

Abstract
Cells use spatial constraints to control and accelerate the flow of information in enzyme cascades and signaling networks. Here we show that spatial organization can be a similarly powerful design principle for overcoming limitations of speed and modularity in engineered molecular circuits. We create logic gates and signal transmission lines by spatially arranging reactive DNA hairpins on a DNA origami. Signal propagation is demonstrated across transmission lines of different lengths and orientations, and logic gates are modularly combined into circuits that establish the universality of our approach. Because reactions preferentially occur between neighbors, identical DNA hairpins can be reused across circuits. Colocalization of circuit elements decreases computation time from hours to minutes compared to circuits with diffusible components. Detailed computational models enable predictive circuit design. We anticipate that our approach will motivate the use of spatial constraints in molecular engineering more broadly, bringing embedded molecular control circuits closer to applications.
Human-engineered systems, from ancient irrigation networks to modern semiconductor circuitry, rely on spatial constraints to guide the flux of materials and information. Cells similarly use spatial organization, through enzyme scaffolds, organelles and chaperones, to perform complex information processing tasks within a crowded intracellular environment. Such spatial constraints play a pivotal role, by accelerating interactions between components that are closer together, and reducing interference between those that are further apart. Not surprisingly, spatial organization has been recognized as a potentially powerful engineering principle for the construction of synthetic molecular circuitry. However, in practice, synthetic circuits have so far relied almost exclusively on interactions between diffusible components guided by chemical specificity1–3. As a result, scaling up such circuits for complex and parallel computation rapidly becomes intractable, due to the limited availability of orthogonal components. Here, we demonstrate a novel design paradigm for realizing scalable molecular logic circuits with a minimal set of orthogonal components, using spatial organization rather than sequence specificity as the main organizing principle.
DNA nanotechnology provides an ideal framework for exploring the use of spatial constraints in molecular circuit design. First, DNA origami forms a uniquely programmable scaffold for the controlled arrangement of molecular circuit elements8. Second, research on DNA-based walking motors corroborates the notion that programmed, multi-step reactions can occur in DNA systems with spatial constraints. Over the last decade such DNA walking motors, mainly powered by enzyme catalysis, have progressed from being able to make a small number of externally triggered steps to autonomously moving along multi-step tracks laid out on a DNA origami9–12. Walking motors could in principle be programmed to perform computation13–15. However, by considering that only information needs to propagate in a computational circuit, rather than a specific motor molecule, we open up a much broader design space for engineering. Third, DNA strand displacement16,17 provides a mechanism for the rational design of complex digital4,18 and analog circuits19,20, neural networks21,22 and reaction diffusion patterns23,24, with quantitatively predictable behaviors. Such DNA strand displacement circuits form a benchmark for success, but also pose a set of challenges that need to be overcome through novel design approaches. In particular, circuit operation is slow at experimentally realistic concentrations. Furthermore, unintentional binding interactions between sequences degrade performance and increase with circuit size, leading to a lack of modularity in circuit design and execution.
Several theoretical papers have proposed DNA circuit architectures that take advantage of spatial constraints to overcome these limitations of speed and modularity25–28. Limiting interactions to spatially proximal circuit elements should result in faster reactions, since spatial localization allows the effective concentrations of components to be substantially increased, compared to components in solution. Moreover, sequences can be reused across components because proximity rather than chemical specificity controls information flow. Recent experimental work has begun to characterize the kinetics of strand displacement reactions with localized components, providing evidence for an increase in circuit speed due to localization29–31, and elementary localized DNA logic gates have also been built32,33. However, an experimental realization of a scalable circuit architecture that exploits the advantages of spatial organization is still lacking.
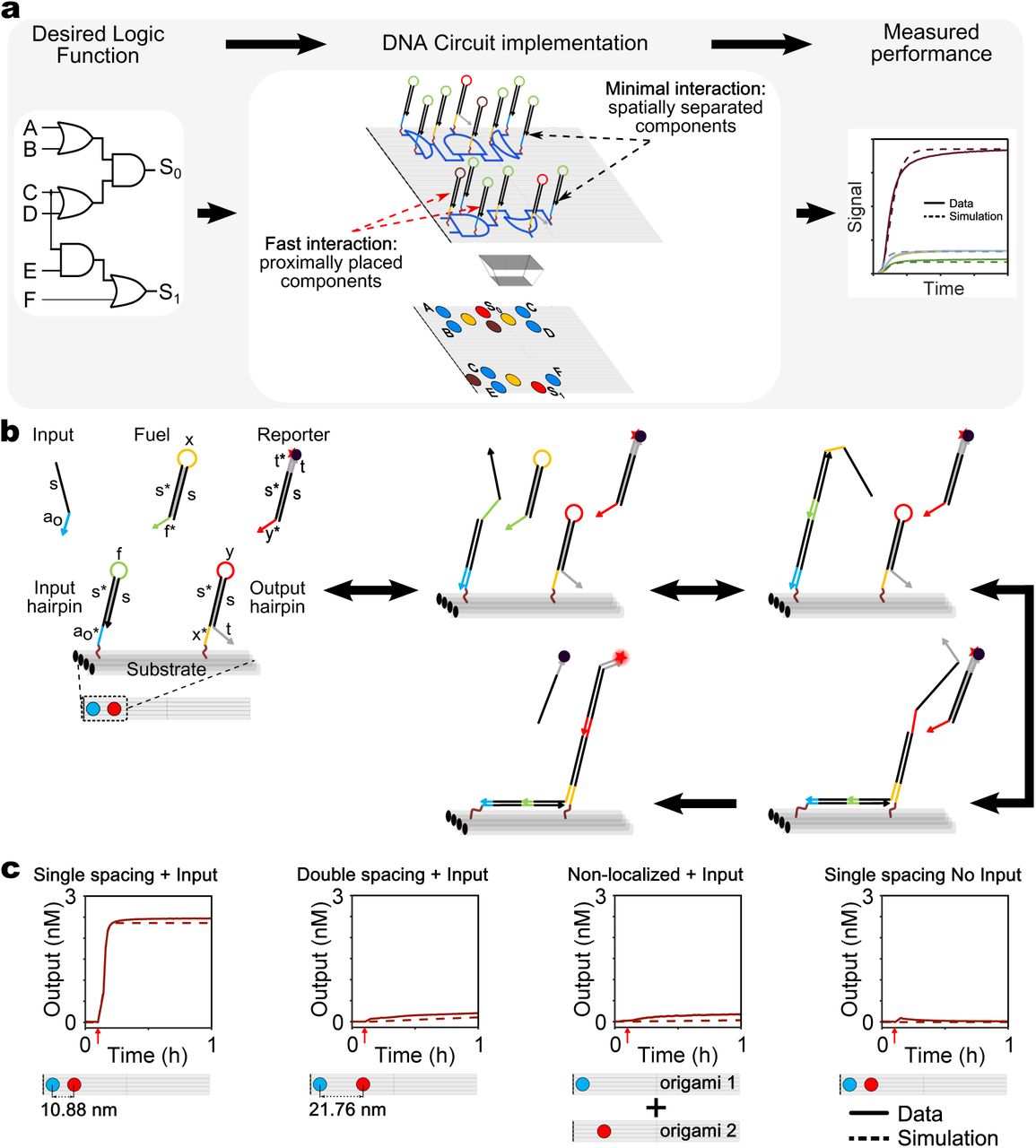
Here we experimentally demonstrate a modular design strategy – the “DNA domino” architecture – that uses spatial organization to realize fast arbitrary logic at the molecular scale. Domino gates and signal transmission lines (wires) are realized with DNA hairpins laid out on a DNA origami scaffold (Fig. 1a, Supplementary Fig. S1). The reaction mechanism underlying circuit operation reimagines the hybridization chain reaction (HCR)34, such that polymerization occurs along designed trajectories on an origami25. All reactions are rationally designed and, unlike most DNA walking motors, no enzyme or ribozyme catalysis is required for operation.
Localized signal propagation mechanism
To illustrate how information is propagated spatially, we consider the “DNA domino effect” in a minimal two-hairpin wire comprised of an Input and Output hairpin attached to a DNA origami scaffold (Fig. 1b). In each reaction step, a hairpin stem is unwound and a toehold that is initially sequestered in the hairpin loop becomes available, to initiate the unwinding of a subsequent hairpin stem. The Input hairpin is opened by binding of an input strand, which enables the capture of a diffusible Fuel hairpin. Requiring a diffusible Fuel ensures that no unwanted reactions can occur between Input and Output hairpins during initial assembly of the origami. In practice, the Fuel can be added in large excess over the concentrations of the other components, and thus does not substantially limit reaction speed. The Output hairpin is opened by the Fuel-bound Input hairpin via a fast local interaction. Finally, the activated Output hairpin displaces the quencher-labeled strand from the diffusible Reporter complex, resulting in increased Reporter fluorescence.

a, High-level abstraction of the circuit design process: An abstract logic circuit (left) is realized using DNA hairpin components arranged on a DNA origami (middle), tested experimentally and compared with model simulation (right). A top view of the localized circuit on origami (middle bottom panel) shows the four basic hairpins reused throughout all circuits in this paper as color-coded circles (Input hairpin (HP): blue; Intermediate hairpin: yellow; Output hairpin: red; Threshold hairpin: brown). b, Reaction mechanism for a two hairpin domino (2HP) wire. Arrows denote 3’-ends. Functional domains are indicated by color and labeled in the first panel (s: stem domain; a0, x, f, y, t: toehold domains; s* represents a domain complementary to s). c, Unquenched fluorophore concentration plots (solid lines) and corresponding simulations (dashed lines) demonstrating signal transfer across localized and non-localized versions of a 2HP wire. Left to Right: Single-spaced 2HP wire; double-spaced 2HP wire; Input hairpin and Output hairpin on different origami; single-spaced 2HP wire without Input. Reactions were carried out at 25°C with 5 nM Origami, 40 nM Reporter, 100 nM Fuel, 50 nM Input in 1X TAE, 12.5 mM Mg++. Red arrows indicate time points when input strands were added.
To demonstrate the benefits of spatial organization, we experimentally compared signal propagation between Input and Output hairpins, positioned at different distances from each other or on different scaffolds (Fig. 1c). We first confirmed that a signal could rapidly propagate across proximally positioned Input and Output hairpins (single spacing) in a two-hairpin wire (t1/2 <3 mins). No observable signal transfer was observed without input addition. We then doubled the distance between Input and Output hairpins (double spacing) on the same origami, and showed that separating the hairpins beyond their theoretical maximum reach resulted in minimal signal transfer (Fig. 1c, Supplementary Fig. S4). Furthermore, we found that interactions between Input and Output hairpins on two different origamis were significantly slower than single-spaced hairpin interactions on the same origami, and comparable to the double-spaced hairpin interactions. Crucially, decreasing the operating concentration of the origamis did not affect the speed of localized intra-origami signal propagation, but significantly reduced the speed of non-localized inter-origami interactions (Supplementary Fig. S5).
We quantified the kinetics of domino circuits by constructing detailed computational models and parameterizing them using experimental data (Supplementary Figures S6 - S18). Interactions involving a diffusible molecule and a tethered hairpin were modelled as bimolecular reactions according to mass action kinetics, while interactions between two complexes tethered to the same origami were converted to unimolecular reactions by scaling with a local concentration27,35,36. We used parameter inference techniques to establish a maximum likelihood parameter set (Supplementary Fig. S15). By ensuring that equivalent interactions in different circuits were parameterized with the same rate constants, we were able to demonstrate consistency in the quantitative behavior of our circuits. In this way, models were used throughout this study for the design and optimization of circuit behavior.
Signal propagation through molecular wires
We created wires of varying lengths and orientations (Supplementary Fig. S1 and Fig. S19) to allow signal propagation over extended distances, by using Intermediate hairpins (Supplementary Fig. S20) as signal relaying components. Multiple identical Intermediate hairpins were positioned at appropriate distances (single spacing) to relay the signal from an Input to an Output hairpin. A schematic of an untriggered eight hairpin domino wire is shown in Fig. 2a. We experimentally observed fast and reliable signal propagation through wires with up to 8 hairpins (spanning over 80 nm) along the origami helical axis (t1/2 < 10 mins.; Fig. 2b), with up to 4 hairpins perpendicular to this axis (t1/2 < 6 mins.; Fig. 2c), and with up to 7 hairpins through a 180 degree turn (t1/2 < 10 mins.; Fig. 2d). We observed significantly reduced signal propagation whenever an Intermediate hairpin was intentionally omitted, and found that signals propagated preferentially via neighbouring hairpins (Supplementary Fig. S21). Signal completion levels varied approximately inversely with track length (Supplementary Fig. S22, Fig. S23), likely due to imperfect incorporation of hairpins into the origami (Supplementary Fig. S24). The non-monotonic decrease in signal can be explained by position-dependent hairpin incorporation efficiency (Supplementary Fig. S25). Signal production due to inter-origami interactions increased with the number of hairpins, but even for an eight-hairpin wire was considerably lower than signal production through localized interactions. As noted previously, lowering the origami concentrations significantly reduced inter-origami interactions, with minimal effect on the speed of localized signal propagation (Supplementary Fig. S26, S27).

a, Schematic representation of an untriggered eight hairpin (8HP) wire along the DNA origami helical axis (domains are labeled with lowercase letters). Bottom panel: Top view of the wire with Input hairpin (blue circle), six identical Intermediate hairpins (yellow circles) and Output hairpin (red circle). Table: Wires of arbitrary lengths can be built with only six unique domains. b-d, Signal propagation along the helical axis (b; 2-8 HP), perpendicular to the helical axis (c; 2-4 HP), and through a 180 degree turn (d; 2-7 HP). Left: Unquenched fluorophore concentration plots (solid lines) and simulations (dashed lines). Experimental conditions were as in Fig. 1. Right: Graphical depiction of all different wires. Hairpin spacing is consistent along the vertical (12 nm), horizontal (10.88 nm) and diagonal (13.5 nm) directions. Black arrows indicate the sequence of signal propagation. Red arrows indicate time points when inputs were added. ds 2HP represents a double-spaced two hairpin wire.
Design and construction of elementary logic gates
As a prerequisite for performing arbitrary logic computation with our domino architecture, we next designed two-input OR and AND gates. The two-input OR domino gate was implemented through a wire fan-in by positioning an Output hairpin close to two orthogonal Input hairpins on the origami (Fig. 3a). Because all hairpin components were constrained to have the same stem, input orthogonality was ensured by using distinct toehold sequences. Experimentally, no significant output fluorescence was observed in absence of both Inputs, but addition of either one or both Inputs resulted in high output fluorescence, consistent with OR logic (t1/2 < 5 mins).

a, Two-input OR; b, Thresholding module; c, Two-input AND. For (a-c), left: Logic gate diagram. second-to-left: DNA domino representation and top view projection. All domains are labeled with lowercase letters and Inputs are distinguished by their toeholds (a0 and b0, blue). The output toehold domain xs* (yellow, 3 or 4nt) is a truncated version of x* (6nt). second-to-right: Graphical summary of circuit operation. For OR and AND gates, the response to different input combinations is depicted. For the threshold module, a circuit with a Threshold hairpin is compared to a circuit without it. Black arrows indicate the (most likely) direction of signal propagation. right: Unquenched fluorophore concentration plots (solid lines) and simulations (dashed lines). For AND and OR gates, legends are represented as truth tables, where presence of an input strand (A,B) or output signal (S) is denoted by “1” and their absence is denoted by “0”. For the Threshold module, the T column indicates whether the system did (+) or did not (-) include the Threshold hairpin, which contains the x* domain (CTGGAC). The xs* column indicates whether the Output hairpin has a 3nt (GAC) or 4nt (GGAC) toehold. Experimental conditions were as in Fig. 1. Red arrows indicate when inputs were added.
To implement a two-input AND domino gate, we used a thresholding strategy to prevent signal propagation when only one of the inputs is present. Specifically, a “Threshold hairpin” was designed to block signal propagation by outcompeting the Output hairpin. We ensured preferential binding to the Threshold hairpin by shortening the Output hairpin toehold, while keeping the Threshold hairpin toehold at 6 nucleotides (Fig. 3b and Supplementary Fig. S28). Once the Threshold hairpin is opened, it captures a partially double-stranded “Threshold block”, making signal inhibition practically irreversible. Experimentally, the extent of preferential binding to the Threshold hairpin depended on the difference in toehold lengths (Fig. 3b), where a 3 nucleotide Output hairpin toehold was found to be optimal. Consequently, all domino logic circuits presented in this study, including the OR gate discussed above, were constructed using Input and Threshold hairpins with 6 nucleotide toeholds, and Intermediate and Output hairpins with 3 nucleotide toeholds.
The two-input AND domino gate was implemented by combining an OR gate with a Threshold hairpin. The Output and Threshold hairpins were positioned at equal distances to two orthogonal Input hairpins (Fig. 3c). When only one of the input strands is added, the Threshold hairpin blocks signal propagation, resulting in low output fluorescence. When both input strands are added, the signal from one of the inputs is blocked, while the signal from the other input successfully propagates to the Output hairpin and generates high output fluorescence (t1/2 < 6 mins.), consistent with two-input AND logic.
The measurements of the two-input OR, threshold and two-input AND gates were included in the parametrization of our computational models. Kinetic parameters for input and fuel binding were shared between all circuits. However, we used distinct local concentration parameters for the wires and logic circuits, since the spacing between hairpins was slightly different in these two settings. The ratio between the binding rates of the Threshold and the Output hairpin was also calibrated to the experimental data, and was approximately 10 to 1 (Supplementary Fig. S15). The simulated model behaviors were consistent with the measured kinetics for the two-input OR, threshold and two-input AND gates (Fig. 3 and Supplementary Fig. S16).
Modular cascading of logic gates to build complex logic circuits
To demonstrate modular circuit design using our domino architecture, we built a three-input AND gate by cascading a pair of two-input AND gates on the same origami. Specifically, we replaced the Output hairpin of an upstream two-input AND gate with an Intermediate hairpin, such that the output of the first gate was relayed to the input of the second (Fig. 4a and Supplementary Fig. S29). Upon addition of any combination of two input strands, the circuit showed low output fluorescence. When all three input strands were added there was high output fluorescence (t1/2 < 6 mins.), consistent with three-input AND logic. Furthermore, the experimental data exhibited the expected Boolean logic. We were able to quantitatively predict the dynamics of the three-input AND gate through model simulation, using the parameters obtained from our previously characterized circuits (Fig. 4a).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
a, Three-input AND gate. b, Two three-input AND gates in parallel. c, Six-input AND gate. d, Two-input dual-rail XNOR gate. left: Logic circuit diagram. middle: Top view of the localized DNA circuit. right: Unquenched fluorophore concentration plots (solid lines) and model predictions (dashed lines). Legends are represented as truth tables, where presence of an input or Output is denoted by “1” and their absence is denoted by “0”. Experimental conditions were as in Fig. 1. Red arrows indicate when inputs were added.
Next, we positioned two distinct three-input AND gates side by side and triggered them simultaneously, to determine whether multiple circuits on the same origami can function concurrently (Fig. 4b). Both gates used the same Intermediate and Threshold hairpins throughout, along with the universal Fuel, but used distinct toeholds for the Input and Output hairpins to ensure orthogonality. Experimental measurements were consistent with model predictions (Fig. 4b).
To further demonstrate scalability, we constructed a six-input AND gate by connecting the outputs of both three-input AND gates, using an additional two-input AND gate (Fig. 4c and Supplementary Fig. S30). When all six inputs were added the circuit produced high output signal (t1/2 ~12 mins.). Conversely, the circuit produced low output for any combination of five or fewer inputs. While the number of hairpins increased from four for the two-input AND gate, to eighteen for the six-input AND gate, all hairpins used the same conserved set of only five domains, plus a separate domain for each input and output. Similar to our experiments on wires, we observed attenuation of the output signal with increasing number of hairpins (Supplementary Fig. S23). This signal loss can in part be explained with a model assuming a fixed non-zero probability of hairpin omission (Supplementary Fig. S24).
To exemplify arbitrary Boolean logic computation using our domino architecture, we built a two-input dual-rail XNOR gate (Fig. 4d and Supplementary Fig. S31). Dual-rail encoding allows an arbitrary Boolean logic function to be realized using only a combination of AND and OR gates, avoiding the need for NOT gates, which are difficult to implement experimentally4. Each input variable A is specified as TRUE by a dedicated signal strand A1, and specified as FALSE by a distinct signal strand A0 (Fig. 4d). Only one of the two strands is present at any given time. Accordingly, our dual rail XNOR gate was implemented as a pair of parallel circuits, computing TRUE and FALSE outputs separately. The complete circuit arranged seventeen hairpins into six two-input AND and OR gates. The experimental data showed signal propagation (t1/2 < 8 mins.) consistent with predictions from the computational model.
Discussion
Overall, the number of unique sequences required for building any complex domino circuit did not depend on the number of constituent gates, but only on the number of inputs and outputs. Our domino circuits also performed significantly faster than previous systems with diffusible components (Supplementary Table S2). For example, a three-input AND domino gate had a half time of 7 min with operating concentration of only 2 nM, compared to 4 hours for an equivalent non-localized circuit with diffusible components operated at 100 nM concentration 4.
The modularity of our design approach enabled us to construct a family of cascaded circuits spanning an entire rectangular tile origami. Even larger and more complex circuit designs could be made with bigger and multilayered stiffer origami scaffolds, however three issues would need to be addressed. First, inputs are currently distinguished only by their toehold domains, and in practice the number of sufficiently orthogonal six nucleotide toeholds is small. This limitation can be overcome by incorporating an additional recognition domain in the Input hairpin, to encode input specificity (Supplementary Fig. S32). Second, we observed signal attenuation with increasing circuit size, likely due to imperfect hairpin incorporation. Such imperfections will become less limiting with continuing improvements in origami assembly protocols. Alternatively, component redundancy could be used to compensate for assembly defects, and signal restoration modules could overcome attenuation. Third, although inter-origami interactions are not currently limiting, they could become significant as circuit size increases. Such interactions could be minimized by using immobilizing techniques to tether origami molecules to a surface, or by using closed 3-dimensional origamis with components tethered to the inner faces.
Circuit architectures with localized components may provide a path towards the delivery of DNA circuits to cells, a long-held ambition of dynamic DNA nanotechnology that only recently started to become reality37–39. Not only do localized circuits enable control of stoichiometry during delivery, but increased speed compared to non-localized circuits may be even more pronounced in the densely packed cellular environment. Moreover, localized circuits could be used to increase the complexity of chemical synthesis reactions achievable with DNA-templated chemistry40,41 or to enhance the specificity of theranostic DNA robots42.
Author contributions
G.C., N. D., R.M., A.P. and G.S. designed experiments and wrote the paper. G.C. performed the experiments. N.D. and A.P. performed the modeling studies.
Acknowledgements
We thank K. Strauss and L. Ceze for their support in initiating this project, and F. Randisi for assistance with oxDNA simulations. This work was supported by NSF grants CCF-1409831, CCF-1317653 and HCC-1212940 and ONR grant N00014-13-1-0880 to G.S. G.C. was partially supported Microsoft Research Ltd.
References




