

Abstract
We discuss here what we feel could be an improvement in future discussions of the brain operating as a Bayesian-Laplacian system, by distinguishing between two classes of priors on which the brain’s inferential systems operate. In one category are biological priors (ß priors) and in the other artefactual ones (α priors). We argue that ß priors are inherited or acquired very rapidly after birth and are much more resistant to varying experiences than α priors which, being continuously acquired at various stages throughout post-natal life, are much more accommodating of, and hospitable to, new experiences. Consequently, the posteriors generated from the two sets of priors are likewise different, being more constrained (i.e., precise) for ß than for α priors.
When, to silent sessions devoted to brain thought,
We summon up formulations from endeavours past,
And sigh the lack of many a principle that we sought,
Because those principles were, in our mind, mis-cast,
Lo, for biological and artefactual priors each has a separate slot
And therefore all priors should not be tied in a single Bayesian knot
A (posterior) Bayesian-Laplacian adaptation from Shakespeare’s Sonnets
I. Introduction
We describe here an approach to the Bayesian-Laplacian system as applied to brain studies which differs from previous approaches, in that it makes a distinction that has not been made before between two fundamental categories of priors. Our hope is that it may constitute a useful contribution to efforts in neuroscience that address the extent to which the brain uses what may be called Bayesian-Laplacian inferential operations. We use the term Bayesian-Laplacian, instead of just Bayesian, because of the very considerable contribution that Pierre-Simon Laplace (1812) made in his treatise entitled Théorie analytique des probabilités (Laplace 1812) to the formulations made in Bayes’ posthumously published paper entitled An essay towards solving a Problem in the Doctrine of Chances (Bayes 1763). By referring to the Bayesian-Laplacian formulation, we thus credit both with this approach.
The Bayesian-Laplacian approach summarizes a fundamental inferential principle in which probabilities of occurrence of events are based on priors which, through experience, lead to posteriors; these, in turn, modify inference (Kersten et al., 2004, Knill and Pouget, 2004, Yuille and Kersten, 2006, Clark, 2013) and behaviour (Friston et al., 2011, Botvinick and Toussaint, 2012, Friston et al., 2015). It is an evolving approach (Laplace, 1812)(Fienberg, 2006), with physiological and philosophical foundations (Helmholtz, 1867)3 (Bovens & Hartmann, 2005) (Talbott, 2008)(Rosenkrantz, 1977) (Gelman & Shalizi, 2013) and probabilistic, statistical, and computational implications (Good et al., 1966) (Bernardo & Smith, 2008) (Gelman et al., 2004).
The application of the Bayesian-Laplacian approach to brain studies promises to shed light on how the brain’s predictive system operates by combining prior knowledge about a phenomenon and modifying it through experimentation and experience. The brain must continually update the hypotheses that it entertains about the world in light of information reaching it and against its current beliefs. Belief, and how it can be modified, thus become fundamental to understanding these operations. One could of course consider all priors under a single category which, subject to experiments or experience, will produce posteriors, as in fact previous discussions of the Bayesian-Laplacian brain have done (Dayan et al., 1995) (Rao & Ballard, 1999) (Lee & Mumford, 2003) (Kersten et al., 2004)(Knill & Pouget, 2004) (Yuille & Kersten, 2006) (Friston et al., 2011) (Botvinick & Toussaint, 2012) (Clark, 2013) (Pouget et al., 2013) (Friston et al., 2015). We, however, propose to distinguish between two sets of priors, the beliefs that they are based on and give rise to, namely: Biological (ß priors) and Artefactual (α priors). The former are regulated largely by inherited brain operations determined by inherited brain concepts (programs) while the latter are subject to acquired (synthetic) brain concepts (Zeki, 2009). This distinction leads us to propose further that the beliefs attached to the two categories of priors must also be distinguished according to the same two categories. The distinction is significant because experience operates on the two categories of priors in different ways. The scope of experience to modify is more limited for ß priors and the beliefs attached to them than for α priors; consequently, the beliefs attached to the ß priors, and the inferences drawn from them, are also more biologically constrained than the ones attached to α priors. In simpler terms, artefactual priors depend upon experience and experiment but biological priors do not. Hence, the central point we are trying to emphasize is that the knowledge-acquiring system of the brain and the predictions that result from it differ qualitatively according to the kind of knowledge being considered.
Our discussion is not exhaustive; rather, we hope that it lays down a basic framework for an alternative approach through which to consider the operations of the brain in a Bayesian-Laplacian context.
II. Definitions
We define the terms we use as follows:
Inherited (biological) priors are the result of brain operations dictated by brain concepts that we are born with; they are resistant to change even with extensive experience and hence must be distinguished from artefactual priors.
Artefactual (acquired or synthetic) priors depend upon concepts which are acquired and formulated postnatally, and which are modified by experience throughout life; they are less constrained than biological priors.
We define the term “concept” separately for the biological and artefactual categories. ‘Concept’ as applied to the former may be defined as an idea dictated and governed by inherited biological laws and used to generate a process that is applied indifferently to incoming signals to generate biological priors. In the example of colour vision, for example, this would be the biologically inherited concept or process of ratio-taking, which is applied to incoming chromatic visual signals and results in a colour category which constitutes the biological prior for colour (see Figure 1). It therefore constitutes a model which generates sensory consequences (colour) from causes. “Colour [category]” in the words of Edwin Land, “is always a consequence, never a cause” (Land, 1985).
Artefactual concepts, for example that of a house, conform more closely to the dictionary definition of “a generic idea generalized from particular instances”. We are not born with the idea of a house nor do we posses a definitive neural system which is dedicated to the generation of houses perceptually, comparable to the ratio-taking process which generates colour categories. Rather, the idea of a “house” is abstract and generated from viewing many houses; the concept continues to grow with experiences derived from viewing new houses. If the brain acquires a generative model of how a “significant configuration” (Zeki 2013) of stimuli corresponding to actual houses generates the perception of a house, it is in a position to recognize these stimuli as corresponding to its previous experience of houses.
Hence, in the Bayesian-Laplacian context, biological priors are much more precise and intransigent than artefactual ones, even though both can lead to almost limitless posteriors. Therefore, the beliefs that are attached to biological priors are more widely shared (i.e., conserved over conspecifics), more independent of culture and learning and also less yielding to experience than artefactual priors.

A schematic representation of the relationship of the ratio-taking process (concept) (R) to the generation of the experience of the colour category (C). The colour category thus generated can then act as a prior for generating a posterior (H) through experimentation (E); this latter posterior (H) differs in the shade of colour (hue) from C. R (the ratio-taking process) determines C because it is independent of any experiment (for further details see text). (Right panel) Representation of the processes involved in generating an artefactual prior. This consists of an initial prior I, for example the chance (50%) that any one of the two candidates will win an election (in the absence of any results)(see text), E represents an “experiment” when one ballot paper has been opened, thus forming a posterior P. When the next ballot paper is opened, this posterior becomes the new prior and along with the result of the second ballot a new posterior will be formed, and so on. Note that, in the left panel no experiment is involved in the appearance of the initial hue H, while in the right an artefactual prior always involves an experiment. Also note that C is determined biologically whereas I is based on learning and assumption.
III. The Bayesian-Laplacian Equation and Its Modification
In the context of colour vision, which we discuss extensively below, the biological (β prior) is a colour category (for example green) generated through a ratio-taking mechanism R, which is essentially identical in all humans. The colour category C, with a specific hue H attached to it, constitutes the prior from which posteriors are generated with experience and experimentation. A modified Bayesian-Laplacian formula to handle cases where experiments are not involved in forming the initial posterior (which is true of all biological priors) is as follows (using colour as an example):

 where
where  , the posterior (percept) of initial hue
, the posterior (percept) of initial hue  depends only upon the prior
depends only upon the prior  (colour category), and hence is independent of E (experiment); and [
(colour category), and hence is independent of E (experiment); and [ |E], the posterior (percept) of colour category, depends only upon [R], the ratio-taking scheme described below, and is independent of any experiment. Further mathematical proofs are given in the Supplementary Materials.]
|E], the posterior (percept) of colour category, depends only upon [R], the ratio-taking scheme described below, and is independent of any experiment. Further mathematical proofs are given in the Supplementary Materials.]
Our equation thus differs from the classical Bayesian-Laplacian equation below:
 (where V and E are two random variables and [V|E] is the probability of event V occurring if E is true; [E|V] is the converse and denotes the probability of observing event E if V is true, [V] being the prior) since the posterior in this equation is dependent on experience.
(where V and E are two random variables and [V|E] is the probability of event V occurring if E is true; [E|V] is the converse and denotes the probability of observing event E if V is true, [V] being the prior) since the posterior in this equation is dependent on experience.
The classical Bayesian-Laplacian approach does not distinguish biological priors from artefactual ones; it can be illustrated by an example taken from a presidential election system that uses two rounds, which can also be used as an example of an artefactual prior. In the first round, any number of candidates is eliminated, leaving the two top scoring candidates for the second round. Since this is a run-off voting system, the elected president always has a majority of the votes. Consider now two presidential candidates A and B, and 20 electoral regions. For simplicity of demonstration, we randomly and sequentially select one ballot from each region, making a total of 20 ballots for predicting the presidential election result (which we wish to do before the results are announced). The initial prior, I, here is the chance (50%) that any one of the two candidates will win an election (in the absence of any results). The opening of one ballot paper constitutes an experiment E through which a posterior P is formed. Let us suppose that when the first ballot is read, the vote goes to A; our belief is now updated about the chance of A winning the election. This new belief then becomes the new prior. Now we read the second ballot, which constitutes a new experiment [E]. If it too is a vote for A, then the updated probability of A wining the presidency becomes higher. We continue to update our expectation with every new vote.
Then the classic Bayesian-Laplacian theorem states that the odds [V] that A will win the presidency increases with every new ballot [E] voting for A. V constitutes the prior (the prior belief about A wining the race) and the outcome of each ballot constitutes the experiment(s) [E]. The posterior [V|E] is the updated guess of the probability of A wining the presidency.
IV. The need for distinguishing biological from non-biological priors
The classification of priors into two broad categories proposed here is based in part on the Kantian system and in part upon our modification of it (Zeki, 2009). Kant wrote in The Critique of Pure Reason (Kant, 1781) that, “perceptions without concepts are blind”, arguing that all inputs into the mind (in our case the brain) must be somehow organized by being interfaced through concepts. In The Critique of Judgment, he nevertheless proposed that some sensory inputs are not interfaced through concepts; among these were signals from objects that could be categorized as beautiful, as opposed to those which had a utilitarian value. With the former the perceiver usually supposed (believed) the operation of a universal belief through which what s/he had perceived to be beautiful would also be perceived to be beautiful by others; in our example of colour as a biological category, we also suppose that, given specific conditions, a perceiver can make the reasonable assumption that the colour that s/he perceives is the same as the colour that others (and indeed all humans) will perceive under the same conditions, and hence that the percept has universal validity. We differ from Kant’s classification by supposing that all percepts, be they biological or not (even including those pertaining to beauty) are interfaced through concepts. But we distinguish percepts that are grounded in inherited β priors from the post-natally acquired a priors, which are acquired postnatally (Zeki, 2009). There is good reason to suppose that the inherited priors, which make (Bayesian - Laplacian) sense of the sensory inputs into our brains, are much more similar between humans and also far less dependent upon culture and learning than the acquired ones. Hence one cardinal distinction between the two sets of priors is that an individual can reasonably suppose that a biological prior, such as colour, is an experience that s/he shares with the great majority of other individuals, regardless of race or culture, a characteristic not shared by the artefactual priors (e.g. appreciation of sushi, or the beauty of a temple or a cathedral).
A distinction of the kind we propose here has not been made before. Our hope is that the differentiation we thus make will be a stimulus for further discussion on how the brain handles these two different categories. There are, indeed, probably other attributes that fall into the biological category, for example that of motion, but we do not discuss these extensively (see, for example, Series & Seitz 2013). Rather, we give examples of what most would agree fall into different categories – colour and faces on the one hand, and the many artefacts such as buildings or cars on the other. Other beliefs, quite distinct from objects, also fall into the artefactual category; we have given a certain system of presidential elections as an example above; another example would be the supposition that a government with a hardline policy on health may, if elected, lead to a rapid change in the value of companies producing medicines. Much of the distinction that we make is based on common human experience. Therefore, although there may not be common agreement on the ontology of biological and artefactual priors, their phenomenology speaks to a clear qualitative distinction. We are effectively claiming that biological priors – that are conserved over generations and cultures – have a greater precision than the more accommodating (empirical, artefactual) priors we call on to assimilate experience that is unique to our time and place.
We address the distinction between biological and artifactual priors in terms of visual perception, about which relatively more is known and with which we are better acquainted. We give two examples, among many, of β priors, those belonging to colour and face perception.
V. Colour vision
V A: Colour as a β prior generated from an inherited brain ratio-taking concept
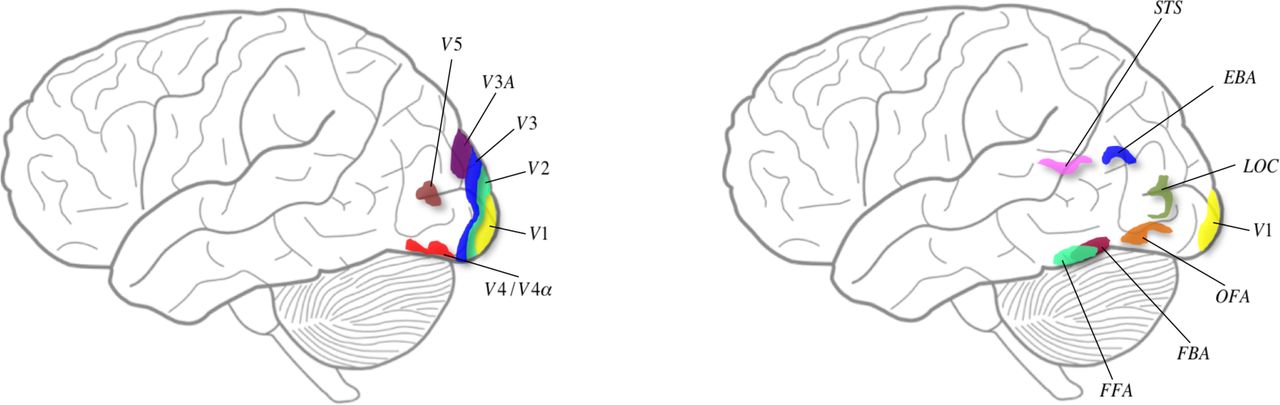
Colour represents perhaps the most extreme form of a β prior which is the result of an operation dictated by an inherited concept, that of ratio-taking, which leads to constant colour categories (see below). Colour is an experience, which we refer to as its β prior and from which hues may be generated which, though belonging to the same colour category, differ in appearance (in shade of colour). These hues become posteriors and act also as new priors from which further hues (posteriors) can be generated through experiments and experience. Colour categories and the hues attached to them are also biological signalling mechanisms allowing the rapid identification of objects, biological or otherwise, by one of their characteristics. If the colour of an object or surface were to change with every change in the illuminant in which it is viewed, then colour would no longer be a useful biological signalling mechanism, because the object can no longer be identified by its colour alone. To make of colours an autonomous identifying mechanism, they must be stabilized and be immune, as far as possible, from the de-stabilizing effect of a change when the wavelength composition of the illuminant in which objects and surfaces are viewed changes. Physiologically, the brain undertakes a ratio-taking operation to render colours constant, and hence stabilize the world. Evidence shows that there are specific brain pathways and a specific visual area, area V4 and the associated V4a, that are crucial for colour perception(Zeki, 1973)(Zeki, Watson, & Lueck, 1991)(Bartels & Zeki, 2000) (see Figure 3), damage to which leads to the syndrome of cerebral achromatopsia (Meadows, 1974)(Zeki, 1990). It is important to emphasize that V4 not only responds specifically to categories of colour but also to hues, or shades of colour (Zeki, 1980)(Stoughton & Conway, 2008) (Brouwer & Heeger, 2013) and that the representation of colour within V4 can be independent of form (Zeki, 1983)(Lafer-Sousa, R, Conway, BR, Kanwisheer, 2016). It is therefore likely that area V4 is pivotal to these operations (Bartels & Zeki, 2000), which is not to say that it acts in isolation; it does so in co-operation with the areas it receives signals from and projects to, together with the reciprocal connections between these areas. Colour is thus an inherited prior which is severely constrained (see below) even though, paradoxically, it also allows an almost unlimited variety of chromatic experiences and therefore posteriors (in terms of hue - see below).
V B. The brain’s ratio-taking system for generating constant colour categories
Colour is a brain construct (Zeki, 1984), which is a prior generated through an operation based on an inherited concept (or program), that of ratio-taking. Ratio-taking compares the wavelength composition of light reflected from one surface with that reflected from surrounding surfaces, thus providing a ratio for light of every waveband reflected from a viewed surface and from its surrounds(Land, 1974) (Land, 1986) (Land & McCann, 1971)(Zeki, 1984). Note that we can refer to the colour category as the prior, i.e. the product of an operation that involves ratio-taking; the hue attached to this prior then becomes the posterior which can act as a prior for generating further hues. We show below that, using the argument presented here and the mathematical proof given in Equation (1) of the supplementary materials, this process is independent of any experiment; namely, a colour category depends only on the ratio-taking operation (see Figure 1).
The ratios thus produced never change (see Figure 1) – they constitute fundamental invariants in a world composed of colourful objects. It is useful to discuss briefly here why this should be so in the context of what the constants in nature are, and how constant colour categories are generated from these constants in nature.

The Bayesian-Laplacian brain operations for colour vision. R refers to the ratio taking scheme leading to our perception of the colour category C, which acts as the biological prior for hue (H) (left panel). When the wavelength composition of the illuminant changes from t0 (corresponding to the initial illumination) to t1 (corresponding to the formation of the color category), the color category C remains the same but the hue changes to H2. For details, see text and for proof of the equations see supplementary material.
The unvarying property of surfaces in terms of colour vision is their reflectance, namely the amount of light of any given waveband – in percentage terms – that a surface reflects in relation to the light incident on it. For a given surface, this percentage never changes. Hence, one finds that the ratio of light of any waveband reflected from a given surface and from its surrounds also never changes, regardless of the variation in the amount of light reflected from an individual surface. If the intensity of light of any given waveband reflected from a surface is increased, the intensity of light of the same waveband coming from the surrounds also increases, and the ratio thus remains the same. By extension, the ratios of light of all wavebands reflected from a surface and from its surrounds also never change.
Take as an example a green surface which forms part of multicoloured (natural) scene, as in Land’s colour Mondrians, and thus surrounded with many patches of other colours, with no patch being surrounded by another patch of a single colour (Figure 1 shows a much simplified version). Let us suppose that the green patch (g) reflects x per cent of the long-wave (red) light, l, incident on it, y percent of the middle wave (green) light, m, and z percent of the short-wave (blue) light, s. The surrounds, having a higher efficiency for reflecting long-wave light, will always reflect more and there will be a constant ratio in the amount of red light (l) reflected from the green surface and from its surrounds. Let us call this ratio gl. The surrounds will have a lower efficiency for reflecting green light (m) and hence there will be another ratio for the amount of green light (gm) reflected from it and from the surrounds, and a third ratio for the amount of blue (s) light gs 4. When the same natural scene is viewed in light of a different wavelength composition, the amount of light of different wavelengths reflected from a surface and from its surrounds will change, often significantly (as, for example, when a scene is viewed successively in tungsten light, in fluorescent light or in sunlight). But the ratios of the amount of light of different wavebands reflected from the centre and from the surrounds remain the same. Formally, let us consider the green patch viewed under two different illuminants, where the first one has gl1, gm1, and gS1 amounts of long, middle, and short wave light reflected from the green patch, and the second one has gl3, gm3, and gS3 reflected from it. The first illuminant results in (gl2, gm2, and gS2) and the second in (gl4, gm4, and gs4) ratios of long-, middle-, and short-wave light reflected from the green patch and from its surrounds, respectively. Then, mathematically, we have:  where the ratios x, y, and z remain the same, regardless of the precise wavelength-energy composition of the light reflected from the green patch and from its surrounds.
where the ratios x, y, and z remain the same, regardless of the precise wavelength-energy composition of the light reflected from the green patch and from its surrounds.
The brain likely uses such a ratio taking operation to construct constant colour categories; these categories are the β priors. There is no physical law that dictates that such ratios should be taken; it is instead an inherited brain law and, given the widespread use of colour constancy in the animal kingdom, we make the assumption that similar mechanisms, or very nearly so, are used in species as far apart as the goldfish (Ingle, 1985) and the human (Land, 1974) and that the results of such operations have universal validity and are therefore independent of culture and learning.
No amount of visual experience can modify the colour categories (β priors); in fact they cannot even be modified by higher cognitive knowledge. For example, green leaves reflect more green than red light at noon and more red light than green at dawn and at dusk; but they are always perceived as green, even in the face of knowledge that they are reflecting more red light under certain conditions. Hence the β priors in colour are extremely stable and un-modifiable with experience. Technically, they are endowed with precision and priors so endowed (as are biological priors) exert a more constrained effect over posteriors. We use precision here in a technical fashion to denote the confidence afforded to priors; precision is the inverse dispersion or uncertainty encoded by a probability distribution (e. g., the variance of a Gaussian distribution). We define precision as 1/variance. In normal colour vision, given the B prior, the variance is very close to 0 and the precision therefore approaches an infinite value.
V C. The experience (experiment) with (constant) colours
This does not mean to say that the shade or hue of the green patch remains constant. The hue will change (a) with every change in wavelength composition and (b) can be much influenced by the surrounds. Thus, if the green surface is reflecting more middle wave light than long and short wave light, it will appear a lighter green than when it is reflecting more long-wave than middle or short wave light, when it will appear a darker green (Figure 1, right). Similarly, through the process of colour induction, the saturation of the green surface can be enhanced by surrounding it with red patches exclusively. In brief, the constant colour category (green) constitutes the primal experience of colour (β prior), while the hue constitutes the posterior derived from it; the direction in which the posterior (hue) changes can be predicted with high accuracy through experience (see below) and it is indeed this knowledge gained through experience and experimentation that artists use constantly. It is interesting to note that the cortical response to colour in pre-linguistic infants (5-7 months old) measured by near infra-red spectroscopy indicates that there is a significant increase in activity in occipito-temporal regions (presumably including area V4) with between-category (colour) alterations but not with within-category (hue) alterations (Yang et al., 2016); similar results have been reported in other studies comparing infants and monkeys (Bornstein et al., 1976). The view expressed here, that constant colour categories are the priors and that hues are the posteriors is consistent with that the results of these experiments.
Another posterior is provided by attaching colours to definite shapes or conditions. Let us take green as a β prior. In our culture, green is linguistically attached to a number of objects and these, through experience, become posteriors. Thus leaves are normally green, but there could be conditions in which they are not. When the Fauvists wanted, in their words, to “liberate” colour to give it its maximal emotional intensity (see Zeki, 1999), they simply attached unusual colours to common objects such as trees or rivers. Here the new and unaccustomed colour-form combination becomes the posterior, with the artist confident in the knowledge that the β priors can be used to generate such posteriors. This usage does not affect the β priors; it simply changes the way in which they are used to modify the posterior.
We propose the following Bayesian-Laplacian Brain Theorem (see Appendix for mathematical details), which summarizes the above example theoretically: the β prior (e.g. the constant colour category generated from the ratio-taking operation detailed above) generates a posterior of any quantity of interest (for example, hue in the above instance). Let us refer to this as  , where the subscript G refers to the green colour category, and the superscript 0 indicates that it has an initial hue. The experiment conducted (for example, adjusting the illuminant in a colour experiment or surrounding the green patch with a patch of a single colour, say red) will lead to a posterior hue which, though still belonging to the prior colour category of green, will differ in its shade of green (and hence hue) from
, where the subscript G refers to the green colour category, and the superscript 0 indicates that it has an initial hue. The experiment conducted (for example, adjusting the illuminant in a colour experiment or surrounding the green patch with a patch of a single colour, say red) will lead to a posterior hue which, though still belonging to the prior colour category of green, will differ in its shade of green (and hence hue) from  . We will refer to this as
. We will refer to this as  . As one experiences different shades of green (different hues) when one views the same scene in different illuminants or, in the example of the green patch above, changes its surrounds, and notes the nature of changes in the illuminant and/or the surrounds of the green patch being studied, so any number of different shades (hues) of green can be generated and experienced. These posteriors (
. As one experiences different shades of green (different hues) when one views the same scene in different illuminants or, in the example of the green patch above, changes its surrounds, and notes the nature of changes in the illuminant and/or the surrounds of the green patch being studied, so any number of different shades (hues) of green can be generated and experienced. These posteriors ( , where i = 0,1,2,…,) can be updated continuously and iteratively in one’s life; knowing more about the illuminant or the spatial configuration of a stimulus, one can therefore make inferences with a high degree of accuracy and reliability.
, where i = 0,1,2,…,) can be updated continuously and iteratively in one’s life; knowing more about the illuminant or the spatial configuration of a stimulus, one can therefore make inferences with a high degree of accuracy and reliability.
V D. The ‘belief’ with respect to colours
A definition of ‘belief’ might be adapted from its ordinary dictionary definition, namely “a feeling of being sure that someone or something exists or that something is true” (Webster’s Dictionary), or “confidence in the truth or existence of something not immediately susceptible to rigorous proof (Dictionary.com) or that “the experience (of colour) will always be true”, even when we are not remotely aware of the operations that lead to the experience. The belief with respect to colours is subtle; it consists of unconscious knowledge and can be illustrated with respect to the green patch referred to above. Helmholtz introduced the concept of the “unconscious inference” to account for this but he qualified it in a way from which we depart, for he supposed that judgment and learning enter into the “unconscious inference”. We believe that the inference is due to an automatic, inherited application of a concept, that of ratio-taking (but also see above). A viewer ‘knows unconsciously’ that the green patch will look green if it reflects more green light than its surrounds, regardless of the actual amount of green light reflected from it and regardless of whether s/he is acquainted with the object or had never seen it before, thus precluding judgment and learning. Anyone armed with such knowledge can predict the colour of a surface, even before seeing it. Thus, a red surface is one that will reflect more red light than its surrounds and a blue surface is one that will reflect more blue light than its surrounds. A white surface will reflect more light of all wavebands than its surrounds while a black one will reflect less light of all wavebands than its surrounds, regardless of the actual amount of light reflected from it. This belief system is quite rigid and not easy to manipulate. Moreover, any perceiver can make the reasonable assumption that the colour s/he perceives under any given condition will be the same for all other perceivers under the same condition, regardless of culture and upbringing.
This can be readily established by matching the colours perceived with Munsell chips (McCann et al., 1976). Hence the belief is universal. Because of this assumed universally shared belief, Kant would no doubt have referred to this as an experience which is not interfaced through a (prior) concept. But, as we have shown, it is in this instance an experience generated through a ratio-taking operation, executed by the brain to stabilize the world in terms of colour categories. We leave out of account here the vexed and unsolved problem of qualia, of whether the quality of green that one person perceives is identical to that perceived by another.
It is also worth pointing out that knowledge that a green leaf reflects more red light (as is common at dawn and dusk) will not, and cannot, modify one’s experience of its colour as green, provided that the leaf is being viewed in a natural context (thus allowing the brain’s ratio-taking system to operate), again calling into question Helmholtz’s supposition that judgment and learning are critical for determining the colour category. In this sense, the experience and the belief attached to it are biologically constrained.
VI. Faces - a category of B priors
It is generally agreed that there are special areas of the brain that are necessary for the perception of faces, including an area located in the fusiform gyrus known as the fusiform face area (FFA) (Sergent et al., 1992)(Kanwisher et al., 1997) (see Figure 3) ) damage to which leads to the syndrome of prosopagnosia. We note that the FFA is active when faces are viewed from different angles, hence implying a certain degree of face constancy (Pourtois et al., 2005). Another area critical for faces is located in the inferior occipital cortex and known as the occipital face area (OFA) (Peelen & Downing, 2007)(Pitcher, 2014) while a third area, located in the superior temporal sulcus, appears to be important for the recognition of changing facial expressions (Haxby et al., 2000). These may not be the only areas that are important for face perception. It has been argued that the recognition of faces engages a much more widely distributed system (Ishai et al., 2005); it has also been argued that cells responsive to common objects, in addition to faces, can be found in an area such as FFA. Whatever the merits of these contrasting views, they do not much affect our argument, given the heightened susceptibility of faces to distortion and inversion and the relative resistance of objects to similar treatment (Zeki & Ishizu, 2013, for a review); this would argue in favour of our general supposition that β priors (to which faces belong) must be separated from α priors (the artefactual category), whether the representation of objects and faces occurs in the same or in different brain areas (for a general review see Zeki & Ishizu, 2013).
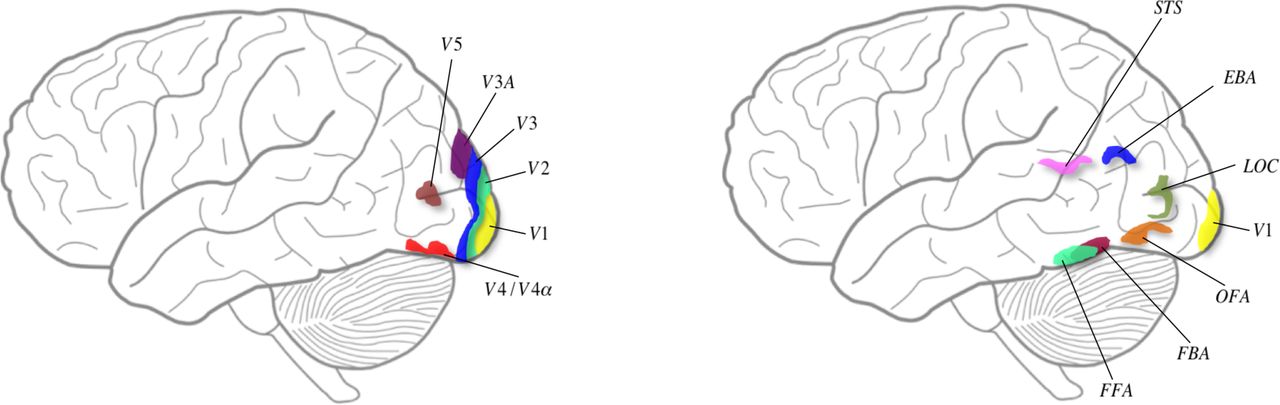
(Left) Schematic diagram depicting some of the classical visual areas on a surface drawing of the brain. The position of the visual areas is approximate. (Right) Schematic diagram to show visual areas considered to be critical for face and body recognition. The position of the areas is approximate.
It is generally also agreed that the capacity to recognize a certain “significant configuration” (Zeki, 2013) as constituting a face is either inherited or very rapidly acquired, within hours after birth (Goren et al., 1975)(Johnson et al., 1991), although there has been much discussion as to what it is in the configuration that is instantly recognizable (see discussion in (Zeki & Ishizu, 2013). It is significant that this preference of the new-born for looking at faces is not found when line drawings of real faces are used (Bushnell et al., 1989) emphasizing the pre-eminence of the biological concept of face.
Any departures, even minor ones, from the significant configuration that constitutes the (possibly) biologically inherited and accepted concept of a face is rejected and never incorporated into the concept of a normal face. The cortical response to faces is itself very exigent in terms of the “significant configuration” that it will respond to optimally; mis-aligning the two halves of an upright face delays and increases the typical N170 negative deflection obtained following facial stimulation, but this delay and increase are not quite as strong for inverted faces (Ishizu et al., 2008), which are immediately classified as having an abnormal configuration, and thus not belonging to the biological category of faces. It is, we think, very difficult to produce a biologically viable posterior, and a belief attached to it, from an inverted face (unlike buildings – or other artefacts – see below) and, even if produced, is unlikely to be durable. The same is true of the expression on a face, with certain expressions being immediately recognizable as comforting or loving and others leading to different emotional apprehensions. The posterior that results from this β prior through experience is thus similarly circumscribed, since any departure (as produced by inversions, for example) would mean that the brain will either not classify it as a normal face, or that it will only be temporarily classified as a face, or that it will be classified as an abnormal face, without leaving a permanent posterior.
Any posterior generated from a face β prior must therefore be strictly linked to what is a normal significant configuration of what constitutes a face. A child, for example, begins to learn to associate certain expressions on a normal face with certain social interactions – whether, for example, someone is enjoying one’s company or is bored by it, whether small inflexions represent doubts or threats, and so on. But it is unlikely to associate the expressions on an inverted face with a permanent posterior, since an inverted face disobeys the inherited brain concept of the significant configuration that constitutes a face. This does not mean that posteriors related to faces cannot be of an unusual nature – for example a continual smile linked to wicked behaviour on the part of an individual may lead the perceiver to establish a different posterior from the same β priors for that individual. But here again, a perceiver is unlikely to form a permanent posterior of a smile linked to wickedness if the face is mis-aligned or inverted.
It is interesting to note that, in his effort to give what he called “a visual shock”, the English painter Francis Bacon subverted the brain’s β priors in terms of faces, and took to painting highly deformed faces, which depart significantly from the significant configurations that constitute the normal β priors for faces (Zeki & Ishizu, 2013); however prolonged the viewing, these never become accepted as normal faces. Indeed, the viewing of stimuli in which inherited concepts of face (and space) are deformed and violated leads to significant activation in frontoparietal cortex, whereas the viewing of “deformed” or unusual configurations of common objects such as cars do not. Even daily exposure to deformed faces for one month does not lead to a significant change in activation patterns produced when viewing them, suggesting that inherited biological concepts of face are stable at the neural level, at least within a time frame of one month (Chen & Zeki, 2011). This neurobiological demonstration is consistent with our proposed subdivisions of priors into the biological and artefactual categories.
VI A. The ‘belief’ attached to faces
We now outline in general terms the biologically based initial ‘belief’ attached to normal faces. It is constrained by the fact that a face must contain a certain number of features such as eyes, nose, mouth etc, set out within certain proportions and symmetrical relations to each other which, together, constitute a significant configuration typical of a face. An absence of any of these features or any significant violation of these proportions or relations will automatically depart from such a belief, and lead to its classification as abnormal. There are of course many ways in which faces can be represented; they can, for example, be represented in terms of straight lines in a drawing. But such, though recognized as representing a face, will be immediately classified as a drawing and therefore not a biological face. This shows how constrained such a belief and the β prior attached to it is. In terms of generality, one person’s belief that the object s/he is seeing is a face and that all other human subjects viewing the same stimulus will also perceive a face in that configuration is a sound one and makes that belief general. Just like colour, it therefore has universal validity and is independent of culture and education.
VII. Artefactual (a) priors
By artefactual priors, we refer to the many constructs - from houses and cars to ordinary utensils and tools - for which there is no inherited brain concept. Instead, the brain acquires a concept of these objects through experience and consequent updating of priors; these are continually modified throughout life and are also strongly culture dependent. In medieval times, people had no concept of a car or a plane. Since their introduction, there have been many modifications of these constructs, and the concepts attached to them have changed accordingly. The concept of a plane that someone living in the 1930s had, for example, did not include jumbo jets equipped with jet engines; these have been added to the overall concept of a plane since. There are, of course many other examples one could give, including the use of knives and forks and chop-sticks, which differ between cultures and times.
We also include in the category of artefactual priors more general ideas pertaining to, for example, economic theories or political beliefs or systems, examples of which we have given above.
The formation of such concepts is strongly dependent on experience and culture, which distinguishes them from biologically inherited concepts (see Figure 2); being culture- and experience-dependent they should, by definition, have greater variance. Crucially, acquired priors emerge de novo and are driven by experiences that are unique to any individual in any given lifetime, although there may be, and usually are, population level similarities. They therefore are necessarily less precise and more accommodating than biological priors. This follows because they are designed to be modified by experience.
It is now generally accepted that there is a complex of areas, known as the lateral occipital complex (LOC) which is critical for object recognition (Grill-Spector, 2003). Even though it has been argued that the so-called face areas may not be as specific to faces as originally supposed (see above), and that cells in them may encode objects as well, including ones which we would classify under artefactual categories, the differential response to faces and objects when inverted suggests that they are processed differently. Moreover, neural sensitivity to faces increases with age in face-selective but not object-selective areas of the brain, and the perceptual discriminability of faces correlates with neural sensitivity to face identity in face selective regions, whereas it does not correlate with a heightened amplitude in either face or object selective areas (Natu et al., 2016). There is no definitive evidence about when infants begin to recognize objects or whether they recognize faces before recognizing objects. Indeed, it has been shown that infants can recognize differences between shapes even at one month where the outside contour/shape is static and identical, but where the inside smaller shapes are different to each other in each image if, significantly, one of the smaller inner shapes is jiggled or moved (Bushnell et al., 1989); this may, in fact, introduce a biological prior, that of motion, into the recognition or inference process (see also Series & Seitz 2013).
Such results, together with common experience, justify a neurobiological separation between the two categories, faces belonging to the biological category and objects to the artefactual.
VIII. A biological prior that makes all artefactual priors possible?
While the emphasis in this article is on separating biological from artefactual priors, it is worth asking whether, at the earliest recorded stages after birth, one can postulate the presence of a general biological prior that leads to artefactual priors, which then assume an autonomy of their own (see also Series & Seitz 2013). The common view is that there is one category of cell in the visual brain, the orientation selective (OS) cell, discovered by Hubel & Wiesel (1962), which is generally considered to be the physiological ‘building block’ of all forms. This is a plausible argument entertained by physiologists as well as artists like Piet Mondrian ( see Zeki, 1999). Evidence from physiological and clinical studies of form perception studies shows that, while the OS cells and hence the machinery for constructing forms must be present at birth, it requires nourishment in the early stages after birth, during a critical period, to mature(Hubel & Wiesel, 1977); depriving cats or monkeys at a critical period after birth blights their visual capacities for considerable periods, perhaps even permanently. Observations in humans deprived of vision at birth through congenital cataracts, with vision restored later in life after successful operations, confirms that visual nourishment during an early ‘ critical’ period is necessary for a normal visual life (for a review, see (Zeki, 1993).
By contrast, a normally nourished visual brain can subsequently recognize and categorize many different shapes, even those that have not been seen before. Hence, one could consider that OS cells constitute the biological priors. In accepting the common supposition that OS cells constitute the physiological ‘building blocks’ from which all categories of objects (including faces) are constructed, one must nevertheless acknowledge that (a) OS cells are widely distributed in different, specialized, visual areas of the brain (Zeki, 1978) and (b) that the OS cells of V1 may not be the sole source for the neural construction of objects. This is especially so since OS cells in visual areas outside V1 survive the destruction of V1 (Schmid et al., 2009), thus showing that their properties are not wholly dependent upon input from V1. Thus, OS cells in different visual areas may contribute to form construction in different ways. Moreover, unlike what is commonly posited, V1 is not the sole source of the ‘feed-forward” visual input to the rest of the visual brain; the specialized visual areas, including areas with high concentration of OS cells as well as areas specialized for face and object perception, receive two further “feed-forward” inputs, from the lateral geniculate nucleus and the pulvinar (Zeki, 2016) and are activated with the same latencies, post stimulus presentation, as V1 (Shigihara & Zeki, 2013)(Shigihara & Zeki, 2014a)(Shigihara & Zeki, 2014b). Hence a strictly hierarchical organization for form (as is commonly supposed), in which cells within the brain’s form system have their sole source in the OS cells of V1 after which they acquire increasingly more complex properties that enable them to respond to complex objects and faces is probably unlikely. Rather, there appears to be multiple hierarchical systems, which operate in parallel and which are task and stimulus dependent (Zeki, 2016).
There is another difficulty in considering OS as being the universal biological prior for the construction of form. There is no single, definitive posterior, that can be built from a single or multiple oriented lines. If faced with either, what would the posterior be? This is quite unlike colour, where certain ratios of wavelength composition of light reflected from a patch and from its surrounds determines, ineluctably, a certain prior in the form of a certain constant colour category, from which posteriors, in the form of hues, can be elaborated. We therefore, believe that we are justified in supposing that orientation selectivity cannot be a biological prior for all forms, as most suppose implicitly. Rather, OS cells in different areas may be used to construct different forms or different categories of form, which then act as distinct biological priors from which different posteriors can be generated. In fact, a line need not be a means toward a more complex form at all; it can exist on its own, as artists have so frequently demonstrated. Moreover, there is no belief that can be attached to single oriented lines, except in the narrow sense that they can constitute, either singly or in arbitrary combination, forms in themselves, as Alexander Rodchenko (1921) argued when he wrote “I introduced and proclaimed the line as an element of construction and as an independent form in painting”. He added, “…the line can be expressed in its own right, as the design of a hypothetical construction [and can have] a status independent of what is actually taking place, and becomes an abstraction” (Rodchenko, 1921) (our ellipsis). Many artists since then have emphasized the primacy of the line in their work.
Hence, there is no universal belief that is attached to how single oriented lines can be combined. There is also no universal belief attached to what significant configuration constitutes a given category of object. The configuration of houses, as places of habitation, differs widely in different cultures - from igloos to huts to skyscrapers and even to inverted pyramidal buildings (see Figure 5 of supplementary material). One cannot make the assumption that huts are the universal mode of habitation or that inverted buildings depart from the concept of habitation. Rather, the latter are absorbed into the concept (i. e. generative models) of habitation through experience.
Conclusion
We have here given a general account of what we believe is an important distinction to be made when considering the brain as a Bayesian-Laplacian system. For simplicity, we have concentrated on extreme examples, ones which we have better knowledge of; namely, that of colours and faces for the β priors and of common artefacts for the α priors. This naturally leaves out of account a vast territory in which both priors may be involved. Laplace himself delved into questions of average mortality and the average duration of marriages. The list can be extended to include social interactions as well as economic activity in which the (unfortunately) un-studied β prior of greed may play a crucial role, in addition to α priors. An example of the latter, which plays a role in economic calculations, is the recognition of political decisions that influence monetary values, which would fall into the artefactual, α, category. In these, and many other human activities that involve making inferences based on a set of beliefs, the distinction between the two categories of priors is, we believe, important.
Finally, the distinction between biological and arteractual priors can also be extended to aesthetics, since aesthetics pertaining to biological entities such as faces or bodies, are similarly constrained by the configurations that constitute them (Zeki 2009)(Zeki, 2013).
We have restricted ourselves here largely to the visual brain, but hope to deal with other brain processes that are subject to Bayesian-Laplacian operations in future papers.
Acknowledgment
We are very grateful to Karl Friston, Will Penny, and Stewart Shipp for their insightful comments on an earlier version of this paper.
Footnotes
↵1 Email: yibing.chen{at}yale.edu
↵2 Email: s.zeki{at}ucl.ac.uk
↵3 In the article, Helmholtz described a view which he called the intuition theory (nativistische Theorie), that “it is necessary to assume a system of innate apperceptions that are not based on experience, especially with respect to space-relations. In the same article, he also stated that “the judgment of the sense may be modified by experience and by training derived under various circumstances, and may be adapted to the new conditions.” He thus came close to distinguishing between the two sets of priors that we discuss here, although his discussion remains vague and does not give or define either a general or specific framework for distinguishing between priors (a term he did not use) or provide a specific framework of how the “unconscious inference” operates in, for example, colour vision, as we do here.
↵4 For brevity, we restrict ourselves to long, middle and short-wave light, without giving the peak values along the spectrum; in practice, and under natural viewing conditions, a surface will reflect light of many wavelengths, but there will be a (constant) ratio for light of any wavelength reflected from the centre and surrounds.
5 Here, O and S are both three-dimensional. For example, 0 = (gl1, gmi, and gSl) and S = (gl2, gm2, and gS2). See Section III B of the article for further details.





{kind=link}
{kind=link}
{kind=link}