

Abstract
Insect nervous systems are proven and powerful model systems for neuroscience research with wide relevance in biology and medicine. However, descriptions of insect brains have suffered from a lack of a complete and uniform nomenclature. Recognising this problem the Insect Brain Name Working Group produced the 1rst agreed hierarchical nomenclature system for the adult insect brain, using Drosophila melanogaster as the reference framework, with other insect taxa considered to ensure greater consistency and expandability (Ito et al., 2014). Ito et al. (2014) purposely focused on the gnathal regions that account for approximately 50% of the adult CNS. We extend this nomenclature system to the sub-gnathal regions of the adult Drosophila nervous system to provide a nomenclature of the so-called ventral nervous system (VNS), which includes the thoracic and abdominal neuromeres that was not included in the original work and contains the neurons that play critical roles underpinning most 2y behaviours.
Background
Insect nervous systems are proven and powerful model systems for neuroscience research with wide relevance for biology and medicine. Although vast anatomical, physiological and molecu lar data are already available, integrating this information into a common analytical framework would generate an even more powerful resource. Computational analysis combined with digital microscopy now make it possible to consolidate data from multiple techniques and transform how we analyse nervous system function (Jenett et al., 2006; Dance, 2015; Boettiger et al., 2016). It is no longer suffcient to use 2D labelled diagrams and photomicrographs to identify and de1ne anatomical structures, as it is now possible to use multilayer microscopy with computational reconstruction to precisely de1ne and allocate boundaries and structures in 3D. This requires a systematic and consistent nomenclature to precisely de1ne anatomical structures and boundaries. Furthermore the de1nitions and nomenclature need to be rationalised as multiple names can be used for the same structure. The precise inter relationships between structures also need to be speci1ed. Once this is complete, new 1ndings can be more easily added to this framework allowing a complete picture of all knowledge to be more readily accessed and enabling consistent analysis of anatomical data from different studies. Recognising this problem, a consortium of neurobiologists studying arthropod brains, the insect brain name working group (Ito et al., 2014) (IBNWG), was established and produced the 1rst agreed hierarchical nomenclature system for the insect brain, using Drosophila melanogaster as the reference framework, with other insect taxa considered to ensure greater consistency and expandability (Ito et al., 2014). Ito et al. (2014) purposely focused on the gnathal regions that account for approximately 50% of the total adult Central Nervous System. Here we extend this nomenclature system (Ito et al., 2014) to the sub-gnathal regions of the adult Drosophila nervous system to provide a nomenclature of the so-called ventral nervous system (VNS), which includes the thoracic and abdominal neuromeres that was not included in the original work (Ito et al., 2014) and contains the neurons that play a critical role in effecting most 2y behaviors. The VNS is an often overlooked but vital part of the insect central nervous system that is necessary for nearly all motor output (Burrows, 1996; Büschges et al., 2008; Robertson et al., 1982), and will be increasingly important as interest in behavior grows (Lemon et al., 2015; Pitmon et al., 2016; Seeds et al., 2014). Power (1948), Merritt and Murphey (1992) and Boerner and Duch (2010) all made previous attempts to provide anatomical frameworks for the 2y VNS. These core works, plus many other additional studies, have created a rich catalogue of anatomical detail but the inconsistent approach to nomenclature and de1nitions has created ambiguity and confusion. The aim of the Drosophila adult VNS working group (DAVWG) was to create a nomenclature, de1nitions and spatial boundaries for the key anatomical entities of the Drosophila VNS.
Organization of the Working Group
The initial phase of work was similar to that adopted by the original Insect Brain Name Working Group (IBNWG) to create the nomenclature for the Drosophila brain (Costa et al., 2013; Ito et al., 2014). We gathered together researchers who are leading relevant work or those who have made recognized historical contributions to the 1eld of Drosophila VNS. We also included some individuals who have an expertise in development, some with knowledge of the VNS in other insect species, as well as representatives of the original IBNWG 2014. Prior to the workshop, a small group compiled a document listing all of the named regions found in the published literature and from the existing Drosophila anatomy ontology (Costa et al., 2013). The group also created additional representative anatomical datasets where required. These datasets and a list of de1nitions were assembled into a working document and shared online prior to the workshop. At the workshop held at Janelia Research Campus on October 2013 the task group systematically reviewed the published anatomical data and unpublished data provided by the participants, while compiling a proposed list of of nomenclature that was agreed upon by all members. Afterwards, the working document and its content were updated and reviewed by the task group. The document was next distributed to a wider constituency of individuals with a broader species based knowledge as well as any interested parties for feedback. Feedback and amendments were agreed, ultimately resulting in this publication.
Establishing the anatomical framework
Establishment of a systematic nomenclature requires two steps: (1) a clear morphological and spatial de1nition of all the structures to be named and (2) a standard and clear naming scheme. The neuropilar regions of the VNS are typically regarded as being ‘unstructured’ or ‘tangled’, or having a 1ne, granular appearance in sections (Merritt and Murphey, 1992) with different regions distinguished only by general spatial terms such as, ‘intermediate’ (Merritt and Murphey, 1992) or ‘dorsal’. Despite this, different volumes of VNS neuropil can be Defined in relation to 1xed landmarks such as the longitudinal tracts, commissures (Shepherd et al., 2016). Recently, Shepherd et al. (2016) have used the primary projections of neuronal hemilineages to provide a vital organizational principle for de1ning the substructure of the neuropil. Although these landmarks may not always correspond to the underlying functional organisation, they provide a consistent means of structurally de1ning neuropil regions.
To provide an initial framework for establishing distinct boundaries within the VNS, we created confocal datasets that reveal various aspects of VNS structure, including tracts and neuropil. An anti alpha tubulin antibody (Figure 1C) was used to reveal 1brous structures such as longitudinal tracts and commissures. An anti-Drosophila N-cadherin (DNx8) was used to visualize neuropils according to the density of an active-zone-speci1c protein, mirroring exactly the structures revealed by nc82 (bruchpilot) immunostaining (Wagh et al., 2006). This allowed us to distinguish between neuropils that are poor in synapses, such as regions occupied by axons, primary neurites, and glial processes, which are left unlabeled (Figure 1A), and synapse-rich regions such as the primary sensory neuropils (Figure 1D). The anti-neuroglian antibody (Figure 1B) was used to reveal the projections of clonally related neurons in neuroblast (NB) hemilineages (Shepherd et al., 2016). The images obtained with these labeling methods are available via the Brain Explorer function of the FlyBrain Neuron Database (hhttp://ndb.2ybrain.org). To ensure consistency with previous and ongoing studies, we took into account the known projection patterns of various neurons, including those of single identi1ed neurons (giant 1ber neuron) and the trajectories of their 1ber bundles (sensory afferents from the halteres). Information about VNS organization in other insect species was integrated wherever relevant.

Frontal section through midsection of the adult VNS. A shows the neuropil marked by N-cadherin expression in the synapses, B shows neuroglian expression demonstrating the high intensity points of the tracts that guide lineage neurite projection, C shows tubulin expression used to highlight tracts and D highlights the dense neuropil regions associated with sensory connections. All levels are inverted for the purposes of clarity in print.
The naming scheme
The key principle for the rationalisation of the nomenclature was to, wherever possible, integrate the current terminology into the newly developed standard. In doing so, we reused the existing terminology as much as possible and changes were only made where there was a need to remove ambiguity. When multiple names for an anatomical entity had been used in the literature, the principle was to give preference to the name that was in most popular usage. Where there was a need to create new terms we developed a standard and clear naming scheme (e.g. naming of the newly identi1ed structures). We also sought consistency with terms used for earlier developmental stages and in other insects but in all cases avoiding the implication of any homology. We were also driven by the need to separate the naming scheme, driven by morphology, from function. Details about the latter, however, have been included in the de1nition, whenever known.
In developing a standard and clear naming scheme, we wished to encourage a more consistent use of terms. We recognize, however, that users may be inclined to continue using the nomenclature of their choice. In acknowledging this issue, we have captured all known synonyms for each term, including references. Users will be able to search for terms in the DAO using any of these synonyms.
Abbreviations
We adopted a systematic approach when developing abbreviations for each named anatomical entity based on the following points. The 1rst was to adopt abbreviations that are unique across the whole CNS, avoiding abbreviations already in use for unrelated regions in the VNS and the brain (for disambiguation purposes, the letter case is ignored). The second was to create a system in which related entities would be easily recognizable (for example, all commissures). Finally, we tried to be consistent with standard neuroanatomical nomenclature e.g. BrainName (Ito et al., 2014). The reasoning behind each abbreviation change was recorded and embedded in the definition.
When referring to the neuromere and related structures, abbreviations were changed from a single letter or number to ‘Pro’,’Meso’ and ‘Meta’. This removed confusion with positional abbreviations such as posterior or medial.
The use of the single letter ‘N’, which is used widely (neuromere, neuropil, nerve, neuron) was reserved, at this level for “nerve” and the other larger gross anatomy structures differentiated with additional letters (e.g. ‘Nm’ for neuromere and ‘Np’ for neuropil). The letter ‘C’ was used to identify commissures.
In cases where multiple abbreviations had developed for speci1c structures the abbreviation that provided the clearest indication with least likelihood of confusion was agreed by consensus.
Known abbreviations were captured as synonyms and references were added whenever possible.
Axis orientation
Unlike the cephalic neuromeres (Ito et al., 2014) the issue of specifying general axis orientation for the VNS is relatively straightforward. The neuroaxis and the body axis are the same, with the prothoracic neuromere being anteriormost and the abdomen (abdominal ganglionic complex) being the most posterior. In the dorsal/ventral plane the tectulum is superior or dorsal and the leg nerves inferior or ventral. The left and right sides are applied as if the sample is viewed from the above (superior/dorsal). The axis is shown in all 1gures by arrows pointing towards the anterior (A), superior/dorsal (S) and the right (R).
Identifying and de1ning structures
Neuromere Boundaries
The thoracic neuromeres consist of three paired enlargements at the anterior end of the VNS, corresponding to the prothoracic, mesothoracic and metathoracic segments, and a single posterior, dorsally located mass, the abdominal neuromeres, that is composed of all the fused abdominal neuromeres.
The scaffold of axon 1bres revealed by neuroglian expression provides a tool to de1ne the internal segmented structure of the neuropils of the VNS. Since each neuromere is founded by a speci1c set of NBs, the neuroglian bundles produced by their progeny create a neuromere speci1c set of anatomical markers that provide a framework to de1ne each neuromere as well as the tectulum and many of the commissures.
Summary of the nomenclature for VNS structures.
Tracts and Commissures
Power (1948) Defined, later volumetrically clari1ed by Boerner and Duch (2010), the major commissures of the adult VNS. Truman et al. (2004) showed that in the larva the postembryonic secondary lineages crossed the midline via speci1c and invariant commissural pathways. Using the neuroglian labelled tracts Shepherd et al. (2016) were able to link the larval commissures to their adult counterparts to reveal the developmental origins of many of the commissures in the adult (VNS). This work not only con1rmed the developmental origins of the commissures identi1ed by Power (1948) but also identi1ed several smaller commissures not previously observed.
The Tectulum
In the transverse plane the neuroglian tracts also help to de1ne another substructure within the VNS: the tectulum. Power (1948) Defined this region as a "distinct subdivision of the thoracic regions of the [VNS]. The region forms a saddle-like structure located dorsally primarily over the mesothoracic neuropil but extending over the posterior-most region of the prothoracic neuromere and the anteriormost region of the metathoracic neuropil". Despite the subsequent work done on the VNS this remained the best de1nition of the tectulum.
The neuroglian framework of lineage derived from primary neurite tracts provides a clear and easily recognisable boundary that more precisely circumscribes the tectulum (Shepherd et al., 2016). The tectulum is the dorsal region of the neuropil posterior to the anterior most limits of the T1 hemilineage 12B neuroglian tract but dorsal to the tracts from the T1 hemilineages 12B, 6A and B, 23 17 and 18. It extends posteriorly through T2 remaining dorsal to the tracts from hemilineages 3B, 6A and B and 23 to the entry point of hemilineage 3A in T3.
The Peripheral Nerves
Most of the peripheral nerves have been historically Defined by Power (1948) in which case the de1nitions were simply collated. However for some of the abdominal nerves we decided to use later terms (Shepherd and Smith, 1996) which names each nerve according to its neuromere of origin, giving them a more consistent naming scheme than the original.
Discussion
In this project, we addressed two primary issues in order to create a clearer understanding of the VNS structure and to promote dialogue among neuroscience researchers. The 1rst was to establish a common anatomical framework to precisely de1ne and describe, textually and spatially, the anatomical organization of the VNS. The second was to create a clear and consistent naming scheme for each anatomical entity. The detailed VNS map we provide will be invaluable for integrating past and future work into a common space, thereby contributing to new lines of investigation. In addition, our effort will also inform researchers working with other insects, providing them with a template that could be adapted to their own model organism.
Although the nomenclature developed in this project will serve as the standard, we acknowledge that to be useful it must be maintained as a ‘living’ process and evolve as our understanding of the VNS structure and function grows. Future revisions and additions will be required and this will be handled via the existing online system for posting anatomy ontology suggestions located at http://github.com/FlyBase/drosophila-anatomy-developmental-ontology/issues and maintained by VirtualFlyBrain.org.
Unlike the brain, the VNS in insects demonstrates extraordinary diversity in its gross organisation and structure (Niven et al., 2008). There is, however, a large anatomical literature for several insect groups that exhibit markedly different VNS structures (e.g. grasshoppers, crickets and moths) that sometimes use the same terms as used for Drosophila. Whilst it is desirable to describe the correspondence of terminology between species, as was done for the more conserved insect brain, for the VNS this is not so readily achievable and only possible for major structures (neuromere, tract, nerve). To create consistent cross species terminology for more detailed structures requires a programme of work to con1rm homology rather than rely on inference from similar structure. For this reason we elected to not expand this nomenclature to other insects. It is intended that our work will provide a framework to then explore cross species homologies in the VNS.
Acknowledgments
We thank Gerald M. Rubin and the staff of the Howard Hughes Medical Institute’s Janelia Research Campus for hosting our workshop. Special thanks to Wyatt Korff and the other members of the organising committee‡ for such a successful workshop.
This work was initially supported in part by grants EP/F500385/1 and BB/F529254/1 for the University of Edinburgh School of Informatics Doctoral Training Centre in Neuroinformatics and Computational Neuroscience (http://www.anc.ed.ac.uk/dtc) from the UK Engineering and Physical Sciences Research Council (EPSRC), UK Biotechnology and Biological Sciences Research Council (BBSRC), and the UK Medical Research Council (MRC). Finally by the Welcome Trust as part of the ’Virtual Fly Brain: a global informatics hub for Drosophila neurobiology’ WT105023MA.
Appendix 1
Overview of nomenclature
Here we provide the agreed anatomy terms, each with a short description. We also highlight any changes we have made from what was considered the classical de1nition with the group’s reasoning for making the change.
Ventral Nervous System (VNS)
The non-cephalic division of the central nervous system consolidated into a single ganglion located in the ventral thorax. The ganglion contains all of the thoracic and abdominal neuromeres (Figure 1). For this reason it was called the thoracicoabdominal ganglion by Power (1948) but is often referred to as the ventral nerve cord (VNC) although the VNC would include the suboesophageal ganglion (Niven et al., 2008). The VNS ganglion has three paired enlargements at its anterior end, thoracic neuromeres, which correspond to the prothoracic, mesothoracic and metathoracic segments, with a single posterior, dorsally located mass, the abdominal neuromeres that contains all the fused abdominal neuromeres.
Changes:
Although the name thoracico-abdominal ganglion (TAG) (Power, 1948) was initially agreed by the group, many who worked in both larvae and adult felt that ventral nerve cord (VNC) was more appropriate and indeed was often used in the adult. However, this structure includes the suboesophageal ganglion. Ultimately, ventral nervous system (VNS) was agreed upon with all alternatives recorded as synonyms.
Synonyms:
VNS (Proposed by group),
thoracico-abdominal ganglion (Power, 1948),
thoracico-abdominal ganglia (Lundquist and Nässel, 1990),
adult ventral nerve cord (Niven et al., 2008),
VNC [BROAD] (Niven et al., 2008),
TAC (Merritt and Murphey, 1992),
TAG (Ito et al., 2014),
ThGng (Miller and Demerec, 1950),
thoracic + abdominal ganglion,
T1 + T2 + T3 + A1 + A2 + A3 + A4 + A5 + A7 + A8,
thoracico-abdominal center (Power, 1948; Merritt and Murphey, 1992),
thoracic nerve center (Miller and Demerec, 1950),
abdominal ganglion RELATED (Power, 1948).
Dense Neuropil
Dense neuropil, as referred to later in this document, indicates more tightly packed synaptic regions. It can be identi1ed by a notably brighter signal than that from the general surrounding neuropil when imaged using standard immuno2uorescence confocal microscopy targeting either NC82 or N-cadherin. See Figure 1(D) for an example.
Thoracic Neuromeres
The three thoracic neuromeres are segmentally homologous and share common structural features. The detailed organisation of serially homologous tracts and neuropil regions within the leg neuropils, as well as the angle of entry of the relevant leg nerve into the VNS, indicate that the thoracic neuromeres have undergone rotation during development. This is such that the prothoracic neuromeres are rotated anteriorly, the mesothoracic and metathoracic and neuromeres rotated posteriorly.

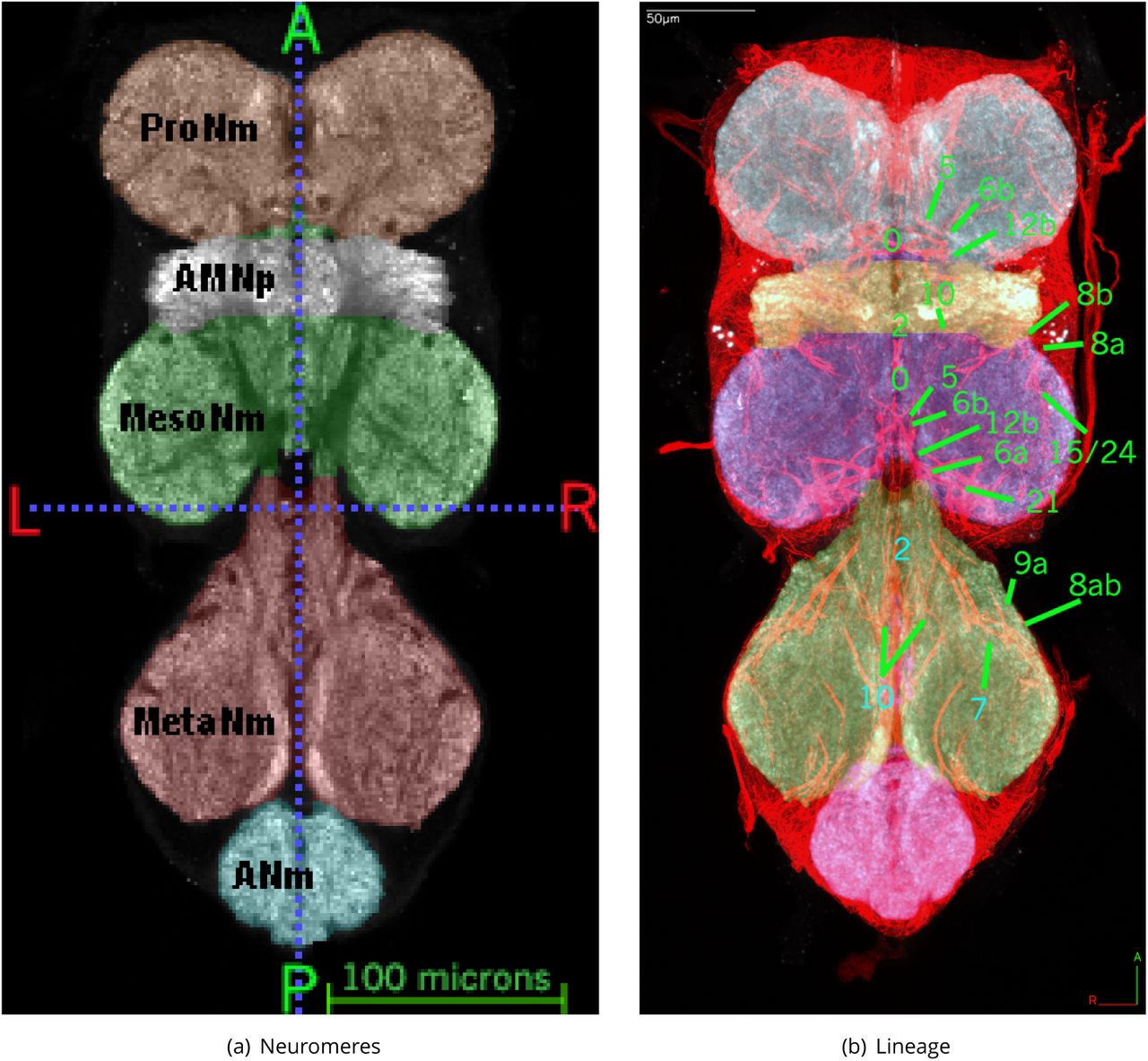
Frontal section through midsection of adult VNS template with painted neuromeres also showing lineage tract boundaries. Shown in Figure 1(a) is the labelled neuromeres: Prothoracic Neuromere ProNm (described in Prothoracic Neuromere (ProNm)), Accessory Mesothoracic Neuropil AMNp (described in Accessory Mesothoracic Neuropil (AMNp)), Mesothoracic Neuromere MesoNm (described in Mesothoracic Neuromere (MesoNm)), Metathoracic Neuromere MetaNm (described in Metathoracic Neuromere (MetaNm)) and Abdominal Neuromeres ANm (described in Abdominal Neuromeres (ANm)). To show the lineage basis for internal boundaries Figure 1(b) shows a thick (22 µm) midsection slice of neuroglian (red) with the labelled neuropil (painted greyscale), showing the painted neuromeres with labelled examples of the lineage tracts de1ning the internal boundaries (see de1nitions in Prothoracic Neuromere (ProNm) Accessory Mesothoracic Neuropil (AMNp) & Metathoracic Neuromere (MetaNm) Abdominal Neuromeres (ANm)).
Prothoracic Neuromere (ProNm)
Defined region shown in Appendix 1 Figure 2
The anteriormost of the 4 major neuropils that makes up the VNS and derives almost completely from the somata and projections of central neurons derived from the prothoracic array of neuroblasts, as well as the axonal projections of sensory afferents from the prothoracic legs and and prothorax. The posterior margin is Defined by the neuroglian tracts from the posterior T1 hemilineages 0, 2, 3, 6, 11 and 19, all of which project anteriorly into the neuromere (Figure 1(b)). The paired neuropil is formed as two more or less spherical masses.
Changes:
The abbreviation was changed to resolve the clash for the original abbreviation PN with those for both the prosternal nerve and the adult antennal lobe projection neuron. The internal boundaries were clari1ed in relation to lineage tracts (Shepherd et al., 2016).
Synonyms:
ProNm (Proposed by group),
Pro (Merritt and Murphey, 1992),
PN (Power, 1948).
Mesothoracic Neuromere (MesoNm)
Defined region shown in Appendix 1 Figure 3
The 2nd (anterior-posterior) of the 4 major neuropils of the VNS that derives almost completely from the somata and projections of central neurons derived from the mesothoracic array of neuroblasts as well as the axonal projections of sensory afferents from the mesothoracic legs and and mesothorax. The mesothoracic neuromere is closely associated with two distinct subdivisions of the thoracic neuropils: the accessory mesothoracic neuromere and the tectulum. The mesothoracic neuromere is contained anteriorly by the neuroglian tracts hemilineages from the anterior T2 hemilineages 2, 7, 8, 9, 10, 15 and 16, all of which project posteriorly into the neuromere and de1ne the anterior margin of the neuropil (Figure 1(b)). Similarly the posterior margin of the neuromere is Defined by the neuroglian tracts from the posterior hemilineages 0, 3, 6, 11, 19, and 21, all of which project anteriorly into the neuromere (Figure 1(b)).
Changes:
The abbreviation was changed for consistency. The internal boundaries were clari1ed in relation to lineage tracts (Shepherd et al., 2016).
Synonyms:
MesoNm (Proposed by group),
Meso (Merritt and Murphey, 1992),
MN (Power, 1948).
Accessory Mesothoracic Neuropil (AMNp)
Defined region shown in Appendix 1 Figure 4
The accessory mesothoracic neuropil (AMNp) is a distinct subdivision of the mesothoracic neuromere. It is formed of dense neuropil (as seen by brighter signal in NC82 or N-cadherin staining) largely from the sensory afferents entering the VNS via the Anterior Dorsal Median Nerve (ADMN) (Power, 1948). The AMNp sits at the interface between the proand mesothoracic neuromeres from which it is morphologically distinct with the tectulum (Tct) forming the dorsal boundary. The AMNp is bounded anteriorly by the posterior T1 hemilineages 0, 5, 6, 11, 19 and 23 and posteriorly by the anterior T2 hemilineages 1, 2, 7, 8, 9, 15, and 16 (Figure 1(b)).
Changes:
After careful consideration it was concluded and agreed by all that the accessory region was indicated by the dense neuropil (see description) and that any area posterior to the prothoracic neuromere lineage boundary but not dense neuropil should be considered part of the mesothoracic neuromere. This also meant that the accessory region should more correctly be labelled neuropil rather than a neuromere. The internal boundaries were clari1ed in relation to lineage tracts (Shepherd et al., 2016).

The proposed volume de1nition for the Prothoracic Neuromere (ProNm). Maximum projections from the labelled orthogonal plane with ProNm shown in green and neuropil (Bruchpilot) shown in magenta. Regions were painted using ITK-SNAP by Robert Court based on literature and group discussions to clarify boundaries.

The proposed volume de1nition for the Mesothoracic Neuromere (MesoNm). Maximum projections from the labelled orthogonal plane with MesoNm shown in green and neuropil (Bruchpilot) shown in magenta. Regions were painted using ITK-SNAP by Robert Court based on literature and group discussions to clarify boundaries.
Synonyms:
AMNp (Proposed by group),
accessory mesothoracic neuromere (Power, 1948),
Acc Meso (Merritt and Murphey, 1992),
AMN (Power, 1948),
ovoid (Merritt and Murphey, 1992).
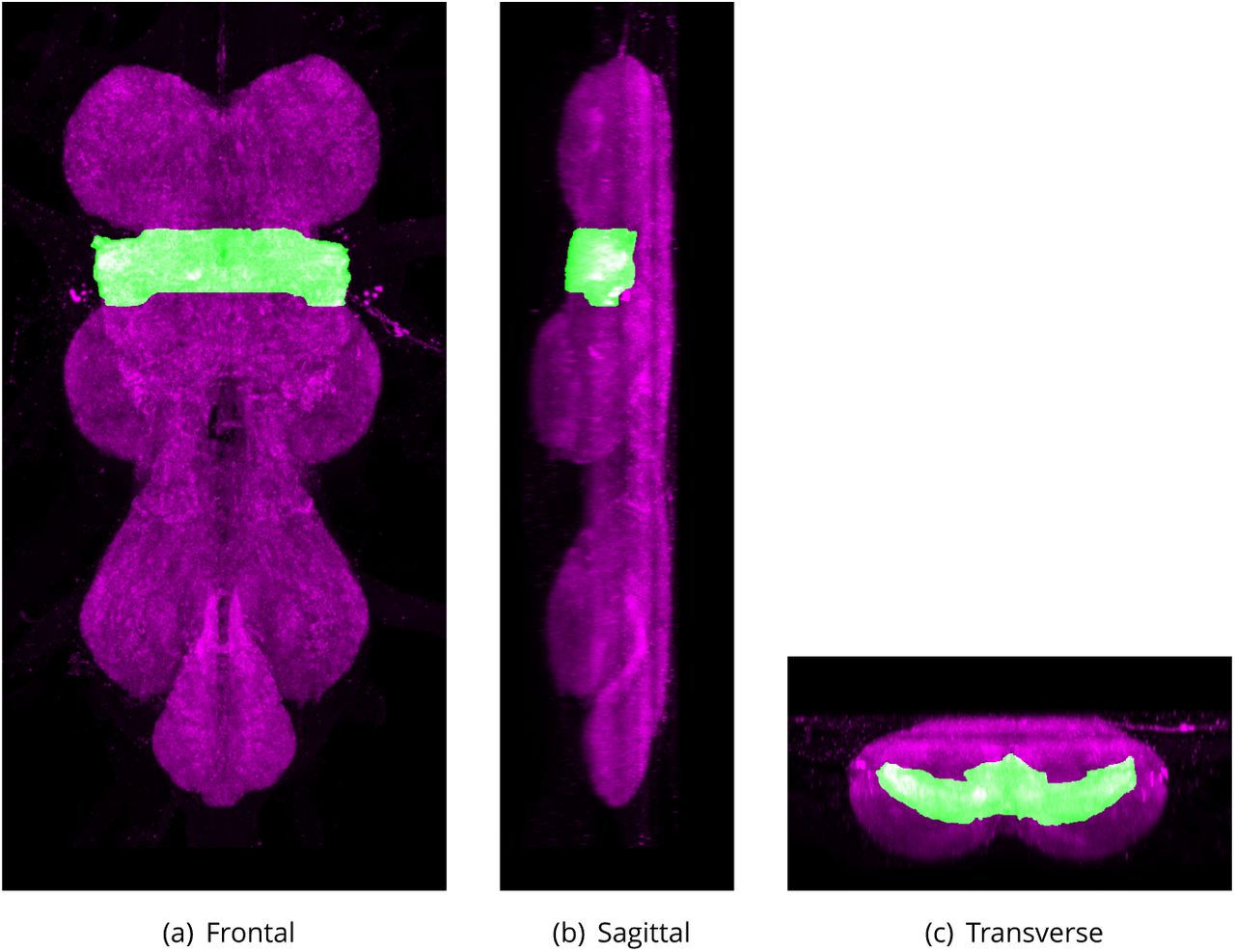
Tectulum (Tct)
Defined region shown in Appendix 1 Figure 5
The tectulum is a distinct subdivision of the thoracic regions of the VNS. The region forms a saddle-like structure located dorsally, primarily over the accessory mesothoracic neuropil (AMNp) and the mesothoracic neuromere (MesoNm), but extending over the posterior-most region of the prothoracic neuromere (ProNm) and the anteriormost region of the metathoracic neuromere (MetaNm). Its internal boundaries within the VNS can be Defined as the dorsal region of the neuropil posterior to the anteriormost limits of the hemilineage 12B neuroglian bundle in ProNm, but dorsal to the bundles from 12B, 6B, 23 17, 18B. Extending posteriorly through MesoNm to the entry point of hemilineage 3 in MetaNm.
Changes:
Abbreviation changed from Power’s ‘T’ (Power, 1948) to avoid potential confusion. The internal boundaries were clari1ed in relation to lineage tracts (Shepherd et al., 2016).
Synonyms:
Tct (Proposed by group),
T (Power, 1948),
2ight neuropil RELATED (Power, 1948; Leise, 1991).
Flight Neuropil (FNp)
Neuropil that contains branches from neurons that drive wing elevators and depressors but also receives projections from 2ight proprioceptors wing hinge receptors and wing sensory hairs (Leise, 1991).
Synonyms:
FNp (Proposed by group).
Metathoracic Neuromere (MetaNm)
Defined region shown in Appendix 1 Figure 6
The 3rd (anterior-posterior) of the 4 major neuropils that make up the VNS (pro-, mesoand metathoracic neuromeres and ANm). It derives almost completely from the somata and projections of central neurons derived from the metathoracic array of neuroblasts as well as the axonal projections of sensory afferents from the metathoracic legs and and metathorax.
The metathoracic neuromere is contained anteriorly by the neuroglian tracts associated with the anterior T3 hemilineages 2, 7, 8, 9, 10, 15 and 16, all of which project posteriorly into the neuromere and de1ne the anterior margin of the neuropil (Figure 1(b)). Similarly, the posterior margin of the neuromere is Defined by the neuroglian tracts from the posterior hemilineages 0, 3, 6, 19, 20/22 and 21, all of which project anteriorly into the neuromere.
Changes:
The abbreviation was changed for consistency. The internal boundaries were clari1ed in relation to lineage tracts (Shepherd et al., 2016).
MetaNm (Proposed by group),
Meta (Merritt and Murphey, 1992),
MtN (Power, 1948).
Abdominal Neuromeres (ANm)

The proposed volume de1nition for the Accessory Mesothoracic Neuropil (AMNp). Maximum projections from the labelled orthogonal plane with AMNp shown in green and neuropil (Bruchpilot) shown in magenta. Regions were painted using ITK-SNAP by Robert Court based on literature and group discussions to clarify boundaries.

The proposed volume de1nition for the Tectulum (Tct). Maximum projections from the labelled orthogonal plane with Tct shown in green and neuropil (Bruchpilot) shown in magenta. Regions were painted using ITK-SNAP by Robert Court based on literature and group discussions to clarify boundaries.
Defined region shown in Appendix 1 Figure 7
The 4th major neuropil situated at the posterior end of the VNS, composed of the fused neuromeres A1 through A8. The lineage composition of the abdominal neuromeres has not been undertaken but the anterior limit of the ANm is Defined by reference to the neuroglian bundles delineating the posterior limit of the MetaNm. The neuropil regions posterior to the neuroglian tracts of hemilineages 0, 5, 6, 19 and 23 de1ne the ANm.
Synonyms:
ANm (Proposed by group),
AbGng (Miller and Demerec, 1950),
ac (Power, 1948),
A1+A2+A3+A4+A5+A6+A7+A8,
abdominal ganglia (Merritt and Murphey, 1992),
abdominal ganglion (Yu et al., 2010),
abdominal center (Power, 1948),
abdominal nerve center (Miller and Demerec, 1950).
The Commissures
The neuroglian tracts and lineage speci1c green 2orescent protein (GFP) expression reveal the developmental origins of many of the commissures in the adult VNS. In the larval VNS there are 5 major commissures in each neuromere: the anterior (aD) and posterior (pD) dorsal commissures, the anterior (aI) and posterior (pI) intermediate commissures, and the anterior ventral (aV) commissure (Truman et al., 2004). The postembryonic hemilineages of the larval VNS cross the midline in speci1c and invariant commissures (Truman et al., 2004). Identi1cation of the postembryonic hemilineages in the adult provides a de1nitive marker to link larval and adult VNS commissures (Power, 1948; Merritt and Murphey, 1992). From this, it is possible to develop a nomenclature for some of the adult commissures that recognises their larval origins (Shepherd et al., 2016).
Prothoracic Anterior Ventral Anterior Commissure (ProAVAC)
ProAVAC is a ventral commissure formed by the axons of the ProNm hemilineage 1A as they cross the midline at the anteriormost margin of the prothoracic neuromere. The ProAVAC is ventrally located and lies at the ventralmost margins of the VNS, outside of the neuropil and cell cortex, crossing the midline anterior to the axon bundles from hemilineage 2A (Shepherd et al., 2016).
Synonyms:
ProAVAC (Proposed by group).
Prothoracic Anterior Ventral Posterior Commissure (ProAVPC)
The ProAVPC is a ventral commissure formed by the axons of ProNm hemilineage 13B as they cross the midline at the anterior of the prothoracic neuromere. The ProAVPC is ventrally located and crosses the midline posterior to the axon bundles from hemilineage 2A (Shepherd et al., 2016).
Synonyms:
ProAVPC (Proposed by group).

The proposed volume de1nition for the Metathoracic Neuromere (MetaNm). Maximum projections from the labelled orthogonal plane with MetaNm shown in green and neuropil (Bruchpilot) shown in magenta. Regions were painted using ITK-SNAP by Robert Court based on literature and group discussions to clarify boundaries.

The proposed volume de1nition for the Abdominal Neuromeres (ANm). Maximum projections from the labelled orthogonal plane with ANm shown in green and neuropil (Bruchpilot) shown in magenta. Regions were painted using ITK-SNAP by Robert Court based on literature and group discussions to clarify boundaries.
Mesothoracic Anterior Ventral Anterior Commissure (MesoAVAC)
The MesoAVAC is a ventral commissure formed by the axons of the MesoNm hemilineage 1A as they cross the midline at the anterior of the mesothoracic neuromere. The MesoAVAC s ventrally located and lies at the ventralmost margins of the VNS, outside of the neuropil and cell cortex, crossing the midline anterior to the axon bundles from hemilineage 2A (Shepherd et al., 2016).
This commissure was originally identi1ed by Power (1948) as the Accessory Prothoracic Commissure.
Synonyms:
MesoAVAC (Proposed by group),
Accessory Prothoracic Commissure (Power, 1948),
APC (Power, 1948).
Mesothoracic Anterior Ventral Posterior Commissure (MesoAVPC)
The MesoAVPC is a ventral commissure formed by the axons of the MesoNm hemilineage 13B as they cross the midline at the anterior mesothoracic neuromere. The commissure is ventrally located and crosses the midline posterior to the axon bundles from hemilineage 2A (Shepherd et al., 2016).
This commissure was originally identi1ed by Power (1948) as the Ventral Accessory Commissure of the Mesothoracic Neuromere.
Synonyms:
MesoAVPC (Proposed by group),
Ventral Accessory Commissure of the Mesothoracic Neuromere (Power, 1948),
VAC (Power, 1948).
Metathoracic Anterior Ventral Anterior Commissure (MetaAVAC)
The MetaAVAC is a ventral commissure formed by the axons of the MetaNm hemilineage 1A as they cross the midline at the anterior of the metathoracic neuromere. The MetaAVAC is ventrally located and lies at the ventralmost margins of the VNS, outside of the neuropil and cell cortex, crossing the midline anterior to the axon bundles from hemilineage 2A (Shepherd et al., 2016).
This commissure was originally identi1ed by Power (1948) as the Accessory Commissure of the Metathoracic Neuromere.
Synonyms:
MetaAVAC (Proposed by group),
Accessory Commissure of the Metathoracic Neuromere (Power, 1948),
ACM (Power, 1948).
Metathoracic Anterior Ventral Posterior Commissure (MetaAVPC)
The MetaAVPC is a ventral commissure formed by the axons of the MetaNm hemilineage 13B as they cross the midline at the anterior of the mesothoracic neuromere. The commissure is ventrally located and crosses the midline posterior to the axon bundles from hemilineage 2A (Shepherd et al., 2016).
Synonyms:
MetaAVPC (Proposed by group).
Prothoracic Anterior Intermediate Commissure (ProAIC)
The ProAIC is a ventral commissure formed by the axons of the ProNm hemilineage 10B as they cross the midline in the anterior of the prothoracic neuromere. The commissure crosses the midline posterior to the axon bundles from hemilineage 2A (Shepherd et al., 2016). The ProAIC is posterior to the ProAVAC and unlike ProAVAC lies within the neuropil.
Synonyms:
ProAIC (Proposed by group).
Mesothoracic Anterior Intermediate Ventral Commissure (MesoAIVC)
The MesoAVIC is a ventral commissure formed by the axons of the MesoNm hemilineage 10B as they cross the midline in the anterior of the mesothoracic neuromere. The commissure crosses the midline posterior to the axon bundles from hemilineage 2A (Shepherd et al., 2016). The MesoAVIC is posterior to the MesoAVAC and unlike MesoVAC lies within the neuropil.
Synonyms:
MesoAIVC (Proposed by group).
Mesothoracic Anterior Intermediate Dorsal Commissure (MesoAIDC)
MesoAIDC is a dorsally located commissure formed by the axons of the mesothoracic hemilineage 18B as they cross the midline in the anterior of the mesothoracic neuromere. The axons cross the midline anterior to the axons from hemilineage 2A (Shepherd et al., 2016).
Synonyms:
MesoAIDC (Proposed by group).
Metathoracic Anterior Intermediate Dorsal Commissure (MetaAIDC)
The MetaAIDC is a dorsally located commissure formed by the axons of the metathoracic hemilineage 18B as they cross the midline in the anterior of the metathoracic neuromere. The axons cross the midline anterior to the axons from hemilineage 2A (Shepherd et al., 2016).
Synonyms:
MetaAIDC (Proposed by group).
Prothoracic Posterior Intermediate Anterior Commissure (ProPIAC)
ProPIAC is a dorsally located commissure formed by the axons of the prothoracic hemilineage 8B as they cross the midline in the anterior of the prothoracic neuromere. The axons cross the midline posterior to the axons from hemilineage 2A (Shepherd et al., 2016).
Synonyms:
ProPIAC (Proposed by group).
Prothoracic Posterior Intermediate Posterior Commissure (ProPIPC)
The ProPIPC is a posterior commissure formed by the axons of hemilineages 6B and 12B as they cross the midline in the prothoracic neuromere (Shepherd et al., 2016).
Synonyms:
ProPIPC (Proposed by group).
Prothoracic Posterior Intermediate Anterior Dorsal Commissure (ProPIDC)
The ProPIDC is a dorsally located commissure formed by the axons of hemilineage 7B as they cross the midline in the anterior prothoracic neuromere (Shepherd et al., 2016).
Synonyms:
ProPIDC (Proposed by group).
Prothoracic Posterior Intermediate Anterior Ventral Commissure (ProPIVC)
The ProPIVC is a commissure at a mid dorso-ventral plane formed by the axons from hemilineage 5B as they cross the midline in the prothoracic neuromere (Shepherd et al., 2016).
Synonyms:
ProPIVC (Proposed by group).
Mesothoracic Posterior Intermediate Anterior Commissure (MesoPIAC)
The MesoPIAC is an anterior commissure formed by the axons of hemilineages 7B and 8B as they cross the midline in the mesothoracic neuromere (Shepherd et al., 2016). This commissure was originally identi1ed by Power (1948) as the Commissure of the Mesothoracic Neuromere.
Synonyms:
MesoPIAC (Proposed by group),
Commissure of the Mesothoracic Neuromere (Power, 1948),
CMN (Power, 1948).
Mesothoracic Posterior Intermediate Posterior Commissure (MesoPIPC)
The MesoPIPC is a posterior commissure formed by the axons of hemilineages 5B, 6B and 12B as they cross the midline in the mesothoracic neuromere (Shepherd et al., 2016).
Synonyms:
MesoPIPC (Proposed by group).
Metathoracic Posterior Intermediate Anterior Commissure (MetaPIAC)
The MetaPIAC is an anterior commissure in the metathorax formed by the axons from hemilineages 7B and 8B as they cross the midline in the metathoracic neuromere. The commissure is posterior to the axons from hemilineage 2A (Shepherd et al., 2016).
This commissure was originally identi1ed by Power (1948) as the Haltere Commissure.
Synonyms:
MetaPIAC (Proposed by group),
Haltere Commissure (Power, 1948),
cHIN (Merritt and Murphey, 1992; Tyrer and Gregory, 1982; Pfluger et al., 1988),
ITD-HC (Boerner and Duch, 2010),
HC (Power, 1948).
Metathoracic Posterior Intermediate Posterior Commissure (MetaPIPC)
The MetaPIPC is a posterior commissure formed by the axons of hemilineages 5B, 6B and 12B as they cross the midline in the mesothoracic neuromere (Shepherd et al., 2016).
Synonyms:
MetaPIPC (Proposed by group).
Prothoracic Posterior Dorsal Commissure (ProPDC)
The ProPDC is a posterior commissure formed by the axons from hemilineage 6A as they cross the midline in the dorsal prothoracic neuromere (Shepherd et al., 2016).
Synonyms:
ProPDC (Proposed by group).
Mesothoracic Posterior Dorsal Commissure (MesoPDC)
The MesoPDC is a dorsal commissure formed by the axons of hemilineage 6A as they cross the midline in posterior mesothoracic neuromere (Shepherd et al., 2016). This commissure was originally identi1ed by Power (1948) as the Posterior Dorsal Mesothoracic Decussation.
Synonyms:
MesoPDC (Proposed by group),
Posterior Dorsal Mesothoracic Decussation (Power, 1948),
PDD (Power, 1948).
Metathoracic Posterior Dorsal Commissure (MetaPDC)
The MetaPDC is a dorsal commissure formed by the axons form hemilineage 6A as they cross the midline in the posterior metathoracic neuromere (Shepherd et al., 2016).
Synonyms:
MetaPDC (Proposed by group).
Commissure of the Fine Fibers of the Intermediate Tract of the Dorsal Cervical Fasciculus (CFF)
A chiasma formed by the inner tracts of the intermediate tract of the dorsal cervical fasciculus (ITD-CFF), as they cross the midline above the mesothoracic neuromere and terminate on the opposite side. The chiasma of 1ne 1bers is anterior to the haltere commissure (MetaPIAC).
Synonyms:
CFF (Power, 1948).
ITD-CFF BROAD (Boerner and Duch, 2010).
In addition to the commissures that can be related to the projections of speci1c hemilineages there are commissures that do not relate to speci1c clusters of central neurons.
Commissure of Prothoracic Neuromeres (CPN)
A transverse bundle of 1bers that runs across the left and right prothoracic neuromeres. The 1bers bow posteriorly and are located dorsal to the dorsal lateral tracts of the ventral cervical fasciculus (DLV). The arms of the bow are directed lateroanteriorly, and extend almost to the lateral borders of the neuromeres.
Synonyms:
CPN (Power, 1948),
prothoracic commissure Bacon and Strausfeld (1986).
Dorsal Accessory Commissure of the Mesothoracic Neuromeres (DAM)
A transverse thin bundle of 1bers that runs across the dorsoposterior region of the left and right mesothoracic neuromeres. It is located ventrally to the roots of the dorsal metathoracic (haltere) nerves, dorsal to the ventral ellipse, and ventroanterior to the haltere chiasma.
synonyms:
DAM (Power, 1948).
Described Tracts
Several tracts are described by Power (1948) and later clari1ed by Merritt and Murphey (1992). They were later put in a volumetric context by Boerner and Duch (2010), who kindly made their data available for inclusion in this project.
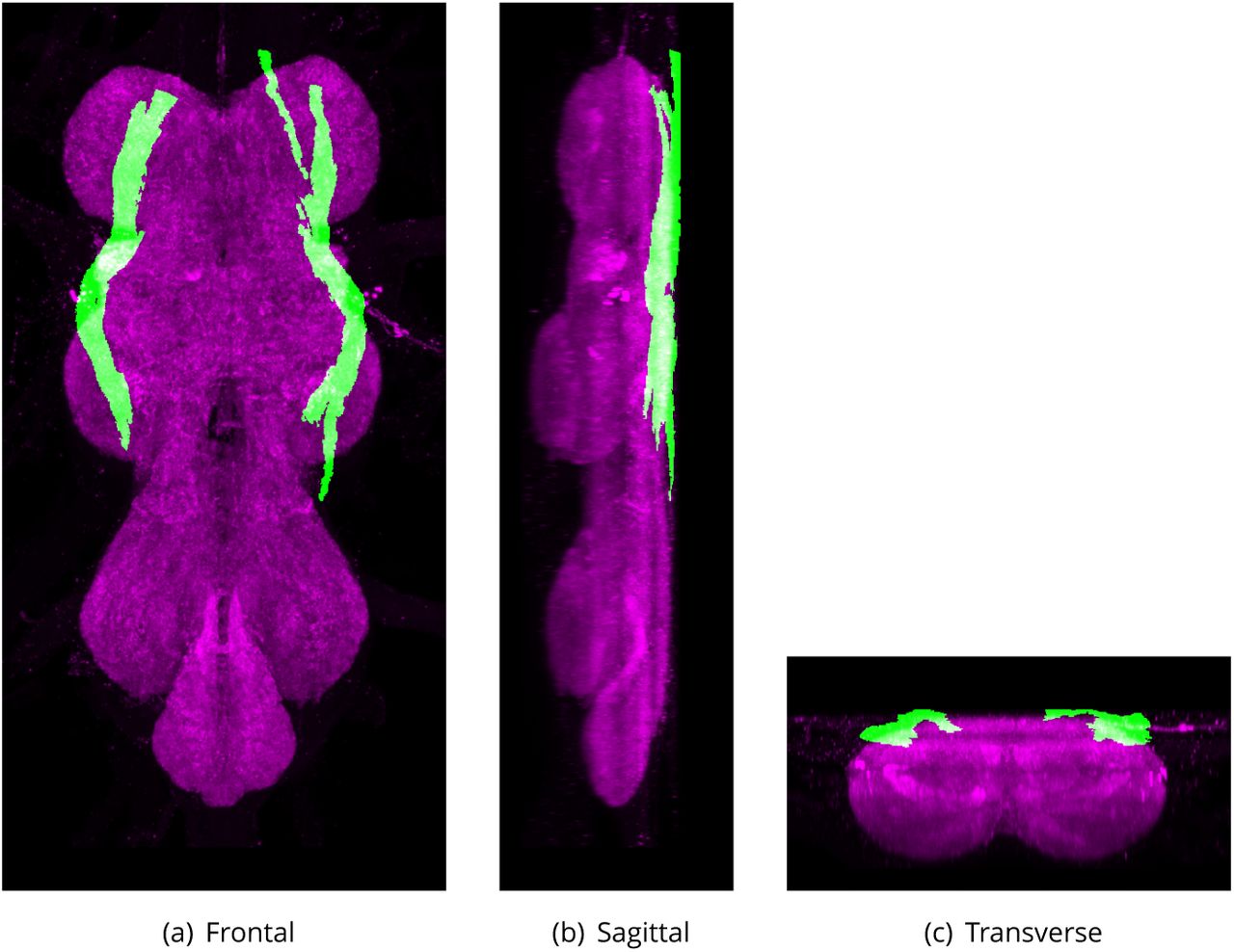
Dorsal Lateral Tract (DLT)
Defined region shown in Appendix 1 Figure 8
The DLT derives from the lateral bundles of the cervical connective. It has coarser 1bres than the other tract derived from the ventral bundles of the cervical connective (DLV, VLT and VTV). The tract projects posteriorly and super1cially at the dorsal lateral edge of the neuropil and terminates in the metathoracic neuromere.
Synonyms:
DLT (Power, 1948)
MTD (Merritt and Murphey, 1992),
Median tract of dorsal cervical fasciculus (Merritt and Murphey, 1992).

The proposed volume de1nition for the Dorsal Lateral Tract (DLT). Maximum projections from the labelled orthogonal plane with DLT shown in green and neuropil (Bruchpilot) shown in magenta. Regions were painted using ITK-SNAP by Robert Court based on data provided by Boerner and Duch (2010) as well as literature and group discussions to clarify boundaries.

The proposed volume de1nition for the Intermediate Tract of Dorsal Cervical Fasciculus (ITD). Maximum projections from the labelled orthogonal plane with ITD shown in green and neuropil (Bruchpilot) shown in magenta. Regions were painted using ITK-SNAP by Robert Court based on data provided by Boerner and Duch (2010) as well as literature and group discussions to clarify boundaries.
Intermediate Tract of Dorsal Cervical Fasciculus (ITD)
Defined region shown in Appendix 1 Figure 9
The intermediate tracts of the dorsal fasciculus derive from the dorsal 1bers in the connective but are much broader than the DLT and located more medial. The tract passes posteriorly, and tends to separate into three adjacent tracts. The inner subdivision (ITD-CFF) turns medially in the mesothoracic neuromere, crosses its homologue of the opposite side, and forms the chiasma of 1ne 1bers of the intermediate tracts of the dorsal cervical fasciculus (CFF). The 1bers of this chiasma pass dorsal to the median tracts of the dorsal fasciculus, and immediately over the very deep roots of the haltere nerves. Some of the 1bers (ITD-HC) bend anterolaterally, while others turn ventrally and straggle into the dorsolateral part of the mesothoracic neuromere where they are quickly lost. There seems to be no consistency in which of the two bundles of the chiasma crosses above the other; sometimes the right tract passes above the left, and in other specimens the left is dorsal to the right, but in all cases, each tract retains its unity. In other words, the component elements of the two do not mix by intermeshing. The ITD as described by Power (1948) includes a third bundle of 1bers (ITD-HT) that enter the VNS via the Dorsal Metathoracic Nerve (Haltere Nerve) and is the tract referred to as the Haltere Tract in Merritt and Murphey (1992). This haltere tract is the most lateral component of the intermediate tract and is composed of many large-diameter 1bres that can be traced as a bundle running into the cervical connective. It is composed of sensory afferent axons (Ghysen, 1980; Strausfeld and Seyan, 1985).
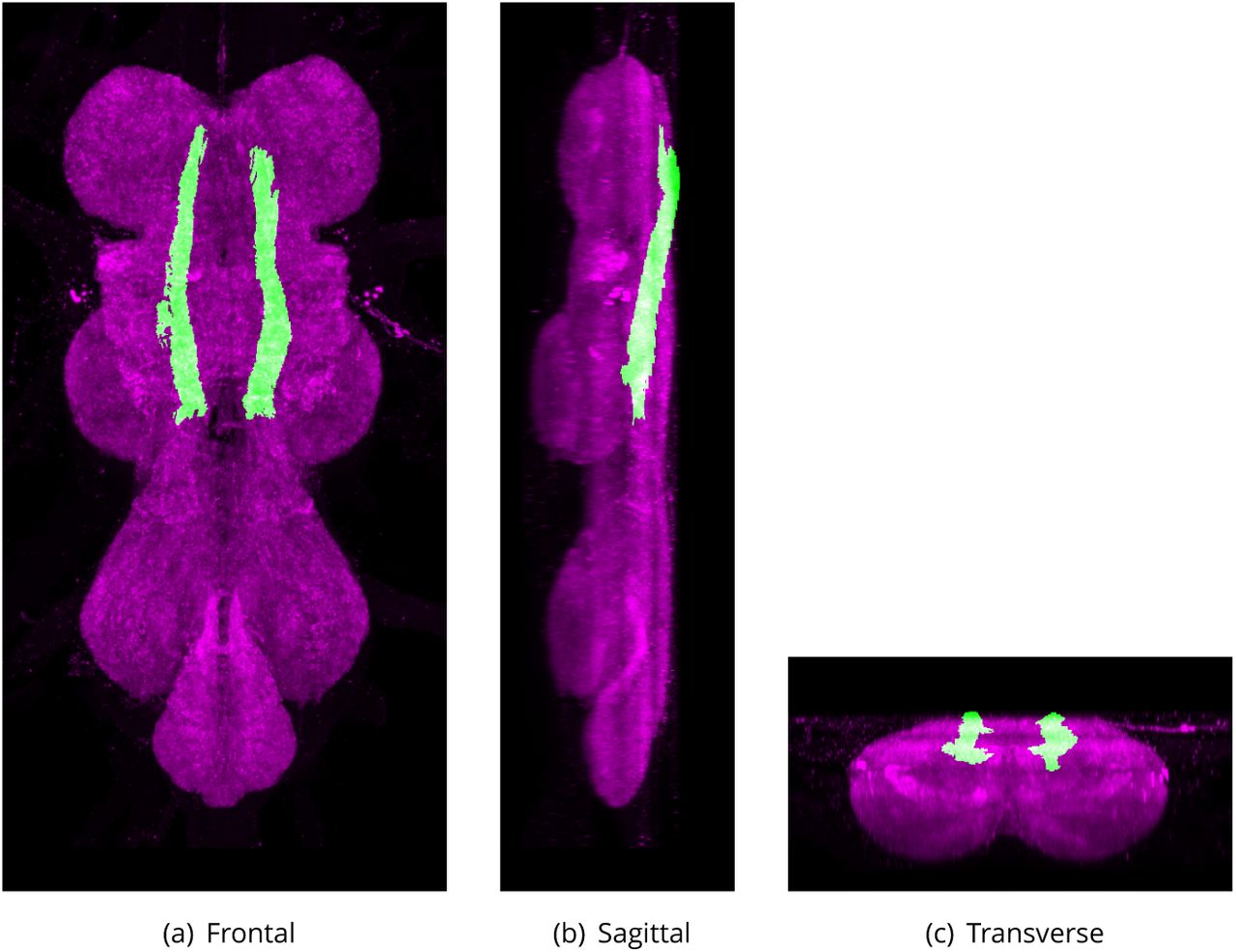
The proposed volume de1nition for the Inner Tracts of the Intermediate Tract of the Dorsal Cervical Fasciculus (ITD-CFF). Maximum projections from the labelled orthogonal plane with ITD-CFF shown in green and neuropil (Bruchpilot) shown in magenta. Regions were painted using ITK-SNAP by Robert Court based on data provided by Boerner and Duch (2010) as well as literature and group discussions to clarify boundaries.

The proposed volume de1nition for the Haltere Tract (HT). Maximum projections from the labelled orthogonal plane with HT shown in green and neuropil (Bruchpilot) shown in magenta. Regions were painted using ITK-SNAP by Robert Court based on data provided by Boerner and Duch (2010) as well as literature and group discussions to clarify boundaries.
Synonyms:
ITD (Power, 1948),
ITD-HC NARROW (Boerner and Duch, 2010),
ITD-CFF NARROW (Boerner and Duch, 2010),
ITD-HT NARROW (Boerner and Duch, 2010).
Inner Tracts of the Intermediate Tract of the Dorsal Cervical Fasciculus (ITD-CFF)
Defined region shown in Appendix 1 Figure 10
An inner subdivision of the Intermediate tract of dorsal cervical fasciculus (ITD) that turns medially in the mesothoracic neuromere, crosses its homologue of the opposite side, and forms the chiasma of 1ne 1bers of the intermediate tracts of the dorsal cervical fasciculus (CFF). The 1bers of this chiasma pass dorsal to the median tracts of the dorsal fasciculus, and immediately over the very deep roots of the haltere nerves.
Synonyms:
ITD-CFF (Boerner and Duch, 2010),
CFF NARROW (Power, 1948),
ITD BROAD (Power, 1948).
Haltere Tract (HT)
Defined region shown in Appendix 1 Figure 11
The haltere tract is formed by the sensory afferent axons (Ghysen, 1980; Strausfeld and Seyan, 1985) from the dorsal metathoracic nerve (Haltere Nerve) entering the metathoracic neuromere and extending anteriorly through the cervical connective (Power, 1948; Merritt and Murphey, 1992). The tract has small arborizations with some of the 1bres bending anterolaterally to become part of the haltere commissure (HC) in the metathoracic neuromere, while others turn ventrally and straggle into the dorsolateral part of the mesothoracic neuromere where they are quickly lost (Power, 1948). The haltere tract is the most lateral component of the intermediate tract (ITD) and is composed of many large-diameter 1bres that can be traced as a bundle running into the cervical connective (Power, 1948).
Synonyms:
ITD-HT (Boerner and Duch, 2010),
ITD BROAD (Power, 1948).
Dorsal Medial Tract (DMT)
Defined region shown in Appendix 1 Figure 12
A tract which derives from the dorsal bundles of the cervical connective. The tract extends posteriorly and bows laterally slightly, in the mesothoracic region, bending again medially, towards each other, in the narrow area between the mesoand metathoracic neuromeres and passing ventrally to the chiasma of 1ne 1bers. The dorsal medial tract turns lateroposteriorly at the level and dorsal to the haltere commissure, enters the metathoracic neuromere, forming collaterals and joins the metathoracic leg nerve (Power, 1948).
Synonyms:
MTD (Power, 1948),
median tracts of the dorsal cervical connective (Power, 1948).
Dorsal Lateral Tract of Ventral Cervical Fasciculus (DLV)
Defined region shown in Appendix 1 Figure 13
A tract which derives from the ventral bundles of the cervical connective. It has coarser 1bres than the ventral tract derived from the ventral bundles of the cervical connective (DLV, VLT and VTV). The tract spreads out as it extends posteriorly, passing dorsally to the commissure of the prothoracic neuromere, until it terminates in the mesothoracic neuromere. Two larger 1bers occupy the lateral and medial borders of the tract. The lateral 1bers send out a branch into the prothoracic neuromere, turn sharply posteriorly and enters the mesothoracic neuromere dorsally, producing a small median branch and joining the mesothoracic leg nerve. The medial 1ber extends posteriorly until the level at which the giant 1bers turn laterally, and turns slightly laterally into the mesothoracic neuromere. Ventral to these projections, the tract forms a 1brous loop (ventral ellipse) which runs from the prothoracic neuromere to the region between the mesoand metathoracic neuromeres. A right and left arm of 1bers turn lateroposteriorly from the posterior end of the ellipse and join the metathoracic leg nerve. Ventral to the ellipse, 1bers extend lateroanteriorly into the proand mesothoracic neuromeres.
Synonyms:
DLV (Power, 1948).
Ventral Lateral Tract (VLT)
Defined region shown in Appendix 1 Figure 14
A thin bundle of 1bers which derives from the ventral bundles of the cervical connective. It is lateral to the ventral ellipse. The ventral lateral tract extends from the prothoracic neuromere posteriorly into the anterolateral sides of the metathoracic neuromere. At the level of the anterior dorsal mesothoracic nerve, the 1bers bow sharply laterally but quickly turn medially and continue extending posteriorly, above the mesothoracic commissure (Power, 1948).
Synonyms:
ventral lateral tract of the ventral cervical fasciculus (Power, 1948),
VLT (Power, 1948).
Ventral Median Tract of Ventral Cervical Fasciculus (VTV)
Defined region shown in Appendix 1 Figure 15
A bundle of 1ne 1bers which derives from the ventral bundles of the cervical connective. They extend posteriorly along the medial plane until they bend dorsally and terminate in the ventroanterior region of the abdominal ganglion. A few 1bers branch out in each neuromere before the tract reaches the abdominal ganglion (Power, 1948).
The proposed volume de1nition for the Dorsal Medial Tract (DMT). Maximum projections from the labelled orthogonal plane with DMT shown in green and neuropil (Bruchpilot) shown in magenta. Regions were painted using ITK-SNAP by Robert Court based on data provided by Boerner and Duch (2010) as well as literature and group discussions to clarify boundaries.

The proposed volume de1nition for the Dorsal Lateral Tract of Ventral Cervical Fasciculus (DLV). Maximum projections from the labelled orthogonal plane with DLV shown in green and neuropil (Bruchpilot) shown in magenta. Regions were painted using ITK-SNAP by Robert Court based on data provided by Boerner and Duch (2010) as well as literature and group discussions to clarify boundaries.

The proposed volume de1nition for the Ventral Lateral Tract (VLT). Maximum projections from the labelled orthogonal plane with VLT shown in green and neuropil (Bruchpilot) shown in magenta. Regions were painted using ITK-SNAP by Robert Court based on data provided by Boerner and Duch (2010) as well as literature and group discussions to clarify boundaries.

The proposed volume de1nition for the Ventral Median Tract of Ventral Cervical Fasciculus (VTV). Maximum projections from the labelled orthogonal plane with VTV shown in green and neuropil (Bruchpilot) shown in magenta. Regions were painted using ITK-SNAP by Robert Court based on data provided by Boerner and Duch (2010) as well as literature and group discussions to clarify boundaries.
Synonyms:
VTV (Power, 1948).
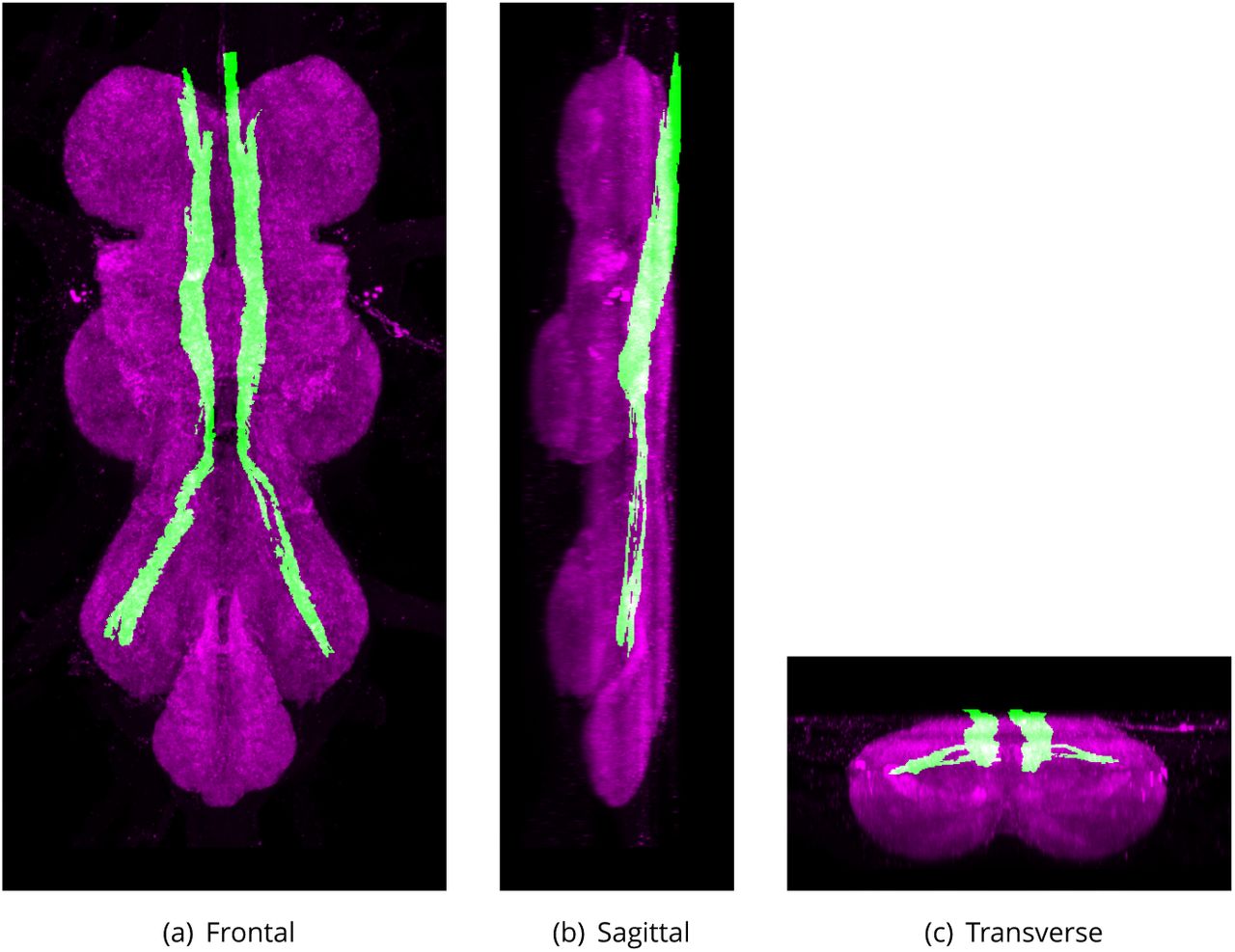
Median Dorsal Abdominal Tract (MDT)
Defined region shown in Appendix 1 Figure 16
The dorsal-most tract, close to midline (Merritt and Murphey, 1992) running along the tectulum (Boerner and Duch, 2010). The median-most pair of three small dorsal tracts which connects the thoracic and abdominal neuromeres. It runs from the dorsal region of the tectulum along through the thoracico-abdominal mass, immediately under the giant 1bers. It continues posteriorly after the giant 1bers have turned laterally, passing above the haltere chiasma into the abdominal neuromeres (Power, 1948).
Synonyms:
MDT (Merritt and Murphey, 1992),
MDA (Power, 1948),
median dorsal tract (Merritt and Murphey, 1992).
Intermediate Dorsal Abdominal Tract (IDT)
The intermediate pair of three small dorsal tracts which connects the thoracic and abdominal neuromeres dorsal of the metathoracic neuromeres (Power, 1948).
Synonyms:
IDT (Proposed by group).
Lateral Dorsal Abdominal Tract (LDT)
The lateral pair of three small dorsal tracts which connects the thoracic and abdominal neuromeres dorsal of the metathoracic neuromeres (Power, 1948).
Synonyms:
LDT (Proposed by group).
Ventral Cervical Fasciculus (VCF)
Ventral fascicle of the dorsal tectulum. It enters the thoracico-abdominal mass along with the dorsal bundles and takes a more ventral course. There are two of these fascicles. Each fascicle passes posteriorly between the prothoracic and at an intermediate position in these neuromeres, dividing into the dorsal lateral (DLV) and ventral median (VTV) tracts (Power, 1948).
Synonyms:
VCF (Power, 1948).
Dorsal Cervical Fasciculus (DCF)
Dorsal fascicle of the dorsal tectulum. It occupies the uppermost region within the anterior part of the dorsal tectulum. The 1bers slope gently ventrally, so that by the time they have passed over the anterior tips of the mesothoracic neuromeres, they have descended to a relatively lower position, below the mesothoracic decussations. There are two of these fascicles. Near the anterior end of the VNS each of the dorsal fasciculus separates forming three poorly Defined longitudinal tracts: the dorsal lateral (DLT), intermediate (ITD) and dorsal median tract (DMT) (Power, 1948).
Synonyms:
DCF (Power, 1948).
Ventral Ellipse (VE)
Flattened annulus of 1bers which lies in a frontal plane and extends from the middle of the prothoracic neuromeres posteriorly to the isthmus between the mesoand metathoracic neuromeres. It lies immediately below the dorsal decussations of the mesothoracic and metathoracic neuromeres. A right and left arm of 1bers extends lateroposteriorly from the posterior end of the ellipse into the respective metathoracic neuromeres, and within them, joins the bundle which runs out into the third leg nerve. At the anterior end, the ventral ellipse incorporates the dorsolateral tracts of the ventral fasciculi (DLV) (Power, 1948).
Synonyms:
VE (Power, 1948).
The peripheral nerves
Most nerves have been historically well Defined by Power (1948) in which case the de1nitions are maintained, however, with some of the abdominal nerves we decided to utilise later terms (Shepherd and Smith, 1996) naming each nerve according to its neuromere of origin to give them a more consistent naming scheme than the original.
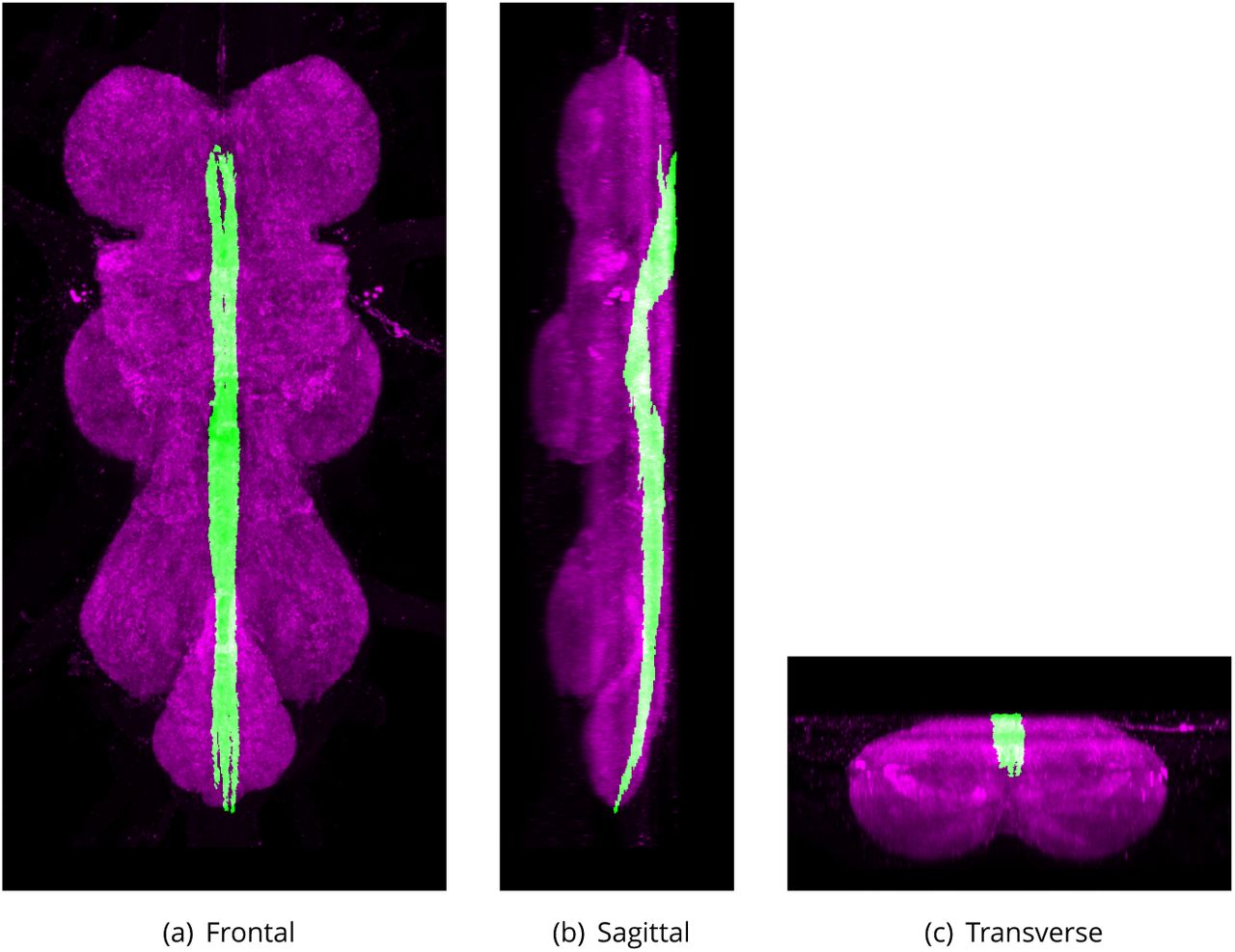
Cervical Connective (CvC)
Defined region shown in Appendix 1 Figure 17
Major axon tract connecting the posterior-most subesophageal ganglion to the VNS in the adult central nervous system (Power, 1948).
Synonyms:
CvC (Power, 1948),
cephalo-thoracic cord (Bodenstein, 1950),
cephalo-thoracic nerve strand,
CV (Ito et al., 2014),
CvCon (Miller and Demerec, 1950).
Cervical Nerve (CvN)
A bilaterally paired nerve that connects laterally to the cervical connective, immediately posterior to where it enters the thorax. Each cervical nerve extends laterally, branching and innervating horizontal muscles of the anterior thorax (Power, 1948).
Synonyms:
CvN (Power, 1948),
nerve to crop (Miller and Demerec, 1950),
CvNv (Miller and Demerec, 1950).
Dorsal nerves directly innervating the neuropil of the prothoracic neuromere
The dorsal prothoracic nerve, prosternal nerve, prothoracic chordotonal nerve, prothoracic accessory nerve and the ventral prothoracic nerve emerge closely adjacent to each other in the anteriolateral corner of the prothoracic neuromere. Their proximity is such that in some preparations they appear to share a common exit point.
Dorsal Prothoracic Nerve (DProN)
Defined region shown in Appendix 1 Figure 18
A nerve that projects latero-anteriorly from the antero-lateral corner of the ventral nervous system (VNS) (prothoracic neuromere). It splits into 4 or more branches before innervating various muscles (Power, 1948).
Synonyms:
DProN (Proposed by group),
ADN (Power, 1948),
anterior dorsal nerve (Power, 1948),
DPN (Merritt and Murphey, 1992),
1rst dorsal nerve (Miller and Demerec, 1950),
DNv1 (Miller and Demerec, 1950).
The proposed volume de1nition for the Median Dorsal Abdominal Tract (MDT). Maximum projections from the labelled orthogonal plane with MDT shown in green and neuropil (Bruchpilot) shown in magenta. Regions were painted using ITK-SNAP by Robert Court based on data provided by Boerner and Duch (2010) as well as literature and group discussions to clarify boundaries.

The proposed volume de1nition for the Cervical Connective (CvC). Maximum projections from the labelled orthogonal plane with CvC shown in green and neuropil (Bruchpilot) shown in magenta. Regions were painted using ITK-SNAP by Robert Court based on literature and group discussions to clarify boundaries.
Prosternal Nerve (PrN)
A slender nerve that projects anteriorly from the ventral nervous system (VNS), medial to the base of the dorsal prothoracic nerve to the prosternal sense organ (Power, 1948).
Synonyms:
PrN (Power, 1948),
Prothoracic Chordotonal Nerve (ProCN)
Very short and thick nerve that arises in the prothoracic neuromere, immediately below the anterior dorsal and prosternal nerves, and connects to each prothoracic chordotonal sense organ (Power, 1948).
Synonyms:
ProCN (Proposed by group),
CN (Power, 1948).
Prothoracic Accessory Nerve (ProAN)
Defined region shown in Appendix 1 Figure 19
A mixed motor-sensory nerve that connects to the prothoracic neuromere, slightly posterior and ventral to the anterior prothoracic chordotonal organ and slightly dorsal to the root of the ventral prothoracic nerves. It extends laterally and dorsally almost to the lateral body wall before branching to innervate muscles (Power, 1948).
Synonyms:
ProAN (Proposed by group),
accessory prothoracic nerve,
PAN (Power, 1948),
1rst accessory nerve (Miller and Demerec, 1950),
AcNv1 (Miller and Demerec, 1950).
Ventral Prothoracic Nerve (VProN)
Defined region shown in Appendix 1 Figure 20
A mixed motor-sensory nerve that carries axons from two clusters of microchaetae on the prothoracic coxa and to motor neuron 1bers from lateral anterior muscles. It connects to the prothoracic neuromere just dorsal to the root of the prothoracic leg nerve and branches about halfway along its length into a motor branch that stays within the body and a sensory branch that projects to the leg (Power, 1948).
Synonyms:
VProN (Proposed by group),
VPN (Power, 1948),
prosternal sense organ (Miller and Demerec, 1950),
PSO (Miller and Demerec, 1950).
Prothoracic Leg Nerve (ProLN)
Defined region shown in Appendix 1 Figure 21
A nerve that carries a mix of motor and sensory axons from the prothoracic leg to the adult prothoracic neuromere. Each nerve extends laterally and slightly anteriorly from the ventral anterior region of the prothoracic neuromere (Power, 1948).

The proposed volume de1nition for the Dorsal Prothoracic Nerve (DProN). Maximum projections from the labelled orthogonal plane with DProN shown in green and neuropil (Bruchpilot) shown in magenta. Regions were painted using ITK-SNAP by Robert Court based on literature and group discussions to clarify boundaries.

The proposed volume de1nition for the Prothoracic Accessory Nerve (ProAN). Maximum projections from the labelled orthogonal plane with ProAN shown in green and neuropil (Bruchpilot) shown in magenta. Regions were painted using ITK-SNAP by Robert Court based on literature and group discussions to clarify boundaries.

The proposed volume de1nition for the Ventral Prothoracic Nerve (VProN). Maximum projections from the labelled orthogonal plane with VProN shown in green and neuropil (Bruchpilot) shown in magenta. Regions were painted using ITK-SNAP by Robert Court based on literature and group discussions to clarify boundaries.

The proposed volume de1nition for the Prothoracic Leg Nerve (ProLN). Maximum projections from the labelled orthogonal plane with ProLN shown in green and neuropil (Bruchpilot) shown in magenta. Regions were painted using ITK-SNAP by Robert Court based on literature and group discussions to clarify boundaries.
Synonyms:
ProLN (Proposed by group),
PLN (Power, 1948),
T1LN (Merritt and Murphey, 1992),
1rst ventral nerve (Miller and Demerec, 1950),
VNv1 (Miller and Demerec, 1950).
Anterior Dorsal Mesothoracic Nerve (ADMN)
Defined region shown in Appendix 1 Figure 22
A mixed sensory-motor nerve that is the thicker of the two dorsal nerves of the mesothorax. The nerve enters the mesothoracic neuromere slightly anterior and dorsal to the smaller PDMN. The ADMN projects anteriorly and dorsally (Power, 1948).
Synonyms:
ADMN (Power, 1948),
second dorsal nerve BROAD (Miller and Demerec, 1950),
DNv2 BROAD (Miller and Demerec, 1950),
wing nerve BROAD (Merritt and Murphey, 1992).
Wing Nerve
A nerve that carries sensory 1bers from the sense organs of the wing, eventually joining the anterior dorsal mesothoracic nerve (ADMN) (Merritt and Murphey, 1992).
Posterior Dorsal Mesothoracic Nerve (PDMN)
Defined region shown in Appendix 1 Figure 23
A nerve that arises from the ventral nervous system (VNS), just posterior to the root of the anterior dorsal mesothoracic nerve. It projects posterolaterally before branching, with one branch innervating the tergal depressor of the trochanter (jump muscle), while the other branch forms further, terminal branches that innervate targets including the dorsal medial muscle (dorsal longitudinal muscle) (Power, 1948).
Synonyms:
PDMN (Merritt and Murphey, 1992),
PDM (Power, 1948)
second dorsal nerve BROAD (Miller and Demerec, 1950),
DNv2 BROAD (Miller and Demerec, 1950).
Mesothoracic Accessory Nerve (MesoAN)
Defined region shown in Appendix 1 Figure 24
A nerve that arises from the lateroposterior side of the mesothoracic neuromere at a point in anterior to the root of the haltere nerve. It extends posteriorly and slightly dorsally and laterally, around the anterior wings of the mesofurca, before branching. One branch innervates the furcoentopleural muscles (muscles 59 and 60) and the other innervates laterally placed muscles, anterior to the halteres (Power, 1948).
Synonyms:
MesoAN (Proposed by group),
MAC (Power, 1948),
accessory mesothoracic nerve (Power, 1948)
second accessory nerve (Miller and Demerec, 1950),
AcNv2 (Miller and Demerec, 1950).
Mesothoracic Leg Nerve (MesoLN)
Defined region shown in Appendix 1 Figure 25
A mixed motor-sensory nerve that arises ventrally from the mesothoracic neuromere. It splits at its base, with a small number of axons innervating a ventral muscle that is posterior-lateral to the tergal depressor of the trochanter (jump muscle) and the rest projecting into the mesothoracic leg (Power, 1948).

The proposed volume de1nition for the Anterior Dorsal Mesothoracic Nerve (ADMN). Maximum projections from the labelled orthogonal plane with ADMN shown in green and neuropil (Bruchpilot) shown in magenta. Regions were painted using ITK-SNAP by Robert Court based on literature and group discussions to clarify boundaries.
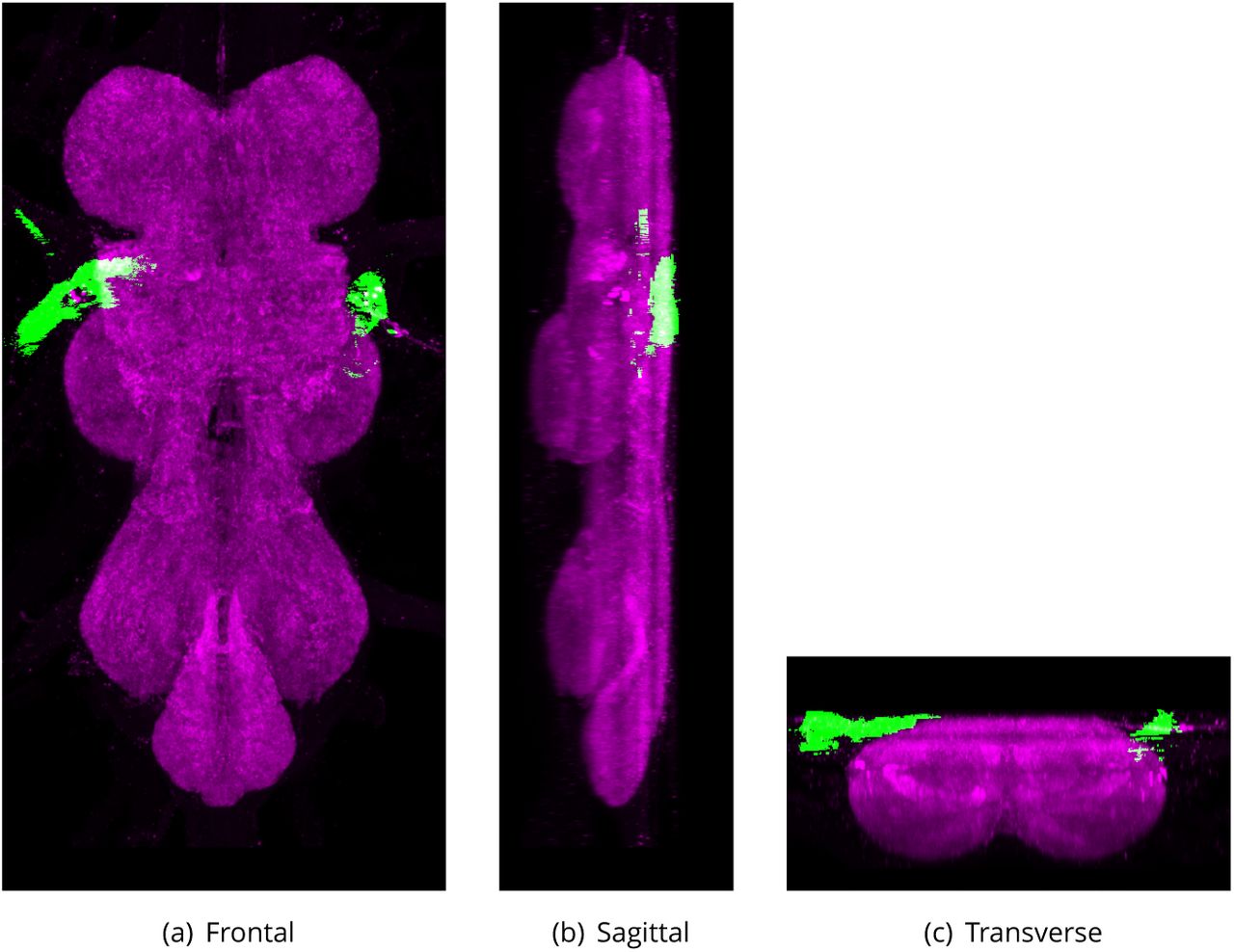
The proposed volume de1nition for the Posterior Dorsal Mesothoracic Nerve (PDMN). Maximum projections from the labelled orthogonal plane with PDMN shown in green and neuropil (Bruchpilot) shown in magenta. Regions were painted using ITK-SNAP by Robert Court based on literature and group discussions to clarify boundaries.

The proposed volume de1nition for the Mesothoracic Accessory Nerve (MesoAN). Maximum projections from the labelled orthogonal plane with MesoAN shown in green and neuropil (Bruchpilot) shown in magenta. Regions were painted using ITK-SNAP by Robert Court based on literature and group discussions to clarify boundaries.

The proposed volume de1nition for the Mesothoracic Leg Nerve (MesoLN). Maximum projections from the labelled orthogonal plane with MesoLN shown in green and neuropil (Bruchpilot) shown in magenta. Regions were painted using ITK-SNAP by Robert Court based on literature and group discussions to clarify boundaries.
Synonyms:
MesoLN (Proposed by group),
T2LN (Merritt and Murphey, 1992),
second ventral nerve (Miller and Demerec, 1950),
VNv2 (Miller and Demerec, 1950),
ventral mesothoracic nerve.
Dorsal Metathoracic Nerve (DMetaN)
Defined region shown in Appendix 1 Figure 26
A thick nerve that primarily carries sensory axons from the haltere to the metathoracic neuromere. It extends anteriorly and somewhat medially to terminate in the center of the metathoracic neuromere. Its 1bers extend anteriorly, contributing to the tectulum (Power, 1948).
Synonyms:
DMetaN (Proposed by group),
haltere nerve (Power, 1948),
HN (Power, 1948),
third dorsal nerve (Miller and Demerec, 1950),
DNv3 (Miller and Demerec, 1950).
Metathoracic Leg Nerve (MetaLN)
Defined region shown in Appendix 1 Figure 27
A large sensory motor nerve that originates in the ventral metathoracic neuromere and innervates the metathoracic leg (Power, 1948).
Synonyms:
MetaLN (Proposed by group),
MLN (Power, 1948),
T3LN (Merritt and Murphey, 1992),
third ventral nerve (Miller and Demerec, 1950),
VNv3 (Miller and Demerec, 1950),
ventral metathoracic nerve.
The abdominal nerves
Emerging from the abdominal neuromeres are four paired nerves that extend posteriorly and laterally into the abdomen.
First Abdominal Nerve (AbN1)
Defined region shown in Appendix 1 Figure 28
The 1rst abdominal nerve apparently emerges from the metathoracic neuromere but contains axons that originate/terminate in the 1rst abdominal neuromere. The nerve exits laterally just dorsal to the exit of the metathoracic leg nerve (Shepherd and Smith, 1996).
Synonyms:
AbN1 (Shepherd and Smith, 1996),
accessory metathoracic nerve,
MA (Power, 1948),
metathoracic accessory nerve (Power, 1948),
nerve of the 1rst abdominal segment (Miller and Demerec, 1950),
Ab1Nv (Miller and Demerec, 1950).

The proposed volume de1nition for the Dorsal Metathoracic Nerve (DMetaN). Maximum projections from the labelled orthogonal plane with DMetaN shown in green and neuropil (Bruchpilot) shown in magenta. Regions were painted using ITK-SNAP by Robert Court based on literature and group discussions to clarify boundaries.

The proposed volume de1nition for the Metathoracic Leg Nerve (MetaLN). Maximum projections from the labelled orthogonal plane with MetaLN shown in green and neuropil (Bruchpilot) shown in magenta. Regions were painted using ITK-SNAP by Robert Court based on literature and group discussions to clarify boundaries.

The proposed volume de1nition for the First Abdominal Nerve (AbN1). Maximum projections from the labelled orthogonal plane with AbN1 shown in green and neuropil (Bruchpilot) shown in magenta. Regions were painted using ITK-SNAP by Robert Court based on literature and group discussions to clarify boundaries.

The proposed volume de1nition for the Second Abdominal Nerve (AbN2). Maximum projections from the labelled orthogonal plane with AbN2 shown in green and neuropil (Bruchpilot) shown in magenta. Regions were painted using ITK-SNAP by Robert Court based on literature and group discussions to clarify boundaries.
Second Abdominal Nerve (AbN2)
Defined region shown in Appendix 1 Figure 29
A nerve that apparently emerges in the dorsal-most region of the metathoracic neuromere but contains axons that originate/terminate in the second abdominal neuromere. The nerve projects postero-laterally to the most posterior-lateral corner of the thorax, where it innervates transverse tubular muscles. This nerve also contains the afferent 1bers from a multiscolophorous organ located on the ventral surface of the second abdominal segment (Shepherd and Smith, 1996).
Synonyms:
AbN2 (Shepherd and Smith, 1996),
extra metathoracic nerve (Power, 1948),
EMN (Power, 1948),
nerve of the second abdominal segment (Miller and Demerec, 1950),
Ab2Nv (Miller and Demerec, 1950).
Third Abdominal Nerve (AbN3)
Defined region shown in Appendix 1 Figure 30
Lateral-most of the two bilaterally paired nerves connected to the abdominal neuropil. It is thin, containing only 1ne 1bers (Shepherd and Smith, 1996).
Synonyms:
AbN3 (Shepherd and Smith, 1996),
1rst lateral abdominal nerve (Power, 1948),
FLA (Power, 1948),
nerve of the third abdominal segment (Miller and Demerec, 1950),
Ab3Nv (Miller and Demerec, 1950).
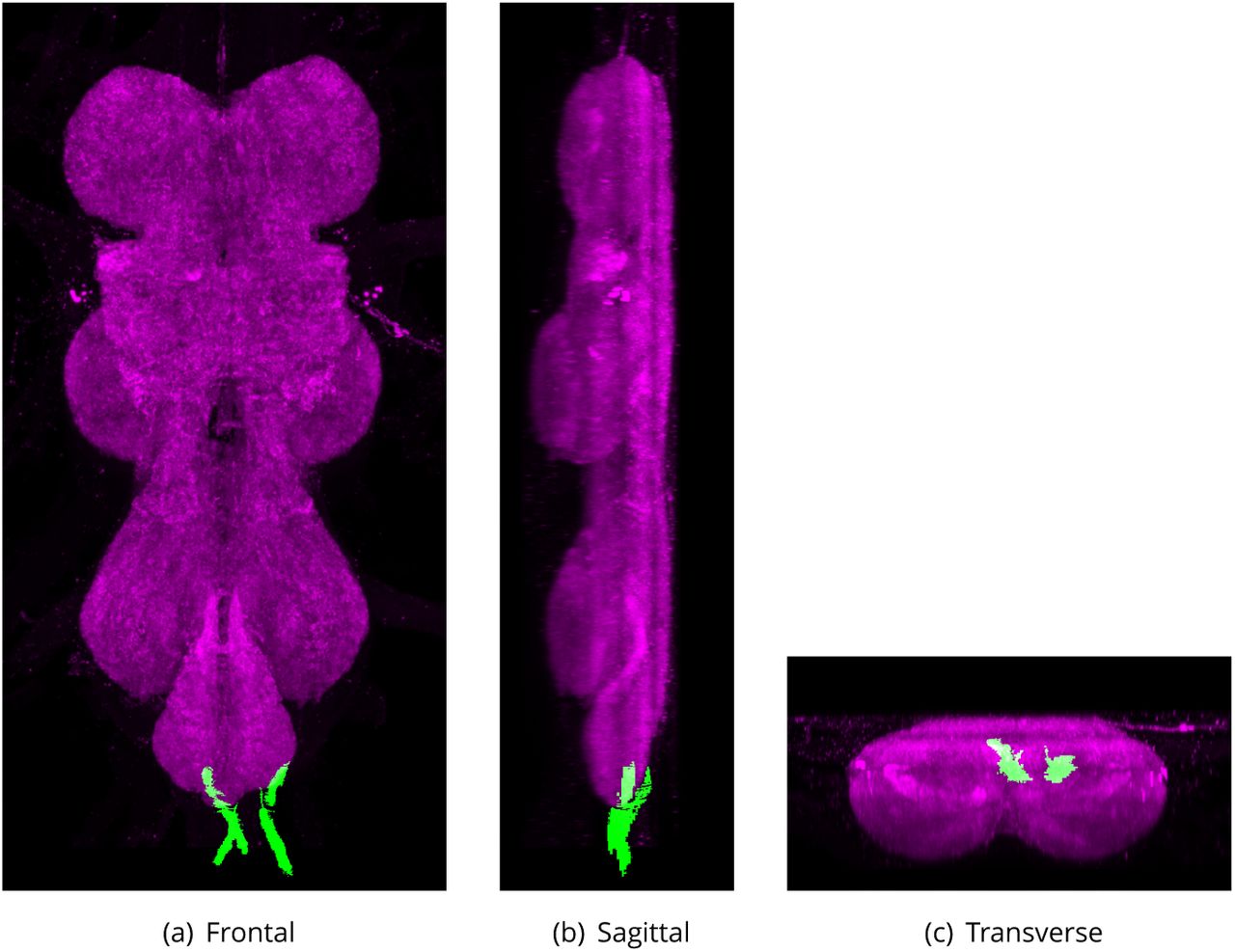
Fourth Abdominal Nerve (AbN4)
Defined region shown in Appendix 1 Figure 31
Medial-most of the two bilaterally paired nerves connected to the abdominal neuropil. It is thin, containing only 1ne 1bers (Shepherd and Smith, 1996).
Synonyms:
AbN4 (Shepherd and Smith, 1996),
second lateral abdominal nerve (Power, 1948),
SLA (Power, 1948),
nerve of the fourth abdominal segment (Miller and Demerec, 1950),
Ab4Nv (Miller and Demerec, 1950).
Abdominal Nerve Trunk (AbNT)
Defined region shown in Appendix 1 Figure 32
A fused terminal nerve that projects posteriorly along the midline from the posterior of the abdominal neuropil (Shepherd and Smith, 1996).
Synonyms:
AbNT (Shepherd and Smith, 1996),
abdominal median nerve trunk,
median nerve trunk (Middleton et al., 2006),
AbNvTr (Middleton et al., 2006),
MAN (Power, 1948),
median abdominal nerve (Power, 1948),
terminal abdominal nerves (Miller and Demerec, 1950),
AbTNv (Miller and Demerec, 1950).

The proposed volume de1nition for the Third Abdominal Nerve (AbN3). Maximum projections from the labelled orthogonal plane with AbN3 shown in green and neuropil (Bruchpilot) shown in magenta. Regions were painted using ITK-SNAP by Robert Court based on literature and group discussions to clarify boundaries.
The proposed volume de1nition for the Fourth Abdominal Nerve (AbN4). Maximum projections from the labelled orthogonal plane with AbN4 shown in green and neuropil (Bruchpilot) shown in magenta. Regions were painted using ITK-SNAP by Robert Court based on literature and group discussions to clarify boundaries.
Individual Neurons
Whilst we did not want to expand our remit to cover individual neurons in this 1rst consolidation we thought it would be a good idea to include an example to demonstrate how easily this could be done.
Giant Fiber Neuron (GF)
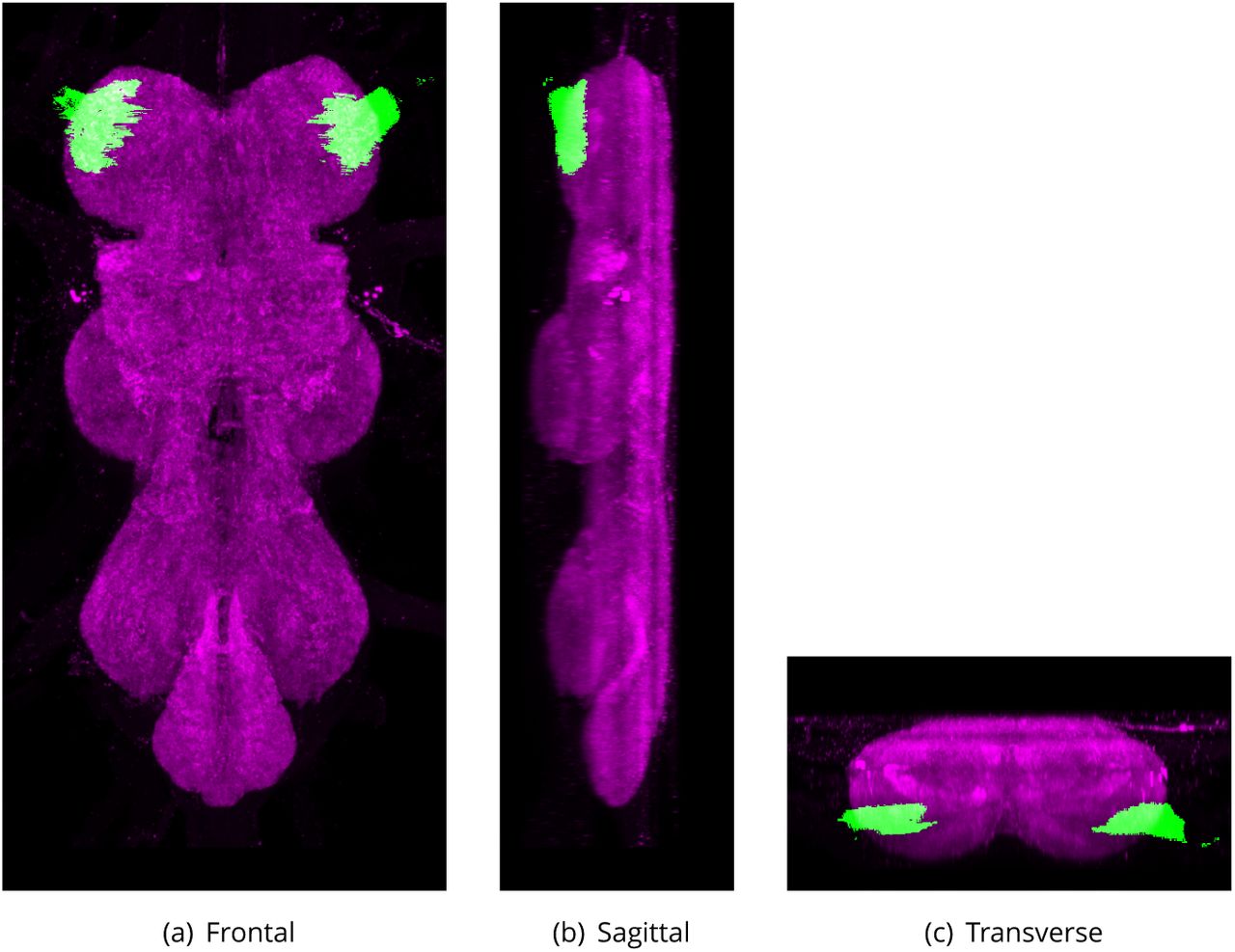
A large, descending neuron that controls jump escape behaviour (Trimarchi and Schneiderman, 1993). Each adult has a bilaterally symmetric pair of these neurons, each with a large cell body at the posterior of the protocerebrum and a long primary neurite that branches to forms an extensive dendritic tree in the brain and an axon that projects, via the cervical connective, to the mesothorax. The axon enters the ventral nervous system (VNS) on the dorsal region and slopes ventrally, reaching the ventral mesothoracic neuromere. Targets of dendritic arborization include zone A of the antennal mechanosensory and motor center (AMMC) (Kamikouchi et al., 2009) and the inferior ventro-lateral protocerebrum. In the mesothorax, the axon forms electrical synapses with tergotrochanteral muscle motor neuron (TTMn) and peripherally synapsing interneurons (PSI)) of the mesothorax that in turn synapse to the motor neurons of the dorso-longitudinal 2ight muscles.
Synonyms:
GF (Power, 1948),
giant 1ber (Power, 1948),
GFN (Kamikouchi et al., 2009),
giant 1bers.

The proposed volume de1nition for the Abdominal Nerve Trunk (AbNT). Maximum projections from the labelled orthogonal plane with AbNT shown in green and neuropil (Bruchpilot) shown in magenta. Regions were painted using ITK-SNAP by Robert Court based on literature and group discussions to clarify boundaries.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}