

Abstract
The ability to powered flight facilitated a great evolutionary success of insects and allowed them to occupy various ecological niches. In addition to primary tasks, wings are often involved in various premating behaviors, such as courtship songs and initiation of mating in flight. These specific implications require certain wing morphology, size, and shape. Although wing properties have been extensively studied in Drosophila, a comprehensive understanding of sexual shape dimorphisms and developmental plasticity in wing morphology is missing for other Diptera. To acquire this knowledge, we applied geometric morphometrics and analyzed wing shape in three dipteran species (Drosophila, Ceratitis, and Musca) raised in different environmental conditions. We extensively studied sexual dimorphism and impact of sex and environment on the adult wing morphology. We present allometric and non-allometric shape differences between males and females and show that wing shape is influenced by rearing conditions in a sex dependent manner. We determine common trends in shape alterations and show that the anterior and posterior crossveins are likely to be plastic regions changing substantially at different environmental conditions. We discuss our data in the light of vein development and hypothesize that the observed shape differences might recapitulate different mating behaviors and flight capabilities.
Introduction
Insects represent the only group of arthropods that developed the ability of a powered flight. This adaptation allowed them to occupy various ecological niches including air and led to a high morphological variation and great ecological success of the entire class. Flying helps insects to surmount long distances in a relatively short time, facilitating basic tasks such as finding mating partners and food resources. In modern insects, wings acquired special significance in other essential processes, e.g. mating and defense. For instance, certain species use their wings to perform courtship songs, which reflect size and vigor of males and help females to choose the right mating partner1,2. Some insects mate in the air while flying3; others initiate the mating process in flight but always land prior to copulation4. These different behaviors together with the intersexual food competition and various reproductive roles cause a constant selective pressure and result in different kinds of sexual dimorphisms, including variation in size and shape of insect wings5,6.
It has been shown that both genetic background and environmental cues contribute to the size variation of distinct body parts, and wings in particularly, across individuals of the same species7–13. Variation in growth rate leads to sexual size dimorphism (SSD), which can be either male- or female-biased, depending on whether males or females are larger14–16. The key component of the SSD is genetically defined in most insects, but the final body and organ size is determined by environmental changes that occur during development, i.e. developmental plasticity17. Moreover, in response to different rearing conditions, organ size may alter in a sex specific manner13,18. For instance, in female-biased systems, females overgrow males under certain environmental conditions. It results in disproportionate growth of certain body parts, e.g. wings, and the whole animal grow to a larger size19,20. In this case, wing size changes are likely to be accompanied by shape changes to assure that the wing remains functional. And indeed, it has been shown that in Drosophila melanogaster size and shape of wings are regulated by similar processes during patterning and differentiation of wing imaginal discs at larval and pupal stages21–23. Due to this tight morphological and developmental coupling, wing size and shape have been considered together for a long time24, and research on variation in shape was inseparably linked to variation in size. At present, however, development of advanced mathematical methods and geometric morphometrics approaches made it possible to disentangle these two parameters and analyze size and shape independently25–29.
Various quantitative techniques were used in order to describe phenotypic variation and possible influence of rearing conditions on wing shape in Drosophila and other insects30–33. Although we have an advanced understanding of sexual shape dimorphism (SShD) in insect wings in general5,6,14,24, sexual specificity of the wing shape changes is still not clear. Thus, we do not know whether shape of male and female wings respond differently to environmental cues and, if yes, to what extent.
We have recently shown that the three dipteran species Drosophila melanogaster, Ceratitis capitata, and Musca domestica exhibit a clear wing SSD and that the response of wing size to different rearing conditions is sex dependent13. Therefore, these three species represent an excellent model to test whether wing shape changes in a similar sex dependent manner. In this study, we applied geometric morphometrics to compare and comprehensively describe variation in wing morphology between Drosophila, Ceratitis, and Musca. Analysis of the total sexual shape dimorphism (SShD) and its non-allometric component revealed that SShD is not uniform across species and that the reaction of wing shape to different rearing conditions varies between sexes. We identified two most variable regions, the radio-medial (r-m) and basal-medial-cubital (bm-cu) crossveins, that changed similarly among the three species in response to various larval densities and temperature regimes. This finding suggests that these regions may represent developmentally less robust wing compartments. We discuss our findings in the light of different mating behaviors in Drosophila and Ceratitis, which produce courtship songs2,34, and Musca, which initiates mating by a strike prior to copulation4.
Materials and Methods
Fly species
For this study, all experiments were performed using three different fly species (Table 1). We used the highly inbred laboratory strain Drosophila melanogaster w1118, which was kept at 18°C on standard food (400 g of malt extract, 400 g of corn flour, 50 g of soy flour, 110 g of sugar beet syrup, 51 g of agar, 90 g of yeast extract, 31.5 ml of propionic acid and 7.5 g of Nipagin dissolved in 40 ml of Ethanol, water up to 5 l). The other two flies were Musca domestica wild type ITA1 collected in Italy, Altavilla Silentia in 2013 (Y. Wu and L. Beukeboom, GELIFES, The Netherlands) and Ceratitis capitata wild type Egypt II (IAEA). The Musca strain was reared at room temperature (RT) (22±2°C) on food composed by 500 g of wheat bran, 75 g of wheat flour, 60 g of milk powder, 25 g of yeast extract, 872 ml of water and 18.85 ml of Nipagin (2.86 g of Nipagin in 10 ml of Ethanol). Adult Musca flies were provided with sugar water. Ceratitis were kept at 28°C, 55 ± 5% RH on a diet composed by 52.5 g of yeast extract, 52.5 g of carrot powder, 2 g of Sodium benzoate, 1.75 g of agar, 2.25 ml of 32% HCl, 5 ml of Nipagin (2.86 g of Nipagin in 10 ml of Ethanol), water up to 500 ml for larvae. For adult flies, we used a 1:3 mixture of sugar and yeast extract.
WCS is shown for landmarks 1 to 11 for each species.
Treatment of experimental groups
To generate a range of sizes for each species, we applied two environmental factors known to influence the overall body size – temperature and density. Prior to the experiment, Drosophila flies were placed at 25°C for two days. On the third day, flies were moved from vials into egg-collection chambers and provided with apple-agar plates. After several hours, we started egg collection by removing apple-agar plates with laid eggs once per hour. Collected plates were kept at 25°C for 24 h to allow embryonic development to complete. Freshly hatched first-instar larvae were transferred into 50 ml vials with 15 ml of fly food. Three vials containing 25 larvae each (low density) and three vials with 300 larvae each (high density) were moved to 18°C; the second set of six vials with the same densities was left at 25°C.
Ceratitis flies were kept at 28°C and allowed to lay eggs through a net into water. Every hour, eggs were collected and transferred on the larval food. After 22 h, first-instar larvae were transferred into small Petri dishes (diameter 55 mm) with 15 ml of the larval food in three densities: 25 (low density), 100 (middle density) or 300 (high density) larvae per plate. Two plates of each density were moved to 18°C. The second set of six plates was left at 28°C for further development.
Eggs of Musca were collected for 24 h in the wet larval food at RT. Next day, all hatched larvae were removed from food, and only larvae hatched within the next hour were transferred into 50 ml vials with 5 g of food. Collection of larvae was repeated several times to obtain two experimental sets with three replicates of three experimental densities 10 (low density), 20 (middle density) or 40 (high density) larvae. One set of nine vials was moved to 18°C, the other was left at RT.
After pupation, individuals of Ceratitis and Musca were collected from food and kept in vials until eclosion with a wet sponge, which was refreshed every second day.
The experimental temperature regimes were chosen for the following reasons. D. melanogaster is known to survive in the range 10 – 33°C, but flies remain fertile at 12 – 30°C with the optimum at 25°C35. Reproduction temperatures in Ceratitis range from 14°C to 30°C with the optimum at 28°C36,37. Opposite to Ceratitis, Musca flies survive at 10 – 35°C 38 with the optimum at approx. 24°C39. The low temperature for our experiment was chose as the one above the survival and fertile minimums for all three species – 18°C. The warm temperature was aimed to be optimal for each species. During analysis, however, we discovered that RT was likely lower than optimal for Musca13. Therefore, results for this species we interpreted with some caution (see below).
Data collection
For each combination of temperature and density, we randomly picked at least five flies of each sex in Musca and 10 flies of each sex in Drosophila and Ceratitis. Both wings were dissected, embedded in Roti®-Histokitt II (Roth, Buchenau) on a microscope slide, and photographed under the Leica MZ16 FA stereo microscope with the Q Imaging Micro Publisher 5.0 RTV Camera.
Wing shape was analyzed using landmark-based geometric morphometric methods40,41. We digitized 11 anatomically homologous landmarks on wings of the three species (Fig. 1). To provide a better coverage of the wing surface of Ceratitis and Musca wings, we included two additional landmarks (12 and 13). The landmarks were the following (nomenclature is given after42): 1, branching point of veins R1 and RS (base of R2+3 and R4+5); 2, branching point of veins R2+3 and R4+5; 3, intersection of veins C and R1; 4, intersection of vein R4+5 and crossvein r-m (anterior crossvein); 5, intersection of crossvein r-m and vein M1+2; 6, intersection of vein M1+2 and crossvein i-m (posterior crossvein); 7, intersection of crossvein i-m and vein M3+4; 8, intersection of M3+4 and the wing margin; 9, intersection of veins C and R2+3; 10, intersection of veins C and R4+5; 11, meeting point of the anal part of the wing and the alula; 12, intersection of veins CuA2 and A1+CuA2 (Ceratitis) or the tip of vein A2 (Musca): 13, intersection of A1+CuA2 and the wing margin (Ceratitis) or the tip of vein A1+CuA2 (Musca).
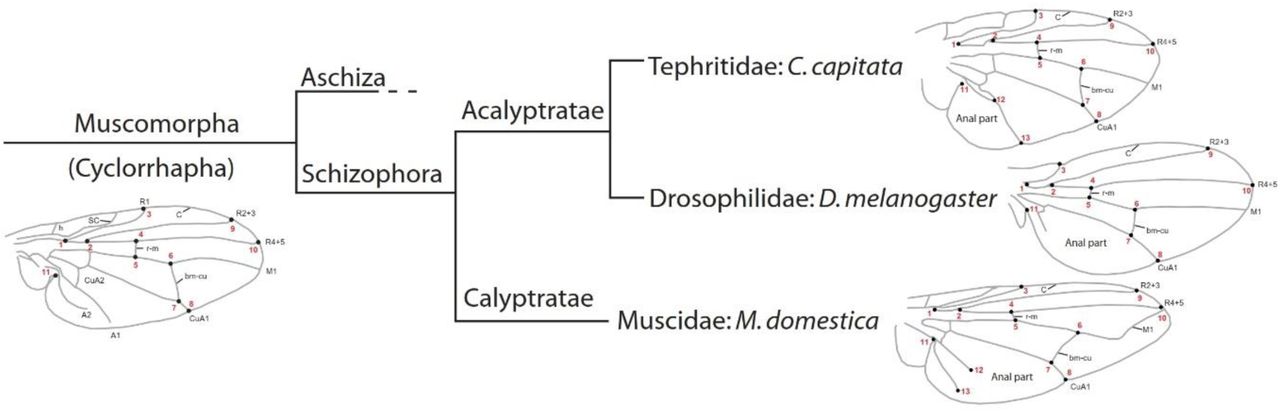
Homologous landmarks 1 to 11 as well as landmarks 12 and 13 in Ceratitis and Musca are shown as black points with the respective number in red. Vein abbreviations: A – anal vein; bm-cu – basal-madial-cubital crossvein; CuA – anterior cubital vein; C – costal vein; h – humeral crossvein; M – medial vein; R – radial vein; r-m – radio-medial crossvein; SC – subcosta. Branch lengths of the tree do not indicate evolutionary time or distance.
Procrustes superimposition and growth trajectories
Wing images were digitized using tpsUtil43,44 and tpsDig245,44 in order to obtain raw x and y landmark coordinates. Using superimposition methods, it is possible to register landmarks of a sample to a common coordinate system in three steps: translating all landmark configurations to the same centroid, scaling all configurations to the same centroid size, and rotating all configurations until the summed squared distances between the landmarks and their corresponding sample average is a minimum scaling46,47. To follow these three steps, we applied the generalized Procrustes analysis (GPA)47,48 in MorphoJ 1.05f49,50. The wings were aligned along the R4+5 vein (landmarks 1 and 10), the mean configuration of landmarks was computed, and each wing was projected to a linear shape tangent space. The coordinates of the aligned wings were the Procrustes coordinates. It has been already shown that fly wings exhibit directional asymmetry51. Because asymmetry was not of interest in this study, we averaged coordinates for the right and left wings of each individual. If only one wing was present, it was used as the mean. The obtained averaged Procrustes coordinates were further used in analyses as the shape variables.
The wing size in this study was quantified as wing centroid size (WCS), computed from raw data of landmarks (11 for Drosophila and 13 for Ceratitis and Musca) and measured as the square root of the sum of squared deviations of landmarks around their centroid26,47,48. Although the extraction of shape from landmarks in GPA removes major variation in size, at this step shape data still contain a size component – the allometric shape variation52,53. This variation accounts for shape changes that occur due to increase in size of the organ. For later analysis, this variation was removed. To determine growth trajectories and characterize morphological changes in response to wing size, we applied a multivariate regression of the Procrustes coordinates on WCS pooling among sub-groups of temperature and density. The amount of shape variation was given as a percentage of the total variation around the sample mean. The percentage numbers were computed to show the relative importance of allometry for shape variation in each species in general and in two sexes separately. A permutation test with 10,000 runs54,55 was applied to test independence between size and shape changes. Additionally, we computed shape scores according to56. These shape scores are the shape variables associated with the shape changes predicted by the regression model. To visualize the association between size and shape, we plotted shape scores against WCS. Similarity between trajectories was estimated with the analysis of covariance (ANCOVA) in R software (aov() package) with WCS being the explanatory variable57.
Interspecies comparison
To compare wing shape variation among species, we created a new dataset with all wings pooled together. All shape comparisons and permutation tests were performed using the MorphoJ software, version 1.05f49,50. To identify and remove the allometric component of the shape variation, we applied a multivariate regression. For the regression, we used WCS computed from the homologous landmarks, 1 to 11 for each species. Subsequently, we performed a Principal Component Analysis (PCA) to visualize the non-allometric component of shape in a scatter plot and visualized morphological differences by thin-plate spline (TPS) deformation grids40,47,58,59. Magnitudes of shape differences between fly wings were computed with the canonical variate analysis (CVA) and expressed in units of Procrustes distance, which is the square root of the sum of squared distances between corresponding landmarks. Significance of the results was tested with permutation tests using 10,000 runs.
Intraspecies comparison: sexual dimorphism, temperature and density effects
Comparison of wing shape within species was performed using 11 landmarks in Drosophila and 13 landmarks in Musca and Ceratitis. Procrustes ANOVA test performed in MorphoJ 1.05f to test whether there were effects of sex, rearing temperature, and density on wing shape. We found clear effects of each parameter and, therefore, continued with more detailed shape analysis.
All following analyses were performed using MorphoJ 1.05f. Sexual shape dimorphism (SShD) was estimated for allometric and non-allometric components of the shape variation together (total SShD) as well as for the pure shape only (non-allometric component). The size correction was performed for each species by using residuals of the allometric regression. Magnitudes of the SShD were estimated with the discriminant function analysis (DFA) and expressed in units of Procrustes distance. DFA identifies shape features that differ at most between groups relative to within groups and it can only be applied to contrast two experimental groups. Therefore, we used this method to define SShD (males and females), effects of the rearing temperature (high and low) in each species and density effect in two Drosophila groups (25 and 300 larvae per plate). To better visualize wing shape changes, we used species specific warped outline drawings with a different scale factor, mentioned in each figure legend. Additionally, we provide discriminant scores for each DFA. To estimate shape changes originated from the three rearing densities in Ceratitis and Musca, we applied canonical variate analysis (CVA), designed to estimate variation among three or more groups. In addition to CVA, we run a DFA for the two groups representing the density extremes (the highest and the lowest number of larvae per plate/vial). For each test, we ran a permutation test with 10,000 random permutations to test for the significance of the result54.
Results
Wing shape variation in dipteran species
To estimate differences in wing shape between Drosophila, Ceratitis and Musca, we digitized 11 landmarks on anatomically homologous points in all three species. The first ten landmarks marked vein intersections and the eleventh landmark was placed on the alula opening (Fig. 1). We averaged shape for the left and right wings for every individual and corrected for evolutionary allometry to exclude size, which is usually the main factor contributing to the shape variation60. Subsequently, we performed a PCA to visualize non-allometric components of shape in a scatter plot (Fig. 2).

Principal component analysis of shape scatter plot (PC1 and PC2) and associated shape change of the non-allometric shape component of side averaged wings. The TPS deformation grids illustrate shape changes indicating the relative shifts of landmarks along the axes with the PC scale factor +/-0.2. The dots on the grids with the respective landmark number (red) indicate the starting point and the blue lines connect them to the final shape. The mean values of shape variance along PC1 and their projections (dashed lines) are shown for Ceratitis males and females, as well as for Musca flies. The inner panel shows morphometric distances between wing shapes in the three species and p-values for them.
With eleven landmarks included in the analysis, we found the largest morphometric distance between Ceratitis and Drosophila (Procrustes distance = 0.3072; p<0.0001) (Fig. 2, the inner panel). Musca wings were more similar to Ceratitis wings (Procrustes distance = 0.1857; p<0.0001) than to those of Drosophila (Procrustes distance = 0.2357; p<0.0001). The PCA revealed that the first two PCs accounted for almost 98% of the variation among species. The main shape difference was reflected in PC1 (80.9% of the variation) and represented the ratio between the proximal and distal parts of the wing. TPS deformation grids showed that Ceratitis wings were broad in the proximal part (landmarks 1-5, 11) and narrow in the distal part (landmarks 6-10). Drosophila wings represented the opposite case with the proximal part being heavily compressed along the anterior-posterior axis. Musca had an intermediate morphology being, however, more similar to Ceratitis (Fig. 2, TPS deformation grids along PC1). Interestingly, along PC1, Ceratitis male wings (PC1mean = -0.17) and Musca wings (PC1mean = -0.04) were almost equidistant from female Ceratitis wings (PC1mean = -0.11), demonstrating a strong sexual dimorphism in Ceratitis, comparable to the interspecies difference along this axis (Fig. 2).
The second significant PC explained 16% of the variation mainly accounting for the ratio between the length and width of the whole wing. In Ceratitis and Drosophila, landmarks 3, 7, and 8 were shifted from the center towards the margin, increasing the wing width (Fig. 2, TPS deformation grids along PC2). At the same time, landmarks 2 and 6 were displaced towards the center, resulting in shorter wings. Landmarks 9 and 10 additionally increased these effects and Ceratitis and Drosophila wings were more compact in comparison with elongated Musca wings. Along the PC2 axis, sexual dimorphism in Ceratitis was less pronounced than the interspecies difference with Musca.
Sexual dimorphism in growth trajectories
In order to estimate the impact of the allometric component on wing shape in the three species, we pooled flies in sub-groups defined by sex, temperature, or density and performed a multivariate regression of shape on size61. In all three species, we found strong static allometry (p < 0.0001; Table 2) but the amount of shape variation explained by variation in size was similar and relatively small in all species (Ceratitis: 4.03%; Musca: 4.71%; Drosophila: 6.04%; Table 2).
The shape variation predicted by each regression is shown as a percentage of the total shape variation. † – non-significant at 5% level.
When we performed the multiple regressions for males and females separately, we found signatures of static allometry for both sexes in Drosophila and Ceratitis (p < 0.0001; Table 2 and Fig. 3). In Musca, we did not observe static allometry in females (p=0.061) but in males it was highly significant (p=0.0008) (Table 2). Sex specific regressions increased the predicted percentage of shape variation explained by size differences up to 7.7 % (Table 2). An ANCOVA showed that growth trajectories were significantly different between sexes in all three species at the 5% confidence level (Table 3 and Fig. 3).

The growth trajectories are shown with shape scores as a function of WCS for males (blue dots and dashed line) and females (red dots and solid line) in the three species.
Sexual dimorphism in wing shape
The PCA and growth trajectory analyses suggested a high level of sexual shape dimorphism (SShD) of wings in the three studied species (see also5,62,63). A Procrustes ANOVA with sex, rearing temperature, or density chosen as the main effects confirmed this finding (Table 4). Therefore, we first applied a DFA and characterized the extent of the SShD in Drosophila, Ceratitis and Musca. Subsequently, we split flies by sex into two groups and examined the effects of temperature and density on shape more closely.
Df – degrees of freedom
The DFA revealed that male and female Drosophila wings were significantly different in shape, and both allometric and non-allometric components contributed to this difference (Fig. 4). The total SShD of Drosophila wings was highly significant (p<0.0001). After size correction, shape difference decreased but remained significant (p=0.01), demonstrating a large impact of wing size on shape in both males and females. Male wings were broader than female wings, radial veins R2+3, R4+5, and M1 were spread apart, but the length of the wing was not affected by the size correction. A comparison of the total SShD and non-allometric SShD in Drosophila suggested that the allometric component introduced variation in the wing length and resulted in shorter and more pointed female wings.

Total and non-allometric SShD in D. melanogaster (scale factor 9, n=168), C. capitata (scale factor 1, n=154) and M. domestica (scale factors 1, n=107). Wing outlines represent differences between male (dark blue) and female (red) average wing shapes. The magnitude of SShD is indicated in units of Procrustes distance with the corresponding p-values (* p<0.05; *** p<0.0001). Histograms with the distribution of the discriminant scores show shape separation into two distinct groups for each species.
In contrast to Drosophila, the extant of the total vs. non-allometric wing SShD was very similar in Ceratitis and Musca (Fig. 4), suggesting that the allometric component has less impact on the shape in these two species. Similar to Drosophila, Ceratitis males had broader wings compared to females. Male wings were also shorter, mainly due to the contraction of the distal anterior region between landmarks 3-5, 9, and 10. Ceratitis wings were wider both proximally and distally, while Drosophila wings were rounded in its distal part only, the proximal part did not change. These results suggest that the major difference between sexes observed in Ceratitis can be explained by the non-allometric component.
In Musca, SShD was opposite to the other two flies: male wings were narrower than those of females, but the anal part of the wing was significantly enlarged. Also, male wings were slightly longer than those of Musca females (Fig. 4).
Sexual dimorphism in response to rearing temperatures
To further characterize the effects of different rearing temperatures on wing shape in males and females separately, we first identified and removed the allometric component of the shape variation. In each species, the DFA analysis clearly assigned wings to one of two rearing temperatures (Fig. 5). In Ceratitis, rearing temperature slightly affected wing shape, but in Drosophila the effect was strong. In general, wings of flies grown at warm temperature (25°C for Drosophila and 28°C for Ceratitis) were broader than wings of those grown at 18°C (Fig. 5, orange and blue wing outlines). The highest plasticity and variation in width was observed in the proximal part of the wing in both species. Another temperature effect was a shortening of wings at warm temperature. In Drosophila and Ceratitis, the direction and strength of the shape change was similar in both sexes. In contrast, shape changes caused by temperature in Musca were not that consistent between sexes. In particular female wings underwent very light changes, while male wings were narrower at higher rearing temperatures (Fig. 5).

Wing shape alterations in response to temperature in D. melanogaster (n(males)=83; n(females)=85), C. capitata (n(males)=81; n(females)=78), and M. domestica (n(males)=56; n(females)=51). Wing outlines represent differences between male cold (light blue) and warm (orange) temperatures with the scale factor 5 for each species. The magnitude of shape variation is indicated in units of Procrustes distance with the corresponding p-values (* p<0.05; ** p<0.001; *** p<0.0001). Histograms with the distribution of the discriminant scores show shape separation into groups and significance of the changes.
In addition to changes in the wing outline in general, we also found displacements of crossveins. A shift of the radio-medial (r-m) crossvein, defined by the landmarks 4 and 5, was similar in all three species (Fig. 1 and Fig. 5). Interestingly, a shift of this vein was the only significant shape alteration in response to different rearing temperatures in wings of Musca females (Fig. 5). Displacement of the basal-medial-cubital (bm-cu) crossvein, defined by the landmarks 6 and 7, was variable in the direction and angle. In Drosophila wings, at higher temperature, the entire bm-cu crossvein was shifted towards the margin of the wing. In Ceratitis, the landmark 7 did not change the position, but the upper part of the vein (landmark 6) was shifted along the M1 vein. The bm-cu crossvein was not affected in Musca females, but in males the landmarks 7 and 8 were displaced towards the wing center and resulted in a narrow wing.
Sexual dimorphism in response to different larval densities
Density is another powerful factor known to influence wing shape in insects25. In order to characterize the impact of the rearing density on wing shape, we grew flies in high and low densities. For Ceratitis and Musca, which are less studied in this respect, we set up intermediate density groups. In the analysis, we first focused on the two density extremes (the high and the low) because we expected to find there the most pronounced shape variation (Fig. 6). Later, we included the intermediate groups as well.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
D. melanogaster males (a(m)) (n=83) and females (a(f)) (n=85), C. capitata males (b(m)) (n=51) and females (b(f)) (n=56) and M. domestica males (c(m)) (n=36) and females (c(f)) (n=34). Wing outlines represent differences between low (green) and high (violet) densities with the scale factor 9 for each species. The magnitude of shape variation is indicated in units of Procrustes distance with the corresponding p-values (* p<0.05; ** p<0.001; *** p<0.0001). Histograms with the distribution of the discriminant scores show shape separation into groups and significance of the changes. CVA scatter plots show distribution of different density groups and the equal frequency ellipses with probability 0.75. The bottom right CVA scatter plot shows distribution of Musca density groups for all flies together (n=107).
In Drosophila, the high rearing density resulted in elongated, narrowed and more pointed male wings with R2+3, R4+5, and M1 veins being close together (Fig. 6A(m)). Drosophila female wings responded in the opposite way: they became more rounded, mainly due to the stretching of the distal part and a shift of the R2+3 and M1 veins (Fig. 6A(f)). The r-m crossvein was displaced in both sexes, and the dm-cu crossvein was shifted in parallel with the r-m crossvein in females only (Fig. 6A). Statistical significance of the difference caused by different densities was confirmed with a permutation test for Procrustes distances run with the DFA (pmale=0.004; pfemale=0.01) and a separate Procrustes ANOVA test (Fmale=10.22, dFmale=18, pmale<0.0001; Ffemale=16.23, dFfemale=18, pfemale<0.0001) (Fig. 6A, discriminant scores).
Unlike Drosophila, Ceratitis male wings were shorter and slightly broader at the high density. The maximum change was observed in the proximal anterior region (Fig. 6B(m)). The same region, together with the anal part of the wing, was enlarged in female wings, but the length remained unaltered (Fig. 6B(f)). Again, we observed a displacement of the r-m and dm-cu crossveins, which was in the direction similar to Drosophila. The observed changes were statistically significant for the density extremes in both sexes (DFA and CVA permutation tests for Procrustes distances: pmale≤0.05 and pfemale≤0.007; Procrustes ANOVA Fmale=2.88, dFmale=22, pmale<0.0001 and Ffemale=3.62, dFfemale=22, pfemale<0.0001) (Fig. 6B, discriminant scores). Wings of flies raised at intermediate densities exhibited an intermediate wing shape (Fig. 6B, discriminant scores and CVA scatter plots).
Musca males and females responded similarly to the high rearing density. The posterior part of the wing was decreased, the landmark 8 was shifted towards the wing center, the r-m and dm-cu crossveins were displaces but no variation in the wing length was found (Fig. 6C). In females, the landmark 3 and the whole C vein were shifted posteriorly and distinguished them from males. Despite the clear wing changes, the observed shape differences between the extreme rearing densities were not statistically significant in the DFA (permutation tests: pmale=0.05 and pfemale=0.1) (Fig. 6C, discriminant scores), while the Procrustes ANOVA test clearly assigned wings in two distinct groups (Fmale=1.74, dFmale=22, pmale=0.01 and Ffemale=1.71, dFfemale=22, pfemale=0.02). Similarly, permutation tests for CVAs gave contradicting results: p-values from permutation tests for Procrustes distances were statistically insignificant (pmale=0.05 and pfemale≤0.1), while p-values for Mahalanobis distances were highly significant for both sexes (pmale and pfemale<0.0001) (Fig. 6C, discriminant scores and CVA scatter plots). Because shape changes were similar in males and females in general, with the only exception of landmark 3, we pooled all wings together to increase the sample size and ran a CVA for 107 flies of three densities (Fig. 6C, bottom CVA scatter plot). This analysis allocated the intermediate density group between two extremes, while the highest and the lowest density were statistically different in shape according to the p-values for Procrustes and Mahalanobis distances (p=0.03; p<0.0001) as well as for the Procrustes ANOVA test (F=2.64, dF=22, p<0.0001).
Discussion
SShD in Drosophila, Ceratitis, and Musca wings
Besides interspecific differences in wing shape (Fig. 2), we found a clear SShD in all three species. In Ceratitis, variation between males and females in the proximal vs. distal part of the wing was so extensive that it was even comparable with the interspecific difference, e.g. between Ceratitis and Musca (Fig. 2, PC1) underpinning a strong sexual dimorphism, which was later tested and confirmed quantitatively (see Fig. 4). A detailed analysis of the SShD did not reveal any general trend among three species.
In Drosophila, we observed a very clear total SShD (Fig. 4). When the allometric component was removed, the difference became less prominent but still statistically significant. One of our observations was that male wings were more rounded and R2+3 and R4+5 veins were spread apart unlike in female wings. This observation is in accordance with previously published data by Bitner-Mathé and Klaczko, 199925. In order to characterize the shape differences in more detail, we carefully analyzed all landmarks and found that male wings were mainly wider in their distal part, while the proximal part and vein intersects of this part were rather similar between sexes.
In Ceratitis, males could be distinguished from female by the variation in the width of the proximal and distal wing parts (Fig. 2, PC1 and Fig. 4). We also found an elongation of female wings. The lack of the wing width variation in our strain contradicts a previously published observation of male wings being shorter and wider than female34. These different conclusions might come from the different definition of the “width”. C. Churchill-Stanland et al. defined the wing width as a distance from the anterior costal bristle of the wing to the point of extreme curvature of the second anal cell34. Our approach includes the overall wing shape and shows that by chance the distance chosen by the authors includes a significantly expand anal region. The difference coming from that region enlarged in male and narrow in females might lead to the initial conclusion of SShD in the wing width.
In Musca, the anterior-posterior variation exceeded the proximal-distal variation (Fig. 2, PC2 and Fig. 4). Thus, Musca female wings were wider in the distal region and male wings were wider in the proximal anal region. The latter was the key difference that allowed to distinguish between sexes (Fig. 4).
Sexual dimorphism in size and shape relationships
Variation in size is often known to entail changes in shape27. Therefore, we tested whether a similar trend was present for wings of the three studied dipteran species. We compared growth trajectories between sexes, which supported the presence of SShD in the three species (Fig. 3, Table 3). In Drosophila, we found a clear contribution of the allometric component to the shape difference between males and females (Fig. 4). For instance, a shift of CuA1 along the wing margin described by25, we could only detect when the allometric component was included. In general, exclusion of the allometric coefficient decreased the SShD, suggesting that most of the observed shape differences resulted from differences in wing size. Interestingly, in Ceratitis and Musca, the impact of the allometric component on wing shape in general was rather weak. Thus, the variation in the wing length that was explained by the allometric component in Drosophila was solely explained by the non-allometric component in Ceratitis and Musca.
Overall, we found a clear sexual dimorphism in wing shape in three species. A tight connection of the shape variation and wing size differences in Drosophila suggests that the growth regulation, patterning, and differentiation processes (e.g. vein placement and axis determination) during the larval wing imaginal disc development may be also tightly linked. These processes, however, may be less connected and independently regulated in the other two species. A potential mechanism for the link between the increase in wing size and shape variation may come from a recent analysis of Ethiopian D. melanogaster populations that showed that increased wing size caused by high altitude was accompanied by a loss of buffering against environmental perturbations64. It has recently been shown that the bone morphogenic protein (BMP) signaling pathways is buffered during embryonic dorsal-ventral axis formation by the action of the BMP-binding protein Crossveinless-2 and Jun N-terminal kinase (JNK) pathway activator Eiger (Egr)65. Since the BMP signaling (e.g. 66–68) and JNK pathway are crucial regulators of wing size in D. melanogaster69, these pathways are excellent starting points to study a potential connection between developmental buffering mechanisms and the concerted size and shape control during development.
Phenotypic plasticity in response to changing environmental conditions
In 1999, Bitner-Mathé and Klaczko used a method of adjusted ellipses for the wing shape analysis in Drosophila and found neither displacement of radial veins nor rounding of wings in response to the rearing temperature25. Based on these findings, they suggested that in this conditions wing shape is more stable than wing size. In this study, we showed that despite the position of the longitudinal veins on the wing margin was only minorly affected, a significant difference in wing shape could be seen in Drosophila flies grown at different temperatures (Fig. 5).
Previous studies described a stronger response to temperature by the distal part of the wing leading to more alterations and displacements relative to the proximal part27. In contrast to this finding, we detected a high variation in proximal landmarks (Fig. 5, see landmarks 1, 3 and 11) and only mild changes in distal ones. One potential explanation for these discrepancies might be the range of rearing temperatures. While Debat et al. raised Drosophila flies at stressful temperatures (12°C and 14°C as the cold temperature and up to 30°C as the high temperature)27, we used intermediate regimes (18°C and 25°C). In their work, they showed the overall wing shape differences along the first two CVs. The variation in wing shape of flies raised at 12-25°C was mainly explained by the first CV, while the second CV distinguished the wings of the extreme temperatures (i.e. 12°C vs. 30°C; see27 and Fig. 3 in there). It would be interesting to know what exact changes in the landmark configuration were explained by these two CVs. If the distal part of the wing was more responsive to stress conditions, which were not covered by our temperature regimes, the variation we saw in proximal landmarks is likely to be somewhat similar to their CV1. We also found pronounced changes in the proximal part of the wing in Ceratitis but not in Musca (Fig. 5, see landmarks 1 and 2). In Musca, the proximal wing region was invariable at different rearing temperatures and all variation was restricted to the distal part (Fig. 5).
Temperature regimes used in this study for the Italian Musca strain might be rather extreme than optimal13. The observed wing shape changes might therefore come from the stress response in Musca, similar to shape changes observed by Debat et al. for Drosophila (Fig. 3 in there)27. Drosophila and Ceratitis flies were reared at favorable temperatures and the pronounced changes in proximal regions observed in these flies might be the actual response to temperature regimes within the normal stress-free reaction norm.
In addition to changes in wing shape along the proximal-distal axis, our shape analysis revealed a high variation in the positioning of the r-m (landmarks 4 and 5) and bm-cu (landmarks 6 and 7) crossveins that was common for all three species. Displacement of these veins was found in all temperature and density groups, suggesting that this region represent a very plastic aspect of wing patterning. Further support for this suggestion comes from the loss of buffering against environmental perturbations in high altitude Ethiopian D. melanogaster populations that showed an increase in wing size. It has been shown that this decanalization resulted in a higher level of abnormal wing development with various defects in crossvein development (i.e. incomplete, missing or additional crossveins)64. Wing vein development requires a proper integration of various central signaling pathways, such as epidermal growth factor receptor (Egfr), Notch, bone morphogenetic protein (BMP), Hedgehog (Hh) and Wnt signaling (reviewed in 70–72). A potential link between these pathways and environmental perturbations has been suggested to be mediated by the heat shock protein Hsp90, since mutations in Hsp83, the gene coding for Hsp90, have been shown to result in various morphological abnormalities in adult traits, including the formation of additional crossveins73. Furthermore, an in-depth analysis of the effect of varying levels of Hsp83 expression on wing shape in D. melanogaster revealed that Hsp90 contributes to the buffering of developmental processes against environmental differences74. In the study by Debat et al. (2006), the placement of the r-m and bm-cu crossveins was affected to different degrees depending on the genetic background of the studied flies. Since crossveins develop later than longitudinal veins72 but still use similar signaling pathways (reviewed in 70), these two vein types probably integrate the stress response differently either via Hsp90 or additional mechanisms. Additional factors are indeed likely because Debat et al. stated that Hsp90 alone cannot explain the entire canalization effect and they also propose the involvement of additional factors74. More targeted experimental and molecular studies are necessary to address the developmental basis of the crossvein plasticity in more detail in insect species other than Drosophila.
Potential functional implications of plasticity and sexual dimorphism in wing size and shape for mating behavior
Both size and shape of wings directly influence specific mating behaviors, such as flying or the generation of mating songs. In many dipteran fly species, male individuals produce species-specific courtship songs by fast and repetitive wing movements. For instance, Caribbean fruit fly females of Anastrepha suspensa judge the size and vigor of a potential mating partner by the intensity of its courtship song1,75,76. Apparently, the intensity and audibility of these songs directly depend on the wing-beat frequency that the fly can afford for certain energy costs and wing fragility. In other fly species, such as M. domestica, the mating process is initiated during flight by an attack (“mating strike”) of a male against the back or side of a female. A successful “mating strike” usually results in the immediate landing and start of copulation4.
Our morphometrics analysis revealed that Musca have wings that are longer and narrower than those of Ceratitis and Drosophila (PC2) (Fig. 2). This implies that Musca has the highest relative wing span (b) among the studied flies. The wing span is proportional to the moment of inertia77 and, thus, the moment of inertia should also the highest for Musca wings. Taking into account that the moment of inertia is inverse to the wing-beat frequency77, we conclude that the wing-beat frequency should be the lowest in Musca compared to the other two species. We also observed male Musca wings being more pointed and even slightly elongated compared to female wings, additionally increasing the moment of inertia and required inertial power. Therefore, in accordance with the “mating strike” behavior in flight, Musca wings may be less suited for buzzing, but their wing shape might be under selection for better flight performance, that is facilitated by long and narrow wings78.
In contrast to Musca, Drosophila and Ceratitis produce courtship songs by buzzing. Usually, their females favor males with a higher audibility2,34. Our study revealed that male wings were wider than female, shorter in case of Ceratitis, and radial veins were more spread apart making wings more compact (Fig. 4). The allometric component of shape additionally increased this difference in Drosophila. The short, wide and rounded wings of males in these species are likely to displace more air and repeat calling song pulses more quickly than long narrow wings, and the wing moment of inertia could be low enough to buzz. Interestingly, these flies produce two different types of wing vibration during the pre-mount courtship: the pulse song and sine song1,79–82. The sine song is a continuous sinusoidal humming generated by small amplitude wing vibrations83–87. In Drosophila, its frequency ranges from 110 to 185 Hz88, with the median value of approx. 160 Hz88 or sometimes 130 Hz89. In Ceratitis, this frequency is similar to Drosophila, 165 Hz90. On the other hand, there is a difference in the pulse songs between these species. In case of Drosophila, it is composed of a series of single pulses (one to three cycles) separated by interpulse intervals. The frequency of these pulses is between 200–280 Hz with the median value of 240 Hz91,92. Instead of pulses, Ceratitis use a continuous vibration of wings when a male looks towards a female and keeps its abdomen bent ventrally80. An average frequency of such buzzing is about 350 Hz1, which is almost half more frequent. Such high frequency of buzzing would require a lower moment of inertia, what could be achieved when the wing mass is concentrated near the axis of rotation93. Intriguingly, our shape analysis revealed major differences between Ceratitis and Drosophila exactly in this region – variation in the width in proximal vs. distal regions (Fig. 2, PC1 axis). Thus, Ceratitis had wider wings in the proximal part appropriate for high frequency buzzing and in Drosophila this part was narrower but perhaps wide enough for low frequency buzzing. Although this hypothesis remains to be tested, it is tempting to speculate that these shape differences may be linked to the mating behavior of the flies and specific properties of their courtship songs.
Author Contributions
N.P. and E.A.W. conceptualized the research; N.S. performed experiments, conducted data analysis, and curated data; N.S. was responsible for data visualization; N.S. and N.P. wrote the original draft and N.P., E.A.W. and N.S. revised the manuscript; N.P. administered the project.
Data availability
All data generated for this work is available from the authors upon request.
Competing Interests
The authors declare that they have no competing interests.
References
- 1.↵
- 2.↵
- 3.↵
- 4.↵
- 5.↵
- 6.↵
- 7.↵
- 8.
- 9.
- 10.
- 11.
- 12.
- 13.↵
- 14.↵
- 15.
- 16.↵
- 17.↵
- 18.↵
- 19.↵
- 20.↵
- 21.↵
- 22.
- 23.↵
- 24.↵
- 25.↵
- 26.↵
- 27.↵
- 28.
- 29.↵
- 30.↵
- 31.
- 32.
- 33.↵
- 34.↵
- 35.↵
- 36.↵
- 37.↵
- 38.↵
- 39.↵
- 40.↵
- 41.↵
- 42.↵
- 43.↵
- 44.↵
- 45.↵
- 46.↵
- 47.↵
- 48.↵
- 49.↵
- 50.↵
- 51.↵
- 52.↵
- 53.↵
- 54.↵
- 55.↵
- 56.↵
- 57.↵
- 58.↵
- 59.↵
- 60.↵
- 61.↵
- 62.↵
- 63.↵
- 64.↵
- 65.↵
- 66.
- 67.
- 68.
- 69.↵
- 70.↵
- 71.
- 72.↵
- 73.↵
- 74.↵
- 75.↵
- 76.↵
- 77.↵
- 78.↵
- 79.↵
- 80.↵
- 81.
- 82.↵
- 83.↵
- 84.
- 85.
- 86.
- 87.↵
- 88.↵
- 89.↵
- 90.↵
- 91.↵
- 92.↵
- 93.↵
Acknowledgments
We thank Y. Wu and L. Beukeboom for providing Musca flies. Many thanks to Micael Reis for critical comments on the manuscript. This work has been funded by a German Academic Exchange Service (DAAD) fellowship number 91540915 to NS, the Göttingen Graduate School for Neurosciences, Biophysics, and Molecular Biosciences (GGNB) and the Volkswagen Foundation (project number: 85 983; to NP).




