

Abstract
A key step in anaerobic nitrate respiration is the reduction of nitrite to nitric oxide, which is catalysed by cd1 nitrite reductase NirS in e.g. the gram-negative opportunistic pathogen Pseudomonas aeruginosa. Each subunit of this homodimeric enzyme consists of a cytochrome c domain and an eight-bladed β-propeller that binds the uncommon isobacteriochlorin heme d1 as an essential part of its active site. Although NirS is mechanistically and structurally well studied, the focus of previous studies has been on the active, heme d1-bound form. The heme d1-free form of NirS reported here, representing a premature state of the reductase, adopts an open conformation with the cytochrome c domains moved away from each other with respect to the active enzyme. Further, movement of a loop around W498 seems to be related to a widening of the propeller, allowing easier access to the heme d1 binding side. Finally, a possible link between the open conformation of NirS and flagella formation in P. aeruginosa is discussed.
Synopsis The crystal structure of heme d1-free cd1 nitrite reductase NirS from Pseudomonas aeruginosa has been determined and provides insight into a premature form of the enzyme.
1. Introduction
Denitrification is the stepwise reduction of nitrogen oxides to dinitrogen and is utilized by many bacteria to replace the terminal oxygen-dependent steps of the respiratory chain under anaerobic conditions. A highly regulated step is the reduction of nitrite to nitric oxide, since both of these molecules are toxic to the cell. Many bacteria utilise the cd1 nitrite reductase NirS to catalyse this step, and the NirS enzymes from Pseudomonas aeruginosa (Pa) and Paracoccus pantotrophus (Pp) are functionally and structurally well studied. Both chains of this homodimeric enzyme possesses a c-type cytochrome domain with a covalently attached heme c functioning as electron entry point and a C-terminal eight-bladed β-propeller that utilizes the uncommon bacteriochlorin heme d1 as an active site cofactor (Figure 1a) (Nurizzo et al., 1997; Williams et al., 1997). Both domains interact via an N-terminal arm, which swaps domains in the case of Pa-NirS (Williams et al., 1997; Nurizzo et al., 1997). Compared to other tetrapyrroles such as heme b, high affinity for anionic molecules such as nitrite and low affinity for nitric oxide has been observed for the ferrous isobacteriochlorine heme d1. This is rooted in the unique carbonyl moieties at ring A and B and a double bond in the propionate sidechain at ring C (Chang et al., 1986; Rinaldo et al., 2011; Fujii et al., 2016). During its reduction, nitrite is coordinated by H327 and H369 on top of the ferrous iron of heme d1 (Figure 1b)(Cutruzzola et al., 2001). After dehydration and reduction of the substrate, the tetrapyrrole heme d1 is then reduced via internal electron transfer to allow efficient replacement of the product nitric oxide by another nitrite anion (Rinaldo et al., 2011). In case of Pa-NirS, the intramolecular electron transfer is the rate limiting step and is allosterically controlled by smaller conformational changes within the cytochrome c domain (Farver et al., 2009; Nurizzo et al., 1999, 1998). Interestingly, when Pp-NirS was crystallised under reducing conditions, a ∼60° rotation of the cytochrome c domain around the pseudo eight-fold axis of the β-propeller was observed (Sjögren & Hajdu, 2001). This rotation was also seen in Pa-NirS after mutation of H327 to alanine (Brown et al., 2001).

(a) Chemical structure of heme d1 with characteristic features highlighted in red. (b) Simplified depiction of the Pa-NirS active site during the reduction of nitrite to nitric oxide.
All structural studies of NirS up to now have focused on the heme d1-bound form of the enzyme and have led to a detailed understanding of the reaction mechanism within the denitrification context. Recently however, following the discovery that Pa-NirS forms complexes with the flagella protein FliC and the chaperone DnaK, an enzyme activity-independent scaffolding function of a heme d1-free form of NirS in flagella formation has been proposed (Borrero-de Acuña et al., 2015). This cofactor-less form is also expected to exist in the course of maturation of NirS, where the enzyme has been demonstrated to transiently interact with NirF and NirN, two proteins involved in the biosynthesis of the heme d1 cofactor (Nicke et al., 2013). We have therefore set out to determine the heme d1-free structure of NirS in order to gain more insight into these processes.
2. Materials and methods
2.1. Macromolecule production
NirS with bound dihydro-heme d1 was isolated from Pseudomonas aeruginosa strain RM361 (nirN::tet) (Kawasaki et al., 1997) after cultivation under anaerobic conditions as previously described (Adamczack et al., 2014; Klünemann et al., 2019). Briefly, purification of NirS with bound dihydro-heme d1, the precursor of heme d1 that accumulates in the absence of NirN, was achieved by two subsequent ion exchange chromatographic steps as first described by Parr and colleagues (Parr et al., 1976). The purified protein was dialysed against 10 mM Tris/HCl pH 8 and loaded onto a Q-Sepharose fast flow anion exchange column. After washing for 4 h with dialysis buffer at a flowrate of 1 ml/min the eluted protein lost dihydro-heme d1 but still contained the covalently attached heme c as indicated by UV/Vis absorption band at 410 nm (indicative of heme c) and no absorption at 630 nm (indicative of dihydro-heme d1) during elution. Interestingly, dihydro-heme d1 does not seem to be eluted during the washing procedure but remained bound to the resin, visible as a greenish coloration of the chromatography column. Prior to crystallisation, the protein was subjected to size exclusion chromatography (Superdex 200, GE Healthcare) in a buffer containing 10 mM Tris/HCl pH 8 with 150 mM NaCl.
2.2. Crystallization
Crystallisation was performed by vapour diffusion in sitting drop 96-3 Intelli plates (Art Robbins Instruments) at room temperature. Plates were set up with a with a HoneyBee 961 pipetting robot (Digilab Genomic Solutions), which mixed 200 nl of protein solution with 200 nl of mother liquor and provided a precipitant reservoir of 60 µl. To get structural insight into possible changes in the dihydro-heme d1 bound form, a screen based on previously published crystallisation conditions for NirS (Tegoni et al., 1994) (Brown et al., 2001) was performed. After two days, plate-shaped green crystals were observed with phosphate as the precipitant (1.9 M K2HPO4 and 0.1 M Tris-HCl pH 8). The green colour hinted at the presence of dihydro-heme d1 in these crystals (Figure 2). Surprisingly, under conditions with PEG4000 or PEG6000, red rod-shaped or tetragonal crystals started to grow. The red coloration indicates that dihydro-heme d1 was lost as a cofactor. As these crystals only diffracted x-ray to 4 Å, sparse matrix screening with the commercially available crystallization suites JCSG+ (Quiagen) and Morpheus (Molecular Dimensions) was used to identify conditions better suited for diffraction experiments. The best-diffracting crystals for heme d1-free NirS occurred with 0.1 M phosphate/citrate pH 4.2 and 40% (v/v) PEG300, with dihydro-heme d1 removed from NirS by the chromatographic procedure described above prior to crystallization experiment. Crystals were flash frozen in liquid nitrogen after cryo-protection with 10% (v/v) (R,R)-2,3-butandiol.

Photographs of representative crystals grown from NirS isolated with (green frame) or without (red frame) bound dihydro-heme d1. The data sets used in this study were collected from crystallization conditions marked with an asterisk (*)
2.3. Data collection and processing
3600 diffraction images, each with an oscillation angle of 0.1°, were collected at beamline P11 at PETRA III (DESY, Hamburg, Germany) on a PILATUS 6M fast detector. Images were processed utilizing the autoPROC pipeline (Vonrhein et al., 2011) executing XDS (Kabsch, 2010), Pointless (Evans, 2011) and Aimless (Evans & Murshudov, 2013). In case of NirS with bound dihydro-heme d1 the crystals diffracted anisotropically, which was accounted for by using STARANISO (Tickle et al., 2018) within autoPROC.
Values for the outer shell are given in parentheses.
2.4. Structure solution and refinement
The crystal structure of heme d1-free NirS was determined by molecular replacement utilizing Phaser (McCoy et al., 2007) and coordinates of the H327A variant of Pa-NirS (NirSH327A; PDB:1hzu (Brown et al., 2001)), which crystalized in the same space group but with approx. 3 Å longer unit cell axes. The structure of NirS with bound dihydro-heme d1 was phased by Fourier synthesis using phases from a published structure of Pa-NirS with bound heme d1 (PDB: 1nir (Nurizzo et al., 1997)). To remove phase bias, both initial models were subjected to ten cycles of refinement with enabled simulated annealing in phenix.refine (Afonine et al., 2012). The final models were constructed by several cycles of manual optimization in Coot (Emsley & Cowtan, 2004) and computational refinement including TLS refinement. Depictions of the final models were prepared with PyMol (Schrödinger, 2015).
Values for the outer shell are given in parentheses.
3. Results
In previous work with dihydro-heme d1 dehydrogenase NirN, we isolated the cd1 nitrite reductase NirS with bound dihydro-heme d1, the final intermediate of heme d1 biosynthesis, which differs from heme d1 by missing a double bond in the propionate chain attached to ring D in comparison to heme d1 itself (Klünemann et al., 2019). It has previously been shown that NirS can utilize both cofactors but is less active with dihydro-heme d1 (Hasegawa et al., 2001; Kawasaki et al., 1997; Adamczack et al., 2014). To determine if the altered activity is caused by structural changes or by inherent properties of dihydro heme d1, NirS with bound dihydro heme d1 was crystalized with conditions published for heme d1-bound NirS (Tegoni et al., 1994; Brown et al., 2001). In this complex, anomalous difference density around the iron centres indicates similar occupancies for dihydro-heme d1 and heme c, the latter being covalently attached to the polypeptide. Comparison with the previously published heme d1-bound structure of NirS reveals no significant differences (Nurizzo et al., 1997)(Cα-rmsd: 0.38 Å; Figure 3+4). This suggests that the reported differences in the activity of NirS loaded with dihydro-heme d1 or heme d1 likely are caused by inherent properties of the cofactor.

Cartoon representation of different conformation observed for cd1 nitrite reductase NirS of P. aeruginosa (heme d1 bound NirS: 1nir (Nurizzo et al., 1997); NirSH327A: 1hzu (Brown et al., 2001)) after superposition of the d1-domains. The N-terminal arm is coloured in red, the linker in blue and the d1-domain in grey. The cytochrome c domain and the tetrapyrroles are coloured individually.
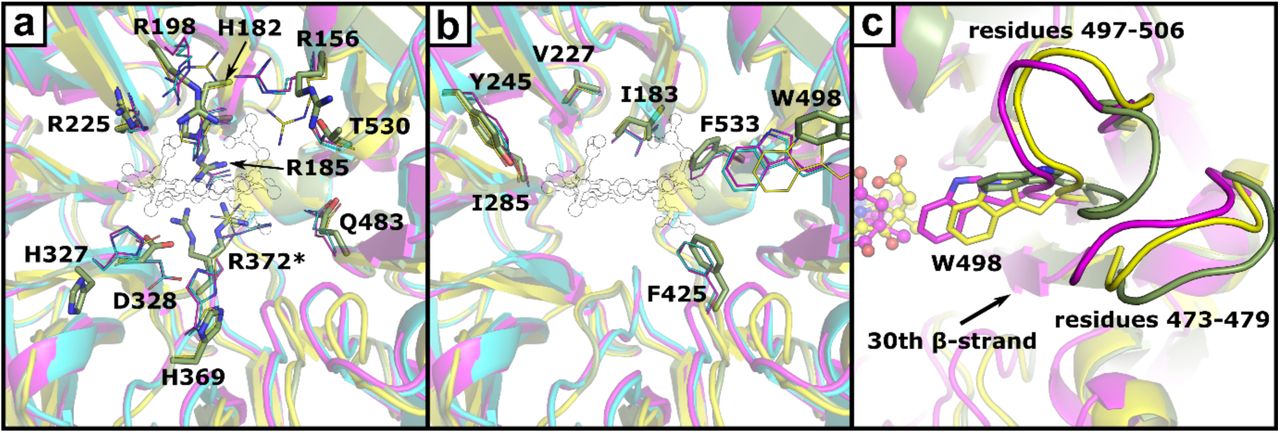
Depiction of hydrophilic (a) and hydrophobic (b) residues in the heme d1 binding pocket. Amino acids are shown as sticks (heme d1-free NirS, green) or lines (heme d1 bound NirS, magenta; NirSH327A, yellow; dihydro-heme d1 bound NirS, cyan). Residues with alternativ conformations are marked with an asterisk (*). The position of heme d1 inside the binding pocket is shown as a thin outline. Panel c depicts the movement of the loops associated with W498 narrowing the binding pocket of heme d1-free NirS after binding a d1-type hemes.
The structure of heme d1-free NirS revealed a similar dimeric structure and domain organization as mature, heme d1-bound NirS (Nurizzo et al., 1997). Both chains have a C-terminal eight-bladed β-propeller, which has previously been termed the d1-domain as it is responsible for binding d1-type tetrapyrroles in NirS and the related proteins NirF and NirN (Figure 3). In NirS, these domains form homodimers by interaction of the 15th β-strand of each subunit, which is consistent with the SEC-data collected during purification (data not shown). The N-terminal domain consists of a cytochrome c fold that is connected via a linker domain to the d1-domain and harbours a covalently attached heme c. Interestingly, the position of the cytochrome c domain differs from the one found in the heme d1 and dihydro-heme d1-bound structures by a rotation of approximately 60° around the pseudo eight-fold axis of the β-propeller. A similar conformation has already been observed for Pa-NirS after amino acid exchange of H327 to alanine or for Pp-NirS crystallised under reducing conditions (Brown et al., 2001; Sjögren & Hajdu, 2001). No density was observed for the N-terminal arm, which resides between the cytochrome c domain and the d1-domain of the opposing subunit in NirS in the closed conformation, indicating that it is flexible in the heme d1-free form of the enzyme.
An overlay of the cytochrome c domain of heme d1-free NirS with other Pa-NirS structures reveals higher similarity with the structure of NirS reduced in crystallo (Cα rmsd: 0.37 Å) as compared to the oxidised form (Cα rmsd: 1.09 Å) (Nurizzo et al., 1998; Nurizzo et al., 1997). The difference is mostly caused by the position of residues 55-64 and consistent with observation previously discussed for NirSH327A (Cα rmsd cytochrome c: 0.35 Å). A similar comparison of the d1-domain reveals a lower Cα rmsd with respect to the H327A variant of Pa-NirS (0.75 Å) than to the oxidised or reduced wildtype protein (1.34 Å and 1.39 Å). NirSH327A has been shown to have a wider β-propeller, allowing better solvent access to heme d1 (Brown et al., 2001). Further, the reported occupancy (0.5) of heme d1 together with the observed change of coloration of these crystals in the course of time, indicative for the loose of heme d1, suggests that the open conformation allows easy access to the ligand binding side in general (Brown et al., 2001). Closer inspection of the heme d1 binding pocket reveals only few distinct changes related to d1-type heme binding (Figure 4). Three arginines that interact with the acetate or propionate sidechains of heme d1 in the heme d1 bound structure (R156, R198, R372) adopt different conformations in the heme d1-free form. Further, H327 and H369, which are both involved in substrate binding and catalysis by NirS, have different positons. Interestingly, H182, which acts as the fifth ligand to the iron cation in the centre of heme d1, shows no conformational changes without bound tetrapyrrole. This stands in contrast to other d1-type heme binding β-propellers of the heme d1 biosynthesis proteins NirN and NirF, where the corresponding histidine rotates with respect to the ligand-free form to coordinate the iron (Klünemann et al., under revision; DOI: 10.1101/2020.01.13.904656) (Klünemann et al., 2019). Another distinctive variation in the d1-domain of heme d1-free and bound NirS is observed for the loop consisting of residues 497-506, which moves upon heme d1 binding to enable W498 to lock the cofactor inside its binding pocket. Interestingly, the neighbouring loop (residues 473-479) also takes a slightly different conformation. This loop is connected to the 30th β-strand, which is inside of the propeller and moved inward upon heme d1 binding, indicating a connection between the propeller widening and the movement of W498 (Fig. 3c).
4. Discussion
It has long been known that the cd1 nitrite reductase NirS adopts an open and a closed conformation, and it has been hypothesized that these states are linked to the catalytic cycle (Brown et al., 2001; Sjögren & Hajdu, 2001). The data presented here, however, suggest that the open conformation rather is related to a native but immature form of the enzyme and is therefore not necessarily associated to the catalytic cycle. This is further supported by the observation that Pp-NirS is unable to undergo a full catalytic cycle in crystallo when crystallised in the open conformation (Sjögren & Hajdu, 2001). In addition, a complex between Pa-NirS in its closed conformation and the nitric oxide reductase (NOR) has been published recently, suggesting an association between NirS and the cytoplasmic membrane that enables substrate channelling (Terasaka et al., 2017). The open conformation would lead to a change of the membrane interface of NirS and therefore cannot support this membrane association, indicating that it is not part of the catalytic cycle.
The open conformation of NirS may, however, have additional physiological importance outside of the denitrification process. It has recently been found that NirS can function as scaffold protein for flagella formation under denitrifying conditions, even in the absence of heme d1 (Borrero-de Acuña et al., 2015). Interestingly, the N-terminal arm and the cytochrome c domain of NirS were shown to be involved in complex formation between NirS, FliC and DnaK in this process (Borrero-de Acuña et al., 2015). Further, kinetic studies with NirS from Pseudomonas stutzeri, which lacks the N-terminal arm, and mutation of a conserved tyrosine in Pa-NirS and Pp-NirS suggest that the N-terminal arm is not required for the enzymatic activity, despite its interaction with heme d1 (Cutruzzolà et al., 1997; Gordon et al., 2003; Wilson et al., 2001). Therefore, the conformational change observed in our study may be envisioned to release the N-terminal arm to allow flagella formation, thereby regulating cell motility. Further investigation will be required to confirm that the conformational changes observed in crystallo can also be found in solution and is associated with distinct functions of NirS in vivo.
Funding information
Deutsche Forschungsgemeinschaft (grant No. 2223/1).
Acknowledgements
We thank the beamline staff at P11 at the PETRAIII synchrotron (Deutsches Elektronensynchrotron DESY, Hamburg, Germany) for letting us use their facilities. This work was supported by grants from the Deutsche Forschungsgemeinschaft (PROCOMPAS graduate school, GRK 2223/1).





{kind=link}
{kind=link}
{kind=link}
{kind=link}