

Abstract
Is activity in prefrontal cortex (PFC) critical for conscious perception? Major theories of consciousness make distinct predictions about the role of PFC, providing an opportunity to arbitrate between these views empirically. Here we address three common misconceptions: i) PFC lesions do not affect subjective perception; ii) PFC activity does not reflect specific perceptual content; iii) PFC involvement in studies of perceptual awareness is solely driven by the need to make reports required by the experimental tasks, rather than subjective experience per se. These claims are incompatible with empirical findings, unless one focuses only on studies using methods with limited sensitivity. The literature highlights PFC’s essential role in enabling the subjective experience in perception, contra the objective capacity to perform visual tasks; conflating the two can also be a source of confusion.
An opportunity to empirically resolve some enduring controversies
Our theoretical understanding of the neural mechanisms of conscious perception remains primitive. To make progress, investigators have begun emphasizing specific, testable hypotheses. One promising topic that has seen renewed interest is the role of prefrontal cortex (PFC) in consciousness. This empirical question can be directly addressed by experiments, providing an opportunity to arbitrate between several theoretical frameworks (Dehaene and Naccache, 2001; Lamme, 2003, 2006, 2010; Dehaene et al., 2006, 2011; de Gardelle and Kouider, 2009; He and Raichle, 2009, 2010; Kouider et al., 2010; Cleeremans, 2011, 2014; Dehaene and Changeux, 2011; Dehaene, 2014; Koch et al., 2016a, 2016b; Sandberg et al., 2016; Tononi et al., 2016). For example, while the Local Recurrency theory predicts that PFC activity is not critical for perceptual consciousness (Lamme, 2006), the Higher Order (Lau and Rosenthal, 2011) and Global Workspace (Baars, 1997, 2005; Dehaene and Naccache, 2001; Dehaene, 2014) theories assert that PFC activity plays a crucial role in enabling conscious perception. Thus, this is currently one of the more tractable and meaningful issues in the field.
The clinical neuropsychology of consciousness: ‘classical’ case studies
Based mainly on old lesion studies (e.g., Hebb and Penfield, 1940; Fulton, 1949; Mettler, 1949; Brickner, 1952), often mentioned indirectly via secondary source (e.g., Pollen, 1999), the role of prefrontal cortex in consciousness has long been debated (Tse et al., 2005; Kouider et al., 2007; Lau and Rosenthal, 2011; Noy et al., 2015). Several investigators have called this topic into question again recently (Tsuchiya et al., 2015; Koch et al., 2016a, 2016b; Sandberg et al., 2016), asserting that PFC plays a negligible role in consciousness.
However, important details about the extent of lesions in older work are often overlooked, leading to false conclusions. For example, Koch et al. (2016a) cite work on one patient with an “almost complete bilateral frontal lobectomy” (p.319, referencing Brickner, 1952) to provide evidence that lesions to PFC do not impair consciousness. While Brickner (1952) claims that most of Patient A.’s frontal lobe is missing, simple visual inspection of the patient’s brain reveals that massive portions of right PFC remain intact (Figure 1).
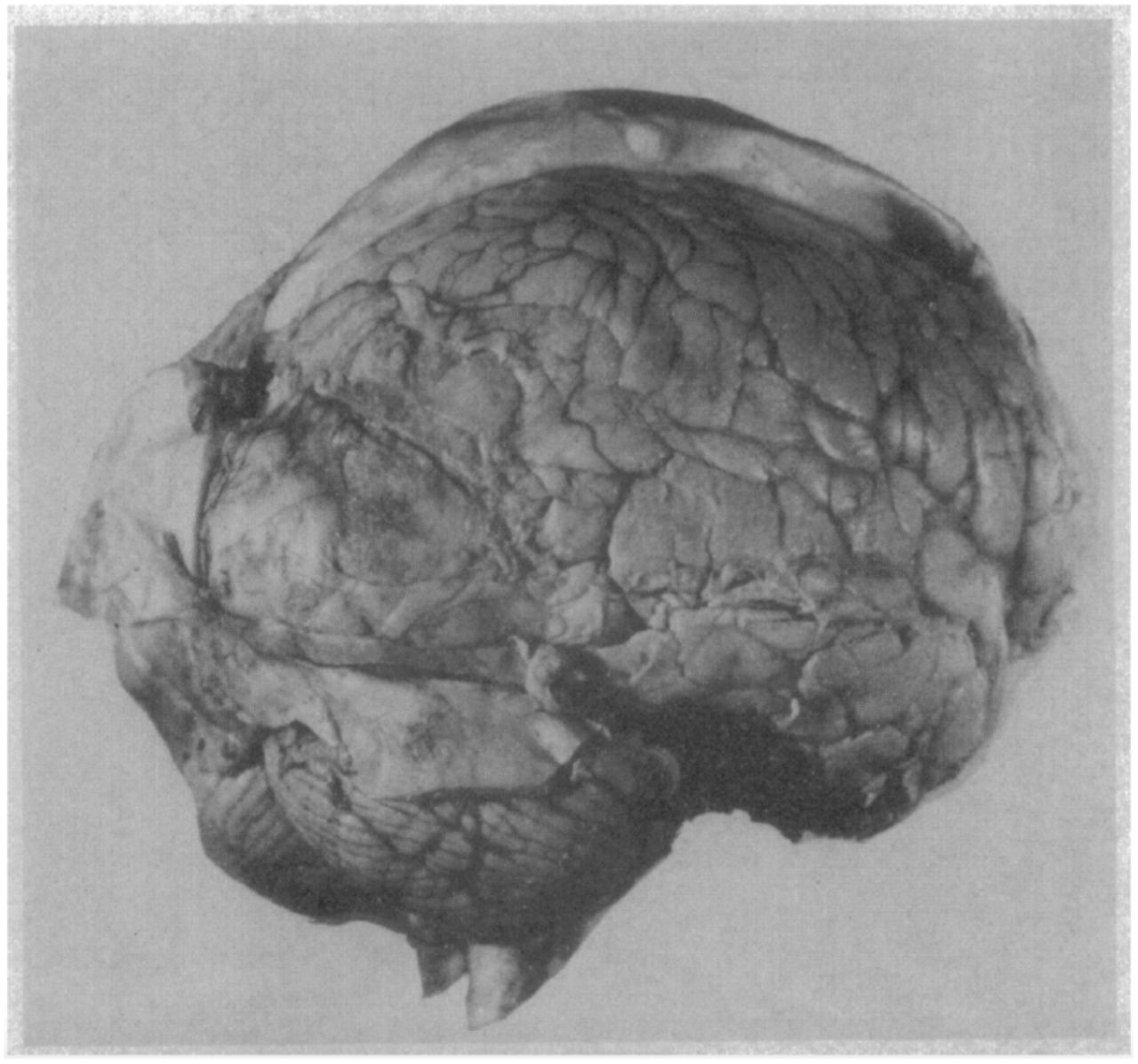
Patient A. was reported as the first human subject to undergo ‘bilateral’ frontal lobectomy. While this patient has previously been described as a classic example of how lesions to PFC do not impair consciousness (Koch et al., 2016), simple visual inspection of this patient’s post-mortem brain image reveals that massive amounts of residual right PFC remained following the surgery. Therefore, the fact that this patient exhibited signs of consciousness following surgery is not surprising. Note also that this patient also had a large posterior meningioma exerting pressure on extrastriate cortices.
Indeed, much older work on psychosurgical patients often cited in discussions of PFC (Hebb and Penfield, 1940; Fulton, 1949; Mettler, 1949) centers on cases with only unilateral or partial removal of frontal cortex, or polar frontal and orbitofrontal resection. For example, Hebb and Penfield (1940) stress a remarkable lack of debilitating symptoms in their patient that had extensive bilateral removal of frontal cortices. While they emphasize that their postoperative report may underestimate the amount of tissue extracted during their procedure, their diagram of the cytoarchitectonic fields mapped by Brodmann (their Fig. 8) shows that much of Broca’s area, superior frontal gyrus, and posterior middle gyrus remained following surgery, which can account for the preserved visual and limited cognitive abilities in the patient. Furthermore, Hebb and Penfield acknowledge only removing approximately one third of the frontal lobes in their summary (p. 438, 1940). Similarly, the patients described by Mettler (pp. 64-77, 1949) are also marked by partial removal of frontal cortical areas. Thus, mischaracterizations of historical data can lead to incorrect conclusions about the role of PFC in consciousness.
Importantly, there are in fact cases where large, near-complete bilateral lesions to the dorsal and ventral lateral PFC fully abolish normal conscious interaction with the external world. One of the authors (RTK) has personally tested patients with such lesions, and often they appear to be almost completely disconnected from - and unaware of - the external world. However, these cases are rarely reported in detail in published scientific articles because such patients are often unable to participate in formal neuropsychological testing (Mettler, 1949). But these patients, similar to patients with bilateral anterior cingulate lesions, often represent the classic clinical observations of severe abulia and perseveration (Barceló and Knight, 2002; Stuss and Knight, 2013), indicating a loss of capacity for conscious interaction with the environment.
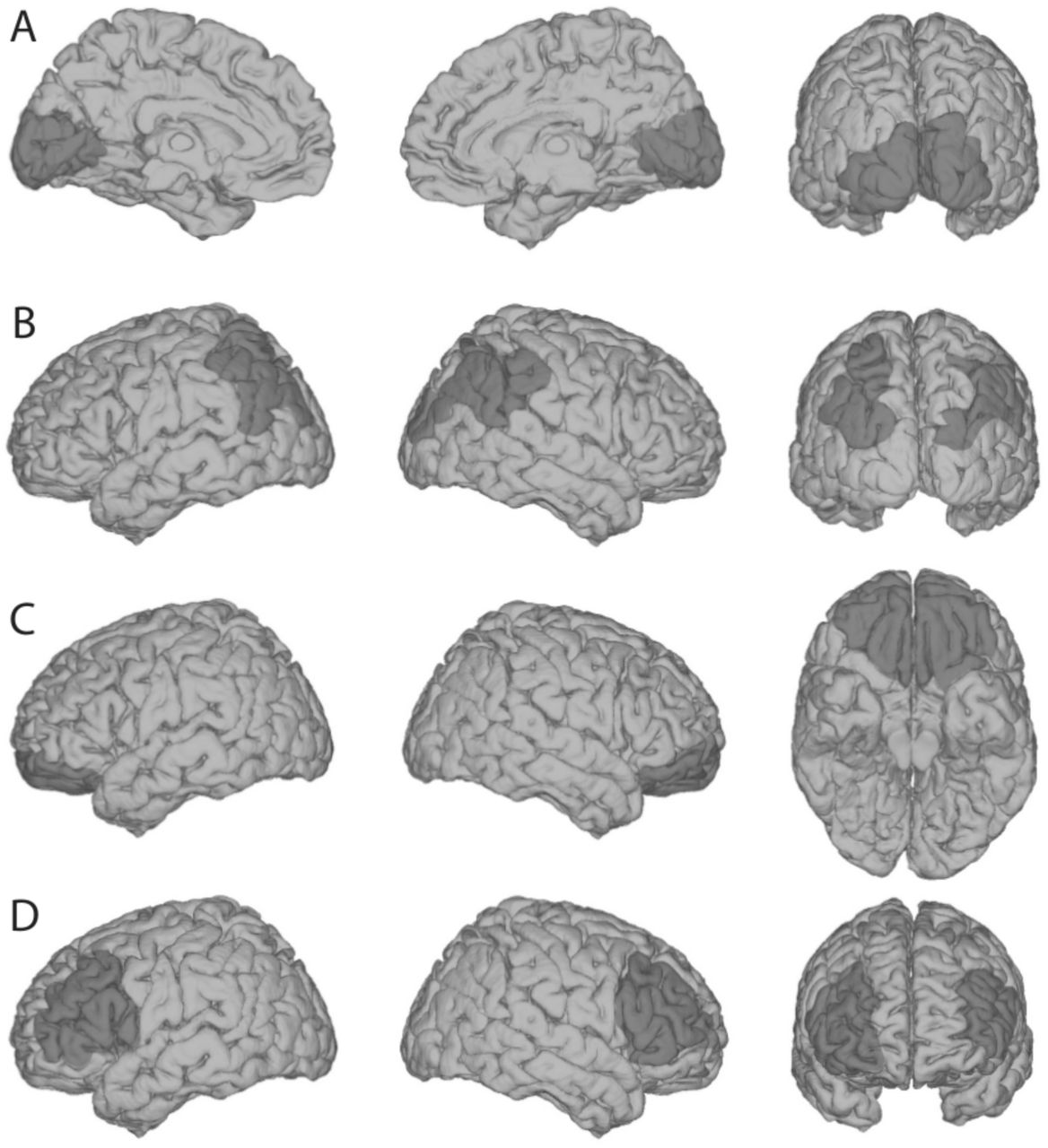
Critically, regarding this general capacity to interact with and be aware of the external world, lesions to areas posterior to the frontal lobes tend to cause minimal, if any, impairments. For example, bilateral occipital lesions (Fig. 2A) can cause cortical blindness, and bilateral parietal lesions (Fig. 2B) can result in impaired visual feature binding (Friedman-Hill et al., 1995) among other visual impairments associated with Balint’s syndrome. However, while these patients have prominent sensory deficits (e.g., in the visual domain), they retain the capacity for being alert, aware and interactive with their surroundings. Similarly, extensive bilateral orbitofrontal lesions cause extensive social regulation deficits (Beer et al., 2003, 2006; Perry et al., 2016), but leave patients cognizant of sensations and fully interactive, albeit often excessively. However, bilateral lateral PFC lesions (Fig. 2D) frequently leave patients abulic, akinetic, unable to consciously process stimuli from various sensory modalities, and unable to have goal-directed interactions with the environment.

Shown here are schematic depictions of typical lesion locations for each of the following types of impairments. (A) Bilateral occipital lesions can cause cortical blindness (Melnick et al., 2016), but preserve an individual’s general awareness of the surrounding world, via other sensory modalities. (B) Bilateral parietal lesions can cause “Balint’s Syndrome,” typically characterized by an inability to perceive multiple objects in the visual field coherently, but again, this leaves basic sensory awareness intact. (C) Bilateral orbitofrontal lesions cause problems with social regulation, but not sensory perception or consciousness. (D) Patients with bilateral lateral PFC lesions are ‘conscious’ in the sense that they are ‘awake’ - as opposed to being in a coma - but they lack the ability to engage meaningfully with the external world with a normal level of conscious awareness. Note that extensive unilateral PFC lesions do not tend to cause these symptoms, highlighting that one functional side of PFC may be sufficient to support general conscious behavior. However, further psychophysical evidence indicates that even unilateral PFC lesioned patients may suffer from specific impoverishment of subjective perceptual experiences (see below; Del Cul et al., 2009; Fleming et al., 2014).
How do lesions to PFC specifically affect subjective perception?
The above discussion highlights that claims about consciousness based on individual case studies can be problematic due to incomplete documentation and limited data, often difficult to access. Fortunately, more recent investigations with carefully controlled psychophysical methods (Del Cul et al., 2009; Fleming et al., 2014) have investigated how PFC lesions specifically impact subjective perceptual experiences in representative samples. Similar to short-term impairments induced by transcranial magnetic stimulation (TMS) (Rounis et al., 2010), lesions to PFC do not fully abolish objective visual task performance capacities, which may explain some previous null findings. But also like TMS, they can render subjective ratings of perception less meaningful (i.e., less diagnostic of actual task performance) (Rounis et al., 2010), by up to 50% (Fleming et al., 2014) (Figure 3). This observed effect was not due to an impairment of general ability for introspection, as it was specific to perception but not memory (Fleming et al., 2014); the specificity of this result was predicted independently based on another study comparing the individual variability of structural correlates for memory and perceptual metacognition in dozens of subjects (McCurdy et al., 2013). Likewise, in another study it has been found that the effect of PFC lesions was more pronounced for subjective ratings than for objective task performance (Del Cul et al., 2009).

In a study by Fleming et al. (2014), healthy controls (HC), patients with lesions to anterior PFC (aPFC), and patients with temporal lobe lesions (TL) were tested in a perception task and a memory task. (A) Overall task performance was equivalent across the three different groups in both tasks. (B) Displayed here (meta-d’ / d’) is a measure of the meaningfulness of subjective confidence ratings, i.e. the degree to which they distinguish between correct and incorrect judgments, expressed on a scale where ‘1’ implies statistical optimality and ‘0’ indicates chance-level trial-by-trial correspondence between confidence and accuracy (Maniscalco and Lau, 2012). This measure showed a nearly 50% decrease in patients with aPFC lesions, compared to patients with temporal lesions and healthy controls, and this effect was specific to perception rather than memory.
Perhaps these studies (Del Cul et al., 2009; Fleming et al., 2014) are written off as idiosyncratic examples, given that both unilateral lesions and disruption by stimulation to lateral PFC tend not to consistently impair objective visual task performance, and this means they should not affect subjective experience, either. For example, in one recent review (Koch et al., 2016a), the authors argue that subjective and objective measures of conscious perception should conform to each other, when “report protocols are implemented judiciously” (p.308, italics added here). However, the distinction between subjective and objective measures is central to the scientific study of consciousness (Merikle et al., 2001; Dienes, 2008; Giles et al., 2016); there is a large literature on how these measures of conscious perception systematically dissociate, and why we need to carefully consider each of them independently (for a review, see Lau, 2008; Lau and Rosenthal, 2011).
Importantly, although objective measures of task performance are often favored because they are easy to quantify, they have relatively limited face validity when it comes to studies of conscious perception. It is well known that blindsight patients (Weiskrantz, 1997) exhibit above-chance objective task performance even though they lack subjective awareness. In this context, that PFC lesions lead to selective impairment to subjective but not objective measures of perception highlights the specific and important roles played by PFC in consciousness. It would be peculiar to write this off, given the current purpose.
Perhaps some may find even a 50% impairment (Fleming et al., 2014) unimpressive, compared to, for example, the effect of lesions to V1, which can often completely abolish ‘conscious’ visual phenomenology (Weiskrantz, 1997; Melnick et al., 2016). However, it is important to note that PFC functions differently from sensory cortices. For instance, neuronal coding in PFC is relatively distributed and shows a high degree of mixed selectivity (Mante et al., 2013; Rigotti et al., 2013). Therefore, to produce a large disruption of perceptual experience, many neurons distributed throughout massive regions of PFC may need to be involved. However, when PFC lesions are large and bilateral, because these same regions support many central cognitive functions (Duncan and Owen, 2000; Miller, 2000; Badre and D’Esposito, 2009; Niendam et al., 2012; Passingham and Wise, 2012), the patients may be so generally impaired that testing them immediately after extensive bilateral PFC damage is often difficult (Mettler, 1949; Knight and Grabowecky, 1995).
Also, the frontal and parietal cortices are densely connected (Barbas and Mesulam, 1981; Petrides and Pandya, 1984; Andersen et al., 1985; Cavada and Goldman-Rakic, 1989; Miller and Cohen, 2001; Croxson et al., 2005). After localized unilateral lesions to PFC, it is known that other parts of the frontal-parietal network can dynamically reorganize to compensate functionally (Voytek et al., 2010). As such, large areas spreading through both frontal and parietal areas may need to be damaged to produce consistently strong effects (Nakamura and Mishkin, 1986). This is compatible with the fact that even for classic ‘textbook’ PFC functions such as working memory, reported studies involving unilateral PFC lesions sometimes only find subtle or null effects (Mackey et al., 2016). Given this context, the highly specific effects of mostly unilateral PFC lesions on subjective conscious perception are striking and convincing (Del Cul et al., 2009; Fleming et al., 2014).
Congruent with the above argument is the fact that chemical inactivations of PFC can sometimes lead to stronger effects, presumably because in these studies the subjects are typically tested right after the PFC is inactivated, leaving little room for alternative circuitries to take over to compensate. For example, one recent study has found that in monkeys, chemical inactivation to the prefrontal cortex impairs memory metacognition (Miyamoto et al., 2017). While this may seem on the surface to be in conflict with Fleming et al.’s (2014) result of lack of memory metacognitive deficits after PFC lesions, one interpretation is that immediately after PFC lesion human patients may show impairment in memory metacognition too, but over time this function recovered, due to activity in the intact remaining side of PFC, or the precuneus, which supports memory metacognition (McCurdy et al., 2013; Richter et al., 2016). In any case, nothing takes away the fact that at the time of testing (Fleming et al., 2014), the metacognitive impairment was specific to perception, not memory, in these patients.
In rodents, it has also been shown that chemical inactivation of orbital PFC selectively impairs subjective confidence judgments without changing olfactory discrimination performance (Lak et al., 2014). In monkeys, chemical inactivation of the pulvinar, specifically the central dorsal region which likely projects to the PFC (Romanski et al., 1997; Shipp, 2003; Pessoa and Adolphs, 2010), also impairs subjective confidence judgements in a visual task, without changing task performance (Komura et al., 2013).
Overall, these results concerning subjective confidence and introspection are congruent with the classical neuropsychological findings discussed in the above section. If a perceptual object is registered in the subject’s mind with no meaningful sense of certainty whatsoever, it is perhaps unsurprising that the subject would fail to act voluntarily in reaction to the relevant object. Conceptually, it is also unclear whether conscious experiences regarding a perceptual object can ever occur without any introspectable sense of certainty that something is presented to us (Lau and Rosenthal, 2011). This is not at all to say that subjective confidence is equivalent to conscious perception, only that the latter likely entails at least some minimal degree of the former.
Finally, there are also numerous studies showing that TMS and lesion to human PFC can affect many other aspects of visual perception (Barceló et al., 2000; Turatto et al., 2004; Ruff et al., 2006; Philiastides et al., 2011; Lee and D’Esposito, 2012; Ritzinger et al., 2012; Chiang et al., 2014; Rahnev et al., 2016), and more recent neuropsychological work has demonstrated that individuals with unilateral damage to PFC often display pronounced deficits on visual tasks. For example, Barceló et al. (2000) showed that individuals with unilateral focal lesions in the lateral prefrontal cortex exhibited impairments in the ability to regulate dependent activity in extrastriate regions and detect visual targets in the visual field contralateral to the damage. Analysis of event-related potentials (ERP) revealed that prefrontal damage reduced stimulus-evoked P1 activity in parietal and temporal-occipital areas, and abolished N2 and P3 activity generated in temporal-parietal cortices, providing critical evidence for prefrontal modulation of visual processing. Indeed, other studies have also demonstrated reduced ERP amplitudes in visual cortices following damage to PFC (Voytek et al., 2010). Thus, suggestions that PFC manipulations do not cause disturbance to conscious perception are grossly incompatible with empirical facts.
Does PFC activity reflect specific perceptual content?
The subjective measures of conscious perception we emphasized above concern subjective visibility and confidence, which are important aspects of conscious perception; these are specifically impaired in blindsight (Weiskrantz, 1997; Persaud et al., 2011; Ko and Lau, 2012; Giles et al., 2016). But to be visually conscious, one also needs to be conscious of specific visual content. It has been argued that because PFC activity does not represent such specific content (Koch et al., 2016a), its role in consciousness is probably limited.
Theoretically, this may not be a critical issue here because in enabling conscious perception, PFC may ‘work together’ with early visual areas where the specific content resides (Lau and Rosenthal, 2011). But importantly, the claim that PFC activity does not reflect specific perceptual content is also empirically false.
The impression that PFC activity may not reflect specific perceptual content is likely due to a selective reading of the fMRI literature, where many studies adopt traditional univariate ‘activation’ approaches. But univariate fMRI is known to have limited sensitivity. Indeed, it has been noted previously that visual content may only be decoded effectively from PFC when careful analysis and advanced modeling strategies are applied (Stokes, 2015; Ester et al., 2015).
Using multivariate ‘decoding’ analyses, Wang et al. (2013) has reported that PFC fMRI pattern activity reflects specific perceptual content, under conditions where the perceptual information was ambiguous, unstable, or needed to be maintained over time. Using similar methods, Cortese et al. (2016) has recently reported that perceptual content can be decoded from PFC even in a simple perceptual decision task. In a somewhat less direct demonstration, Hebart et al. (2016) also reported that the ‘decision variable’ supporting specific perceptual decisions can be decoded from PFC fMRI pattern activity.
Perhaps one may be tempted to dismiss these studies by pointing out that they are individual experiments restricted to specific task conditions, and in some cases the results were statistically weak (e.g., indeed, Cortese et al.’s decoding of perceptual decision from PFC was only significant before Bonferroni correction); such may be the nature of the limited sensitivity of fMRI decoding in these brain regions, given that coding at the neuronal level in PFC may be complex (Mante et al., 2013; Rigotti et al., 2013). But we also have to be aware that vision researchers often do not pay as much attention to PFC as opposed to visual areas. Whereas retinotopic mapping methods are often carefully applied to delineate early visual areas, for PFC, researchers often use statistically inefficient ‘searchlight’ methods based on brain images imperfectly aligned across participants. Sometimes, PFC data are not even acquired or analyzed. Other times positive results are reported without much emphasis. For example, Albers et al. (2013) reported positive PFC decoding findings for visual short-term memory and imagery, but only in the supplementary results section. All of these factors may contribute to the impression of the relative paucity of positive evidence regarding whether fMRI activity in PFC reflects specific perceptual content.
Fortunately, we are not limited to fMRI studies. There are numerous single- and multi-unit recording studies in non-human primates, clearly demonstrating that specific perceptual decisions are represented in PFC (Kim and Shadlen, 1999; Mante et al., 2013; Rigotti et al., 2013). Overall, these studies are compatible with the view that PFC plays a key role in forming perceptual decisions (Heekeren et al., 2004; Philiastides et al., 2011; Szczepanski and Knight, 2014) via ‘reading out’ perceptual information from sensory cortices. Importantly, such decisions are central parts of the perceptual process itself (Green and Swets, 1966; Ratcliff, 1978); they are not ‘post-perceptual’ cognitive decisions. These mechanisms contribute to the subjective percept itself (de Lafuente and Romo, 2006), and have been linked to specific perceptual illusions (Jazayeri and Movshon, 2007).
Is PFC Activity Only Related to Explicit Perceptual Reports?
The last point may not be immediately convincing to some critics of PFC theories of consciousness, as the studies mentioned all involved the subjects’ explicit reporting of perceptual content. It has been claimed that in these studies, activity in PFC does not reflect conscious perception per se; rather, it is confounded by the task demand to report the stimulus (Tsuchiya et al., 2015; Koch et al., 2016a). Such claims are inspired by null findings in recent studies not requiring subjects to explicitly make these reports (Frassle et al., 2014; Tsuchiya et al., 2015).
Proponents of PFC theories are generally aware of this possible criticism, because this line of inquiry is not new (Lumer and Rees, 1999; Tse et al., 2005; Kouider et al., 2007). However, in at least one previous study involving no explicit reports (Lumer and Rees, 1999), positive results were indeed found in PFC. Nevertheless, this still raises the question of why some studies failed to find positive results when demands for report were removed.
Again, the abovementioned issues of limited sensitivity and relatively imprecise analysis methods typically used in fMRI studies, specifically for univariate analysis concerning the PFC, are relevant here. Using more sensitive methods such as direct intracranial electrophysiological recording in human surgical epileptics (electrocorticography, or ECoG), activity related to perceptual awareness in PFC has been reported even when subjects were not required to respond to the stimulus (Noy et al., 2015). It is perhaps noteworthy that the aforementioned study was conducted by a lab that has been critical of PFC theories of consciousness (Malach, 2011), and they reported the positive PFC finding all the same.
More importantly, in direct neuronal recordings in monkeys, it has also been shown that PFC activity reflects specific perceptual content, even when the animal was viewing the stimulus passively (Panagiotaropoulos et al., 2012). One may argue that even under passive viewing, an over-trained animal may still be preparing a report even though it was no longer required (which may have driven prefrontal activity). But in another study, PFC multi-unit activity was found to reflect an unreported feature of a stimulus, even when the animal had to report on a different, orthogonal stimulus feature (Mante et al., 2013). It is unlikely that the animal would implicitly prepare to report on both features, given that only one report was actually required and the task was challenging with near-threshold stimuli (Fig. 4). There was no immediate task-related benefit to represent the irrelevant feature in the brain, and yet they did so in the PFC.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
In a multi-unit recording study (Mante et al., 2013), monkeys had to make saccades to indicate either the direction or the color of the majority of moving dots, depending on the color of a fixation cue. Task-irrelevant / unattended information (e.g., color) was decoded from neuronal activity in the frontal eye field, almost as well as task-relevant information (e.g., motion).
This is not to say that in studies of conscious perception, the demand to make explicit reports does not further drive activity in PFC. It probably does for signals obtained with most imaging methods, and this is congruent with some previous null findings, as well as the comment by Noy et al. (2015) that their positive ECoG findings in PFC were subtle when no report was required. But in the multi-unit recording study mentioned above (Mante et al., 2013), even unreported stimulus features could be robustly decoded, almost at the same level as attended and reported features.
Thus, the recent excitement surrounding the use of so-called ‘no-report’ paradigms may be misguided; PFC activity in conscious perception had already survived such tests, well before their recent advocacy.
Concluding remarks
While debates about consciousness are traditionally dominated by authoritative opinions, the relevant empirical literature has also flourished (Block et al., 2014). It is understandable that keeping up with this growing literature has become difficult, which may explain why many of the studies mentioned here, including those published in venues of the highest profile, or discussed regularly at conferences such as the meetings of the Association for the Scientific Study of Consciousness, were omitted in some recent reviews (Tsuchiya et al., 2015; Koch et al., 2016a). However, consciousness is such an important yet controversial topic, on which weighty claims need to be made with the utmost care. Specifically, individual null findings obtained with one measure, even if true, may not generalize to other measures. Therefore, in reviewing the literature we need to take into account research reflecting different perspectives, as well studies conducted with different methods. When we do so, we find that the specific role of PFC in conscious perception resists even the sternest challenges.
References




