bacteriocin blpK and PblpT, controlling expression of the functionally uncharacterized gene blpT. The PblpK and PblpT promoters have previously been shown to be co-regulated (21) and act as reporters for BlpR activation across the tested pneumococcal strains (as there is almost no variation in the DNA-binding motif of different BlpR). The reporter constructs PblpK-luc-gfp-lacZ and PblpT-luc-gfp-lacZ (a tripartite reporter cassette) integrated into the non-essential cep-locus of S. pneumoniae D39, has been described previously (21).
Luciferase assays
Luciferase assays were performed essentially as described before (21, 50). Briefly, S. pneumoniae cultures pre-grown to OD600 0.4 were diluted 100-fold in C+Y medium (pH 6.8) with 340 μg/ml luciferin. Luc-activity was measured in 96-well plates at 37°C, and OD600 and luminesencence (as relative luminescence units, RLU) were recorded every 10 minutes using Tecan Infinite 200 PRO. Synthetic peptides (BlpCs) were purchased from Genscript (Piscataway, NJ). Different concentrations of BlpCs were added to the culture wells after 100 min or in the beginning of the experiment, depending on the experiment. The data was plotted as RLU/OD over time to analyze induction of blp expression.
LacZ assays on agar plates
LacZ assays were performed on C+Y agar plates (pH 7.2) covered with 40 μl of 40 mg/ml solution X-gal (spread on top of the plates). All strains were pre-grown to OD600 0.4, before 2 ?l of the wild-type strains (BlpC producers) were spotted and allowed to dry. Then 2 ?l of the different reporter strains were spotted next to the dried spot. The plates were incubated at 37°C over-night.
For induction with synthetic BlpC, C+Y agar plates were covered with 40 μl of 40 mg/ml solution X-gal and 5 μl 1 mg/ml BlpC (spread on top of the plates), and different reporter strains were spotted on top. The plates were incubated at 37°C over-night.
Stochastic Model
We built a spatial, stochastic model in which cells are modeled individually and interact in a grid. At each discrete time point, each cell can divide with probability 80%, which produces an identical offspring in an empty, randomly chosen adjacent position on the grid. Each cell also secretes one of four signaling molecules, which accumulate and diffuse in the space around the cell (defined as “diffusion area”). When the amount of signaling molecules within the sensitivity area (which is an area with half the radius of the diffusion area) of a cell reaches a defined threshold, this cell becomes induced and starts producing bacteriocins. A bacteriocin-producing cell can kill up to six neighboring cells depending on their genotype, as explained below. Induced cells, which produce bacteriocins, grow 20% slower than uninduced cells. Every cell has a 0.1% probability of death at each time point.
We modeled four genotypes, which differ in the signaling molecule and bacteriocins that they produce as well as in the number and identity of signals that they respond to (Table S1). Bacteriocins produced by genotypes 1 and 2 specifically could kill genotypes 3 and 4 and vice versa. Signals produced by genotype 1 could induce genotypes 1 and 2 and similarly, signals produced by genotype 3 could induce genotypes 3 and 4; we therefore classify genotypes 2 and 4 as “eavesdropping genotypes”. Genotypes 1 and 3 can only respond to their own signal, as “signal-faithful genotypes”. All four genotypes have equivalent growth rates, which are only variable depending on if a cell is induced or uninduced. Eavesdropping cells respond to signals that they do not produce with certain degrees of affinity. If we consider the affinity of a cell to its own signal as 100%, we ranged the affinity to the other signals in the case of eavesdropping genotypes as 0% - 90% for different simulations.
We used an initial grid of 400 by 400 positions, and we started with 10% of the grid randomly populated with eavesdropping and signal-faithful phenotypes at a range of proportions. As cells grow and interact, the center of the grid remains occupied, while the cell population can expand on the boundaries past the initial 400 by 400 grid. Simulations were run for until the number of cells reached 110% of initial grid size. We calculated fitness as the difference between relative frequencies of eavesdropping cells at the last time point and at the initial time point, i.e.:

Funding
This work was supported by the Biotechnology and Biological Sciences Research Council (grant number BB/J006009/1) to DER and ISR and by the Wellcome Trust (105610/Z/14/Z) to the University of Manchester. MA is supported by the Biotechnology and Biological Sciences Research Council (grant number BB/M000281/1). Work in the Veening lab is supported by the EMBO Young Investigator Program, a VIDI fellowship (864.12.001) from the Netherlands Organisation for Scientific Research, Earth and Life Sciences (NWO-ALW) and ERC starting grant 337399-PneumoCell. MK is supported by a grant from The Research Council of Norway (250976/F20).
Acknowledgements
We would like to thank Frank Lake for technical assistance.
References




