

ABSTRACT
The intestine is maintained by stem cells located at the base of crypts and distinguished by the expression of LGR5. Genetically engineered mouse models have provided a wealth of information about intestinal stem cells, while less is known about human intestinal stem cells due to difficulty detecting and isolating these cells. We established an organoid repository from patient-derived adenomas, adenocarcinomas, and normal colon, which we analyzed for variants in 71 colorectal cancer (CRC) associated genes. Normal and neoplastic colon tissue organoids were analyzed by immunohistochemistry and fluorescent-activated cell sorting for LGR5. LGR5-positive cells were isolated from 4 adenoma organoid lines and were subjected to RNA-sequencing. We found that LGR5 expression in the epithelium and stroma was associated with tumor stage, and by integrating functional experiments with LGR5-sorted cell RNA-seq data from adenoma and normal organoids, we found correlations between LGR5 and CRC- specific genes, including DKK4 (dickkopf WNT signaling pathway inhibitor 4) and SMOC2 (SPARC related modular calcium binding 2). Collectively, this work provides resources, methods and new markers to isolate and study stem cells in human tissue homeostasis and carcinogenesis.
INTRODUCTION
In adult mammals, the intestine is a site of rapid cellular turnover, mediated by a population of intestinal stem cells (ISCs) that reside at the base of intestinal crypts (Barker et al., 2008). These stem cells are identified by expression of several genetic markers in the mouse (van der Flier et al., 2009, Montgomery et al., 2011, Powell et al., 2012, Yan et al., 2012), and genetically modified mice provide a robust toolbox for isolating and manipulating these ISCs (Barker et al., 2007, Snippert et al., 2010, Sato et al., 2009). Lgr5 (leucine-rich repeat-containing g-protein coupled receptor 5) is one of the best characterized of these markers. The LGR family of proteins (LGR4, 5, 6) code for receptors of the secreted R-spondin proteins (Rspo1- 4). Together, Lgr/Rspo acts to potentiate Wnt pathway signaling (Carmon et al., 2012, de Lau et al., 2011). Lineage tracing experiments have demonstrated that differentiated cell lineages in mouse small intestine and colonic crypts are clonally derived from Lgr5(+) ISCs (Barker et al., 2007). Additionally, lineage tracing studies have revealed that Lgr5 marks a population of stem-like cells within precancerous adenoma tissue that drive adenoma growth (Schepers et al., 2012), and human colorectal cancers overexpress LGR5 (Junttila et al., 2015). Previous efforts to expand, isolate, and experimentally characterize primary human LGR5 cells have been hampered by two distinct issues: (1) The difficulty in obtaining cultures highly enriched for epithelial stem cells (Wang et al., 2015b), and (2) a paucity of specific reagents to detect and isolate live LGR5(+) human cells (Barker, 2014). Recent efforts have successfully used gene editing techniques to create human organoid LGR5 reporter lines (Shimokawa et al., 2017), however, this approach does not allow isolation from primary (unmodified) tissue and is not broadly useful across many cell lines. Previous studies have also reported varied localization of Lgr5 within the normal crypt using antibody-based methods (Becker et al., 2008, Kleist et al., 2011, Fan et al., 2010, Takahashi et al., 2011, Kobayashi et al., 2012, Kemper et al., 2012). Efforts have also utilized in situ RNA hybridization strategies to detect LGR5+ cells in human tissue, however, this approach does not permit isolation of cells (Jang et al., 2013, Baker et al., 2015). Collectively, these studies highlight the need for approaches that can be widely adopted to identify and isolate human intestinal stem cells.
Intestinal organoid culture is a robust system to grow tissue-derived cells that maintains the cellular heterogeneity in the intestinal epithelium (Sato et al., 2011a, Dedhia et al., 2016). Here, we establish an organoid biobank of normal, precancerous adenoma and colon adenocarcinoma from human tissue. We report detailed and robust methods to identify and isolate human colon LGR5(+) cells from normal human intestine and primary adenomas growing in long-term organoid culture using commercially available antibodies. Using these methods, we quantify Lgr5 protein expression in human intestinal tissues, including a colon adenocarcinoma tissue microarray (TMA). Using magnetic bead and fluorescent activated cell sorting (FACS) to enrich and isolate LGR5(+) and LGR5(−) cells from organoids, we conducted RNA sequencing and defined the expression profile of human LGR5(+) adenoma cells. We generated a TCF/LEF-GFP reporter adenoma organoid line to test the relationship between WNT reporter activity and Lgr5 expression. Through our analysis, we find that LGR5 protein expression is associated with progressive stages of cancer and we demonstrate that isolated human LGR5+ cells are enriched for TCF/LEF-GFP expression, SMOC2, and DKK4, a gene not detectable in normal colon tissue but associated with colorectal cancer. The methods and datasets presented are powerful resources for basic biological studies of the role of LGR5(+) cells in human colon homeostasis, as well as translational studies in chemoprevention and precision medicine designed to target LGR5(+) cell populations.
RESULTS
Isolation, Culture and genomic characterization of human adenoma organoids
We have developed an ongoing repository (Table 1) of organoids from patient-derived adenomas (n=17; including 2 high-risk sessile serrated adenomas), adenocarcinomas (n=4; including 1 colitis- associated cancer), and normal colon (n=9). All have been cryopreserved at early passage, reestablished from frozen stock, validated as matching their source tissue by short tandem repeat profiling and genomic sequencing (see below sequencing methods), are mycoplasma-free, and have demonstrated long-term culture (i.e., > 6 months in continuous culture). The neoplasm-derived organoids have been genomically characterized for variants in a panel of common CRC driver mutations across 71 genes (Table 1).
A targeted colorectal cancer DNA sequencing panel was used to determine the presence of variants for 71 different oncogenes and tumor suppressor genes often mutated in colorectal cancers. Stop codon (*); frame shift (FS).
LGR5 immunohistochemical specificity in human colon and intestine
Two antibody clones for human Lgr5 were used in this study: a rabbit monoclonal antibody generated against a peptide sequence of LGR5 (clone STE-1-89-11.5) and a rat monoclonal antibody generated against a full-length LGR5 protein (clone 22H2.8). At the onset of this study these antibodies were in development by Miltenyi Biotec, but now both antibodies are commercially available (Miltenyi Biotec GmbH, Bergisch Gladbach, Germany; see Methods and Supplemental Methods). Details of their development have been presented previously (Agorku et al., 2014, Agorku et al., 2013), including the use of human LGR4 and LGR6 stable transfectants to demonstrate lack of cross-reactivity with these close homologues.
LGR5 immunohistochemical (IHC) expression was localized with clone STE-1-89-11.5 to the crypt base columnar (CBC) cells in normal formalin fixed paraffin embedded (FFPE) colon tissue (Fig. 1A1). At high magnification this staining pattern marked thin cells (Fig. 1A2), consistent with the morphology of CBC cells. From the same patient, an adenoma (found in the adjacent margins of an adenocarcinoma tissue resection, 10cm from the histologically normal tissue) showed intensified staining at the dysplastic crypt bases (Fig. 1A3) and sporadic focal staining throughout the more disorganized epithelial component. Interestingly, stromal staining was pronounced in this cancer-associated adenoma (Fig. 1A3). Supportive LGR5 in situ hybridization (ISH) staining was observed in the normal CBC cells (Fig. 1B top panel); in the dysplastic epithelium (Fig. 1B, arrow-1) and in the associated stroma (Fig. 1B, arrow-2).
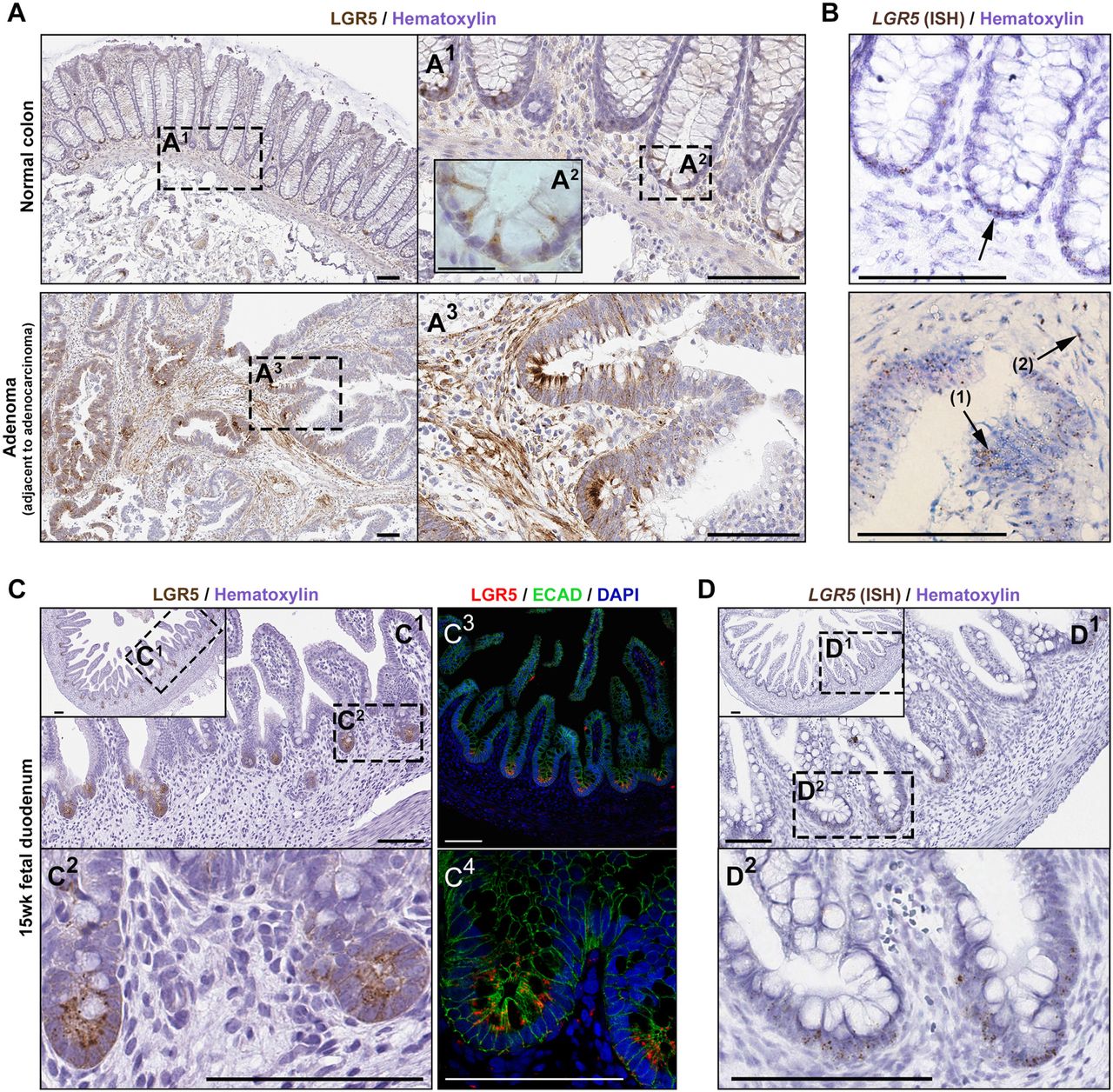
(A) LGR5 IHC staining in normal human colon (one of five representative patients) at low (A1) and high (A2) magnification, as well as adenoma (A3) from the same patient (high-grade dysplasia; adjacent to adenocarcinoma; specimen 14881). (B) Lgr5 expression by in situ hybridization provides a conventional reference for the LGR5 IHC staining in normal crypts (upper panel) and in the adenoma (bottom panel); glandular Lgr5 (arrow-1) and stromal expression (arrow-2) in adenoma; (C) LGR5 IHC (C1, C2) and IF staining (C3, C4) in fetal duodenum, and (D) ISH expression in the same duodenum specimen. Scale bars, 100μM; A2, 25μM.
The human fetal small intestine has been shown to express high levels of LGR5 mRNA relative to adult by RNA-sequencing (Finkbeiner et al., 2015). Consistent with this, robust and specific Lgr5 protein staining by IHC and immunofluorescence (Fig. 1C), in conjunction with LGR5 ISH (Fig. 1D), was observed in the proliferative zone of the 15-week fetal gut. In contrast, IHC and IF staining in adult duodenum (Fig. S1A) showed weak punctate LGR5(+) staining in cells present between Paneth cells marked by DefensinA5 (DEFA5), consistent with published ISH and RNA-sequencing data (Finkbeiner et al., 2015).
Clone STE-1-89-11.5 was further demonstrated to be specific for human LGR5 by Western blotting. Mouse 1881 lymphoma cells that were previously transfected with human LGR5 served as a positive control [1881(+); provided by Miltenyi Biotec]. Transfection stability was confirmed by mRNA expression analysis (Fig. S1B). The antibody showed strong reactivity against the human LGR5 1881(+) cell line, as well as measurable activity against one adenoma organoid (specimen #14881) (Fig. S1C).
LGR5 immunohistochemical expression levels, and localization, are associated with human colon cancer stage
LGR5 IHC staining was performed in a FFPE TMA, which included 2 normal tissues, 3 low grade small adenomas (well differentiated), and 70 adenocarcinomas. We also included staining of 5 additional normal colon autopsy samples obtained for our studies in this analysis (Fig. 2). Tissue sections were scored for staining intensity in the glandular epithelium and separately in the stroma. Unlike normal colon, which showed staining in CBCs at the base of the crypt, the adenomas stained intensely in distinct zones of epithelium or for the entire dysplastic crypt (Fig. 2A). Relative to normal tissue, the adenocarcinomas showed increased LGR5 staining in both the epithelium and in the stroma (Figure 2A-C).

(A) Representative images of LGR5 staining in the epithelial and stromal components in normal colon (n=5 normal autopsy samples) and from a CRC tissue microarray (n=2 normals, 65-68 neoplasm). Scale bar, 50μM. (B) Association between epithelial or (C) stromal staining intensity, and stage/grade of the tissue tested by generalized linear modeling, with staining intensity as the dependent variable and stage or grade as the independent variable.
Isolating Lgr5(+) cells from patient-derived adenoma organoids using magnetically-activated cell sorting (MACS) and FACS
Fig. 3A outlines the approach developed for isolation and analysis of LGR5(+) cells from human adenoma. Adenoma organoids were established in culture (approx. 2 months) and genomically characterized (Table 1). The organoid cultures used in this study (14881, 282, 584, 590) were established and cultured in ‘reduced medium’, KGM-Gold™ (Lonza, Walkersville, MD; KGMG), a serum-free epithelial medium containing epidermal growth factor (EGF) and pituitary extract (Dame et al., 2014). We chose this medium to establish/expand non-normal epithelium, similar to approaches used in recent studies which have demonstrated that mutations allow cells to grow independent of particular ISC niche factors (Li et al., 2014, Matano et al., 2015, Drost et al., 2015). When organoid cultures grown in reduced medium were transferred to medium replete with ISC niche factors (L-WRN medium-see methods) (Miyoshi and Stappenbeck, 2013), they generally transitioned from budding structures (Fig. 3B, middle column) to thin-walled spherical cysts (Fig. 3B, right column). Accompanying this change in morphology was an increase in Lgr5 mRNA expression in L-WRN-cultured organoids compared to organoids cultured in reduced medium, as assessed by qRT-PCR (Fig. 7A, left graph). Flow cytometric analysis revealed that L-WRN-cultured organoids also had an increase in the number of Lgr5 positive cells (Fig. 3C, comparing “stain” in the first and second row; Fig. S2A). Even with L-WRN-enhanced LGR5 expression, a small proportion of cells were LGR5(+) based on flow cytometry, and therefore magnetic bead positive enrichment was used prior to FACS to increase the number of cells obtained.

(A) Graphical representation of complete procedure. (B) Culture of adenoma organoids for LGR5 enrichment. LGR5 IHC staining (left column) in biopsied large adenoma (>10mm; two representative specimens are shown). Organoids with budding morphology (middle column) derived from these specimens cultured in the reduced medium KGMG. The same cultures with cystic morphology (right column) after being transferred for 3-4 weeks to L-WRN medium. Scale bars, 100μM. (C) Representative scatterplots of LGR5 expression in organoids cultured in KGMG or L-WRN (specimen 282). Live, LGR5(+) human adenoma cells were obtained after LGR5-magnetic bead enrichment by isolating DAPI(−) and LGR5(+) cellular populations.
Mouse 1881 cells stably expressing Lgr5 were used to validate the magnetic bead positive separation protocol and the high specificity of antibody clone 22H2.8. 1881-LGR5 expressing and nonexpressing cells were mixed in varying proportions, magnetically separated, and analyzed by flow cytometry (Fig. S2B). The initial ratio of 1881 LGR5(+) to 1881 LGR5(−) cells was predicted as shown by the pre-magnet flow histograms (Fig. S2B, left column). The post-magnetic separation shows almost complete separation of 1881 LGR5(+) cells in the positive fraction (Fig. S2B, right column) from 1881 LGR5(−) cells in the negative fraction (middle column). After magnetic bead based enrichment of the adenoma organoid cells, the unbound flow-through negative population was used to set the Lgr5 FACS gate (Fig. 3C bottom row). Magnetic bead-bound LGR5(+) cells from organoids were isolated via FACS, and importantly, we observed that while only a small percentage of cells bound to beads were LGR5(+) (Fig. 3C bottom row), on average, magnetic bead pulldowns of LGR5(+) cells led to an 11.3-fold enrichment in the number of LGR5(+) cells (cultured in L-WRN medium) obtained compared to samples without magnetic enrichment (specimens 282, 584, 590; SE=6.55; n=3 sorts no magnet, n=7 sorts with magnet; Table S1).
Transcriptomic profiling of LGR5(+) adenoma cells
The gene expression signature of human LGR5(+) intestinal cells is essentially uncharacterized, which has significantly limited our understanding of the role of these cells in human intestinal stem cell biology and colorectal cancer progression. Using the methods described above, we isolated LGR5(+) cells from four different adenoma-derived organoid lines grown in L-WRN media and conducted RNA-seq on three sorted cellular populations from each (Fig. 4, Fig. S3): (1) magnetic column flow-through cells, termed LGR5(FT-); (2) cells that bound to the magnet, but were Lgr5 negative based on FACS, termed LGR5(−); and (3) cells that bound to the magnet and were LGR5 positive based on FACS, termed LGR5(+). Genomic variant analysis showed that each of these patient specimens has functionally significant mutations within commonly mutated colon cancer genes, confirming the adenoma, and not normal, identity of these organoids (Table 1). Expression profiles of the LGR5(−) and LGR5(FT-) were found to be largely similar at both the gene expression and pathway level (Fig. S3C, D). Due to transcriptional similarity between negative populations, our analysis focused on comparisons between the magnetic bead-bound cells deemed to be LGR5(+) and LGR5(−) by FACS. Multidimensional scaling on the top 500 most variably expressed genes revealed that samples clustered distinctly by the patient of origin, not by the expression of Lgr5 (Fig. 4A, Fig. S3A). Despite this, we identified 519 differentially expressed genes (FDR P-value < .05) between the two cell populations across four genetically diverse adenomas (Fig. 4B and Table S2). Expression values for all differentially expressed genes for each specimen, as well comparisons between LGR5(+) cells and LGR5(FT-) cells, are reported in Table S2. We determined that LGR5 had the highest level of statistical enrichment in the LGR5(+) cells (FDR=3.8E- 21) and was expressed an average of 5.5-fold higher compared to LGR5(−) cells, lending confidence to the specificity of the LGR5 antibody and the separation procedure. Unsupervised hierarchical clustering analysis using the top 50 most differentially expressed genes showed clear separation between the LGR5(+) and LGR5(−) samples from three of the four enteroid lines (Fig. 4C), suggesting both commonalities and heterogeneity in the LGR5(+) cell gene expression signature between specimens. When the expression signature of LGR5(+) cells were clustered with those from both the LGR5(−) and LGR5(FT-) cells, however, there was clear separation of the LGR5(+) cells across all four organoid lines (Fig. S3B). Stem cell markers associated with the colon as well as other tissue-specific stem cells including BMI1, MEX3A, and SMOC2, were upregulated in LGR5(+) cells, while known markers of colonic differentiation, including MUC2, TFF3, and KRT20, were down-regulated (Fig. 4D). KEGG pathway analyses (Kyoto Encyclopedia of Genes and Genomes) (Kanehisa and Goto, 2000) revealed differential expression in LGR5(+) cells for genes involved in metabolism, including the pathways, glycolysis and gluconeogenesis, biosynthesis of amino acids, metabolic pathways, carbon metabolism, fructose/mannose metabolism, glycosphingolipid biosynthesis, and central carbon metabolism in cancer (Fig. 4E; Fig. S3C). Additionally, LGR5(+) cells had significantly differentially expressed genes involved in HIF-1 signaling, cytokine-cytokine receptor interaction, hematopoietic cell lineage, MAPK signaling, PI3K/AKT signaling, and pathways in cancer (Table S2; Fig. 4E, Fig. S3C, Fig. S4-9).

(A) Multidimensional scaling plot of the LGR5(+) and LGR5(−) cells, based on the top 500 most variable genes. Patient identifiers #14881, 228, 584, and 590. (B) False discovery rate (FDR) volcano plot of the log(2) ratio of gene expression between the LGR5(+) and LGR5(−) cells. (C) Unsupervised hierarchical clustering heatmap of the 50 most differentially expressed genes by fold change between the LGR5(+) (red) and LGR5(−) (green) populations. (D) Log(2) fold change in gene expression between LGR5(+) and LGR5(−) cells for known markers of colon stem (red) and differentiated (green) cells. (E) The top 10 most enriched KEGG pathways for differentially expressed genes between the LGR5(+) and Lgr5(−) cells.
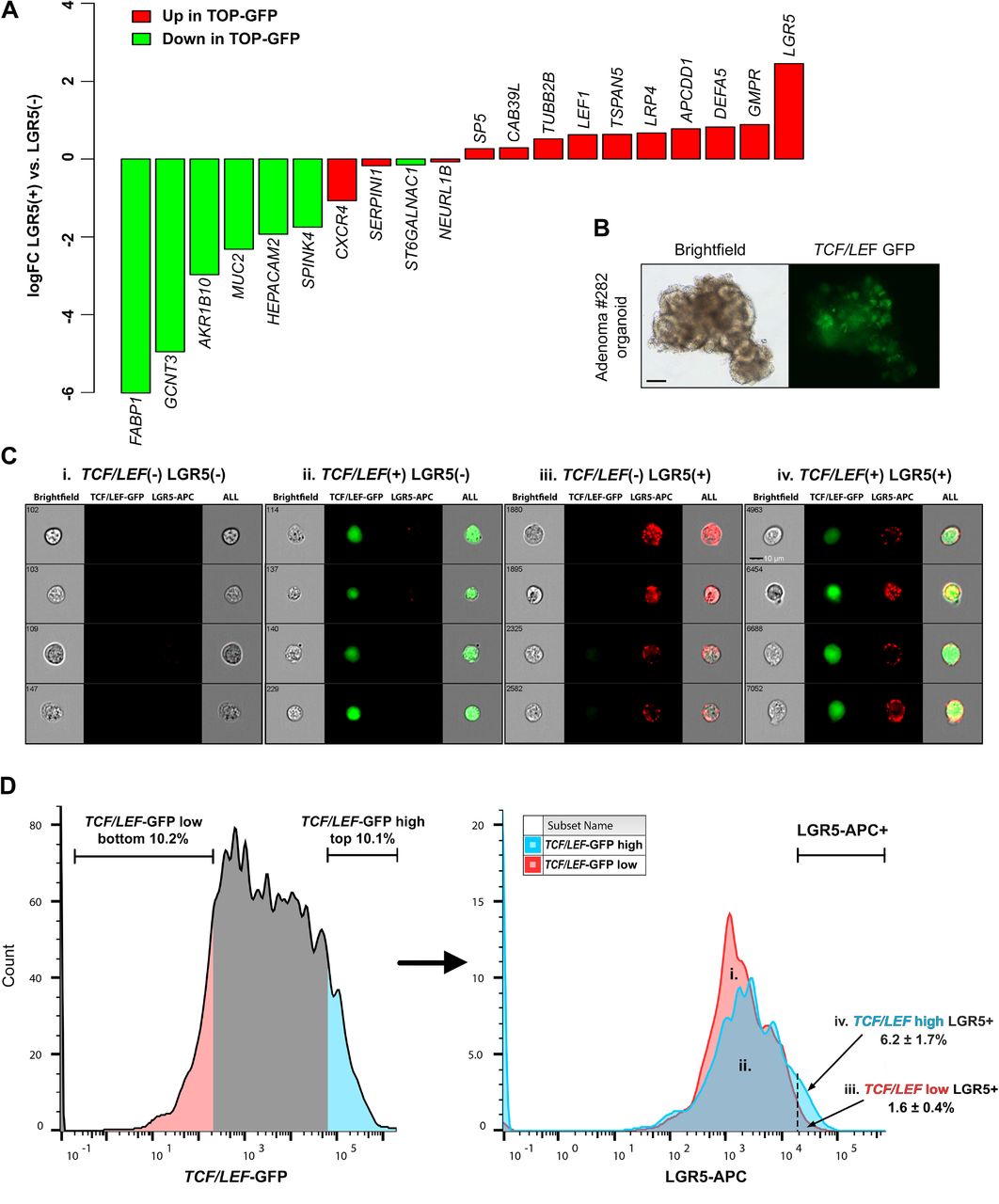
We also identified a high level of concordance between LGR5(+) cells and a previously reported set of genes associated with high levels of Wnt signaling in mouse cancer stem cells (based on TOP-GFP Wnt reporter activity (Vermeulen et al., 2010) (Fig. 5A). To functionally validate this overlap, we transduced organoids (line 282) with a lentiviral TCF/LEF-GFP reporter (Figure 5B). TCF/LEF-GFP organoids were stained with the Lgr5 antibody for analysis by ImageStream flow cytometry. ImageStream analysis visually established the specificity of the fluorescently tagged Lgr5 antibody and TCF/LEF-GFP expression to viable cells (Fig. 5C). Based on the same experimental paradigm established by Vermeulen et al., we compared LGR5 expression in the top 10% highest and lowest GFP expressing cells. We identified a significant enrichment of LGR5(+) cells in the TCF/LEF-GFPHI cells relative to low cells. (average 3.8 fold enrichment of Lgr5; 6.2% vs. 1.6%).

(A) Concordance of adenoma LGR5(+) cell gene expression with gene expression associated with Wnt signaling in colon cancer stem cells, based on TOP-GFP Wnt reporter activity (Vermeulen et al., 2010). (B) Brightfield and fluorescence images of the 282 adenoma organoid line lentivirally transduced with a TCF/LEF-GFP reporter for Wnt signaling. Scale bar, 50μM. (C) Representative ImageStream flow cytometry results of TCF/LEF-GFP organoids stained with the LGR5- APC antibody (n=3 experimental replicates). Scale bar, 10μM. (D) Comparison of LGR5 expression in the 10% highest and 10% lowest TCF/LEF-GFP expressing cells; values in iii and iv represent mean % ± s.e.m.
To identify a “human adenoma LGR5(+) cell gene signature” with more stringent parameters, we compared genes upregulated in LGR5(+) cells to both LGR5(−) and to LGR5(FT-) cells to which we identified 33 genes (Table 2). A comparison between these 33 genes in the “human adenoma LGR5(+) cell gene signature” and the previously published “murine intestinal stem cell signature,” generated from isolated Lgr5HI ISCs (Munoz et al., 2012), identified 4 overlapping genes that were enriched in both populations, including the canonical ISC markers LGR5, SMOC2, and CDCA7 (Fig. 6A). This analysis also revealed a large number of genes in the human adenoma LGR5(+) cell signature that did not overlap with the mouse Lgr5 stem cell signature.

(A) Gene expression overlap between genes upregulated in LGR5(+) vs. LGR5(−) cells and the Lgr5 mouse intestinal stem cell signature reported in Munoz et al (Munoz et al., 2012). (B) Comparison of gene expression between LGR5(+) adenoma cells and previously published RNA-seq data from normal human colon (Uhlen et al., 2015) for genes in the “human adenoma LGR5+ cell gene signature. (C) The top ten most differentially expressed genes by magnitude between LGR5(+) adenoma cells and normal human colon (all genes; unbiased analysis) (Uhlen et al., 2015). (D) An analysis of TCGA colorectal cancer gene expression data through Oncomine™ of DKK4, comparing colorectal adenocarcinoma expression with normal colon and rectum.
Given that our transcriptional analysis compared cells, LGR5(+) vs. LGR5(−), isolated from adenoma-derived organoid cultures (as opposed to intact tissue biopsies), we reasoned that it was possible that LGR5(+) enriched genes might be masked by comparing fractions which, in whole, were cultured in medium that promotes high levels of WNT signaling (L-WRN). Therefore, we also compared LGR5(+) cells with previously published RNA-seq data from whole-thickness normal human colon (Uhlen et al., 2015) (Fig. 6). When we compared the expression of normal colon genes with the 33 genes from the “human adenoma LGR5(+) cell gene signature” (Table 2), we observed a strong enrichment for a majority of the 33 genes (Fig. 6B), including the Lgr5 surrogate marker gene SMOC2. In an unbiased comparison of all LGR5(+) expressed genes, one of the most upregulated in LGR5(+) adenoma cells compared to normal colon was the Wnt pathway inhibitor DKK4 (dickkopf WNT signaling pathway inhibitor 4) (Fig. 6C). DKK4 was virtually undetectable in the normal colon (Table S2). An analysis of TCGA colorectal cancer gene expression data (The Cancer Genome Atlas, 2012) revealed that DKK4 is significantly upregulated in colorectal tumors compared to normal tissue (Fig. 6D). To follow up on these correlations between LGR5, SMOC2 and DKK4 identified via RNA-seq analysis, we performed additional functional and expression profiling experiments. Three adenoma organoid lines were grown in reduced medium or in L-WRN media to drive a more “stem-like” phenotype in the cells, and organoids from both conditions were analyzed for LGR5, SMOC2 and DKK4 by qRT-PCR. We observed a significant increase in the expression of LGR5 (4.6-fold), SMOC2 (78-fold), and DKK4 (6992-fold) across the three adenomas (Fig. 7A). We further validated the protein expression of LGR5 (Fig. 7B; Fig. S10) as well as the co-localization in adjacent histological sections of LGR5 and SMOC2 mRNA expression in colon organoids (Fig. 7C) and the co-localization of LGR5 and DKK4 in colon adenocarcinoma tissue (Fig. 7D) by in situ hybridization.
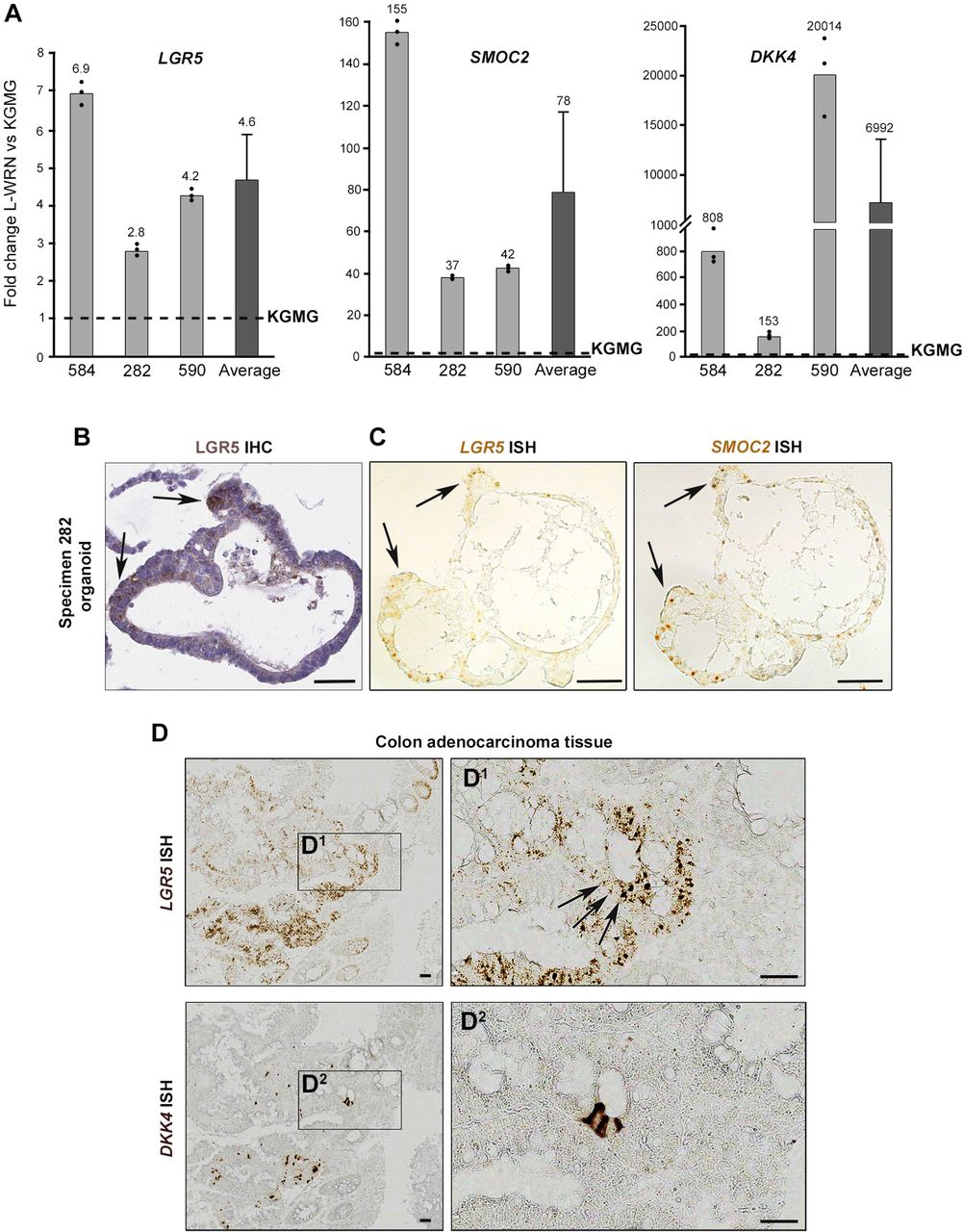
(A) Relative LGR5, SMOC2, and DKK4 mRNA expression in L-WRN vs KGMG adenoma organoid cultures. Individual specimen qRT-PCR technical replicates; represents the mean±s.e.m. of n=3 biological replicates. (B) LGR5 immunohistochemistry staining and (C) SMOC2 and Lgr5 mRNA in situ hybridization (ISH) of formalin fixed paraffin-embedded (FFPE) adenoma 282 organoids cultured in L- WRN. Arrows designate localization of Lgr5 and SMOC2 in corresponding regions of consecutive serial sections. Scale bar, 50μM. (D) DKK4 and Lgr5 ISH images in consecutive FFPE cuts of a colon adenocarcinoma (specimen 815). Scale bar, 50μM.
Because we isolated enriched LGR5(+) cells from L-WRN-cultured adenoma organoids, we also wanted to isolate LGR5(+) cells directly from normal colon tissue to demonstrate robustness of the isolation and purification methods. Using the same dissociation methods as were employed for the adenoma organoid cultures, dissociated single cells from 3 independent normal cadaveric colon specimens were sorted for EpCAM (epithelial cell adhesion molecule) positivity and Lgr5 expression. LGR5(+) cells comprised 2.35%, 0.63%, and 0.76% of the EpCAM(+) cells in independent experiments. A representative distribution of LGR5(+) staining from a normal colon, demonstrates that fluorescent intensity for LGR5(+) events were 2-3 orders of magnitude above baseline (Fig. 8A). LGR5(+) cells were collected and subsequently analyzed by qRT-PCR for LGR5 and the intestinal stem cell marker OLFM4 (olfactomedin 4), which was recently shown to be expressed in normal human colon, and in adenoma (Jang et al., 2016). We identified an average of 7.5-fold increase in LGR5 expression and 2.3-fold increase in OLFM4 mRNA expression in the sorted LGR5(+) normal colon cells relative to the LGR5(−) cells (Fig. 8B). Further, protein expression of OLFM4 was confirmed by IF, which showed staining at the bottom 1/3rd of the normal colon crypts (Fig. 8C).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
(A) Flow cytometry gating strategy for LGR5(+) cells isolated directly from warm autopsy normal colon tissue (specimen 84, a representative scatter plots of 3 patients; Table 1). Includes EpCAM(+) inclusion criteria, using a phycoerythrin (PE)-conjugated antibody (not shown). To set the LGR5-APC(+) gating, either magnet flow-through (specimen 81 & 83), as done with organoid sorts, or rigorous so-called fluorescence- minus-one controls [FMO; DAPI(−), EpCAM(+); specimen 84] were used. (B) mRNA expression by qRT-PCR of Lgr5 and OLFM4 across all 3 patient samples. Values represent mean ± SE. (C) OLFM4 immunofluorescent staining of normal colon crypts. Scale bar, 100μM. (D) Normal colon organoid cultures were established from warm autopsy resections. LGR5(+)/(−) cells were isolated from early passage organoids, sorted into culture medium and seeded into Matrigel to access organoid-forming efficiency. Mean organoid-forming efficiency from two normal colon organoid cultures (specimen 78, passage 5; and specimen 85, passage 3) sorted for Lgr5(−) vs. LGR5(+) cells (n=4 wells per fraction), error bars represent s.e.m. Representative images (right panels; specimen 85) of organoids formed from Lgr5(−) and LGR5(+) cells shown at day 14 and day 52 (passage 5). Scale bars, 200μM top panel; 1mm bottom panel.
Isolated LGR5(+) and LGR5(−) cells were sorted into medium and seeded into Matrigel to access organoid-forming efficiency (Fig. 8D). Those cells isolated directly from patient tissue did not initiate organoids under single cell culture conditions (n=3; data not shown). Instead, we established normal organoid cultures from the colon cadaveric resections (Table 1), and isolated LGR5(+) and LGR5(−) cells from early passage organoids. Organoids formed from sorted LGR5(+) and LGR5(−) cells and were passaged up to five times before terminating the experiment. Both LGR5(+) and LGR5(−) cells could form organoids with equal ability. The nearly equivalent capacity to form organoids between Lgr5(+) and LGR5(−) cells suggest that LGR5(−) cells, especially when cultured in a medium which drives WNT signaling (L-WRN), are capable of much plasticity, as other recent studies have recently elaborated (Richmond et al., 2015, Beyaz et al., 2016, Tetteh et al., 2016, Sasaki et al., 2016, Shimokawa et al., 2017, Yan et al., 2017, Jadhav et al., 2017).
DISCUSSION
LGR5 is one of the few cell surface markers available for the identification and isolation of cycling stem cells in the colonic crypt. Efforts to generate effective antibodies to isolate these cells have largely been unsuccessful (Barker, 2014). Recently antibody drug conjugates (ADCs) have been developed to target cells expressing LGR5, demonstrating therapeutic potential (Gong et al., 2016, Junttila et al., 2015), but as part of these applications primary human cell isolation and characterization was limited (Hirsch and Ried, 2016). Thus, the role of Lgr5(+) stem cells in human intestinal development, homeostasis, and carcinogenesis remains poorly understood. The identification and isolation of human LGR5(+) adenoma cells, as well as the establishment of methods for the long-term culture and propagation of organoids enriched for these rare cells, as described in this study, provides some of the first insight into the characteristics of these cells in humans. Further, we confirm that our methods are robust for the identification and isolation of LGR5(+) cells from normal human colon tissues. Ideally, methods and data sets presented in this work will be widely utilized to overcome a major hurdle in the field of colorectal cancer stem cell biology.
Human primary intestinal organoid culture systems represent an unparalleled tool to study colorectal cancer biology (Matano et al., 2015) and to develop precision medicine platforms for targeted therapeutics (van de Wetering et al., 2015). Here, we report a living biobank of human normal colon, adenoma, and adenocarcinoma tissues. We used this biobank to study and isolate(+) cells from four genetically diverse adenoma-derived organoids. We found that LGR5 is expressed at higher levels in organoids in response to Wnt3a, Rspondin-3, and Noggin, and that this is associated with a cystic phenotype, consistent with other reports (Sato et al., 2011b, Farin et al., 2012, Matano et al., 2015, Drost et al., 2015, Onuma et al., 2013). While this report was in preparation, a study eloquently demonstrated that Mex3a marks a subset of LGR5(+) slow cycling intestinal cells (Barriga et al., 2017). Indeed, we show that MEX3A is among the upregulated genes in isolated human colon LGR5(+) cells. Interestingly, while gene expression associated with LGR5 expression in human adenoma significantly overlapped with Lgr5HI stem cells in the mouse (Munoz et al., 2012), there were several examples where human expression patterns were opposite to that of the mouse. For example, SEMA3B and PLCE1 are enriched in mouse Lgr5HI cells, but were downregulated in human LGR5(+) adenoma cells (FDR=0.04 and 0.002 respectively). Determining whether differential expression of these genes reflects differences in human compared to mouse biology or adenoma compared to normal human colon biology represents an important future direction for this research.
Histological approaches that compared LGR5(+) cells from adenoma and adenocarcinoma samples with normal colon revealed a number of unexpected biological findings. While LGR5 expression significantly correlated with advanced stage in colorectal cancers, we surprisingly identified that high grade, poorly differentiated, tumors had lower LGR5 expression compared to lower grade tumors. This result corroborates a recent report of LGR5 mRNA expression, quantified by in situ hybridization, in human colorectal cancers which also reported a decrease in LGR5 expression in poorly differentiated tumors (Martin et al., 2017). We also identified the presence of stromal LGR5 staining, and confirmed the specificity of this signal with Lgr5 mRNA ISH. We did not however identify obvious stromal staining in normal tissue. We identified that stromal staining of LGR5 was associated with cancer stage based on a cohort of independent samples on a tissue microarray. Others have identified the presence of multipotent LGR5(+) stromal cells in the oral mucosa and tongue of adult mice expressing the LGR5-EGFP reporter with a phenotype resembling neural crest cells (Boddupally et al., 2016). Further, Lee et al eloquently demonstrated recently through lineage tracing experiments in mice that LGR5(+) cells located in the lung alveolar mesenchyme support differentiation of the alveolar epithelium (Lee et al., 2017). Our finding is likely of high relevance in light of the identification of stromal gene expression having a strong correlation with colorectal cancer subtypes (Calon et al., 2015). Future work should focus on understanding the role of these putative LGR5 expressing stromal cells in colorectal carcinogenesis.
The ability to isolate LGR5(+) and LGR5(−) cells from human samples will enable future studies aimed at understanding human stem cells and cancer. In our functional studies, we found that single FACS-isolated LGR5(+) cells from normal colon organoids did not have increased organoid forming efficiency compared to LGR5(−) cells. These results are in line, however, with a recent report of the organoid forming efficiency of human LGR5(+) cells labeled genetically using Cas9/CRISPR, where the authors demonstrated a reemergence of LGR5-GFP(+) cells from the sorted Lgr5(−) fraction, and noted that organoid forming efficiency was not increased in the LGR5(+) cells when compared with LGR5(−) cells (Shimokawa et al., 2017), further highlighting the inherent plasticity of intestinal cells (Richmond et al., 2015, Beyaz et al., 2016, Tetteh et al., 2016, Sasaki et al., 2016, Yan et al., 2017, Jadhav et al., 2017). We anticipate that these varied approaches will help to move our understanding of human biology forward.
Transcriptional comparisons of LGR5(+) adenoma stem cells with published normal colonic RNA-seq data (Uhlen et al., 2015), identified genes strongly upregulated in LGR5(+) adenoma cells that were virtually unexpressed in the normal human colon, the highest being DKK4. DKK4 is a negative regulator of Wnt signaling that has been reported as both upregulated (Matsui et al., 2009, Pendas-Franco et al., 2008) and downregulated in CRC (Baehs et al., 2009). A recent study showed that DKK4 acts selectively to inhibit a subset of WNT ligands, but is proteolytically inactivated (Sima et al., 2016). Elevated DKK4 expression is linked with CRC chemoresistance (Ebert et al., 2012) and metastasis (Chen et al., 2015), both processes associated with cancer stem cells. We identified that DKK4 is significantly upregulated in colorectal cancers, compared to normal colon, in data from TCGA. The association with LGR5 expression suggests that DKK4 could be a novel biomarker of colon adenoma stem cells or cells with high Wnt activity, and may prove useful for colon cancer prevention or treatment efforts.
Notably, our experiments used a two-step enrichment methodology to obtain purified cells (magnet followed by FACS), yet we still encountered a relatively small population of LGR5(+) cells. This could be explained, in part, by work showing rapid internalization of LGR5 from the plasma membrane to the trans-golgi network (Snyder et al., 2017). Thus, it is possible that our antibody-based methods will only recognize rare cells expressing the highest levels of LGR5 protein on the cell surface. On the one hand, this may preclude us from isolating cells that express LGR5 mRNA but that do not have abundant protein localized to the cell surface. On the other hand, this same caveat would enhance specificity for the sorted population of cells, as supported by our gene expression profiling.
Our study presents a blueprint for the identification, in vitro culture, isolation, and experimental characterization of human LGR5(+) cells. We anticipate our methods can be readily adapted for the isolation and characterization of the LGR5(+) cell populations in other human tissues including cochlea (McLean et al., 2017), hair (Jaks et al., 2008), kidney (Barker et al., 2012), liver (Huch et al., 2013), lung (Lee et al., 2017), mammary gland (De Visser et al., 2012), oral mucosa (Boddupally et al., 2016), prostate (Wang et al., 2015a), and stomach (Barker et al., 2010, Simon et al., 2012). Additionally, we expect that the organoid culture methods described here will provide an experimental platform for the development of novel chemopreventive and chemotherapeutic agents that target LGR5(+) stem cells.
MATERIALS AND METHODS
Establishment of organoid cultures from human colonic adenomas
Isolation of human colonic crypts and adenomas, and culture and maintenance of adenoma organoid cultures, were performed using our previously described protocol (Dame et al., 2014). Adenoma tissue was acquired by endoscopy and adenocarcinoma tissue was collected from colonic resections according to protocols approved by the University of Michigan Institutional Review Board (IRB; HUM00064405/0038437/00030020), with informed consent obtained from all subjects. Normal colonic tissues were collected from deceased donors through the Gift of Life, Michigan (HUM00105750). Deidentified human fetal intestinal tissue was obtained from the University of Washington Laboratory of Developmental Biology and approved by University of Michigan IRB (HUM00093465). Growth media used for organoid cultures included KGM-Gold™ medium (Lonza) (Dame et al., 2014) and L-WRN medium. KGMG is a serum-free epithelial medium containing a reduced calcium concentration of 0.15mM, hydrocortisone, epinephrine and pituitary extract, as well as some components common to L- WRN media, epidermal growth factor (although 1:1000 of L-WRN; 0.1ng/mL), insulin, and transferrin. L-WRN conditioned medium (Miyoshi and Stappenbeck, 2013) contains high levels of Wnt3a, R- spondin-3 and Noggin, with added 10mM Nicotinamide (Sigma-Aldrich). Adenoma organoid cultures were propagated long-term in KGMG. To drive organoids from a budding to cyst morphology, and to enrich for a “stemness” phenotype, cultures were switched from the reduced medium, KGMG, to the stem cell support medium, L-WRN, for 3-4 weeks. Consistent with previous reports, organoids formed cystic structures in the presence of Wnt ligand provided by L-WRN medium (Sato et al., 2011b, Farin et al., 2012, Matano et al., 2015, Drost et al., 2015, Onuma et al., 2013). Cultures were propagated in Matrigel (Corning) which was made to 8mg/mL in growth media, in 6-well tissue culture plates. Cultures were passaged every 4-7 days by digesting Matrigel in cold 2mM EDTA and plated on the first day with 10μM Y27632 (Miltenyi Biotec), a Rho-associated protein kinase (ROCK) inhibitor. For further organoid line details, including media components and concentrations, subculturing methods, genomic characterization, cell line authentication and mycoplasma monitoring, see supplementary Materials and Methods.
Single Cell Isolation and Magnetic separation for LGR5(+) and LGR5(−) cells
Single cell suspensions of adenoma organoids or normal intact colon tissue were generated using the Tumor Dissociation Kit (human; Miltenyi Biotec) in combination with the gentleMACS Dissociator (Miltenyi Biotec) with protocol modifications. On the day of sorting, organoid cultures were treated with 10μM Y27632 for 2.5 hrs prior to harvest. The enzymes designated “H” and “R” were prepared in HBSS modified to 0.13mM calcium and 0.9mM magnesium (10% standard HBSS concentrations) to minimize differentiation of the epithelial cells while supporting enzymatic activity. All plasticware, including cell strainers and columns, were 0.1% BSA-coated, and buffers contained 5-10μM Y27632. Cells were labeled with anti-LGR5 MicroBeads (human; Miltenyi Biotec) and the LGR5(+) cells were enriched by Magnetic- Activated Cell Sorting (MACS). Cells were applied through a cold BSA-coated 20μM cell strainer (CellTrics) to LS Columns (Miltenyi Biotec) in the above HBSS buffer containing 200 Kunitz units/mL DNAse (Sigma-Aldrich), 0.5% BSA in DPBS (Sigma-Aldrich), and 5μm Y27632. The magnet-bound positive and flow-through negative fractions were analyzed and isolated by FACS. For further details, regarding single cell preparation and MACS, see supplementary Materials and Methods.
Flow Cytometry
Cells were analyzed on a LSRII cytometer (BD Biosciences) and sorted on a MoFlo Astrios (Beckman Coulter). Events first passed through a routine light-scatter and doublet discrimination gate, followed by exclusion of dead cells using 4’,6-diamidino-2-phenylindole (1μM DAPI dilactate; Molecular Probes). Gating strategy set the APC-positive cell population at 0.05-0.1% of the viable MACS flowthrough cells, LGR5(FT+) (Fig. 3C bottom panels; all FACS RNA sequencing-derived data; Fig. 8A, specimen 81, 83). Gating strategy set the APC-positive cell population at 0.05-0.1% of the viable FMO- APC check reagent control for the analysis of samples that were not MACS processed (Fig. 3C top and middle rows; Fig. S2A). Flow cytometric data analysis was performed using Winlist 3D software (Verity) and FlowJo vX.0.7 (Tree Star). See supplementary Materials and Methods for further details.
High Throughput RNA Sequencing (RNA-seq)
RNA was extracted from sorted cells using the RNeasy Micro Kit (Qiagen) with on-column DNase digestion. Due to the small number of cells following FACS and corresponding low level of input RNA, we depleted ribosomal RNAs with RiboGone (Takara Clontech) and prepared sequencing libraries utilizing the SMARTer Stranded RNA-Seq kit (Takara Clontech,). Libraries were multiplexed over 2 lanes and sequenced on a HiSeq sequencer (Illumina). For further details, including alignment and quality control, differential expression testing, and KEGG pathway analyses, see supplementary Materials and Methods.
Comparison to previously published datasets
RNA-seq data from normal colon (Tissue Based Map of the Human Proteome Science REF) and differential gene expression between the LGR5(+) cells and normal colon were calculated as described above (Fig. 6B, C). Overlap between genes overexpressed in LGR5(+) cells with the previously reported Lgr5+ ISC gene signature (Muñoz et al., 2012) was calculated by identifying the overlap between genes present in both the Munoz et al ISC gene expression signature and genes identified as upregulated in LGR5(+) adenoma cells (Fig. 6A; Table 2; Table S2). Statistical significance of the overlap between these gene signatures was calculated using the hypergeometric distribution.
LGR5 Immunohistochemistry
Formalin fixed, paraffin sections were cut at 5-6 microns and rehydrated to water. Heat induced epitope retrieval was performed with FLEX TRS High pH Retrieval buffer (pH 9.01; Agilent Technologies, 154 #K8004; Santa Clara, CA) for 20 minutes (Fig. 1A, C and Fig. 3A). After peroxidase blocking, the antibody LGR5 rabbit monoclonal clone STE-1-89-11.5 (Miltenyi Biotec) was applied at a dilution of 1:50 (Fig. 1A, C) or 1:100 (Fig. 3A) at room temperature for 60 minutes. The FLEX HRP EnVision System (Agilent Technologies) was used for detection with a 10 minute DAB chromagen application. Note, sections freshly cut were compared to those that were stored at room temperature for 4 weeks, and showed more robust LGR5 staining (data not shown). The colon cancer tissue microarray (Fig. 3A; 2 normal, 3 adenoma, 70 adenocarcinoma; 2 core samples per specimen) was freshly cut and provided by BioChain Institute, Inc.
LGR5 Immunohistochemistry scoring for staining intensity in the epithelium and in the stroma
The TMA, along with five additional FFPE normal colon samples from warm cadaveric colon resections, were scored for staining intensity in both the epithelium and then separately in the stroma (Fig. 3). Scoring was conducted by two independent viewers on blinded samples at 8X and 20X magnification. Scoring key: 0 = non-specific or < 1%; 1 = 1-10% or only evident at 20X magnification; 2 = 10-50% or light diffuse staining >50%; 3 = >50%. Stage I (n=24), Stage II (n=24), and Stage III (n=21) tumors were compared to normal colon (n=7) and adenoma (n=3). TMA cancers with grades I & I-II were grouped, termed “Grade I”, and cancers grade II-III & III were grouped, termed “Grade III” for further analyses. LGR5 stromal and epithelial staining for adenomas (n=3), cancer Grade I (n=20), II (n=38), and III (n=10) were compared to normal colon tissue (n=7). For additional details regarding IHC staining and scoring see supplementary Materials and Methods.
LGR5 Immunofluorescence
Rehydrated sections were retrieved with R-Buffer B (pH 8.5; Electron Microscopy Sciences) in a pressurized Retriever 2100 (Electron Microscopy Sciences) overnight. Slides were blocked for 1 hour with 0.5% Triton X-100 and 5% donkey serum and were incubated overnight at 4°C with the following antibodies in 0.05% Tween 20 and 5% donkey serum: anti-LGR5 clone 89.11 (Fig. 1C3C4); anti-OLFM4 antibody (Abcam) (Fig. 8C); anti-defensin5a (Abcam) (Fig. S1A); anti-E-cadherin (BD Bioscience) (Fig. 8C); and anti-E-cadherin (R&D) (Fig. S1A), followed by 1 hour room temperature incubation with the following secondaries: LGR5 plus DAPI (Fig. 1C3/C4; Fig. 2A) and OLFM4 (Fig. 8C) secondaries (biotinylated secondary donkey anti-rabbit; Jackson Immuno Research Labs); defensin5a (Fig. S1A) and E-cadherin secondary (Fig. 8C) (donkey anti-mouse 488; Jackson Immuno Research Labs); E-cadherin secondary (Fig. S1A) (donkey anti-goat 647; Jackson Immuno Research Labs). LGR5 and OLFM4 were amplified with the SK4105 TSA Kit with Alexa 594 tyramide (Invitrogen). For additional IF staining details see supplementary Materials and Methods.
Further Specificity Studies of LGR5 Antibodies and Development of Procedures for Enrichment of LGR5(+) cells by Magnetic-activated Cell Sorting (MACS)
Western analysis was performed with 1881 LGR5(+), wild type 1881 LGR5(−), specimen 282 and 14881. After protein transfer from a Tris-Glycine 4-20% gradient gel, the membrane was incubated with primary antibody, 0.1μg/mL rabbit anti human LGR5 STE-1-89-11.5 (Miltenyi Biotec), in 10mL TBS/Tween/milk powder overnight at 4°C, followed by incubation with secondary HRP goat anti rabbit IgG (Cell Signaling Technology). Flow cytometry spike-in experiments (Fig. S2B) were performed with 1881 LGR5(+) and 1881 LGR5(−) by mixing at varying proportions and analyzing on a LSRII cytometer (BD Biosciences) before and after magnetic separation with rat monoclonal antibody anti-human LGR5 clone 22H2.8 magnetic bead-conjugate (Miltenyi Biotec), with the allophycocyanin (APC) anti-bead check reagent to recognize the bound magnetic beads (Miltenyi Biotec). Detailed information regarding the culture and transfection of the 1881 cell lines, validation of 1881 LGR5 transfection stability by qRT- PCR, flow cytometry/MACS methods, and western analysis, see supplementary Materials and Methods.
Transduction of organoids with lentiviral TCF/LEF GFP reporter and ImageStream Analysis
Colon adenoma specimen 282 organoids were transduced with a lentiviral TCF/LEF GFP reporter (Qiagen) based on Koo et al (Koo et al., 2013) with the major modifications being that 1) organoids were mechanically dissociated in the absence of enzymatic digestions (to avoid single cells), and 2) that incubations were done at 4°C. Organoids were passaged under puromycin selection and transitioned to cystic morphology with L-WRN medium. Single cells were analyzed and imaged with the Amnis ImageStreamX Mark II (EMD Millipore) for co-expression of TCF/LEF-GFP and LGR5-APC fluorescence (Fig 5B,C). For further transduction and ImageStream details see supplemental Materials and Methods.
Association of SMOC2 and DKK4 expression with LGR5 expression
LGR5, SMOC2, and DKK4 mRNA expression were assessed by qRT-PCR and presented as increased expression in L-WRN relative to KGMG, with three adenomas organoids 282, 584, and 590, after transitioning from KGMG to L-WRN. In situ hybridization (ISH) was preformed to assess of localization of SMOC2 and DKK4 relative to LGR5. ISH was performed using the RNAscope 2.0 HD detection kit (Advanced Cell Diagnostics, ADC) according to the standard provided protocol (Fig. 1B, D; Fig 7C, D), and all probes were designed by ADC. See supplemental Material and Methods for further qRT-PCR and ISH details including primer/probe sequences.
Normal Colinic Crypt LGR5(+) Cell Isolation and Culture
Crypts were isolated from three warm autopsy normal colon specimens (Table 1) and processed as previously described (Dame et al., 2014), a modification of Sato et al (Sato et al., 2011a), and normal colon organoid cultures were established (Table 1). LGR5(+)/(−) cells were isolated from early passage organoids (P3 and P5) and sorted (without MACS) into growth medium containing 0.25mg/mL Matrigel. The flow cytometry strategy was as above, with the added gating/analysis for EpCAM expression using a phycoerythrin (PE)-conjugated antibody (BioLegend). The medium composition was roughly based on previous mouse small intestine/colon single cell organoid-forming efficiency studies (Yin et al., 2014, von Furstenberg et al., 2014, Wang et al., 2013, Shimokawa et al., 2017, Jung et al., 2015). Sorted cells were seeded into 4 wells at 500 cells/28μL Matrigel/well (24-well plate). At day 14-17, images were taken and organoids over 100μm were counted to access organoid-forming efficiency (Fig. 8D). Cultures were passaged for approximately 2 months with cryopreservation before ending. For further details including FACS of normal cells and single cell medium components, see supplemental Material and Methods.
Statistical Analyses
For IHC analyses, LGR5 stromal or epithelial staining intensity categories were plotted by tumor stage and grade using boxplots. Differences in LGR5 staining in the stroma or epithelium by stage or grade were quantified by linear regression, treating epithelial or stroma staining intensity (0, 1, 2, or 3) as a continuous dependent variable and either tumor stage or grade as a categorical independent variable, setting normal tissue as the reference group. For qRT-PCR analyses, differences between biological replicates across experimental conditions were assessed using t-test. For both IHC and qRT-PCR analyses, differences were considered statistically significant at p < 0.05. Statistical methodology for genomic variant characterization and RNA-sequencing differential expression analyses can be found in the Supplemental Information.
Data Access: The RNA-seq and genomic variant raw data are publically available at ArrayExpress under accession number E-MTAB-4698.
Disclosures
Olaf Hardt, David Agorku, and Andreas Bosio are full-time employees of Miltenyi Biotec GmbH. Julie Laliberte and Jay Stoerker are full-time employees of Progenity, Inc. The remaining coauthors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Author contributions
Conceptualization: MD, DA, SM, JC, JS, YS, XX, JS, BS; Methodology: MD, DA, SM, OH, PO, DAg, AB; Investigation: MD, DA, SM, PD, PO, AC, XX, JL, EK, GN, DH, AM, YT, DAg, CA; Analysis: JC, JS, HA, JL, PO; Resources: LS, OH, JS, CA; Original Draft: JC, MD, DA; Review-Editing: JS, YS, DB, LS, MW, JV; Supervision JC, JS, YS, DB, MW, JV. All authors had access to the study data and reviewed and approved the final manuscript.
Funding
This study was supported by University of Michigan Comprehensive Cancer Center (M.S.W.) and UMCCC Fund for Discovery Pilot Project Award (J.A.C.) P30CA046592; MCubed (J.A.C.), a seed funding program at the University of Michigan; NIDDK 5P30DK034933 [The University of Michigan Center for Gastrointestinal Research (UMCGR)] and NIH NIAID U19AI116482 (J.R.S.); NCI CA148828 (Y.M.S.); NIH 5R01GM068848 and NIH P50CA130810 (GI SPORE, D.E.B., PI), the Kutsche Family Endowed Chair in the Department of Internal Medicine and the Geriatrics Research Education and Clinical Center at the Ann Arbor VA Medical Center (D.E.B.); NCI 3P30CA046592-26-S3 (M.S.W.); NIH R21 CA181855-01/02/03A1 and NCI R21CA201782-01(J.V.); American College of Surgeons, T32HD007505 (P.H.D.); X.X. was supported by the Research Scholar Award from American Gastroenterological Association; and A.M. was supported by the NIH Cellular and Molecular Biology Training Grant T32GM007315; the University of Washington Laboratory of Developmental Biology was supported by NIH 5R24HD000836 (Ian Glass, PI) from the Eunice Kennedy Shriver National Institute of Child Health & Human Development.
Acknowledgments
We thank Kelly D. Maynard, Veda Yadagiri, Bodrul Islam, Kevin Kim, and Jessica Zhang for their support of the GISPORE Organoid Core facility; Missy Tuck, Brian Kleiner and Kim Gonzalez, clinical coordinators, the University of Michigan Hospitals, for their assistance in obtaining adenoma biopsies; Kristina Fields and Alan Burgess of the Research Histology and IHC Core; Dave Adams and Michael Dellheim of the UM BRCF Flow Cytometry Core; Deborah Postiff and Jackline Barikdar of the Tissue Procurement Core; Jeanne Geskes, Robert Lyons, and Melissa Coon, University of Michigan DNA Sequencing Core Facility; Ashwini Bhasi of the University of Michigan Bioinformatics Core; Robin Kunkel for graphic design, Department of Pathology; and Kaycee White and Muhammad N Aslam of the Histomorphometry Core, Department of Pathology, for their ScanScope service.
REFERENCES
SUPPLEMENTAL REFERENCES




