

Abstract
In physiological settings, all nucleic acids motor proteins must travel along substrates that are crowded with other proteins. However, the physical basis for how motor proteins behave in these highly crowded environments remains unknown. Here we use real–time single molecule imaging, kinetic Monte Carlo simulations, and Molecular dynamics simulations to determine how the ATP-dependent translocase RecBCD travels along DNA occupied by tandem arrays of high affinity DNA–binding proteins. We demonstrate that RecBCD forces each protein into its nearest adjacent neighbor, causing rapid disruption of the underlying protein–nucleic acid interface. This mechanism is not simply the same way that RecBCD disrupts isolated nucleoprotein complexes on otherwise naked DNA. Instead, molecular crowding itself completely alters the mechanism by which RecBCD removes tightly bound protein obstacles from DNA.
Significance statement Chromosomes are crowded places, and any nucleic acid motor proteins that act upon DNA must function within these crowded environments. How crowded environments affect motor protein behaviors remains largely unexplored. Here, we use single molecule fluorescence microscopy visualize the ATP-dependent motor protein RecBCD as it travels along crowded DNA molecules bearing long tandem arrays of DNA-binding proteins. Our findings show that RecBCD can push through highly crowded protein arrays while evicting the proteins from DNA. Molecular dynamics simulations suggest that RecBCD forces the proteins into once another, causing rapid disruption of the protein-DNA interface. These findings may provide insights into how other types of motor proteins travel along crowded nucleic acids.
Introduction
Long stretches of naked DNA do not exist in living cells. Instead, chromosomes are bound by all of the proteins that are necessary for genome compaction, organization, regulation and maintenance. DNA polymerases, RNA polymerases, helicases, translocases, chromatin-remodeling complexes, must all travel along the highly crowded nucleic acids that exist within these physiological settings. There is a growing appreciation that ATP-dependent motor proteins are required to either remove or remodel nucleoprotein complexes that may otherwise block normal processes related to nucleic acid metabolism, including DNA replication, transcription and DNA repair (1–9). Despite this importance, there remains almost no detailed mechanistic information describing how molecular motor proteins of any type behave on highly crowded nucleic acids.
RecBCD is a large (330–kDa) heterotrimeric complex that has served as an important model system for understanding the properties of nucleic acid motor proteins (10–13). RecBCD processes double–stranded DNA breaks (DSBs) during homologous recombination and replication fork rescue in Escherichia coli, and also degrades linear chromosome fragments to prevent aberrant DNA replication or recombination (10, 11, 14). Interestingly, RecB and RecC are the only two recombination proteins necessary for cell viability when head-on replication-transcription collisions are exacerbated by inversion of the ribosomal RNA operon (15). In addition to its roles in protecting genome integrity, RecBCD is also a self–defense enzyme that degrades foreign invaders, such as bacteriophage, and the resulting DNA fragments are incorporated into the CRIPSR locus, providing immunity against further infection (16). RecBCD possesses two ATP-dependent Superfamily 1 (SF1) molecular motor proteins, the 3’ →5’ SF1A helicase RecB (134-kDa), and the 5’→3 SF1B helicase RecD (67-kDa) (13). The RecC subunit (129-kDa) holds the complex together and coordinates the response to 8-nucleotide cis-acting Chi (crossover hotspot instigator) sequences (5’-dGCTGGTGG-3’). RecD is the lead motor before Chi, RecB is the lead motor after Chi and Chi recognition is accompanied by a reduced rate of translocation corresponding to the slower velocity of RecB (17, 18). RecB also contains a nuclease domain necessary for DNA-end processing, and recognition of Chi results in the production of 3’ single-stranded DNA overhangs onto which the recombinase RecA is loaded (10, 14).
All biological functions of RecBCD require it to travel along DNA that will be occupied by other proteins, and as such RecBCD has been used as a model for studying how motor proteins respond to DNA–bound obstacles (19, 20). Single–molecule observations have revealed that RecBCD can disrupt a variety of tenaciously bound nucleoprotein complexes, including EcoRIE111Q, RNA polymerase (RNAP), and lac repressor. RecBCD does not slow or pause during collisions with individual proteins. Instead, RecBCD appears to evict each of these different proteins through a common mechanism in which probability of protein dissociation is directly proportional to the number of steps they are forced to take as RecBCD pushes them from one nonspecific site to the next (19). RecBCD is even capable of stripping nucleosomes from DNA, highlighting it as an extremely powerful molecular motor (19, 20).
Here we sought to establish whether RecBCD could translocate along DNA substrates that were occupied by the high-affinity DNA-binding proteins EcoRIE111Q or E. coli RNA polymerase holoenzyme. We demonstrate that under crowded conditions, RecBCD quickly and sequentially clears these nucleoprotein complexes from DNA by pushing adjacent proteins into one another. Our results suggest a model in which RecBCD uses DNA-bound proteins as molecular levers to generate torque, which rapidly destabilizes the underlying protein-nucleic acid interfaces. This unique mechanism of sequential protein disruption from the tandem arrays is entirely distinct from how RecBCD removes isolated protein complexes from DNA, indicating that molecular crowding itself can alter the mechanism by which ATP-dependent molecular motor proteins respond to nucleoprotein obstacles. These findings provide new insights into how molecular motors behave while traveling along nucleic acids in crowded physiological settings.
Results
Models for protein eviction in crowded environments
As an initial step towards understanding how motor proteins might behave in crowded settings, we first considered three generalized scenarios describing potential outcomes of RecBCD collisions with protein arrays (Fig. 1A). In the (i) accumulation model, RecBCD pushes each protein into its nearest neighbor without dislodging any of the proteins from the DNA, resulting in greater resistance as proteins continue to accumulate in front of the translocase. For the (ii) sequential model, RecBCD actively evicts each protein as it is encountered. In the (iii) spontaneous model, the proteins spontaneously dissociate according to their intrinsic dissociation rate constants, and RecBCD must wait for these dissociation events before moving forward. We include the spontaneous model as a formal possibility, although we note that this model is unlikely to be correct for RecBCD because previous experiments have shown that RecBCD can quickly push EcoRIE111Q, RNAP and lac repressor off of their respective cognate binding sites with no evidence of either slowing or pausing (19). Importantly, the accumulation and sequential models are not mutually exclusive. Indeed, RecBCD readily pushes isolated proteins for extended distances along DNA (19), suggesting that it might also push proteins into one another on crowded DNA. These observations suggest that some variation of the accumulation model could apply for RecBCD acting in crowded environments. To account for this possibility we also considered an alternative variation of the sequential model, in which small numbers proteins can accumulate in front of RecBCD before the accrued resistance leads to sequential dissociation (SI Appendix, Fig. S1A).

(A) Schematic depictions of three generalized models (accumulation, sequential, and spontaneous) for RecBCD movement through extend protein arrays. Details of each model are presented in the main text. (B) Results from kinetic Monte Carlo simulations for each different model showing the predicted relationships between RecBCD pause durations (〈Δt〉) and protein array size. (C) Predicted postarray RecBCD velocities for each model.
Monte Carlo simulations of protein eviction by DNA translocases
We next performed Kinetic Monte Carlo (KMC) simulations as a means to predict potential experimental outcomes for each of the different models. For these KMC simulations, we modeled the behavior of RecBCD on DNA substrates bearing 5-50x tandem arrays of the high-affinity DNA-binding protein EcoRIE111Q. This protein is a catalytically inactive version of the EcoRI restriction endonuclease that binds tightly to DNA, but is unable to cleave its cognate target site (21). We chose EcoRIE111Q for our computational and experimental studies because it is one of the highest affinity DNA–binding proteins known to exist, with site–specific and non–specific dissociation constants (Kd) of ~2.5 femtomolar (fM) and ~4.8 picomolar (pM) (21), respectively. EcoRIE111Q is also a highly potent block to both the transcription (22–24) and DNA replication machineries (25, 26). In addition, wild–type EcoRI can withstand up to ~20–40 pN of applied force (27). EcoRIE111Q binds to its cognate target ~3000–fold more tightly than wild–type EcoRI, therefore, we infer that EcoRIE111Q can resist at least as much force as the wild–type protein.
Within each KMC simulation, the DNA–bound proteins must either slide or dissociate upon collision with RecBCD. The accumulation model is realized by prohibiting dissociation of the obstacle proteins, which are instead always pushed by RecBCD. The sequential model requires RecBCD to provoke protein dissociation prior to moving forward. In the spontaneous model, RecBCD must wait until proteins dissociate according to their intrinsic dissociation rate constants. Each model predicts that RecBCD will slow or stall upon encountering the array; these events should be revealed as an experimentally observable pause coinciding with the location of protein array (SI Appendix, Fig. S1A). Importantly, the accumulation, sequential, and spontaneous dissociation models all yield distinct predictions for the relationship between apparent pause duration (〈Δt〉) and array size (Fig. 1B and SI Appendix, Fig. S2B). The accumulation model predicts an exponential increase in pause duration with increasingly large protein arrays, the sequential model predicts a linear relationship between pause duration and array size, and the spontaneous model predicts a logarithmic variation in pause duration for the different array sizes. The spontaneous dissociation model also predicts RecBCD will traverse the array orders of magnitude more slowly than the other models because RecBCD must wait for each protein to dissociate (Fig. 1B). In addition, the accumulation model predicts that RecBCD will experience a persistent reduction in velocity after traversing the array due to the accumulated resistance of the proteins it must push as it continues to move along the DNA (Fig. 1C). Finally, if small numbers of EcoRIE111Q can build up in front of RecBCD prior to dissociating, then the pause duration will scale approximately linearly with array size as up to five proteins accumulate in front of RecBCD, similar to expectations for a purely sequential model (SI Appendix, Fig. S1B). However, pause duration begins to scale exponentially if larger numbers of proteins can accumulate in front of RecBCD before dissociation, yielding results that would be more similar to the pure accumulation model (SI Appendix, Fig. S1B).
Visualizing removal of unlabeled EcoRIE111Q from crowded DNA
We next sought to establish an experimental approach for directly testing the predictions for each of the different models. To accomplish this we engineered λ-phage DNA molecules bearing tandem arrays of 5, 10, 30 or 50 EcoRI binding sites. We then used single molecule DNA curtain assays to experimentally determine whether quantum dot (Qdot)–tagged RecBCD could traverse these protein arrays (Fig. 2A), and if so, determine which of the models presented above might most closely reflected the experimental data.

(A) Schematic of the DNA curtain assay used to assess RecBCD behavior during passage through protein arrays. The DNA contains four native EcoRI binding sites (as indicated) and a cloned array of 5–50x EcoRI binding sites. The reactions were initiated by addition of 1 mM ATP into the RecBCD buffer (40 mM Tris–HCl [pH 7.5], 2 mM MgCl2, and 0.2 mg ml−1 Pluronic). (B) Representative kymographs showing RecBCD movement through 5–50x arrays in the presence of saturating EcoRIE111Q. Gaps in the RecBCD trajectories result from Qdot blinking. (C) Experimentally observed pause distributions for each array. Each data set is fitted by a Gaussian distribution to derive the average pause duration. (D) Experimental pause duration (〈Δt〉) plotted against array size. Error bars represent s.d. derived from a bootstrap analysis. (E) Scatter plot showing RecBCD pre– and post–collision velocities; color-coding is the same as in (B) and (C).
In the absence of EcoRIE111Q, Qdot–tagged RecBCD displayed high processivity and monotonic translocation with two peaks in the velocity distribution, corresponding to 745±37 (13.6%) and 1,368±18 bp sec−1 (86.4%), in good agreement with reports for the properties of unlabeled RecBCD on naked DNA (SI Appendix, Fig. S3A and B) (18, 28, 29). We next asked whether the presence of EcoRIE111Q affected the translocation behavior of RecBCD. Remarkably, RecBCD was still able to processively translocate along the DNA in the presence of saturating EcoRIE111Q concentrations (Fig. 2B and SI Appendix, Fig. S4). However, as predicted by the KMC simulations, RecBCD exhibited a pronounced pause upon encountering each of the protein arrays (Fig. 2B and C), and the average pause duration (〈Δt〉) scaled linearly with array size (Fig. 2D). Control experiments with unlabeled RecBCD and unlabeled EcoRIE111Q were in close agreement with results from Qdot-tagged RecBCD, arguing against the possibility that the Qdot might alter the outcomes of the collisions (SI Appendix, Fig. S5). In addition, RecBCD slowed to an average apparent velocity of just 27.3±3.4 bp sec−1 while traversing the EcoRE111Q arrays, but resumed its pre–collision velocity once beyond the array. This finding indicates that encounters with the tandem high-affinity nucleoprotein complexes had no lasting impact on the translocation properties of the enzyme (Fig. 2E and SI Appendix, Fig. S3C). Together, these results are in closest agreement with expectations for the sequential eviction model, or a variation of the sequential model involving the accumulation of a small number of EcoRIE111Q dimers in front of RecBCD (Fig. 1B and C and SI Appendix, Fig. S1B). We do not yet know precisely how many EcoRIE111Q dimers might accumulate in front of RecBCD before they start dissociating. However, DNA curtain experiments have revealed that RecBCD can concurrently push at least two EcoRIE111Q dimers (SI Appendix, Fig. S1C), whereas the tandem arrays of five EcoRIE111Q dimers cause a pronounced pause in RecBCD translocation (Fig. 2). Therefore, we conclude that between two to five EcoRIE111Q dimers might accumulate in front of RecBCD before the combined resistance leads to their sequential eviction.
Sequential disruption of fluorescent EcoRIE111Q arrays
The KMC simulations and experimental results presented above, together with the observation that RecBCD readily pushes isolated molecules of EcoRIE111Q (19), all suggest that RecBCD sequentially removes proteins from crowded DNA, and that it may do so by pushing the proteins into one another. This model makes at least two important predictions that can be experimentally tested using DNA curtain assays. First, the sequential models predict that passage of RecBCD through a fluorescently tagged protein array should coincide with a linear decrease in the array signal as the proteins are evicted, but the last protein(s) within the array should be pushed for long distances along the naked DNA because they will encounter no resistance from more distal obstacles (19) (Fig. 3A). To test this first prediction, we labeled a λ-DNA substrate bearing a 50x EcoRI binding site array with Qdot -tagged EcoRIE111Q, and then asked whether and how unlabeled RecBCD passed through these arrays. As anticipated, eviction of the fluorescent proteins from the 50x array was initially observed as a linear decrease in the overall fluorescence signal intensity as unlabeled RecBCD moved slowly through the array (Fig. 3B). Also as predicted, upon reaching the end of the array, RecBCD resumed its normal velocity and pushed the remaining protein(s) towards the end of the DNA molecule (Fig. 3B).

(A) Model describing predicted outcome for RecBCD collisions with the 50x EcoRIE111Q arrays; details are presented in the main text. (B) Representative kymograph and corresponding signal intensity profile showing unlabeled RecBCD traversing a 50x array of Qdot–tagged EcoRIE111Q. Reactions were initiated by addition of 1 mM ATP into the RecBCD buffer (40 mM Tris–HCl [pH 7.5], 2 mM MgCl2, plus 0.2 mg ml−1 Pluronic). Gaps in the EcoRIE111Q trace result from Qdot blinking.
The sequential models also predicts that if RecBCD pushes a single proximal EcoRIE111Q into a more distal EcoRIE111Q array, then the resulting collision should coincide with preferential eviction of the proximal protein as it is driven into the larger array (Fig. 4A). To test this prediction, we performed two-color single molecule experiments in which separate aliquots of EcoRIE111Q were labeled with either green (Qdot 605) or red (Qdot 705) quantum dots. The differentially labeled proteins were bound to λ-DNA bearing the 5x EcoRI binding site array. We then visually identified DNA molecules with appropriately dispersed mixtures of red and green EcoRIE111Q bound to the native EcoRI sites within the λ-DNA and the engineered 5x arrays (Fig. 4B). As predicted by sequential models, when unlabeled RecBCD pushed Qdot–tagged EcoRIE111Q into a 5x array, the proximal protein rapidly dissociated from the DNA upon encountering the 5x array in ~93% of experimentally observed collisions (N=25/27) (Fig. 4B). We also compared the fates of isolated EcoRIE111Q molecules bound to the native EcoRI sites located either upstream or downstream of the engineered 5x array (Fig. 4C). As predicted by the sequential model, the upstream proteins most commonly dissociated from the DNA upon being pushed into the 5x EcoRIE111Q arrays (Fig. 4C and D). In striking contrast, the downstream proteins were pushed for much longer distances and often survived until reaching the chromium (Cr) barrier at the tethered ends of the DNA molecules (Fig. 4C and D). Taken together, these experimental findings all support a model in which RecBCD clears crowded DNA of tightly bound EcoRIE111Q by pushing the proteins into one another, resulting rapid and sequential removal of proteins from the DNA.

(A) Model describing the predicted outcome for two-color labeling experiments designed to test for sequential protein eviction by RecBCD; details are presented in the main text. (B) Examples of two–color kymographs showing Qdot–tagged EcoRIE111Q (Qdot 705; magenta) being pushed into a 5x array bound by Qdot −tagged EcoRIE111Q (Qdot 605; green). Reactions were initiated by addition of 1 mM ATP into the RecBCD buffer (40 mM Tris–HCl [pH 7.5], 2 mM MgCl2, plus 0.2 mg ml−1 Pluronic). Gaps in the EcoRIE111Q traces result from Qdot blinking. (C) Schematic illustration of the experiment used to assess the fates EcoRIE111Q bound to native target sites in the λ-DNA located either upstream or downstream of the 5x EcoRI array (upper panel), and resulting dissociation positions of the upstream and downstream EcoRIE111Q molecules (lower panel). (D) Survival probability plots for EcoRIE111Q molecules located upstream and downstream of the 5x EcoRI arrays.
Molecular dynamics simulations of protein-protein collisions
The molecular events that take place during the collisions would be extremely difficult, if not impossible, to analyze experimentally due to the short spatial and temporal scales. Therefore, we used Molecular Dynamics (MD) simulations to more closely examine what might take place when RecBCD encounters with EcoRIE111Q in crowded settings. The MD simulations utilized coarse–grained representations of the DNA and EcoRIE111Q, and collisions were conducted on 100-or 150-bp DNA fragments bearing either two or three EcoRI binding sites, respectively (Fig. 5). The interaction parameters for the proximal EcoRIE111Q molecule(s) recapitulated the experimentally observed specific and non–specific EcoRIE111Q dissociation constants (SI Appendix, Fig. S6), and the distal EcoRIE111Q was bound more tightly to the DNA to emulate the accumulated resistance of several nucleoprotein complexes. RecBCD was modeled as a simple torus placed at one end of DNA, and exerted a constant force of 100 pN as it pushed against the DNA–bound molecules of EcoRIE111Q. The force that RecBCD can exert upon colliding with a DNA-bound obstacle is unknown, so we chose 100 pN for the MD simulations because this value is within an order of magnitude of several of the most powerful molecular motors that have been characterized (30). The expectation that RecBCD would be capable of exerting a force comparable to the most powerful molecular motor proteins is consistent with the finding that RecBCD can clear DNA bound by 50x arrays of EcoRIE111Q, and is also consistent with the finding that RecBCD readily overwhelms the hexameric motor protein FtsK, which itself exhibits a stall force of ~65 pN (31).

(A) Snapshots from a representative simulation trajectory involving two EcoRIE111Q molecules (also see SI Appendix). RecBCD begins at one end of the DNA and an arrowhead indicates its direction of travel. The proximal and distal proteins are labeled A and B, respectively. Magenta arrowheads and lettering highlight protein eviction. (B) The time trajectories showing the distance between the two EcoRIE111Q molecules (top), the DNA bending score (middle), and interaction energy between the proximal EcoRIE111Q and DNA (bottom). The zero time point corresponds to the initial collision of proteins A and B. (C) Snapshots from a representative simulation trajectory involving three EcoRIE111Q molecules, labeled A, B and C, as indicated. (also see SI Appendix). (D) Time trajectories showing the distance between the EcoRIE111Q molecules (top; A–B distance, green line; B–C distance, magenta line), DNA bending score (middle), and interaction energy between the EcoRIE111Q molecules (A, green line; B, magenta line) and DNA (bottom). The zero time point corresponds to the initial collision of proteins A, B and C.
A representative MD simulation trajectory of RecBCD traveling on DNA bound by two EcoRIE111Q molecules is shown in Fig. 5A and SI Appendix. As anticipated, RecBCD pushed the proximal EcoRIE111Q along the DNA major groove until it collided with the distal EcoRIE111Q. The proximal protein then dissociated from the DNA as it was forced against the distal EcoRIE111Q in all simulations (N=15/15) (Fig. 5B). The simulation time scale for each dissociation event was on the order of ~1.0 μsec (Fig. 5B), corresponding to ~108–fold rate enhancement relative to spontaneous dissociation and ~106–fold relative to RecBCD–induced dissociation of single proteins in the absence of crowding (Fig. 5B and SI Appendix, Fig. S7)(19). A representative MD simulation trajectory of RecBCD traveling on DNA bound by three EcoRIE111Q molecules is shown in Fig. 5C and SI Appendix. Remarkably, all MD simulations (N=15/15) involving RecBCD collisions with three molecules of EcoRIE111Q resulted in sequential eviction of the first and then second proteins (Fig. 5D). Together, these MD simulations provide further support for a model in which RecBCD dislodges EcoRIE111Q from crowded DNA by forcing the proteins into one another, leading to an accumulation of resistance that ultimately causes sequential eviction of individual proteins from within the tandem arrays.
Protein eviction through torque-generated DNA deformation
We next sought to determine more precisely why the proteins dissociated from the DNA when forced into one another by RecBCD. Detailed analysis of the MD simulation trajectories revealed extensive DNA deformation as EcoRIE111Q molecules were pushed together (Fig. 5A to D). Deformation occurs because the EcoRI center–of–mass (COM) is ~24 Å away from the DNA axis, so the force (F) exerted by RecBCD on EcoRIE111Q when it is pushed into the distal obstacle gives rise to torque (F × d), where d is the distance between the DNA and the protein COM. This torque bends the DNA, which destabilizes the interface between EcoRIE111Q and the DNA causing the protein to rapidly dissociate into solution (Fig. 5B and D). We conclude that EcoRIE111Q dissociates from the DNA due to torque-induced deformation of the underlying protein-nucleic acid interface.
It is possible that the extensive DNA bending observed for collisions involving tandem molecules of EcoRI was a unique outcome arising because of the specific geometry of the protein-protein interface that is formed when the two molecules of EcoRI are pushed into one another. To test this possibility, we conducted MD simulations using EcoRI embedded within spherical shells. We varied the radius and COM of the spherical shells to eliminate any protein surfaces or binding geometries that may be specific to EcoRI, allowing us to emulate a series of “generic” high affinity DNA-binding proteins of varying sizes (SI Appendix, Fig. S8A). In all simulations, the proteins still dissociated rapidly from the DNA (SI Appendix, Fig. S8B) and each dissociation event was correlated with extensive deformation of the underlying protein-DNA interfaces (SI Appendix, Fig. S8C). Taken together, these MD simulations suggest that similar eviction mechanisms may be operative for other high affinity DNA-binding proteins, even those with different radii, different molecular interfaces or binding geometries, and different COMs. Interestingly, 87% of all protein-DNA complex structures in the protein data bank (PDB) have a COM that is offset from the DNA axis by ≥5 Å (SI Appendix, Fig. S8D) (32), suggesting the possibility that torque induced DNA deformation might be a general mechanism allowing RecBCD to clear DNA of crowded obstacles.
RNA polymerase rapidly dissociates when forced into EcoRIE111Q
Our results demonstrate that RecBCD sequentially removes EcoRIE111Q from DNA by pushing the proteins into one another. We next sought to determine whether similar principles apply to other nucleoprotein roadblocks. RNA polymerase is perhaps the most abundant and formidable nucleoprotein roadblock that will be encountered by RecBCD in living cells. A single Escherichia coli cell contains ~2,000 molecules of RNAP, and under typical growth conditions ≥65% of these polymerases are bound to the bacterial chromosome (33). RNAP is also of particular interest because it is a high-affinity DNA-binding protein (Kd ~10–100 pM) and a powerful translocase capable of moving under an applied load of up to ~14–25 pN (34). RNA polymerases can survive encounters with replication forks and stall fork progression in head-on collisions (35–39). Indeed, the highly transcribed ribosomal RNA genes are a potent blockade to DNA replication (40–42). We have previously shown that RecBCD can disrupt individual E. coli RNAP complexes, including core RNAP, RNAP holoenzyme, stalled elongation complexes and actively transcribing polymerases in either head-to-head or head-to-tail orientations (19). RecBCD pushes isolated RNAP for long distances over naked DNA, and dissociation takes place as RecBCD forces the polymerase to step from one nonspecific binding site to the next (19).
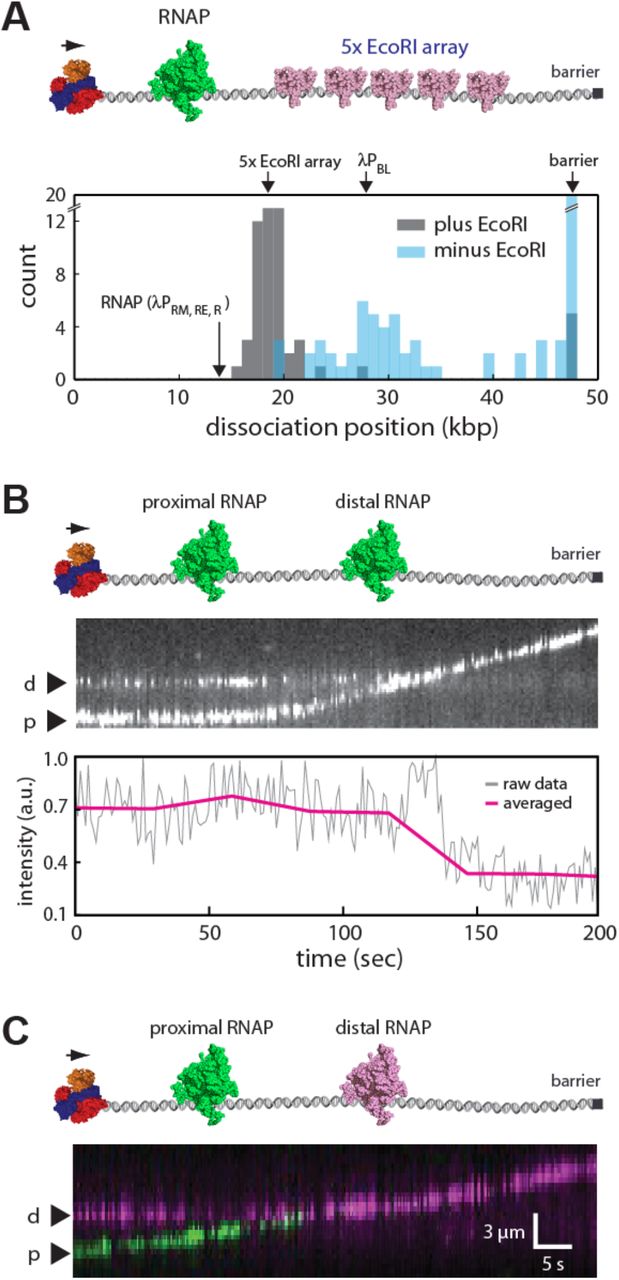
To determine whether and how RecBCD might disrupt RNAP on crowded DNA, we first sought to establish what happens when RecBCD pushes RNAP into tandem 5x arrays of EcoRIE111Q (Fig. 6A). If the sequential model applies to RNAP, then this model predicts that RNAP should rapidly dissociate from the DNA upon being forced into the EcoRI array by RecBCD. For these experiments, Qdot-tagged RNAP holoenzyme was bound the native phage promoters (19, 43), and unlabeled RecBCD was loaded onto the free ends of the DNA molecules. RecBCD translocation was then initiated by the addition of ATP. Remarkably, RNAP dissociates from the DNA almost immediately upon being pushed by RecBCD into the EcoRIE111Q array for all observed collisions (N=22/22) (Fig. 6A). Control experiments confirmed that RNAP dissociation at the 5x EcoRI array position was entirely dependent upon the presence of EcoRIE111Q, and when EcoRIE111Q was absent, many of the polymerases were instead pushed to the ends of the naked DNA molecules (Fig. 6A). We conclude that RNA polymerase rapidly dissociates from the DNA when pushed into other high affinity DNA-binding proteins by RecBCD, in good agreement with the sequential eviction model.

(A) Experimental schematic (upper panel) and resulting dissociation position distribution data (lower panel) for Qdot-tagged RNAP pushed into 5x EcoRI arrays by RecBCD. (B) Representative one-color kymograph showing two Qdot -tagged RNAP complexes (promoter-bound holoenzyme) being pushed into one another by unlabeled RecBCD, and the corresponding graph showing the cumulative fluorescence intensity of the Qdot-tagged proteins. (C) Representative two-color kymograph showing two Qdot –tagged RNAP complexes (promoter bound holoenzyme) being pushed into one another by unlabeled RecBCD; similar outcomes were observed in most (N=50/52) two–color RNAP collisions, and in the remaining cases, RecBCD pushed both proteins (N=2/52). In (B) and (C), reactions were initiated by addition of 1 mM ATP into the RecBCD buffer (40 mM Tris–HCl [pH 7.5], 2 mM MgCl2, plus 0.2 mg ml−1 Pluronic) and gaps in the RNAP trajectories result from Qdot blinking.
Sequential eviction of tandem RNA polymerases
Interestingly, when EcoRIE111Q was absent from the reactions, a second population of RNAP dissociated at a position coinciding with the location of the λPBL promoter (Fig. 6A)(43). One possible explanation for this observation is that the Qdot-tagged RNAP might be encountering unlabeled RNAP bound to the λPBL promoter as it is pushed along the DNA by RecBCD, and the resulting collisions with the unlabeled proteins may have provoked rapid eviction of the Qdot-tagged protein from the DNA. Therefore, we next sought to directly examine what happens when RecBCD pushed two RNA polymerases into one another. To accomplish this, we relied upon the eight native promoters present in the λ phage genome, which allow multiple RNAP complexes to be loaded onto the same DNA molecule (43). We first performed DNA curtain experiments using unlabeled RecBCD and promoter-bound RNAP open complexes, which were labeled with a single color Qdot (Qdot 705) (Fig. 6B). Remarkably, analysis of the cumulative fluorescence intensity of the DNA-bound polymerases suggested that only one of the two polymerases remained on the DNA when RecBCD pushed them into one another (Fig. 6B). We conclude that although RecBCD readily pushes single RNAP complexes along DNA, it does not appear to push two RNAPs at the same time. Instead, as predicted by the sequential eviction model, one of the two polymerases quickly falls of the DNA when they collide with one another.
The results described above provide evidence that the sequential eviction model may apply to RecBCD encounters with RNA polymerases in crowded settings. Importantly, the sequential eviction model specifically predicts that the proximal polymerase should be preferentially evicted when pushed by RecBCD into the distal polymerase. We next sought to verify this prediction by determining which of the two polymerases dissociated from the DNA when forced into one another by RecBCD. We therefore conducted two-color DNA curtain assays in which separate aliquots of RNAP were labeled with either green (Qdot 605) or red (Qdot 705) quantum dots. The differentially labeled polymerases were then mixed together and bound to the native phage promoters, unlabeled RecBCD was bound to the free DNA ends and translocation was initiated by the injection of ATP. These experiments revealed that for ~96% of observed collisions (N=50/52) involving RecBCD and two tandem molecules of RNAP, the proximal polymerase almost immediately dissociated from the DNA upon being pushed into the distal polymerase (Fig. 6C). We conclude that RecBCD can rapidly and sequentially evict RNAP from crowded DNA, and that it does so specifically by forcing the polymerases into one another.
Discussion
Our results demonstrate that RecBCD rapidly and sequentially removes crowded nucleoprotein complexes from crowded DNA by pushing them into one another. These findings reveal that molecular crowding itself can have a crucial and unanticipated influence on how molecular motor proteins clear nucleic acids of bound obstacles.
Molecular crowding alters the mechanism of protein eviction by RecBCD
The mechanism by which RecBCD removes high affinity nucleoprotein complexes from DNA is dependent upon molecular crowding (Fig. 7). When RecBCD encounters isolated molecules of either of EcoRIE111Q or RNA polymerase on otherwise naked DNA, it is able to push these proteins over average distances of 13,000 ± 9,100 bp and 10,460 ± 7690 bp, respectively (19). The proteins eventually dissociate as they are forced to step between successive nonspecific binding sites, and the probability (P) of dissociation is directly proportional to the number of steps (n) that EcoRIE111Q or RNA polymerase are forced to take while being pushed along the DNA (Fig. 7A) (19). However, the probability of dissociating during any given step is low, and as a consequence RecBCD is able to push isolated proteins for extended distances on naked DNA (Fig. 7A). This outcome is in marked contrast to what takes place in crowded environments, where EcoRIE111Q and RNA polymerase both dissociate almost immediately when pushed into more distal proteins or protein arrays (Fig. 7B). This much more rapid eviction takes place on crowded DNA specifically because RecBCD pushes the proteins into one another. Molecular dynamics simulations of collisions involving tandem molecules of EcoRIE111Q suggest that when crowded proteins are forced into one another, RecBCD uses the most proximal protein as a molecular lever to generate torque, which in turn distorts the DNA and destabilizes the underlying protein-nucleic acid interface of the proximal protein (Fig. 7B, and see below). The striking differences between the outcomes of isolated collisions involving single nucleoprotein complexes, compared to the outcomes of collisions on crowded DNA, highlights the dramatic and unexpected impact that molecular crowded has on the mechanism by which RecBCD interacts with nucleoprotein obstacles that it encounters while traveling along DNA. This sequential mechanism for protein eviction may likely reflect what takes place in vivo where long tracts of naked DNA are unlikely to exist (44, 45).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
(A) RecBCD can push isolated proteins for extended’ distances along the otherwise naked DNA, and the probability (P) of dissociation scales proportionally with the number of steps (n) that the protein is forced to take as its pushed by RecBCD (19). (B) In contrast, when RecBCD encounters proteins in crowded environments, it pushes each protein into its nearest adjacent neighbor. The resulting collisions generate torque on the most proximal protein to RecBCD, which distorts the DNA and disrupts the underlying protein–DNA interface resulting in rapid dissociation of the protein. Additional details of the models are presented in the main text.
Sequential removal of RNA polymerase from DNA by RecBCD
Our model for torque-dependent sequential disruption of crowded nucleoprotein complexes has only three basic requirements: (i) a high-affinity DNA-bound protein whose center of mass (COM) is offset from the DNA axis; (ii) a motor protein capable of generating sufficient force to push the DNA-bound protein in question; and (iii) a downstream nucleoprotein complex to provide additional resistance. These simple requirements highlight the generality of the proposed sequential dissociation mechanism, and suggest its potential applicability to other nucleic acid motor proteins and different types of nucleoprotein obstacles. The general applicability of the sequential disruption model is also supported by the similarities between the experimental findings for EcoRIE111Q and E. coli RNA polymerase. Molecular dynamics simulations of collisions involving RNAP are not yet feasible, so we cannot determine whether RNA polymerase dissociation also coincides with extensive deformation of nucleoprotein interface. However, the E. coli RNA polymerase center of mass is offset from the DNA axis by ~26.5 Å (46), suggesting that sequential eviction of RNA polymerase may also be occurring through a torque-dependent mechanism similar to that described for EcoRIE111Q. Future work will be necessary to more precisely determine the extent to which DNA deformation contributes to sequential disruption of RNA polymerase by RecBCD.
Interestingly, our work suggests that a small number of EcoRIE111Q complexes can accumulate in front RecBCD before they start dissociating from the DNA. In contrast to EcoRIE111Q, it does not appear as though multiple molecules of RNA polymerase can accumulate in front of RecBCD. Instead the proximal polymerase dissociates from the DNA almost immediately upon being pushed into a distal protein. These observations indicate that RecBCD removes RNA polymerase from crowded DNA much more easily than it removes EcoRIE111Q. This difference may reflect the fact that RNA polymerase is a naturally occurring obstacle that will likely be encountered whenever RecBCD acts on DNA in a cellular environment. We speculate that co-evolution of RNA polymerase and RecBCD may have tuned to relative binding strengths of these two nucleoprotein complexes to ensure that RNA polymerase cannot impede the movement of RecBCD.
Mechanisms of protein dissociation
Through-DNA allosteric communication can influence the dissociation of stationary proteins that are bound in close spatial proximity to one another (47). Previous studies have shown that protein pairs, including glucocorticoid receptor and BamHI, or lac repressor together with either EcoRV or T7 RNAP, exhibited up to 5-fold changes in dissociation rates when the corresponding partner was bound to a nearby DNA site (47). These experimental findings have been attributed to through-DNA allosteric communication based upon the long-range oscillatory changes in DNA major and minor groove widths observed in MD simulations (47–49). Similarly, we find that a static RecBCD complex positioned immediately adjacent to an EcoRI site causes approximately a 2-fold reduction in DNA cleavage by EcoRI (SI Appendix, Fig. S9). This reduction in cleavage is comparable in magnitude to the effects previously ascribed to the through-DNA allosteric model. However, this 2-fold effect is substantially less than the ~106-fold enhancement in dissociation rates observed as RecBCD traverses an array of EcoRIE111Q, suggesting that through-DNA allostery may not be a predominant factor affecting protein displacement by RecBCD. In addition, our experimental data demonstrate that under crowded conditions, RecBCD causes rapid dissociation of the most proximal protein, but only when it is pushed into a more distal obstacle, indicating that crowded environments enhance protein dissociation by RecBCD relative to isolated collisions. The substantial increase in rate enhancement when RecBCD is moving through a protein array, together with the dependence of proximal protein dissociation on the presence of a more distal obstacle, suggests that the through-DNA allosteric model cannot account for RecBCD-mediated protein displacement under crowded conditions. Instead our MD simulations suggest that RecBCD can push proteins into one another, which is consistent with our experimental observations, and that the resulting collisions give rise to torque that results in rapid destabilization of the proximal protein obstacle. Although the torque-based model comes from MD simulations, the model itself is in good agreement with our experimentally observed finding that RecBCD provokes sequential protein dissociation under crowded conditions. Future work will be essential to further evaluate the torque-based model for protein eviction from crowded environments, and to determine whether other types of DNA translocases may act similarly
Nucleoprotein obstacles and genome integrity
Nucleoprotein complexes are the primary source of replication fork stalling (50) and their presence represents a major challenge to genome integrity (1, 40, 41, 51–53). Indeed, prokaryotic and eukaryotic replisomes both require accessory helicases to clear tightly bound proteins from DNA (25, 54–58). For instance, the E. coli replisome requires the accessory helicases Rep and UvrD to prevent replication fork collapse upon encountering RNA polymerase and other types of high affinity nucleoprotein complexes (25, 50). Similarly, Bacillus subtilis requires the accessory helicase PcrA to facilitate replication fork progression through highly transcribed genes (56). The physical basis by which Rep, UvrD and PcrA assist the replisome remain unknown, however, all three proteins are 3’→5’ SF1A helicases and are closely related to RecB (13), suggesting the possibility that they may strip proteins from crowded DNA through mechanism similar to that used by RecBCD. In addition, both Rep and UvrD can push isolated ssDNA-binding proteins along single-stranded DNA (59) using a mechanism that is similar in many respects to what takes place during RecBCD collisions with isolated proteins (19). Future work will be essential for further establishing how these replication accessory helicases and other types of motor proteins disrupt tightly bound nucleoprotein complexes on crowded nucleic acids.
Conclusion
We have presented a model describing the ability of RecBCD to sequentially clear crowded DNA of nucleoprotein complexes. The key feature of the sequential model is that RecBCD provokes rapid disruption of crowded nucleoprotein complexes by pushing these obstacles into one another. This model suggests that molecular crowding itself alters the mechanism by which RecBCD removes proteins from DNA, and the proposed mechanism is supported by a combination of experimental data, kinetic Monte Carlo simulations and molecular dynamics simulations. The sequential model, which represents the first detailed mechanistic description for the behavior of any motor protein in a highly crowded environment, strongly suggests that ATP-dependent nucleic acid motor proteins can respond differently to encounters with isolated nucleoprotein complexes compared to encounters involving multiple nucleoprotein complexes. The general principles revealed from our studies with RecBCD may also apply to behavior of other types of motor proteins as they travel along crowded nucleic acids.
Acknowledgments
We thank members of the Greene laboratory for discussion and assistance throughout this work, and we thank Shoji Takada, Jayil Lee, Daniel Duzdevich, Fabian Erdel, Corentin Moevus, Kyle Kaniecki, Chu Jian Ma, Johannes Stigler, Justin Steinfeld, Luisina De Tullio, and Mayu S. Terakawa for comments on the manuscript. This work was supported by a Japan Society for the Promotion of Science fellowship to T.T., an NIH Grant (R35GM118026) to E.C.G., and NIH Fellowship (F32GM101819) to T.D.S. The data described in this manuscript are archived in the laboratory of E.C.G. in the Department of Biochemistry and Molecular Biophysics, Columbia University.
References
SUPPLEMENTAL REFERENCES




