

Abstract
MRSA is Methicillin-resistant Staphylococcus aureus and they are widespread and making trouble in treatment in communities and surgical areas. MRSA have been adapted to antibiotics so that they can block the access of antibiotics physically or chemically deactivate it or modify the precursor of the target. Flavonoids are secondary metabolites which are naturally produced by plant or fungus and they are acting generally as pigment, quorum sensing molecules, antibiotics to other competitive microorganisms. Their natural origins and multiple activities have drawn much attention to be developed as the potential drugs since flavonoids could be a good candidate to overcome antibiotic resistant bacteria. Among various flavonoids, we found out naringenin has antibacterial activity on MRSA and Δagr mutants which are more resistance than MRSA to beta-lactam antibiotics by decreasing biofilm formation dramatically and decreasing the secretion of fatty acid. It also showed high synergetic activity with oxacillin to both antibacterial activity and biofilm inhibition. Considering the number of flavonoids, our experiments expand the possibility of the use of flavonoids to MRSA.
Introduction
MRSA stands for Methicillin-resistant Staphylococcus aureus and they are widespread and troublesome in the communities and hospital area (1). They occasionally cause pneumonia, urinary tract infection, surgical site infection, soft tissue infection and necrotizing pneumonia (2). The long-term overuse of antibiotics to treat them contributed to the multi-drug resistance in Staphylococcus aureus (3). MRSA have been developing their own way for antagonizing or blocking antibiotics. For instances, they have developed to have biofilm, peptidoglycan and membrane barrier to block the access of the antibiotics (4, 5). Additionally, antibiotics have been modifying targets of antibiotics to bypass them and chemical degradation or modifications of antibiotics have been made (6, 7).
Flavonoids are the secondary metabolites produced from plant and fungus and they fulfill various kinds of functions in them. For example, they act as a colorizing agent in plant and are involved in the filtration of UV (8). Additionally, they can be used as messengers as physiological regulators and cell cycle inhibitors in signal cascades in plant or fungus gene (9). Nowadays, flavonoids have drawn attention with their anti-bacterial activities to pathogenic bacteria (10). Since they derived from the natural origins, they seem safer to consumers than the chemically synthesized drugs. Also, they have advantages over antibiotics that they have multiple activities acting on the cells, so it is hard for bacteria to develop resistance to them (11). Thus, many approaches have been carried out to seek natural origin flavonoids having anti-bacterial effect and develop to make them have stronger efficacy (12). For example, catechins can disrupt the bacterial membrane by interacting with membrane and inhibiting enzymes (13). Isovitexin (apigenin-6-glycoside 14), epicatechin, and 5, 7, 5’-trihydroxyflavanol were active against Staphylococcus aureus having anti-biofilm activity (11). Naringenin, quercetin, sinensetin and apigenin have been shown as inhibitors of autoinducer-mediated cell-cell signaling (14). Synthetic 3-arylidene avanones acted as inhibitors of biofilm formation to Staphylococcus aureus and Enterococcus faecalis (15).
In this study, plant derived flavonoids including naringenin, chrysin, daidzein, genistein and apigenin were screened for the MRSA and MRSA mutant with higher resistance than USA300 (LAC). Naringenin decreased biofilm formation and fatty acid secretion was analyzed by GC-MS to see how they are distributed and affecting antibiotic resistance. We found out naringenin could decrease growth of MRSA, MRSA mutant and clinically isolated MRSAs decreasing biofilm formation at the same time. This showed the possibility of flavonoids as an antibacterial material or synergetic molecules with other well-known antibiotics.
Result
Flavonoids compound screening toward MRSA LAC and Δagr mutant
Flavonoids have anti-bacterial activities to MRSA, and they can be simultaneously used with antibiotics for synergetic effect (16). And they have been tested for the MIC test toward pathogenic bacterium and suggested possible mechanisms but there are still detail explanations required to be discussed and the efficacy of the flavonoid are dependent on which bacteria they are applied to (17). Therefore, five flavonoids including naringenin, chrysin, daidzein, genistein and apigenin which have been previously studied to have been effective toward MRSA were selected to be tested for the Δagr mutant with higher resistance. The chemical structures of five plant-derived flavonoids are presented in (Figure 1A). As an initial experiment, 100 μg/ml of flavonoids were applied to the LAC (WT) and Δagr strain w/o or with oxacillin for synergetic effect confirmation by checking the comparative analysis of cell growth and biofilm formation.
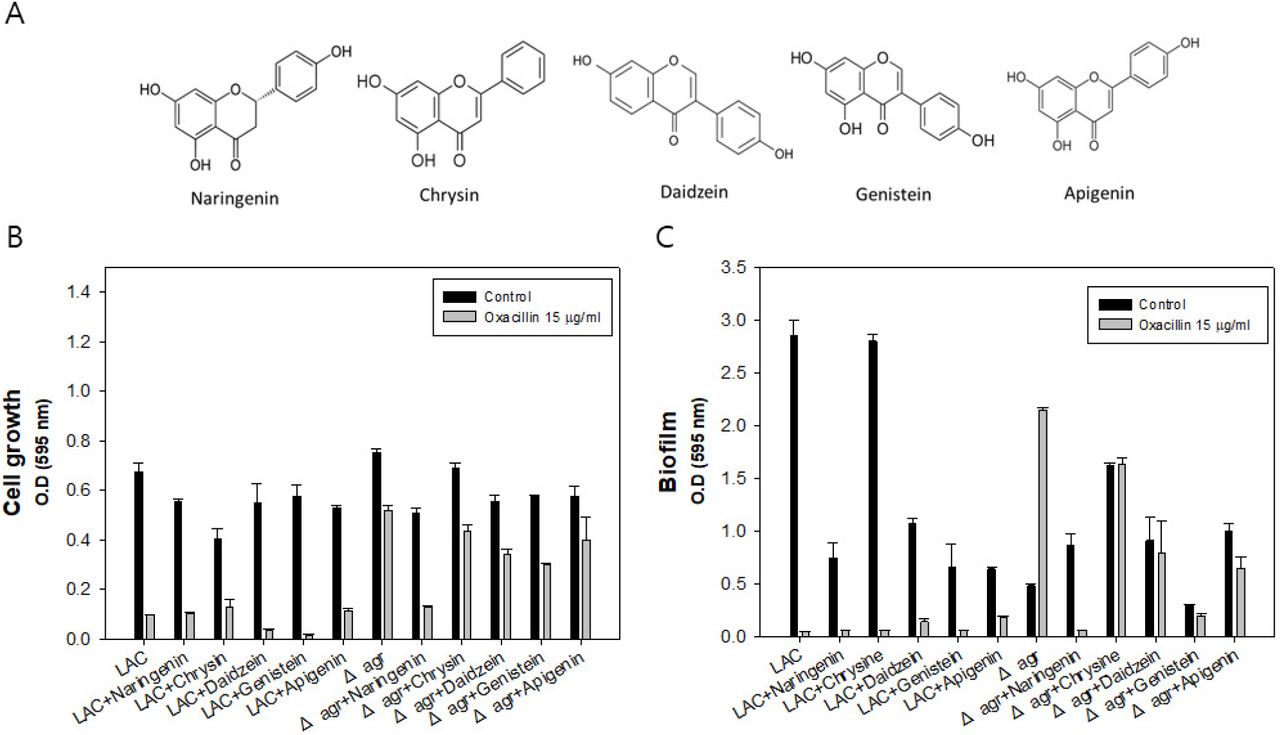
Chemical structures of plant-derived flavonoids, Investigation of five selected flavonoids efficacy to LAC and Δagr strain. A: Cell growth of LAC and Δagr strain with flavonoid w/o oxacillin or with oxacillin addition. B: Biofilm formation of LAC and Δagr strain with flavonoid w/o oxacillin or with oxacillin addition. The error bars represent the standard deviation of three replicates.
As a result, all the flavonoids were effective to LAC and Δagr strain. Among them, the most potent efficacy was shown by naringenin with oxacillin in that the combination has synergistically decreased biofilm and completely inhibited the growth of both LAC and Δagr strain (Figure 1B and 1C). Biofilm thickening is occasionally happening in the MRSA mutant with agr, and biofilm blocks the passage of the antibiotics thereby acquiring higher resistance. Thus, biofilm formation is one of the key factors affecting antibiotic resistance which should be dealt with when it comes to antibiotic treatment. Also, biofilm thickening was discovered especially when oxacillin was treated in Δagr strain. In conclusion, the naringenin came out to be the most effective flavonoid to MRSA inhibiting growth and biofilm formation at this level though still mechanism and efficacy are dose dependent.
Investigation of optimal concentration of naringenin and expression level of biofilm formatting genes
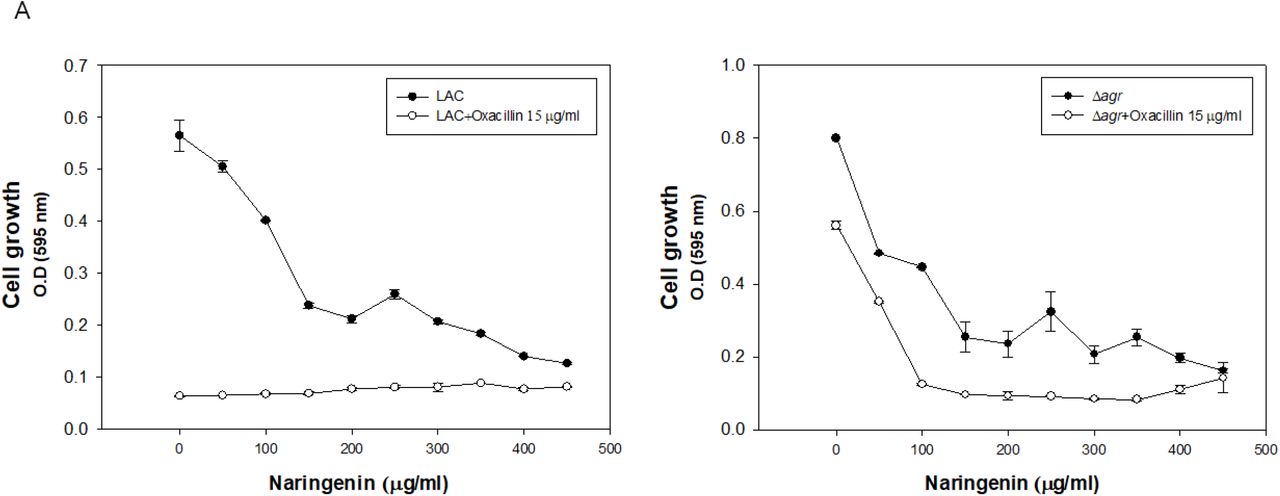
To set up the optimally effective concentration of naringenin to MRSA strains, different concentrations of naringenin were applied to MRSA strains. MIC50 value of the naringenin was similar between two groups so that 200 μg/ml of naringenin was used for further study unless stated otherwise. The problem of Δagr strain is biofilm thickening when they grow since they lack PSMs which have surfactant activity causing lipid shedding and biofilm dispersion (5). Therefore, they have higher resistance to β-lactam anbitiotics. With addition of 200 μg/ml of naringenin with oxacillin, mutant strain was not able to grow, and they produce almost no biofilm (Figure 2).


Investigation of optimal naringenin concentration to LAC and Δagr strain. A: Cell growth of LAC and Δagr strain with naringenin w/o oxacillin or with oxacillin addition. B: Biofilm formation of LAC and Δagr strain with naringenin w/o oxacillin or with oxacillin addition. The error bars represent the standard deviation of three replicates.
To see any effect on mRNA expression, expression levels of icaAD, mecA, nanK were compared to see how they act on increased antibiotic susceptibility (Figure 3). icaAD genes are biofilm formation genes and mecA is a PBP2a protein coding gene increasing resistance of β-lactam anbitiotic. Finally, nanK is an indicator of persister cell formation so that the expression levels of between wild type and mutant groups were compared. In the case of mecA and nanK, there was no change in expression level with naringenin addition. When it comes to biofilm formation, the expression level of icaAD was less with naringenin addition compared to reference. As a result, with naringenin, decreased level of biofilm genes led to the higher susceptibility to oxacillin.

Semi-quantitative PCR of icaAD, mecA and nanK.
Monitoring of synergetic effect by naringenin
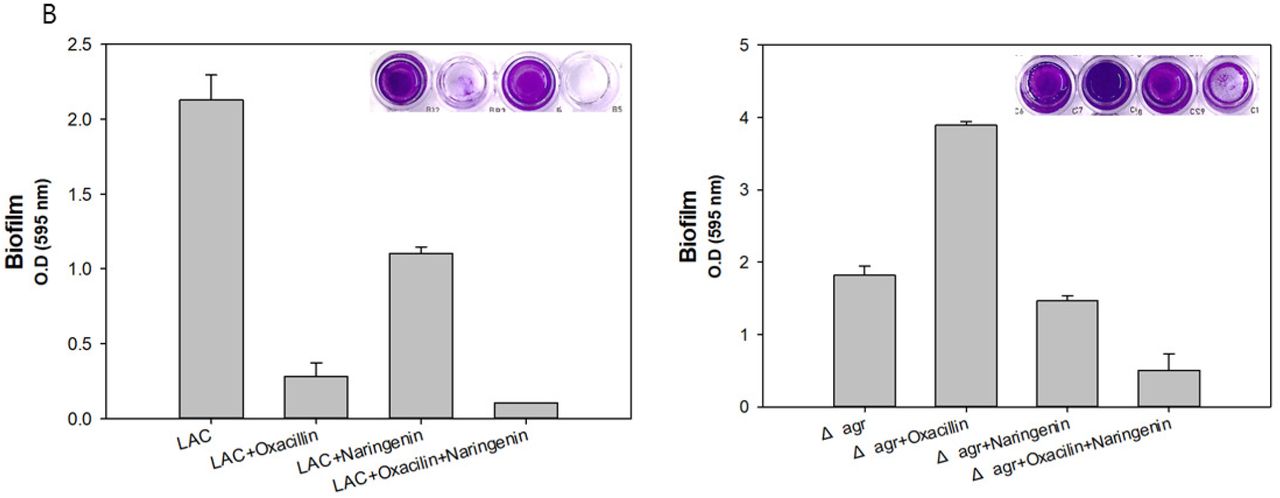
To see the synergetic effect of naringenin, we applied oxacillin, naringenin and combination of two to both LAC and Δagr mutant. In LAC strain, naringenin showed antibacterial activity resulting the decrease of growth and biofilm formation (Figure 4A). In Δagr mutant, the effect of naringenin was critical this is because the expression of icaAD was higher in Δagr mutant than LAC with naringenin treatment. Combination therapy showed higher antibacterial activity than oxacillin alone and its activity on biofilm inhibition was much higher than oxacillin alone (Figure 4B). Considering oxacillin increasing the production of biofilm, the effect of naringenin to decrease the biofilm was quite useful and its combinatorial use to Δagr mutant clearly showed the dramatic decrease of biofilm formation with antibacterial activity. When we monitored biofilm formation with SEM at different time points by treating oxacillin, naringenin and both, we could see the clear differences. As detected by absorbance of growth and biofilm formation, SEM data showed the less formation of biofilm and low and dispersed population of Δagr strains by treating both oxacillin and naringenin (Figure 5A, 5B). As a result, naringenin could work as a sole antibiotic material and anti-biofilm material, but it could be also used as a synergetic molecule with known antibiotic, oxacillin.


Growth and biofilm formation profile of MRSA with higher resistance compared to LAC. A: Cell growth of LAC and Δagr strain with naringenin w/o oxacillin or with oxacillin addition. B: Biofilm formation of LAC and Δagr strain with naringenin w/o oxacillin or with oxacillin addition. The error bars represent the standard deviation of three replicates.


Scanning electron microscopy (SEM) images of with or w/o oxacillin and naringenin addition. A: SEM images of Δagr strains with or w/o oxacillin and naringenin treatment at 24 hr. B: SEM images of Δagr strains with or w/o oxacillin and naringenin treatment at 48 hr.
Changes in fatty acid metabolism and expression level of biofilm formation genes
Naringenin has been known to be inhibit quorum sensing limiting biofilm formation and can reduce fluidity of the bacterial membrane (18). In addition to that, naringenin can target FAS II system which is essential pathway for bacteria survival.
In Δagr mutant there was lower level of secreted fatty acid since fatty acid are known to have anti-bacterial and anti-biofilm activities and external addition of fatty acid increase the antibacterial activity and antibiofilm activity (19). Thus, it is favorable for them to keep lower amount of especially C16 and C18 fatty acids for their own survival. Microenvironment control of MRSA is quite related to control of antibiotic resistance so that fatty acid distribution profile of mutant strains was conducted w/o or with naringenin addition. We performed the analysis of secreted fatty acid at different time points with two LAC and Δagr strains. Preparation of materials were explained in Material and methods section. Surprisingly, with naringenin addition, total amount of fatty acids was decreased in both strains (Figure 6A). Although total amount of fatty acids by cell weight was increased in LAC with naringenin, but there was growth limitation so that it is out of consideration (Figure 6B). Also, total fatty acids production was lower when treated naringenin (Figure S1). Considering flavonoids are the inhibitor of fatty acid synthesis, and secreted fatty acid could affect antibiotic activity, this is quite interesting result and explained the functional mode of naringenin. This have led to the higher effect of oxacillin and lower level of biofilm formation to kill both LAC and Δagr strains.

Analysis of secreted fatty acid w/o or with naringenin in MRSA mutants. The error bars represent the standard deviation of three replicates.
Application of synergetic effect of naringenin and oxacillin to clinically isolated strains
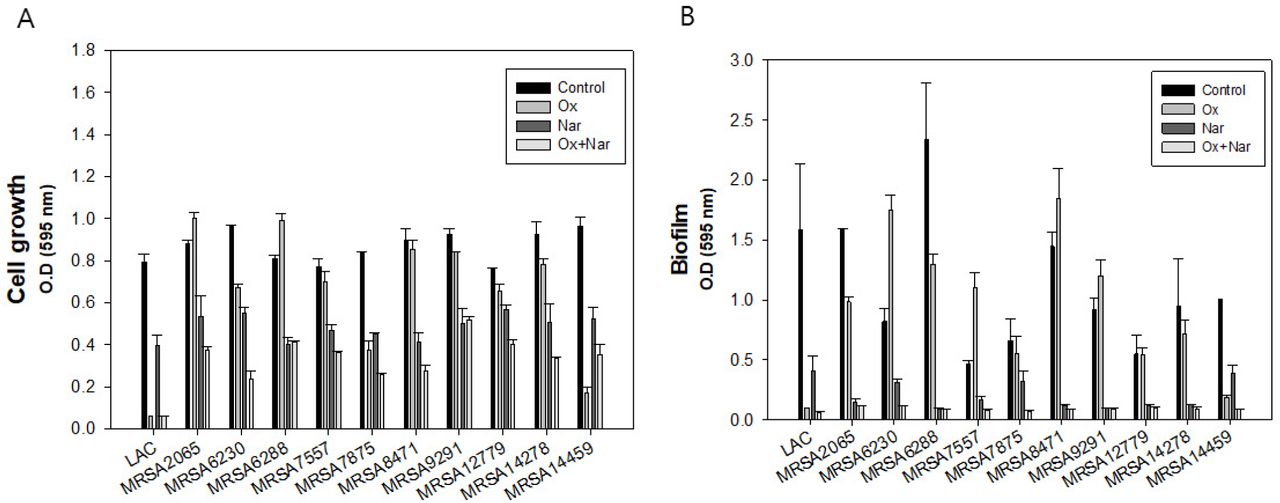
Naringenin and oxacillin had synergetic effect toward LAC and Δagr but still they can be classified as CA-MRSA which has MIC level about >32 μg/ml. Thus, we further explored the effect of combination of naringenin and oxacillin to 10 clinically isolated MRSAs. The concentrations of naringenin and oxacillin were set same for the comparison to CA-MRSA data. Surprisingly, the synergetic effect of the oxacillin and naringenin still remained in the most of clinically isolated strains except MRSA14459 (Figure 7A). Oxacillin alone was able to treat MRSA14459 but not with naringenin combination. In the case of biofilm, all the strains did not produce biofilm when treated with oxacillin and naringenin simultaneously (Figure 7B). To validate the case of MRSA14459, we further investigated the MIC of each strains. However, in this time, MRSA14459 was able to tolerate the antibacterial effect of oxacillin even over 200 μg/ml. Except MRSA 6230 of which MIC is about 150 μg/ml, MIC level of oxacillin of other strains were over 200 μg/ml (Table 1). Inequal effect of oxacillin might be from induction of persister formation or from unknown mechanism (data not shown). In addition, it was surprising that even MRSA6230 and MRSA14459 have higher resistance to oxacillin since they are SCCmec type IV which is a characteristic of CA-MRSA. However, synergetic effect of oxacillin and naringenin still exist even at the low concentrations of oxacillin for both strains. Therefore, utilization of naringenin and oxacillin together is an effective way to treat most of the MRSAs though the range of concentration should be increased depending on strains for optimal combination therapy.
Types of clinically isolated MRSAs and Oxacillin MIC for each strains.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Application of synergetic effect of naringenin and oxacillin to clinically isolated strains. A: Cell growth of LAC and clinical strains with oxacillin and naringenin B: Biofilm formation of LAC and clinical strains with oxacillin and naringenin. The error bars represent the standard deviation of three replicates.
Discussion
MRSA is notorious for its widespread existence in surgical area and resistance toward antibiotic treatment (20). The overuse of antibiotics for long term had led to the findings of new anti-bacterial agent such as plant derived flavonoids (21). To be more specific, flavonoids are safe as they originated from natural origin and they can be modified to have more efficacy (11). Flavonoids are secondary metabolites which are naturally produced by plant or fungus and they are acting generally as pigment, quorum sensing molecules, antibiotics to other competitive microorganisms (22). Flavonoids have drawn attention from their natural origin and multiple activities which is making bacteria hard to become resistant to flavonoids. For example, they can block respiratory chain, fatty acid synthesis, gyrase activity, quorum sensing molecules and increase membrane permeability etc. (11). However, its functional mechanism on antibacterial activities are not well known. The use of naringenin affected not only distribution of fatty acid in the cell or supernatant but also reducing the expression of biofilm formation genes causing change in biofilm formation and antibiotic sensitivity. Also, cell population became more dispersed when combination therapy was used. As β-lactam antibiotics widely were used, there came out many MRSA strains with much higher or multi-drug resistance. Therefore, there were not many ways of control it but dose dependent treatment of antibiotics. In that point, combinatorial use of flavonoids with antibiotics could be one way to overcome those difficulties especially with higher resistance strains.
Materials and Methods
Bacterial strains, media, and culture conditions
For cell preparation, the wild-type strain Staphylococcus aureus USA300 (LAC, ATCC® BAA1756) (23) and the mutant strain Δagr (24) were cultured in tryptic soybean broth (TSB) agar and/or liquid broth. For pre-culture, a single colony of the strain from a TSB agar plate was used to inoculate 5 mL of TSB medium. Then, 1% (v/v) of the cell culture suspension was inoculated in a 96-well plate for the antibiotic resistance test and the cell cultivation was conducted overnight in incubator at 37 °C without shaking unless stated otherwise.
Antibiotic and phytochemical
Oxacillin and a total of 5 selected phytochemical powders from flavonoid group i.e., naringenin, chrysin, daidzein, genistein, apigenin were commercially purchased from Sigma-Aldrich (St. Louis, Minneapolis, USA). Stock solutions of these agents were prepared in sterile dimethyl-sulphoxide (DMSO) solvent to various concentrations.
Analysis of cell growth and biofilm formation
The cell growth was measured in terms of cell density using a 96-well microplate reader (TECAN, Switzerland). Biofilm formation was analyzed crystal violet staining using the protocols (25). Briefly, supernatant was discarded, and biofilm was fixed with methanol and air dried. Then, biofilm was stained with 200μl of 0.2 % crystal violet solution for 5 min. Next step was to discard crystal violet and wash with distilled water and air-dry step. Finally, biofilm was analyzed at 595 nm using 96-well microplate reader.
Reverse transcription for cDNA synthesis and semi-quantitative real time PCR
Pre-culture was conducted using 5 ml of TSB with a single colony from agar plate in shaking incubator at 37 °C and 200 rpm during overnight. Cell cultivations were carried out using 5ml of TSB with 1 % inoculum in shaking incubator at 37 °C for 24 h to extract total RNA w/o naringenin or with naringenin addition. Cells were harvested using centrifuge at 3500 rpm for 20 min. Then, total RNA was prepared using TRIzol™ Reagent and reverse transcription was performed with Superscript IV Reverse Transcriptase (Invitrogen Co., Carlsbad, CA) to generate the cDNA following the instructions manuals. Primer design was done using Primer express software v3.0.1 from Thermo Fisher Scientific (Waltham, MA, USA), and these primers can generate 150 bp PCR product for the comparison of gene expressions. Before semi-quantitative PCR, cycle number was optimized to set up the saturated gene expression level of gyrB (endogenous control) for each template. After optimization, 25 cycle came out to be the optimal cycle number and further comparative analysis of gene expression became possible. Then, Semi-quantitative PCR was conducted using LA taq with GC buffer I (Takara medical co. ltd) using the methods in manual.
Fatty acid analysis
Gas chromatography-mass spectrometry (GC-MS) was used for the detection and quantification of total fatty acids and supernatant fatty acids, according to a previously described method with slight modification (26). For total fatty acids analysis, cell cultivation was conducted using 5ml of TSB with 1 % inoculum in shaking incubator at 37 °C and 200 rpm. Then, cell was collected at 24 h and 48 h followed by centrifugation at 3500 rpm for 20 min and washed twice with Milli Q water. Harvested cell was freeze-dried for further methanolysis to analyze total fatty acids. For the analysis of supernatant fatty acids in the medium, spent culture supernatant was incubated in shaking incubator for 2 h at 37 °C and 200 rpm after adding 5 ml of methanol and chloroform to extract fatty acids. The chloroform phase was collected and slowly evaporated under compressed N2 at 50 °C in heating block. Fatty acids were re-solubilized with 1 ml of chloroform for further steps. For methanolysis of fatty acids, approximately 10 mg of freeze-dried cells were weighed and placed in Teflon-stoppered glass vials, and then 1 mL chloroform and 1 mL methanol/H2SO4 (85:15 v/v %) were added to the vials. After incubation at 100°C for 2 h, the vials were cooled to room temperature, and then incubated on ice for 10 min. After adding 1 mL of ice-cold water, the samples were thoroughly mixed by vortexing for 1 min and then centrifuged at 3500 rpm. The organic phases (bottom of the vials) were extracted by a pipette and transferred to clean borosilicate glass tubes containing Na2SO4. GC-MS was then performed with a Perkin Elmer Clarus 500 gas chromatograph that was connected to a Clarus 5Q8S mass spectrometer at 70 eV (m/z 50-550; source at 230 °C and quadruple at 150 °C) in EI mode with an Elite 5 MS capillary column (30 m × 0.32 mm × 0.25 μm film thickness; J&W Scientific, USA). Helium was used as the carrier gas at a flow rate of 1.0 mL/min. The inlet temperature was maintained at 300 °C, and the oven was programmed to start at 150 °C for 2 min before increasing to 300 °C at a rate of 4 °C/min, and the temperature was maintained for 20 min. The injection volume was 1 μL, with a split ratio of 50:1.
The structural assignments were based on interpretation of the mass spectrometric fragmentation and confirmed by comparison with the retention times and fragmentation patterns of the authentic compounds along with spectral data obtained from the online libraries of Wiley (http://www.palisade.com) and NIST (http://www.nist.gov). The internal standard was 1 μL of methyl heneicosanoate (10 mg/mL) and Bacterial acid methyl ester (BAME) mix (Merck-Millipore, Burlington, MA, USA) was used to identify the each peak of fatty acids and analytical standards for each fatty acid were used for quantification.
Acknowledgements
This paper was supported by Konkuk University in 2019.
References




