

Abstract
The laboratory reference strain 630Δerm of the anaerobic human pathogen Clostridioides difficile is characterized by a remarkable high oxygen tolerance. We show that an amino acid exchange in the DNA binding domain of the hydrogen peroxide sensor PerR results in a constitutive derepression of PerR-controlled genes and thus in an oxidative stress response even under anaerobic conditions. This questions the model status, strain 630Δerm claims in C. difficile research.
Introduction
Clostridioides difficile (C. difficile) is a Gram-positive, anaerobic, spore-forming pathogen causing primarily hospital-acquired, but increasingly also community-acquired infections, which turned the bacterium into one of the most problematic pathogens in human health care nowadays. C. difficile infections (CDIs) are often associated with broad-spectrum antibiotic therapy. Clinical symptoms of CDI vary from light diarrhea to acute infections like pseudomembranous colitis (1).
Due to its anaerobic lifestyle, oxygen (O2) and reactive O2 species in the human intestine represent a challenge for C. difficile. Remarkably, a high tolerance to O2 was recently reported for a sporulation deficient mutant of C. difficile 630Δerm (2). Strain 630Δerm is an erythromycin-sensitive and laboratory-generated derivative of the original patient-isolated strain 630 and is commonly used by C. difficile researchers as reference strain for the generation of gene knock out mutants (3). Although the oxidative stress response is vital for an intestinal pathogen infecting its host, knowledge on the molecular details of oxidative adaptation mechanisms in C. difficile is still limited (4, 5).
Results and Discussion
In previous studies, we observed a high abundance of oxidative stress-related proteins in C. difficile 630Δerm already at conditions devoid of any oxidizing agents (6) and no significant induction when the bacterium was shifted to micro-aerobic conditions (7). Several of the corresponding genes are encoded at one genetic locus comprising a rubrerythrin (rbr1), the transcriptional repressor PerR (perR), a desulfoferrodoxin (rbo) and a glutamate dehydrogenase with an N-terminal rubredoxin fold (CD630_08280). Rbr1 even represents the second most abundant protein after the S-layer protein SlpA (6). We enquired, why these genes are highly expressed in the absence of any oxidative stress and focused on the repressor protein PerR, which regulates its own transcription and the one of genes involved in oxidative stress and metal homeostasis as described in Bacillus subtilis and in other Gram-positive bacteria (8, 9). PerR is a member of the ferric uptake regulator (Fur) family and senses H2O2 stress by metal-catalyzed histidine oxidation (10), (Fig. 1A). Due to the permanently high cellular concentration of proteins encoded in the rbr1 operon, we hypothesized a constitutive expression of the operon possibly caused by failure of PerR-mediated gene repression under anaerobic conditions.

A] Mode of PerR function is shown schematically. PerR bound to the DNA represses expression of the rbr1 operon. H2O2 treatment leads to PerR oxidation, a conformational change and release of the DNA promoter resulting in rbr1 expression. B] Alignment of amino acid sequences of the PerR protein of 13 different C. difficile strains. C] Structure of C. difficile 630Δerm PerR deduced from the homolog of B. subtilis. The DNA binding site including the HTH-motif and the dimerization domain are marked. D] Amino acid sequence of the DNA binding site of C. difficile 630Δerm PerR. The T41A mutation is marked red. E] Alignment of the promoter sequences of the rbr1 gene in C. difficile strains 630 and 630Δerm.
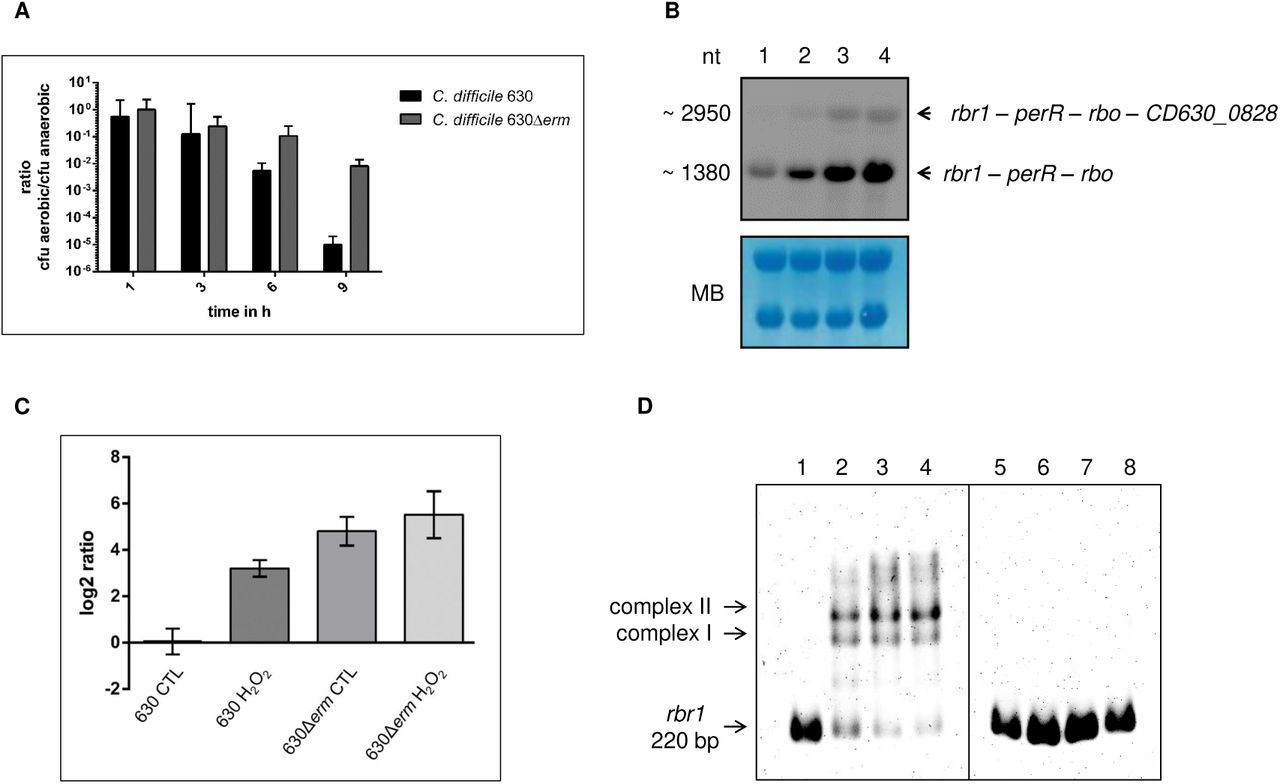
It was reported before, that the lab-generated strain C. difficile 630Δerm features several genome alterations compared to its parental strain C. difficile 630 (11, 12). We aligned perR-sequences of C. difficile 630 and C. difficile 630Δerm and found a single nucleotide polymorphism (SNP, A > G), resulting in an amino acid conversion from threonine to alanine at position 41. An alignment of the PerR amino acid sequence with 11 other clinically relevant C. difficile strains revealed that the T41A substitution is unique to the laboratory strain 630Δerm (Fig. 1B). A comprehensive sequence alignment of over 900 proteins of the Fur family from different species showed that the threonine in position 41 is highly conserved and present in over 80% of the investigated proteins. More than 90% of the proteins contain a threonine or serine at this position (Dataset S1). A structural comparison of previously investigated DNA binding domains of Fur and PerR homologues in Escherichia coli, B. subtilis, Staphylococcus epidermidis and Streptococcus pyogenes to the C. difficile 630Δerm PerR sequence indicates the T41A mutation to be located in a helix of the helix-turn-helix motif of the DNA binding domain (Fig. 1C and D). The DNA promoter sequences upstream of rbr1 are identical between strain 630 and 630Δerm (Fig. 1E). We therefore hypothesized that the amino acid substitution in PerR is the reason for loss of binding of the repressor to PerR boxes on the DNA and possibly causes increased O2 tolerance of strain 630Δerm compared to other C. difficile strains including its parental strain 630. To investigate differences in O2 tolerance between C. difficile 630 and 630Δerm we counted colony forming units (CFU) for both strains after cells have been exposed to atmospheric O2 concentrations (Fig. 2A). Strain 630 showed a significantly higher susceptibility to O2 than its derivative 630Δerm, of which a substantial number of cells survived even after 9 h of challenge.

{kind=link}
{kind=link}
CFU counting, transcriptional analyses and DNA-PerR interaction. A] Survival of C. difficile strains 630 and 630Δerm in the presence of O2. Cell numbers are related to the cell number determined before the aerobic shift. B] Northern blot analysis of expression of rbr1 operon: 630 control (1) and 630 H2O2 induced (2), 630Δerm control (3) and 630Δerm H2O2 induced (4). Total RNA levels were monitored by methylene blue staining (MB). C] Transcription of the rbr1 gene was quantified by RT-qPCR analysis and related to C. difficile 630 control. D] EMSA analyses were carried out with 2 ng of the rbr1 promoter fragment (220 bp), concentrated 147 nM, from strain C. difficile 630. In lane 1 and 5, the DNA fragment was incubated without protein. The DNA fragment was incubated with purified PerR 630 (lane 2 to 4) or PerR 630Δerm (lane 6 to 8) with increasing protein amount (360 ng, 420 ng and 480 ng, respectively).
To prove that the higher O2 tolerance of C. difficile 630Δerm was caused by the missing binding of PerR to its cognate DNA binding site, we analyzed transcriptional levels of the rbr1 operon under hydrogen peroxide (H2O2) stress and control conditions (Fig. 2B and C). Transcription of the operon was inducible by H2O2 in strain 630 by a factor of 9, whereas transcript levels in 630Δerm were permanently very high and almost not inducible by H2O2 (factor of 1.6).
To confirm that the T41A exchange in PerR of strain 630Δerm is the sole reason that hampers PerR-box binding, we performed electrophoretic mobility shift assays (EMSA) excluding any other cellular factors. PerR proteins from strains 630 and 630Δerm were recombinantly produced, purified and incubated with a labelled 220 bp upstream promoter fragment of rbr1. While PerR from strain 630 led to a clear shift of the DNA band, no shift was detectable for PerR from C. difficile 630Δerm at any tested protein concentration (Fig. 2D).
This study demonstrated the constitutive derepression of genes involved in the oxidative stress response in strain C. difficile 630Δerm caused by only one SNP in the DNA sequence of the transcriptional repressor PerR. Since this strain is used as reference in many laboratories for the construction of gene inactivation mutants, researchers should be aware of its permanent oxidative stress response.
Materials and Methods
Bioinformatic methods
Structural analysis of the C. difficile PerR protein was performed using the Phyre2 web portal (13). For the alignment of over 900 Fur family proteins MView was used (14). The alignment of promoter sequences was carried out with Clustal Omega (15). PerR Boxes were identified using Virtual Footprint Version 3.0 (16).
Bacterial strains and growth conditions
C. difficile strains were obtained from the German Collection of Microorganisms and Cell Cultures GmbH (DSMZ, Braunschweig, Germany) and cultured as previously described (17). For Northern Blot and RT-qPCR analyses, C. difficile 630 and C. difficile 630Δerm were grown to an A600 of 0.4, before cultures were split and one of the two subcultures stressed with 0.4 mM H2O2 for 10 min. Samples were taken to allow for a later preparation of RNA (18). For CFU counting experiments, 10 mL of the C. difficile culture at A600 of 0.4 were transferred to a 92 × 16 mm petri dish and aerobically incubated. Samples were taken before and 1, 3, 6 and 9 h after oxygen exposure in three biological replicates, and dilution series incubated anaerobically. For overexpression of PerR, Escherichia coli BL21 grown in LB medium was used.
RNA preparation
For cell lyses and RNA isolation TRIzolTM Reagent provided by Invitrogen (Thermo Fisher Scientific; Waltham, Massachusetts, USA) was used according to the manufacturer’s protocol (19). RNA solubilized in DEPC-treated water was stored at −70 °C.
Transcriptional profiling
A PCR fragment of gene rbr1 was prepared using chromosomal DNA of C. difficile 630 as a template with primers 5’-AATGGCAGGATTTGCAGGAG-3’ and 5’-CTAATACGACTCACTATAGGGAGATGGATGGTCACATACTGGGC-3’. Digoxygenin (DIG)-(18) labeled RNA probes were obtained and Northern Blot analyses carried out as previously described (18). Rbr1 transcription was quantified by RT-qPCR in three biological replicates with three technical replicates each using above mentioned primers. The codY gene with forward primer 5’-ATTAGGAACATTGGTACTTTCAAGAT-3’ and reverse primer 5’-TTGAACTACAGCTTTCTTTCTCATT-3’ served as reference. cDNA synthesis and qPCR was performed as described elsewhere (20). The qPCR reactions were performed on a qTOWER 2.2 quantitative PCR thermocycler (Analytik Jena). Quantitative data analysis was based on the Pfaffl method (21).
Overexpression and purification of PerR from C. difficile 630 and C. difficile 630Δerm
Slightly modified guidelines for protein overproduction were followed as previously reported (22). Firstly, synthesized gene variants (ThermoFisher, Darmstadt, Germany) were introduced into the pGEX6P1 allowing for GST-mediated affinity purification and subsequent tag excision. E. coli BL21 cells harboring recombinant genes were induced (0.1 mM IPTG) at an OD600nm of 0.5, grown aerobically for 4 h and shifted to anaerobiosis for 2h.
Electrophoretic mobility shift assay (EMSA)
Shift assays were conducted as previously specified (23) with minor variations. The 220 bp rbr1 promoter region was amplified via PCR using forward primer 5’-TTGCAATAGGTATAGCGACAAG-3’ and reverse primer 5’-TGCAATAGGTATAGCGACAAG-3’. The EMSA was performed under anaerobic conditions.
Legend of Dataset S1
Dataset S1
Sequence alignment of Fur family proteins of different bacteria
Sequences of over 900 proteins of the Fur family were aligned using a PSI-Blast via Phyre2 (13). Conserved amino acid residues are indicated at the end of the table.
Acknowledgments
This work was funded by the German Research Foundation (231396381/GRK1947), the „Ministerium für Bildung, Wissenschaft und Kultur Mecklenburg-Vorpommern” (UG16001), the Federal State of Lower Saxony, Niedersächsisches Vorab CDiff and CDInfect projects (VWZN2889/3215/3266).
Footnotes
Competing Interest Statement: The authors declare no competing interests.
References




