

Abstract
A critical component controlling bacterial virulence is the delivery of pathogen effectors into plant cells during infection. Effectors alter host metabolism and immunity for pathogen benefit. Multiple effectors are phosphorylated by host kinases, and this posttranslational modification is important for their activity. We sought to identify host kinases involved in effector phosphorylation. Multiple effector phosphorylated residues matched the proposed consensus phosphorylation motif of the plant calcium-dependent protein kinase (CDPK) and Snf1-related kinase (SnRK) superfamily. The conserved Pseudomonas effector AvrPtoB acts as an E3 ubiquitin ligase and promotes bacterial virulence. We identified a member of the Arabidopsis SnRK family, SnRK2.8, that associated with AvrPtoB in yeast and in planta. SnRK2.8 was required for AvrPtoB virulence functions, including facilitating bacterial colonization, suppression of callose deposition, and targeting the plant defense regulator NPR1 and flagellin receptor FLS2. Mass spectrometry revealed AvrPtoB was phosphorylated at multiple serine residues in planta, with S258 phosphorylation reduced in the snrk2.8 knockout. AvrPtoB phospho-null mutants exhibited compromised virulence functions and were unable to suppress NPR1 accumulation, FLS2 accumulation, or inhibit FLS2-BAK1 complex formation upon flagellin perception. These data identify a conserved plant kinase utilized by a pathogen effector to promote disease.
Introduction
Plants are exposed to diverse pathogens and rely on both passive and active defenses in order to restrict infection. Passive plant defenses include a waxy cuticle, pre-formed antimicrobial compounds, and the cell wall (Gu et al., 2017). Inducible defenses are often triggered by membrane-localized pattern recognition receptors (PRRs) as well as intracellular nucleotide-binding site leucine-rich repeat proteins (NLRs) (Boutrot and Zipfel, 2017; Lolle et al., 2020). Common plant immune responses after active pathogen perception include the production of reactive oxygen species (ROS), callose deposition, activation of mitogen-activated protein kinases, and global transcriptional reprogramming towards defense (Bigeard et al., 2015; Peng et al., 2018).
To colonize their hosts, pathogens rely on the ability to secrete effector proteins. Characterized effectors act to suppress plant immune responses, alter host developmental processes, and affect host metabolism to promote pathogen infection (Toruño et al., 2015). Bacterial pathogens have the most well-characterized effector repertoires. For example, the Pseudomonas syringae effectors AvrPto and HopAO1 directly interact with the cytoplasmic domains of the PRRs FLAGELLIN-SENSING 2 (FLS2) and ELONGATION FACTOR-Tu RECEPTOR (EFR), and inhibit PRR kinase activity (Xiang et al., 2008; Macho et al., 2014). Effectors are also capable of manipulating plant hormone processes. The P. syringae effector HopX1 and its Ralstonia solanacearum homolog RipE1 are cysteine proteases that degrade multiple JASMONATE-ZIM DOMAIN (JAZ) transcriptional repressors to activate jasmonic acid signaling, suppress plant salicylic acid (SA)-mediated defense responses, and open stomatal apertures (Gimenez-Ibanez et al., 2014; Nakano and Mukaihara, 2019). Multiple Xanthomonas oryzae pv. oryzae transcriptional activator-like effectors, PthXo1, TalC, AvrXa7, PthXo3, and Tal5, induce the expression of sucrose transporter genes OsSWEET11 or OsSWEET14, which could alter metabolism or nutrient acquisition (Streubel et al., 2013; Zhou et al., 2016; Tran et al., 2018). The diverse functions of pathogen effectors make them excellent molecular probes to investigate plant biological processes.
Despite the collective importance of effectors for pathogen virulence, higher-order effector deletions are normally required to observe strong effects. Two P. syringae effectors, AvrPto and AvrPtoB, are conserved in many P. syringae pathovars and are collectively required for pathogen virulence and bacterial growth in planta (Lin and Martin, 2005). AvrPto and AvrPtoB are partially redundant. For example, in Arabidopsis both AvrPto and AvrPtoB associate with the flagellin receptor FLS2 and interfere with downstream defense signaling (Shan et al., 2008). In tomato, both AvrPto and AvrPtoB interact with the Pto kinase, leading to recognition by the NLR Prf (Kim et al., 2002; Xiao et al., 2007a). However, compared with AvrPto, AvrPtoB has a more complex structure. The N-terminus of AvrPtoB comprises multiple target-binding domains and its C-terminus is a U-box type E3 ubiquitin ligase, which mediates the degradation of target proteins through the host proteasome (Abramovitch et al., 2006). AvrPtoB associates with both FLS2 and its co-receptor BAK1, disrupts flagellin 22 (flg22) induced FLS2-BAK1 complex formation, and mediates the degradation of FLS2 (Göhre et al., 2008; Shan et al., 2008). NPR1 (NON-EXPRESSER OF PR GENES 1) is an essential regulator of salicylic acid (SA)-dependent defense gene expression (Withers and Dong, 2016; Chen et al., 2019). AvrPtoB associates with NPR1 and mediates the degradation of NPR1 in the presence of SA (Chen et al., 2017). In addition, AvrPtoB also targets the protein kinase Fen, the chitin receptor CERK1, and the exocyst complex protein EXO70B1 (Rosebrock et al., 2007; Gimenez-Ibanez et al., 2009; Wang et al., 2019). AvrPtoB’s diverse targets demonstrate its prominent role in P. syringae infection.
Although effectors are produced in the pathogen, they are activated upon delivery and function inside their host. For instance, the P. syringae effector protease AvrRpt2 requires the plant protein folding catalyst and chaperone cyclophilin for activation (Coaker et al., 2005). Several pathogen effectors target plant nuclei and rely upon host importin-α for nuclear translocation, including the Xanthomonas bacterial effector AvrBs3 and the Hyaloperonospora oomycete effector HaRxL106 (Szurek et al., 2001; Bai et al., 2009; Schornack et al., 2013; Wirthmueller et al., 2014). Collectively, these studies highlight diverse plant proteins that are required for mediating the activation, localization, and virulence activities of pathogen effectors.
Pathogen effectors also rely upon host phosphorylation to promote their virulence activity (Xing et al., 2002; Bhattacharjee et al., 2015). Truncated AvrPtoB1-307 is phosphorylated in different plants including tomato, Nicotiana benthamiana, and Arabidopsis thaliana. The substitution of the serine 258 phosphorylated residue on AvrPtoB1-307 to alanine results in significantly attenuated virulence on susceptible tomato genotypes (Xiao et al., 2007a). The P. syringae effectors AvrPto, AvrB, and HopQ1 as well as the Rhizobium effectors NopL, NopP, and the cyst nematode effector 10A07 also target and recruit host kinases to promote their virulence (Desveaux et al., 2007; Li et al., 2013; Hewezi et al., 2015; Tahir et al., 2019). With the exception of the nematode effector 10A07, the host kinases utilized by these pathogen effectors remain unknown.
In this study, we investigated the identity of the host kinase that phosphorylates AvrPtoB. SnRK2.8, a member of sucrose non-fermenting-1(SNF1)-related kinase (SnRK) family in Arabidopsis, is involved in NPR1 mediated systemic acquired resistance and several abiotic stress signaling pathways (Umezawa et al., 2004; Fujii et al., 2011; Lee et al., 2015). Here, we show that SnRK2.8 interacts with and phosphorylates AvrPtoB on three conserved serine residues in planta. AvrPtoB requires these three phosphorylated residues as well as SnRK2.8 for virulence. The snrk2.8 knockout and AvrPtoB phospho-null mutations blocked AvrPtoB virulence activities, including the ability to inhibit NPR1 and FLS2 protein accumulation as well as disrupt ligand-induced FLS2-BAK1 complex formation. Taken together, our results identify a conserved plant kinase recruited by a core P. syringae effector to promote virulence.
Results
The Pseudomonas syringae effector AvrPtoB interacts with the plant kinase SnRK2.8
To investigate which plant kinases may be capable of phosphorylating pathogen effectors, we analyzed the sequence of previously identified effector phosphorylation sites (Supplemental Table 1). Interestingly, most effector phosphorylation sites, including AvrPtoB’s phosphorylated residues, matched the proposed consensus phosphorylation motif of the sucrose non-fermenting-1 (SNF1)-related kinases (SnRKs) and calcium-dependent protein kinases (CDPKs) (R-X(2-3)-S/T or S/T-X(1-2)-R) (Klimecka and Muszyńska, 2007; Vlad et al., 2008) (Supplemental Figure 1). SnRK-CDPK family is conserved in plants and involved in a range of metabolic and stress signaling pathways, such as carbohydrate biosynthesis, abscisic acid (ABA)-induced signaling, salinity tolerance, cold stress, and response to pathogen infection (Coello et al., 2011; Hulsmans et al., 2016). Transcriptional profiling data from the BAR Expression Angler revealed that multiple SnRK members are induced upon activation of plant defense, including application of SA, the immunogenic flagellin epitope flg22, or infection with the P. syringae type III secretion mutant △hrcC (Boudsocq et al., 2010; Gao et al., 2013) (Figure 1A).

(A) Phylogeny of the Arabidopsis SnRK family and transcript expression of SnRK members after immune activation and pathogen infection. Phylogeny was determined by maximum likelihood method (1000 bootstrap replicates). The heat map shows fold change in leaf transcript expression one-hour post-treatment with SA, one hour post-treatment with flg22, and six hours post-infiltration with Pseudomonas syringae pv. tomato strain hrcC. The bar graph represents the SnRK expression in leaf tissue. Data were obtained from BAR Expression Angler. Blue dots represent the SnRK members that were cloned for yeast-two hybrid. hrcC = P. syringae pv. tomato DC3000 hrcC mutant defective in Type III secretion system.
(B, C) Yeast two-hybrid assay of AvrPtoB and SnRK-CDPKs. AvrPtoB was co-expressed with Pto, SnRK3.1, SnRK1.1, SnRK2.6, SnRK2.8, as well as a C-terminal deletion of CPK4 (CPK4△C) and CPK5 (CPK5△C) in yeast. Empty vector (EV) and AD-mCherry were included as negative controls. Colony growth on SD -Leu/-Trp media confirms the presence of AD and BD vectors. Growth on SD -Leu/-Trp/-His media indicates protein-protein interaction. AD = activation domain vector, BD = binding domain vector.
(D, E) Co-immunoprecipitation (Co-IP) of AvrPtoB-GFP with SnRK/CPK△C kinase dead (KD)-HA variants in N. benthamiana. AvrPtoB-GFP under control of a dexamethasone (Dex)-inducible promoter was co-expressed with 35S::SnRKs-KD-HA and 35S::CPKs△C-KD-HA in N. benthamiana using Agrobacterium-mediated transient expression. Expression of AvrPtoB-GFP was induced by 15 μM DEX for three hours at 24 hours post-infiltration. Protein extracts were subjected to anti-GFP immunoprecipitation (IP). The IP and input proteins were immunoblotted with anti-HA and anti-GFP antibodies.
Next, we analyzed the ability of diverse SnRK/CDPK members to interact with AvrPtoB by yeast two-hybrid. The plant protein kinase Pto was used as a positive control (Figure 1B). We analyzed 32 SnRKs for the ability to interact with AvrPtoB by yeast-two hybrid (Supplemental Figure 2). SnRK1.1, SnRK2.6, and SnRK2.8 specifically interacted with AvrPtoB (Figure 1B and C). Some CDPK subgroup Ⅰ members are involved in flg22 signaling and NLR immune responses, so we cloned these CDPK members (CPK2, 4, 5, 6, 11 and 26) and three other members from subgroups which can induce defense gene expression after flg22 application (Boudsocq et al., 2010; Gao et al., 2013). Auto-active CDPKs were generated by deleting their C-terminal Ca2+ regulatory and auto-inhibitory domains (CPKs△C) for yeast-two hybrid screening (Klimecka and Muszyńska, 2007). CPK4△C and CPK5△C specifically interacted with AvrPtoB (Figure 1B, C and Supplemental Figure 2). The expression of AvrPtoB, SnRKs, and CDPKs in yeast were verified by western blotting (Supplemental Figure 3). AvrPtoB, SnRKs, and CPKs△C did not induce auto-activity in yeast (Figure 1C).
To validate the association of AvrPtoB with the SnRK/ CDPK members identified by yeast-two hybrid, we performed co-immunoprecipitation (Co-IP) in Nicotiana benthamiana. To enhance the transient interaction between kinase and substrate, kinase-dead variants (KD) of five SnRK/CPK△C members were generated by mutating the lysine residue (K) in their ATP binding pocket to alanine (A). AvrPtoB-GFP under the control of a dexamethasone (Dex)-inducible promoter was co-expressed with each 35S:: SnRKs-KD-HA/CPKs△C-KD-HA in N. benthamiana and immunoprecipitation (IP) was performed with anti-GFP agarose beads. Immunoblotting results demonstrated that SnRK1.1, SnRK2.6, CPK4△CA, and CPK5△C weakly associated with AvrPtoB (Figure 1D and E). Only SnRK2.8 strongly associated with AvrPtoB (Figure 1D and E). The strong association of SnRK2.8 with AvrPtoB in yeast and in planta suggests that SnRK2.8 may play an important role in AvrPtoB phosphorylation.
AvrPtoB is phosphorylated by SnRK2.8
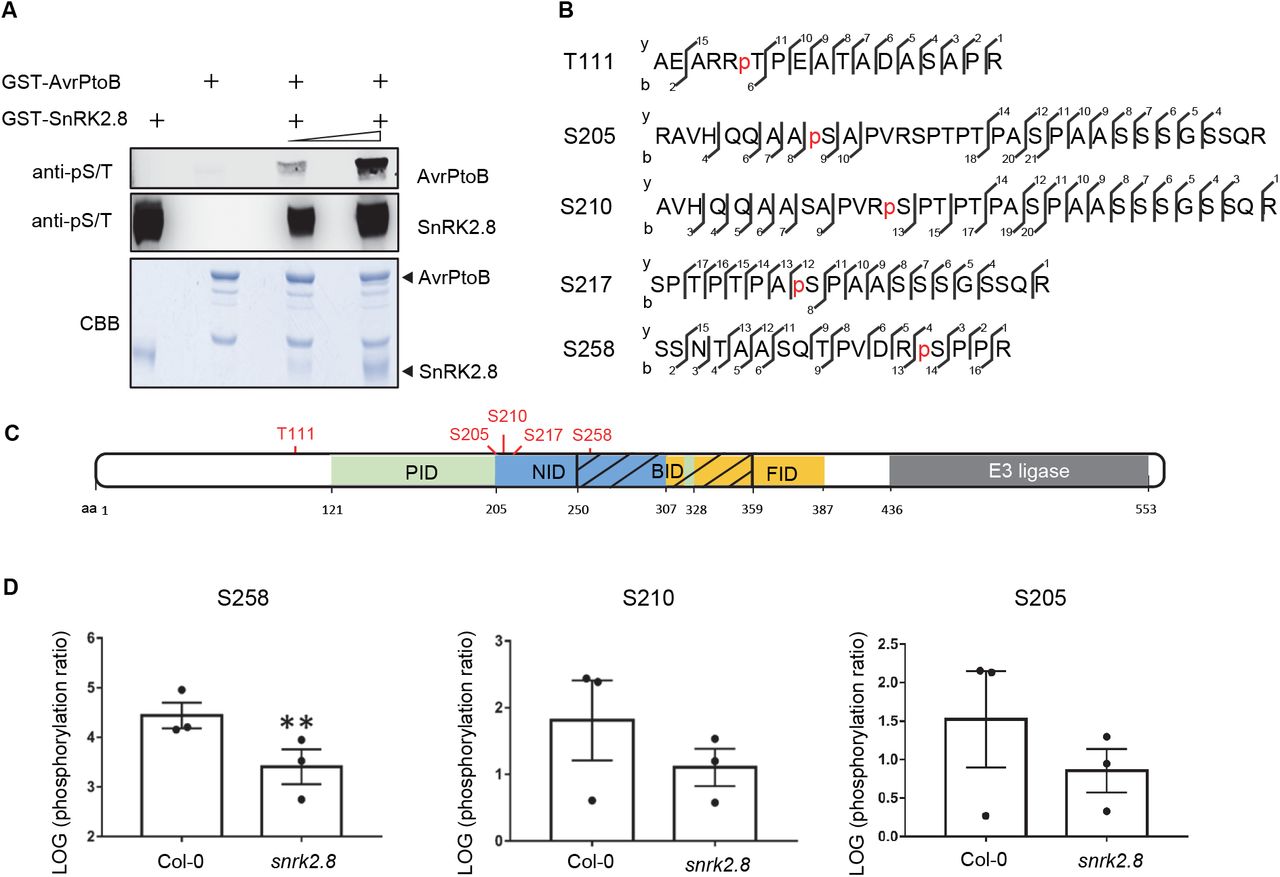
To investigate whether SnRK2.8 phosphorylates AvrPtoB, we tested the phosphorylation by SnRK2.8 in vitro. Recombinant GST-AvrPtoB and GST-SnRK2.8 proteins were purified from E. coli, subjected to kinase activity assays, and protein phosphorylation was detected by immunoblot with antibodies recognizing pSer/pThr residues. Purified SnRK2.8 is an active kinase and exhibits strong auto-phosphorylation (Figure 2A). No phosphorylation was detected in the sample only containing purified AvrPtoB. However, after incubation with SnRK2.8, AvrPtoB phosphorylation was detected in SnRK2.8 dose-dependent manner (Figure 2A). These results indicate that SnRK2.8 is able to phosphorylate AvrPtoB in vitro.
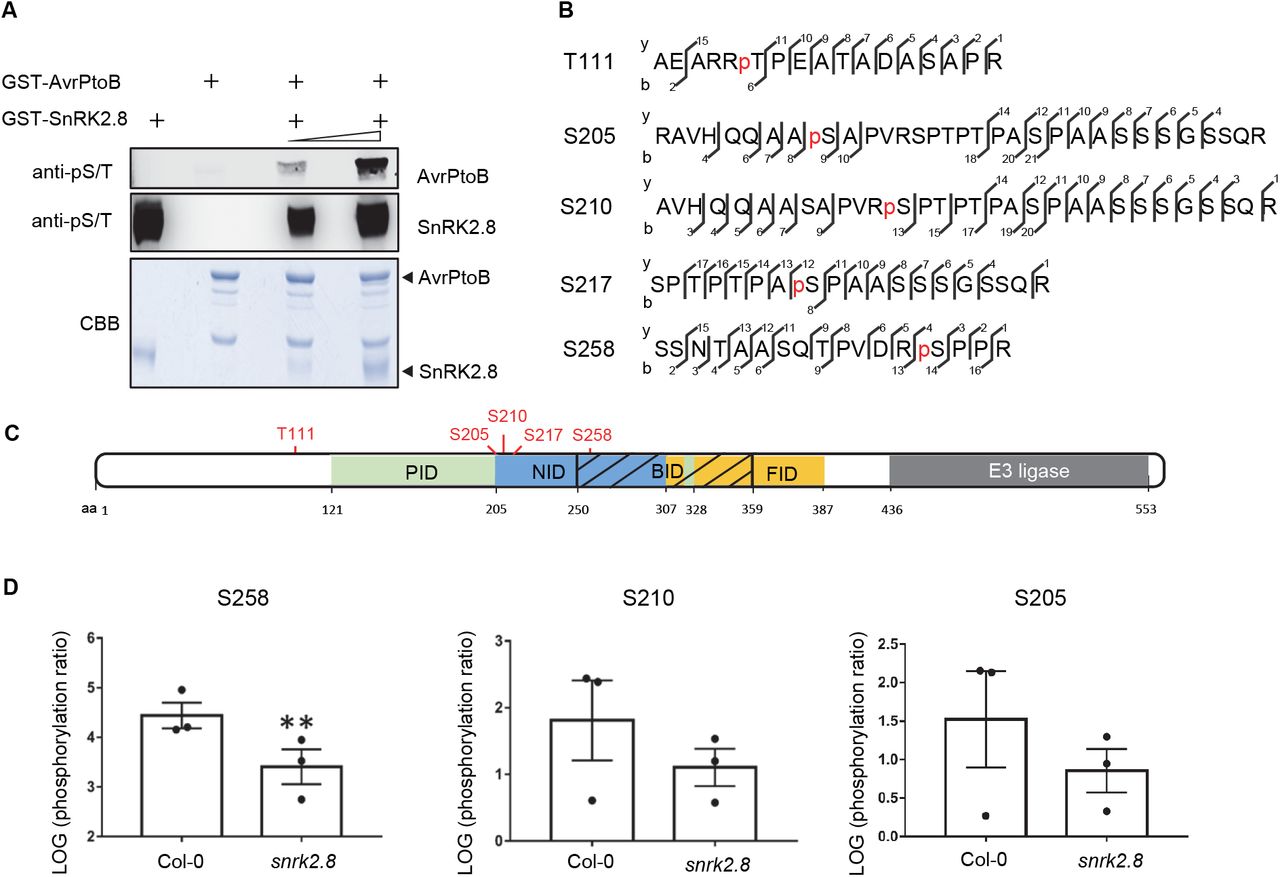
(A) In vitro kinase activity assay of AvrPtoB and SnRK2.8. Recombinant GST-AvrPtoB and GST-SnRK2.8 proteins were purified from E. coli and incubated with ATP in kinase buffer for 30 min. Phosphorylation was detected by anti-pSer/Thr (anti-pS/T) immunoblot. Coomassie Brilliant Blue (CBB) staining shows protein loading.
(B) AvrPtoB phosphorylation sites identified in vivo by LC-MS/MS. AvrPtoB-GFP was transiently expressed in Arabidopsis Col-0 protoplasts and total proteins were subjected to anti-GFP IP followed by trypsin digestion. Phosphorylated peptides were detected by LC-MS/MS. The observed y and b ions are numbered.
(C) Diagram of phosphorylation sites and host protein interaction domains in AvrPtoB. PID (green) indicates the Pto interaction domain, NID (blue) indicates the NPR1 interaction domain, BID (black slash) indicates the BAK1 interaction domain and FID (yellow) indicates the Fen interaction domain. Numbers correspond to amino acid (aa) residues.
(D) Quantification of S258, S210 and S205 phosphorylation. AvrPtoB-GFP was expressed in Col-0 and snrk2.8 protoplasts and total proteins were subjected to anti-GFP IP followed by tryptic digestion. Phosphorylated peptides were detected by LC-MS/MS with the parallel reaction monitoring method. Peptide phosphorylation ratios (phosphorylated/non-phosphorylated) were determined using Skyline software. Data are means ± SE of three biological replicates (separate transfections). Asterisks indicate significant differences (Student’s t-test, **p<0.01).
Next, we investigated AvrPtoB phosphorylation in vivo. AvrPtoB-GFP was expressed in Arabidopsis Col-0 protoplasts, enriched by anti-GFP IP, and samples were subjected to trypsin digestion and mass spectrometry analyses (LC-MS/MS). Five phosphorylated AvrPtoB residues (T111, S205, S210, S217, and S258) were identified by LC-MS/MS (Figure 2B). Previously, AvrPtoB was demonstrated to be phosphorylated at S258 in tomato, indicating that phosphorylation of this residue is conserved across diverse plant species (Xiao et al., 2007b). Aside from T111, all other phosphorylated residues map to specific domains required for binding the target proteins NPR1 (S205, S210, S217, and S258) and BAK1 (S258) (Figure 2C). The region surrounding S258 matches the proposed consensus phosphorylation motif of the SnRKs/CDPKs (Supplemental Table 1).
To determine the importance of SnRK2.8 for AvrPtoB phosphorylation in planta, AvrPtoB-GFP was expressed in Arabidopsis protoplasts from both wild-type Col-0 and the snrk2.8 knockout. AvrPtoB-GFP was then immunoprecipitated after transfection, subjected to trypsin digestion and LC-MS/MS using parallel reaction monitoring (PRM). Using PRM, we were able to detect phosphorylation of the five previously identified AvrPtoB residues. The phosphorylation ratios (phosphorylated/nonphosphorylated peptides) of T111 and S217 were similar between Col-0 and snrk2.8 (Supplemental Figure 4A and B). However, the phosphorylation ratios of the other three residues were lower in snrk2.8 compared to Col-0. Phosphorylation of S258 was significantly reduced by over 90% in snrk2.8 and phosphorylation of S205 and S210 were reduced by ~50% (Figure 2D). Notably, S205, S210, and S258 are highly conserved as either S or T residues across 27 Pseudomonas AvrPtoB homologs (Supplemental Figure 5). Taken together, these results indicate that SnRK2.8 can directly phosphorylate AvrPtoB in vitro and in vivo.
SnRK2.8 is required for AvrPtoB virulence
AvrPtoB’s phosphorylated residues are located within regions known to bind to plant targets, which led us to investigate whether SnRK2.8 is required for AvrPtoB virulence (Figure 2C). Since AvrPto and AvrPtoB are redundant with respect to virulence, a Pseudomonas syringae pv. tomato DC3000 avrPto and avrPtoB double knockout strain was used (Lin et al., 2005). DC3000ΔavrPtoΔavrPtoB (DC3000 −/−) was transformed with a plasmid expressing AvrPtoB or the empty vector (EV) control. Col-0 and snrk2.8 plants were inoculated with DC3000 and DC3000 −/− strain variants by syringe infiltration. Four days post-inoculation DC3000 caused severe disease symptoms, while DC3000 −/− EV caused weak disease symptoms on both Col-0 and snrk2.8 (Figure 3A). DC3000 −/− carrying AvrPtoB caused much more severe symptoms compared to DC3000 −/− carrying EV on Col-0, but not the snrk2.8 knockout (Figure 3A). Consistent with the disease symptoms, the bacterial titer of DC3000 −/− AvrPtoB was significantly higher than that of DC3000 −/− EV in Col-0, but not in the snrk2.8 knockout (Figure 3B). These results indicate that SnRK2.8 is required for AvrPtoB virulence in Arabidopsis.

(A) Disease symptoms of Arabidopsis Col-0 and the snrk2.8 knockout after infection with P. syringae pv. tomato DC3000 (DC3000) variants. DC3000 and DC3000ΔavrPtoΔavrPtoB (−/−) carrying empty vector (EV) or expressing AvrPtoB on a plasmid were syringe infiltrated into Arabidopsis Col-0 and snrk2.8 at a concentration of 2 × 105 CFU mL−1. Disease symptoms were observed 4 days post-inoculation (dpi).
(B) Bacterial populations in Col-0 and snrk2.8 leaves 4 dpi. Bacterial inoculations were performed as described in (A). LOG CFU/cm2 = log colony forming units per cm2 of leaf tissue. Data are means ± SD (n ≥ 9 plants for day 0, n ≥ 18 plants for day 4). Different letters indicate significant differences (two-way ANOVA, Tukey’s test, p < 0.05).
(C) Callose deposition in Arabidopsis Col-0 and snrk2.8 after inoculation with DC3000, −/− EV, and −/−AvrPtoB. Leaves were inoculated with P. syringae at a concentration of 1 × 108 CFU mL−1 and harvested 16h later. Leaves were stained by 1% aniline blue and imaged by fluorescence microscopy. Scale bar, 100 μm.
(D) Quantification of callose deposits. Data from three independent experiments were used for statistical analysis. Data are means ± SD (n = 24 images from 24 leaves). Different letters indicate significant differences (two-way ANOVA, Tukey’s test, p < 0.05).
AvrPtoB suppresses plant defense triggered by PAMP recognition including the deposition of callose, a β−1–3 linked glucan polymer, that is defense induced and acts as a structural barrier (Torres et al., 2006). To investigate the impact of SnRK2.8 on AvrPtoB virulence, we examined callose deposition after inoculation with DC3000 −/− carrying EV or AvrPtoB. DC3000 −/− carrying EV elicited strong callose deposition in both Col-0 and snrk2.8 (Figure 3C and D). As expected, DC3000 −/− carrying AvrPtoB did not elicit strong callose deposition in Col-0. However, AvrPtoB was unable to suppress callose deposition in the snrk2.8 knockout (Figure 3C and D). These data indicate that AvrPtoB requires the plant kinase SnRK2.8 for suppressing callose deposition in Arabidopsis.
Phosphorylated AvrPtoB serine residues are required for virulence
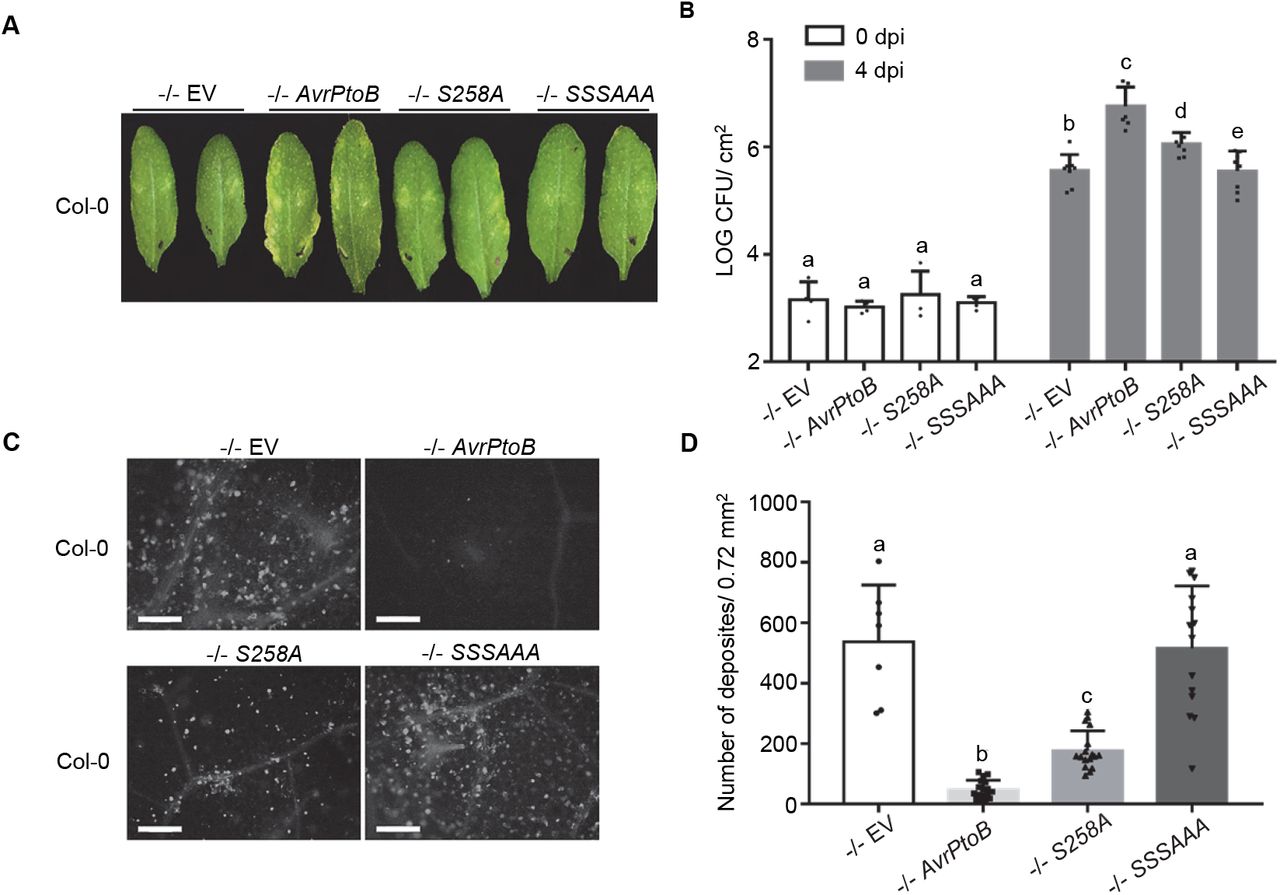
To test the functional relevance of AvrPtoB phosphorylation, we analyzed the virulence of AvrPtoB phospho-null mutants. We focused on S205, S210 and S258 due to their decrease in phosphorylation in the snrk2.8 knockout (Figure 2D). These serine residues were replaced by alanine to create single and triple phospho-null mutations. Previous research demonstrated that the alanine substitution of S258 resulted in a loss of virulence of AvrPtoB1–307 on the susceptible tomato genotype Rio Grande 76S (lacking the resistance gene Prf) (Xiao et al., 2007b). We first tested the ability of full-length AvrPtoBS258A to promote bacterial growth on Rio Grande 76S and Arabidopsis Col-0. The DC3000 −/− strain expressing AvrPtoBS258A was able to increase bacterial growth in both tomato and Arabidopsis, but not to the same extent as wild-type AvrPtoB (Supplemental figure 6A and B). In contrast, the DC3000 −/− strain expressing the 1-307 truncation of AvrPtoBS258A was unable to increase bacterial growth in both tomato and Arabidopsis (Supplemental Figure 6A and B). These results indicate that the phosphorylation of S258 is important, but not sufficient, for the virulence of full-length AvrPtoB.
Next, we tested the virulence of the phospho-null mutant AvrPtoBS205AS210A258A compared to wild-type AvrPtoB and AvrPtoBS258A. Arabidopsis Col-0 was syringe infiltrated with DC3000 −/− carrying EV, AvrPtoB, AvrPtoBS258A, and AvrPtoBS205AS210A258A. Disease symptoms, bacterial titers, callose deposition were analyzed. As expected, infection with DC3000 −/− carrying AvrPtoB resulted in leaf disease symptoms, increased bacterial growth, and suppression of callose deposition compared to EV (Figure 4). Infection with DC3000 −/− AvrPtoBS258A resulted in an intermediate phenotype, with slight disease symptoms, a partial increase in bacterial growth compared to EV, and partial suppression of callose compared to EV (Figure 4). However, DC3000 −/− carrying AvrPtoBS205AS210A258A was unable to induce disease symptoms, grew to the same level as the EV control, and was unable to suppress callose deposition (Figure 4). Western blot analyses demonstrated that all AvrPtoB variants were equally expressed in DC3000 −/− (Supplemental Figure 6C). Collectively, these results demonstrate that the combined phosphorylation of S205, S210, and S258 play a critical role in AvrPtoB virulence.

(A) Disease symptoms of Arabidopsis Col-0 after inoculation with P. syringae pv. tomato DC3000ΔavrPtoΔavrPtoB (−/−) variants. DC3000 −/− carrying empty vector (EV) or plasmids expressing wild-type AvrPtoB, AvrPtoBS258A, or AvrPtoBS205AS210AS258A (SSSAAA) were syringe infiltrated into Arabidopsis Col-0 at a concentration of 2 × 105 CFU mL−1. Disease symptoms were observed 4 days post-inoculation (dpi).
(B) Bacterial populations in Col-0 leaves 4 dpi. Bacterial inoculations were performed as described in (A). LOG CFU/cm2 = log colony-forming units per cm2 of leaf tissue. Data are means ± SD (n = 4 plants for day 0, n = 8 plants for day 4). Different letters indicate significant differences (two-way ANOVA, Tukey’s test, p < 0.05).
(C) Callose deposition in Arabidopsis Col-0 after inoculation with −/− carrying EV, AvrPtoB, S258A, and SSSAAA. Leaves were inoculated with P. syringae at a concentration of 1 × 108 CFU mL−1 and harvested 16h later. Leaves were stained by 1% aniline blue and imaged by fluorescence microscopy. Scale bar, 100 μm.
(D) Quantification of callose deposits. Data are means ± SD (n = 8 images from 8 leaves for −/− EV, n =18 images from 18 leaves for rest variants). Different letters indicate significant differences (one-way ANOVA, Tukey’s test, p < 0.05).
AvrPtoB-mediated inhibition of NPR1 accumulation requires the plant kinase SnRK2.8
AvrPtoB associates with and ubiquitinates NPR1 in the presence of SA (Chen et al., 2017). To further investigate the relevance of SnRK2.8 for promoting AvrPtoB virulence, we examined AvrPtoB-mediated inhibition of NPR1 accumulation in Col-0 and snrk2.8. Both Col-0 and snrk2.8 were pretreated with 0.5mM SA for 6 hours and then syringe inoculated with DC3000 −/− EV and DC3000 −/− AvrPtoB. NPR1 accumulation was detected by anti-NPR1 immunoblotting. Infection with DC3000 −/− EV induced high accumulation of NPR1 in both Col-0 and snrk2.8 (Figure 5A and B). However, infection with DC3000 −/− AvrPtoB inhibited NPR1 accumulation in Col-0, but not in the snrk2.8 knockout, indicating that SnRK2.8 is required for AvrPtoB-mediated inhibition of NPR1 accumulation (Figure 5A and B).

(A) NPR1 accumulation in Arabidopsis Col-0 and the snrk2.8 knockout after inoculation with P. syringae pv. tomato DC3000ΔavrPtoΔavrPtoB (−/−) variants. DC3000 −/− carrying the empty vector (EV) or a plasmid expressing wild-type AvrPtoB were syringe infiltrated into Col-0 and snrk2.8 at a concentration of 1 × 108 CFU mL−1 and proteins extracted after four hours. Protein extracts were subjected to anti-NPR1 immunoblotting. Coomassie Brilliant Blue (CBB) staining shows equal protein loading.
(B) Quantification of NPR1 band intensity in (A). NPR1 bands intensities were quantified by Image Lab 6.0.1 (BIO-RAD). The values were normalized first by Rubisco bands and subsequently by the intensities of “0h” bands. Data are means ± SD (n = 14 leaves). Different letters indicate significant differences (one-way ANOVA, Tukey’s test, p < 0.05).
(C) NPR1 accumulation in the presence of AvrPtoB phosphorylation mutants in N. benthamiana after Agrobacterium-mediated transient expression. 35S::NPR1-HA was co-expressed with FLAG-tagged Dex-inducible AvrPtoB, AvrPtoBS258A, AvrPtoBS205AS210AS258A (SSSAAA), AvrPtoBS258D or AvrPtoBS205DS210DS258D (SSSDDD). The Dex inducible EV was used as a control. The expression of AvrPtoB-FLAG phosphorylation variants was induced by 15 μM DEX for 5 hours 24h post-Agrobacterium infiltration. Protein extracts were subjected to anti-HA and anti-FLAG immunoblotting. CBB staining shows protein loading.
(D) Quantification of NPR1-HA band intensity in (C). NPR1-HA bands intensities were quantified by Image Lab 6.0.1. The values were normalized first by Rubisco bands and subsequently by the intensities of “EV” treatment. Data are means ± SD (n = 6 leaves). Different letters indicate significant differences (one-way ANOVA, Tukey’s test, p < 0.05).
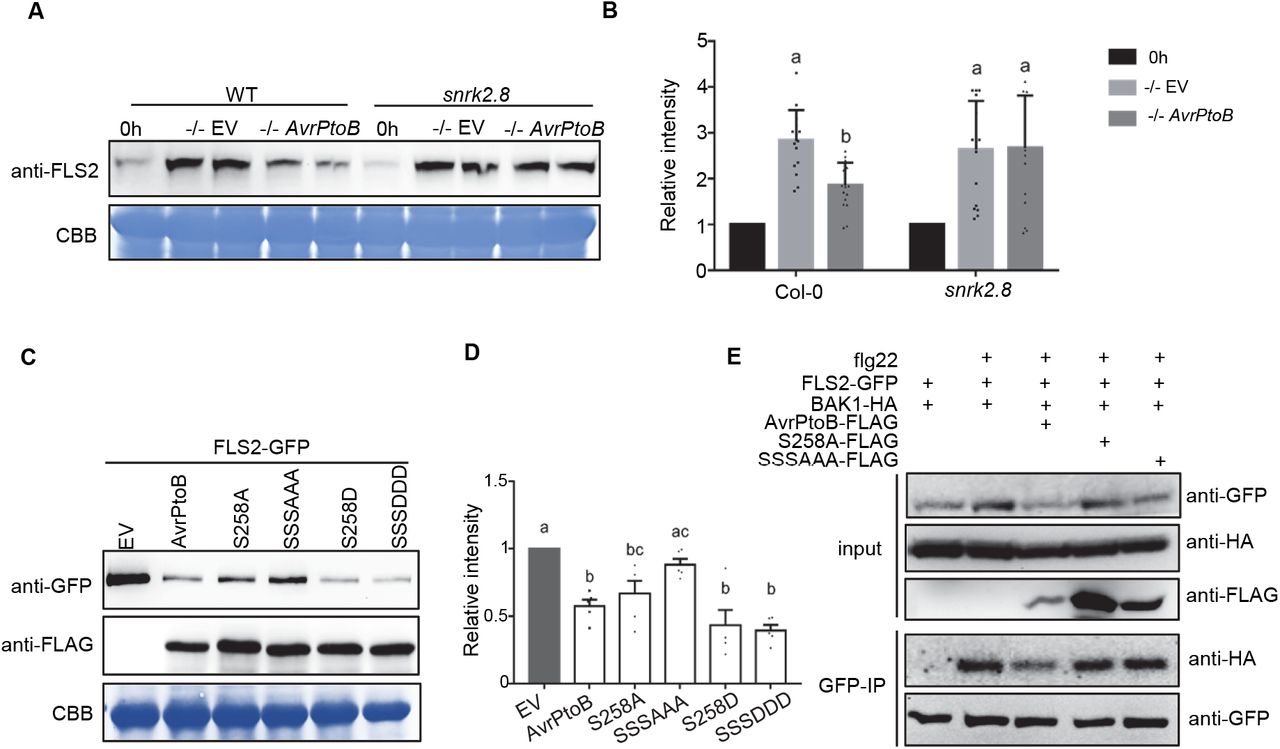
Next, the importance of AvrPtoB phosphorylated residues for inhibition of NPR1 accumulation was examined. We co-expressed 35S::NPR1-HA and DEX inducible AvrPtoB-FLAG, AvrPtoB phospho-null, and AvrPtoB phospho-mimetic mutants in N. benthamiana. Wild-type AvrPtoB, AvrPtoBS258D, and AvrPtoBS205DS210DS258D significantly decreased NPR1 accumulation compared to the EV control (Figure 5C and D). Both AvrPtoBS258D and AvrPtoBS205DS210DS258D were able to strongly suppress NPR1 accumulation, indicating that mimicking phosphorylation also promotes virulence of AvrPtoB (Figure 5C and D). AvrPtoBS258A was still able to suppress the accumulation of NPR1, indicating that the phosphorylation of serine 258 is not sufficient for this function of AvrPtoB (Figure 5C and D). In contrast, AvrPtoBS205AS210A258A was unable to suppress NPR1 accumulation (Figure 5C and D). Collectively, these results show that AvrPtoB-mediated inhibition of NPR1 accumulation requires the plant kinase SnRK2.8 and the SnRK2.8-mediated phosphorylated residues.
AvrPtoB-mediated inhibition of FLS2 accumulation and FLS2-BAK1 complex formation requires the plant kinase SnRK2.8
The flagellin receptor FLS2 is another host target of AvrPtoB (Gómez-Gómez and Boller, 2000; Lu et al., 2011). AvrPtoB directly associates with FLS2 and ubiquitinates FLS2 (Göhre et al., 2008). To investigate the relevance of SnRK2.8 with AvrPtoB-mediated inhibition of FLS2 accumulation, we compared FLS2 accumulation after infection with DC3000 −/− carrying EV or AvrPtoB in different genetic backgrounds. Col-0 and the snrk2.8 knockout were syringe inoculated and FLS2 accumulation was detected by anti-FLS2 immunoblotting. FLS2 accumulation was enhanced after infection with DC3000 −/− EV in both Col-0 and snrk2.8 (Figure 6A and B). In contrast, AvrPtoB was able to suppress FLS2 accumulation in Col-0 but not in the snrk2.8 knockout, which demonstrates that SnRK2.8 is required for AvrPtoB-mediated suppression of FLS2 accumulation (Figure 6A and B).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
(A) FLS2 accumulation in Arabidopsis Col-0 and the snrk2.8 knockout after inoculation with P. syringae pv. tomato DC3000ΔavrPtoΔavrPtoB (−/−) variants. DC3000 −/− carrying the empty vector (EV) or a plasmid expressing wild-type AvrPtoB were syringe infiltrated into Col-0 and snrk2.8 at a concentration of 1 × 108 CFU mL−1 and proteins extracted after 8h. Protein extracts were subjected to anti-FLS2 immunoblotting. Coomassie Brilliant Blue (CBB) staining shows equal protein loading.
(B) Quantification of FLS2 band intensity in (A). FLS2 band intensities were quantified by Image Lab 6.0.1. The values were normalized first by Rubisco bands and subsequently by the intensities of “0h” bands. Data are means ± SD (n = 14 leaves). Different letters indicate significant differences (one-way ANOVA, Tukey’s test, p < 0.05).
(C) FLS2 accumulation in the presence of AvrPtoB phosphorylation mutants in N. benthamiana after Agrobacterium-mediated transient expression. 35S::FLS2-GFP was co-expressed with FLAG-tagged Dex-inducible AvrPtoB, AvrPtoBS258A, AvrPtoBS205AS210AS258A (SSSAAA), AvrPtoBS258D, or AvrPtoBS205DS210DS258D (SSSDDD). The Dex inducible EV was used as a control. The expression of AvrPtoB-FLAG phosphorylation variants was induced by 15 μM DEX for 5 hours 24h post-Agrobacterium infiltration. Protein extracts were subjected to anti-GFP and anti-FLAG immunoblotting. CBB staining shows protein loading.
(D) Quantification of FLS2-GFP band intensity in (C). FLS2-GFP bands intensities were quantified by Image Lab 6.0.1. The values were normalized first by Rubisco bands and subsequently by the intensities of “EV” treatment. The data from two independent experiments were used for statistical analysis. Data are means ± SD (n = 6 leaves). Different letters indicate significant differences (one-way ANOVA, Tukey’s test, p < 0.05).
(E) Analyses of AvrPtoB-mediated inhibition of FLS2 complex formation. AvrPtoB-FLAG variants, BAK1-HA, and FLS2-GFP were co-expressed in the indicated combinations in N. benthamiana using Agrobacterium-mediated transient expression. The expression of AvrPtoB-FLAG phosphorylation variants was induced by 15 μM DEX for 3 hours 24h post-Agrobacterium infiltration. Leaves were infiltrated with 10 μM flg22 peptide 15 minutes before harvesting protein extracts for GFP immunoprecipitation and western blotting. Protein inputs and immunoprecipitated samples were detected by immunoblot.
To test if AvrPtoB phosphorylated residues are required for inhibition of FLS2 accumulation, we co-expressed 35S::FLS2-GFP and DEX inducible AvrPtoB-FLAG, AvrPtoB phospho-null, and AvrPtoB phospho-mimetic mutants in N. benthamiana. Wild-type AvrPtoB, AvrPtoBS258D, and AvrPtoBS205DS210DS258D significantly decreased FLS2 accumulation compared to the EV control (Figure 6C and D). AvrPtoBS258A was still able to decrease the accumulation of FLS2, indicating that the phosphorylation of serine 258 is not sufficient for this AvrPtoB function (Figure 6C and D). In contrast, the phospho-null mutant AvrPtoBS205AS210AS258A was unable to suppress FLS2 accumulation (Figure 6C and D). Collectively, these results demonstrate that AvrPtoB phosphorylated residues and the plant kinase SnRK2.8 are required for inhibition of FLS2 accumulation.
FLS2-mediated recognition of flg22 induces the formation of an FLS2-BAK1 complex, which is required for downstream immune signaling (Chinchilla et al., 2007; Shan et al., 2008). We investigated the importance of AvrPtoB phosphorylated residues for disrupting FLS2-BAK1 complex formation. We examined flg22-induced FLS2-BAK1 association in the presence of AvrPtoB and AvrPtoB phospho-null mutants after transient expression in N. benthamiana. As shown in Figure 6E, wild-type AvrPtoB effectively diminished FLS2-BAK1 association in the presence of flg22, but AvrPtoBS258A and AvrPtoBS205AS210AS258A were unable to suppress FLS2-BAK1 complex formation. These results suggest that phosphorylation of S258 is required and sufficient for disrupting FLS2-BAK1 complex formation.
Discussion
Post-translational modification (PTM) is a rapid and specific protein regulation mechanism that plays important roles in plant growth, development, and disease resistance (Withers and Dong, 2017). Recent comprehensive proteome analyses in Arabidopsis reveal that 47% of the identified proteins are phosphorylated under specific conditions(Withers and Dong, 2017; Mergner et al., 2020). Many pathogen effectors require PTM in planta for their virulence activity. Effectors from the bacterial pathogens Pseudomonas syringae, Xanthomonas, and Rhizobium are phosphorylated in plants (Li et al., 2013; Kim et al., 2014; Tahir et al., 2019). The phosphorylation of effectors by plant kinases, in many instances, enhances their virulence-promoting activity (Tahir et al., 2019). However, the host kinases involved in effector phosphorylation are largely unknown. Our study reveals that the AvrPtoB is phosphorylated on five serine/threonine residues, three of which (S205, S210, and S258) are required for AvrPtoB virulence in Arabidopsis and conserved across P. syringae AvrPtoB. One SnRK family member, SnRK2.8, interacts with AvrPtoB, is primarily responsible for phosphorylating AvrPtoB, and is required for AvrPtoB virulence promoting activities.
Sucrose non-fermenting-1 (SNF1)-related protein kinases (SnRKs) are the plant homologs of SNF1 kinase in yeast and AMP-activated protein kinase (AMPK) in mammals, which are key regulators of carbon metabolism and energy status (Coello et al., 2011; Hardie et al., 2012). The plant SnRK family can be subdivided into three subfamilies: SnRK1, SnRK2, and SnRK3. The SnRK2 subfamily members have diverged from Snf1 and AMPK and are plant specific. SnRKs act as metabolic sensors, integrating multiple developmental processes and very diverse stress conditions including pathogen infection (Halford and Hey, 2009; Hulsmans et al., 2016) An increasing number of studies show the key roles of SnRKs in plant responses to different types of pathogens. Tomato SnRK1 phosphorylates the pathogenic protein βC1 of the tomato yellow leaf curl virus to limit viral infection (Shen et al., 2011). Tomato SnRK1 is also involved in cell death elicited by Xanthomonas effectors and may be related to Adi3-mediated suppression of cell death (Szczesny et al., 2010; Avila et al., 2012).
SnRK2.8, an Arabidopsis SnRK2 member, is transcriptionally induced by infection with P. syringae DC3000 and mediates SA induced systemic acquired resistance (Lee et al., 2015). SnRK2.8 phosphorylates NPR1 in the cytoplasm and mediates the nuclear import of NPR1 (Lee et al., 2015). Since both SnRK2.8 and AvrPtoB associate with NPR1, we assume these two proteins are likely in close contact with one another. In the snrk2.8 knockout, the phosphorylation of S258 is reduced by over 90% and the phosphorylation of S205 and S210 is reduced by ~50%. These results indicate that SnRK2.8 is one of the main kinases involved in AvrPtoB phosphorylation. However, since some phosphorylation of S205/S210/S258 remain in the snrk2.8 knockout and SnRK2.8 does not influence phosphorylation of T111 and S217 in planta, other kinases are also involved phosphorylating AvrPtoB. AvrPtoB weakly associates with SnRK1.1, SnRK2.6 and another two CDPK members in planta. These kinases may also be capable of AvrPtoB phosphorylation.
Previously, only one kinase has been identified that is capable of phosphorylating a plant pathogen effector. The cyst nematode effector 10A07 interacts with an Arabidopsis autophagy related (Atg) 1 kinase, Atg1C, in planta (Hewezi et al., 2015). Atg1C phosphorylates 10A07 on two serine residues, which are required for nuclear localization of this effector. An inactive variant of Atg1C reduced the plant susceptibility to cyst nematode (Hewezi et al., 2015). Similarly, AvrPtoB virulence promoting activities are blocked in the snrk2.8 knockout and SnRK2.8 phosphorylated residues on AvrPtoB are required for effector function. Both SnRKs and Atg kinases are widely conserved among eukaryotes and are involved in plant immune responses (Hayward et al., 2009; Coello et al., 2011). However, these kinases could be a double-edged sword as they are also exploited by pathogen effectors during infection.
Phosphorylation can impact protein-protein interactions by creating an intricate binding surface (Tarrant and Cole, 2009). Effectors from human pathogens, including the E. coli effector Tir (Translocated intimin receptor) and Helicobacter pylori effector CagA (Caytotoxin-associated antigen A) are subjected to host phosphorylation. Phosphorylation of Tir and CagA are required for the binding of host adaptor proteins for bacterial adhesion (Nougayrede et al., 2003; Higashi et al., 2004). The plant pathogen effectors HopQ1, XopN, and XopE2 are phosphorylated in plants, which results in their associations with host 14-3-3 proteins and phosphorylated residues are required for effector virulence (Taylor et al., 2012; Li et al., 2013; Dubrow et al., 2018). In our study, the phosphorylation of AvrPtoB is required for the effector-mediated inhibition of NPR1/FLS2 accumulation and FLS2-BAK1 complex formation. Three critical AvrPtoB phosphorylated residues are not present in the AvrPtoB E3 ligase domain, so they should not directly affect the AvrPtoB ubiquitination activity. Thus, the phosphorylation of AvrPtoB may be required for the interactions with its targets. S258 is located in both the NPR1 and BAK1 binding domains of AvrPtoB, while S205 and S210 are present in the NPR1 binding domain. The single phospho-null S258A mutant was still able to inhibit NPR1/FLS2 accumulation. However, the triple phospho-null mutant completely lost these functions, indicating that the phosphorylation of S205 and S210 is required for AvrPtoB-mediated target degradation (Figure 5C, D and Figure 6C, D). However, the S258A was unable to disrupt FLS2-BAK1 complex formation. These data are consistent with previous observations that AvrPtoB is a multifunctional protein with different interfaces involved in interacting with diverse host proteins (Martin, 2011).
Despite the importance of effector phosphorylation, the plant kinases involved remain largely elusive. Here, we identified a member of the SnRK-CDPK family, SnRK2.8, whose phosphorylation of the conserved bacterial effector AvrPtoB promotes effector virulence activities. The SnRK-CDPK family is conserved in plants and most reported effectors phosphorylated residues are consistent with the SnRK-CDPK phosphorylation site preferences. Thus, we hypothesize that different SnRKs and CPDKs may be involved in phosphorylation of diverse effector proteins. Future investigation of the role of SnRK-CDPKs in phosphorylation of diverse pathogen effectors will provide a more comprehensive understanding of how pathogen effectors exploit their hosts to facilitate disease development.
Methods
Plant material and growth conditions
Arabidopsis thaliana seeds were stratified for two days at 4℃ in the dark before sowing. Plants were grown in a controlled environmental chamber at 23℃ and 70% relative humidity with a 10-hr-light /14-hr-dark photoperiod (100 μM m−2 s−1). Four-to-five-week old plants were used for all experiments. A confirmed snrk2.8 knockout T-DNA insertion line (SALK_073395) in the Columbia (Col-0) background was obtained from the Arabidopsis Biological Resource Center (ABRC) (Shin et al., 2007).
N. benthamiana was grown in a growth chamber at 26℃ with a 16-hr-light/8-hr-dark photoperiod (180 μM m−2 s−1). Three-to-four-week old plants were used for Agrobacterium-mediated transient protein expression.
Yeast two-hybrid screen
The GAL4 based Matchmaker yeast two-hybrid system was used for the AvrPtoB-kinase interaction screen (Clonetech). AvrPtoB and a mCherry negative control were cloned into the pGADT7 vector fused to the GAL4 activation domain and HA tag. Arabidopsis SnRKs and CDPKs were cloned into the pGBKT7 vector, which contains the GAL4 DNA binding domain and N-terminal Myc epitope tag. In order to detect interactions with CDPKs, their C-terminal Ca2+ regulatory and auto-inhibitory domains were removed prior to clone into the pGBKT7 vector. The pGADT7-AvrPtoB and pGADT7-mCherry plasmids were separately co-transformed with each pGBKT7-SnRK/CPKΔC plasmid into the yeast strain AH109, colonies were selected on SD -Leu/-Trp dropout media and tested for interactions on SD -Leu/-Trp/-His dropout media containing X-α-Gal. Yeast transformation and media preparation were performed per manufacturer instructions (Clontech). To confirm protein expression, yeast proteins were extracted as described previously and subjected to anti-HA HRP (Roche #12013819001; 1:2,000) and anti-Myc (Clontech #631206; 1: 2,000) immunoblotting (Zhang et al., 2011). Primers used in all experiments are listed in Table S2.
Phylogeny and expression of SnRKs
A total of 38 Arabidopsis SnRK protein sequences were obtained from the Arabidopsis information resource (TAIR, http://Arabidopsis.org). Multiple sequence alignments were performed with MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms (Kumar et al., 2018). The phylogeny was determined by maximum likelihood method (1000 bootstrap replicates). Data of SnRK transcriptional expression in leaves were obtained from BAR Expression Angler (Austin et al., 2016).
Co-immunoprecipitation assays
To confirm the association between AvrPtoB and SnRK/CDPKs in planta, kinase dead variants were generated and tested for their ability to associate with wild-type AvrPtoB after transient expression in N. benthamiana. SnRK1.1-KD (K71A), SnRK2.6-KD (K50A), SnRK2.8-KD (K33A), CPK4ΔC-KD (K54A) and CPK5ΔC-KD (K126A) kinase dead variants were generated by PCR-based site-directed mutagenesis and fused with a C-terminal HA tag in the binary vector pGWB414 (NAKAGAWA et al., 2007). Primers are listed in Table S2. AvrPtoB fused with a C-terminal GFP tag was cloned into the dexamethasone (Dex)-inducible binary vector pTA7001 (Gu and Innes, 2011). Binary vectors were transformed into Agrobacterium tumefaciens GV3101. Agrobacterium suspensions were co-infiltrated into N. benthamiana leaves at an OD600 = 0.4 for AvrPtoB and an OD600 = 0.6 for each kinase. Twenty-four hours post-inoculation, 15 μM DEX and 0.01% Silwet L-77 were sprayed to induce the expression of AvrPtoB-GFP. Two grams of leaf tissue per sample were collected three hours post-DEX application.
For immunoprecipitations, N. benthamiana leaf tissues were ground in liquid nitrogen and re-suspended in 2 mL IP buffer (50mM Tris-HCL ph7.5, 150mM NaCl, 0.1% Triton, 0.2% NP-40, 1× complete protease inhibitor (Thermo Fisher Scientific #A32963) and 1× phosphatase inhibitor (Thermo Fisher Scientific #A32957), 1mM DTT, 40uM MG132, 0.5% PVP). Samples were centrifuged at 14,000 rpm for 15 min and filtered to remove debris using a poly-prep chromatography column (10mL, Bio-Rad). The supernatant was incubated with 25 μL of anti-GFP agarose beads at 4℃ for 1.5 hr. Beads were washed once with IP buffer by centrifuging at 3000 rpm for two min and twice by filtration through a pierce centrifuge column (0.8 mL, Bio-Rad). Proteins were eluted from the beads by boiling in 2× Laemmli buffer for five min. Proteins were separated by SDS-PAGE and immunoblotted with anti-HA HRP (Roche #12013819001; 1:2,000) and anti-GFP HRP (Miltenyi Biotec #130-091-833; 1:3,000).
To test the ability of AvrPtoB phosphorylation mutants to disrupt FLS2-BAK1 complex formation, AvrPtoB phospho-null mutants (S258A and S205AS210AS258A) were generated by PCR-based site-directed mutagenesis. Primers are listed in Table S2. AvrPtoB variants were fused with C-terminal 3× FLAG tag in binary vector pTA7001, and FLS2 and BAK1 were separately fused with C-terminal GFP tag and HA tag in binary vector pEarleyGate 103 and pGWB14 (Earley et al., 2006; NAKAGAWA et al., 2007). The binary vectors were transformed into Agrobacterium tumefaciens GV3101. FLS2, BAK1, and AvrPtoB variants Agrobacterium suspensions were co-infiltrated into N. benthamiana leaves, 15 μM DEX and 0.01% Silwet L-77 were sprayed to induce the expression of AvrPtoB at 24 hpi for three hours, two grams of leaf tissue for each sample were collected after 15 min treatment with 5 mM MgCl2 or 10 μM flg22. Immunoprecipitation was performed as described above.
Recombinant protein purification
AvrPtoB and SnRK2.8 were cloned in E.coli expression vector pDEST15 (Invitrogen) fused with N-terminal GST tag, the constructs were transformed into E.coli BL21 (DE3). 200 mL of E.coli culture was grown at 28 ℃ until OD600 = 0.5. Protein expression was induced with 0.5mM IPTG at 16 ℃ for 12 hr. Cells were harvested by centrifuging at 5000 g 4 ℃ for 10 min and washed once with buffer A (0.1 M Tris-HCl pH 7.5, 150 mM NaCl, 1 mM PMSF, 1 × CPI, 10 mM DTT and 10 uM MG132). Cell pellets were resuspended in 3 mL of buffer A with 15 μg/mL lysozyme and incubated on ice for 30 min. Total protein was released by sonication and incubated with Glutathione Sepharose 4B (GE Healthcare #GE17-0756-01) at 4 ℃ for one hour. Agarose beads were washed three times with buffer A by centrifuging at 5000 g for 5 min. Proteins were eluted by incubating with buffer B (50 mM Tris-HCl pH8, 10 mM reduced Glutathione) for 10 min at room temperature (RT).
Kinase activity assay
An in vitro kinase activity assay was performed with recombinant proteins, 3 μg of GST-AvrPtoB and 0.3-1 μg GST-SnRK2.8 were mixed in kinase buffer (20 mM Tris-HCl pH7.5, 10 mM MgCl2, 1 mM CaCl2, 100 μM ATP, 1 mM DTT). The kinase reaction was performed at 30 ℃ for 30 min and stopped by 3 × Laemmli buffer. Protein samples were separated in SDS-PAGE and immunoblotted with anti-pSer/Thr antibody (Sigma #P3430; 1:1000) and anti-GST antibody (Sigma; 1: 3000).
Phosphorylation site identification and quantification
To identify AvrPtoB phosphorylation sites in vivo, AvrPtoB-YFP in the pBluescript vector was transiently expressed in Col-0 and snrk2.8 protoplasts, protoplast preparation and transient transformation were performed as previously described (Yoo et al., 2007). One mL of protoplasts were transfected with 100 μg of plasmid and collected after 9 hr. Proteins were released in IP buffer (without 0.5% PVP) and subject to GFP-IP as described above. Protein peptides were generated by in-solution trypsin digest as previously described and subjected to LC-MS/MS run by Orbitrap Fusion Lumos mass spectrometer (Thermo Fisher Scientific) at the UC Davis Proteomics Core (Minkoff et al., 2013). LC-MS/MS data were analyzed in MaxQuant (Tyanova et al., 2016).
To quantify the phosphorylated peptides, an inclusion list of phosphopeptide and control peptides, including the Mono-isotopic precursor (m/z) and charge state (z), was generated by software Skyline based on previous MS data, as shown in Table S3 (MacLean et al., 2010). The peptide samples were scanned by Orbitrap Fusion Lumos mass spectrometer (Thermo Fisher Scientific) with a parallel reaction monitoring (PRM) method. The PRM data were analyzed by MaxQuant and Skyline, the peptides peak areas were exported for the quantification analysis.
Bacterial growth assay
The broad host range plasmid pBAV226 was used to express AvrPtoB variants in Pseudomonas syringae. pBAV226 harboring AvrPtoB or its derivatives were transformed into the P. syringae pv. tomato DC3000ΔavrPtoΔavrPtoB (DC3000 −/−) strain by electroporation (Lin and Martin, 2005). DC3000, DC3000 −/−, and DC3000 −/− carrying plasmids expressing AvrPtoB variants were syringe infiltrated into five-week-old Arabidopsis Col-0 or snrk2.8 leaves at a concentration of 2 × 105 CFU mL−1 (OD600 = 0.0002). Bacterial titers were determined as colony forming units (CFU) per cm2 at four days post inoculation as previously described (Liu et al., 2009).
Callose staining and quantification
Four-week-old Arabidopsis plants were inoculated with Pst DC3000, DC3000 −/−, or DC3000 −/− carrying plasmids expressing AvrPtoB variants at a concentration of 1 × 108 CFU mL−1 (OD600 = 0.1) for 16 hr. The infected leaves were fixated and destained in 95% ethanol until they were cleared of chlorophyll. The cleared leaves were washed twice with 70% ethanol and three times with distilled water and stained with 1% aniline blue in 150 mM K2HPO4 (pH 9.5) for 30 min in dark at RT. Callose deposition was imaged by fluorescence microscope (Leica CMS GmbH) using a DAPI filter. Image J was used for the quantification of the number of callose (Collins, 2007).
NPR1 and FLS2 accumulation
To test the ability of AvrPtoB to inhibit NPR1 and FLS2 accumulation in Arabidopsis Col-0 and the snrk2.8 knockout. DC3000 −/− carrying the empty vector (EV) or a plasmid expressing wild-type AvrPtoB were syringe infiltrated into Col-0 and snrk2.8 at a concentration of 1 × 108 CFU mL−1 and proteins extracted after 4h (NPR1) and 8h (FLS2). The immunoblot was performed by anti-NPR1 (Agrisera #AS12 1854; 1:1000) and anti-FLS2 (Agrisera #AS12 1857; 1:5000) primary antibody followed by anti-rabbit-HRP (BioRad #170-5046; 1:3000) secondary antibody. Bands intensities of immunoblotting and Coomassie Brilliant Blue (CBB) staining were quantified by Image Lab 6.0.1 (BIO-RAD). The values were normalized first by Rubisco bands and subsequently by the intensities of “0h” bands.
To test the ability of AvrPtoB phosphorylation mutants to inhibit NPR1 and FLS2 accumulation, AvrPtoB phospho-null mutants (S258A and S205AS210AS258A) and AvrPtoB phospho-mimic mutants (S258D and S205DS210DS258D) were generated by PCR-based site-directed mutagenesis. Primers are listed in Table S2. AvrPtoB variants were fused with C-terminal 3× FLAG tag in binary vector pTA7001, and NPR1 and FLS2 were separately fused with C-terminal HA tag and GFP tag in binary vector pEarleyGate 103 and pGWB14 (Earley et al., 2006; NAKAGAWA et al., 2007). The immunoblot was performed by anti-HA HRP (Roche #12013819001; 1:2,000), anti-FLAG (Sigma #A8592; 1:3000), and anti-GFP HRP (Miltenyi Biotec #130-091-833; 1:3,000). Bands intensities of immunoblotting and Coomassie Brilliant Blue (CBB) staining were quantified by Image Lab 6.0.1 (BIO-RAD). The values were normalized first by Rubisco bands and subsequently by the intensities of “EV” bands.
Plant protein extraction and immunoblotting
Plant tissues were ground in liquid nitrogen and homogenized in Protein Extraction Buffer [(PEB: 50 mM Tris-HCl, pH 7.5, 1 mM EDTA, pH 8.0, 150 mM NaCl, 0.1% Triton X-100, 0.5% IGEPAL, 5% glycerol, 1 mM PMSF, 3 mM DTT, 1 × CPI (Thermo Fisher Scientific), 1× PPI (Thermo Fisher Scientific), 50 mM MG132 (Sigma-Aldrich)] (Chen et al., 2017). The homogenate was cleared by centrifuging at 14,000 rpm for 15 min at 4 ℃, and boiled with 5 × SDS buffer (250 mM Tris-HCl pH 6.8, 6% SDS, 0.5 M DTT, 30% glycerol, 0.08% bromophenol blue) for 10 min.
Statistical analyses
Statistical analyses were performed by Prism 7 software (GraphPad). The data are presented as mean ± SD or ± SEM as indicated in figure legends. For quantification of phosphorylated peptides and quantification of NPR1 or FLS2 band intensity, n represents the number of experimental replicates. For bacterial growth curve assay and callose deposition, n represents the number of individual plants. Student’s t test was used to compare means for two groups. One-way ANOVA with Turkey’s multiple-comparison test was performed to compare means from several groups against a control group mean. Two-way ANOVA with Turkey’s multiple-comparison test was performed to compare means between groups. Statistical analyses, p-values, and the exact value of n are described in detail in the figures and figure legends.
Funding
This work was supported by grants from United States Department of Agriculture: USDA-NIFA 2015-67013-23082 and National Institutes of Health R01: R01GM092772 awarded to G.C.
Author Contributions
G.C. conceived the study, L.L. and G.C. designed experiments, and L.L. performed most experiments. D.M.S. analyzed the AvrPtoB homologs. L.L. and G.C. wrote the manuscript. All authors approved of the final manuscript.
Supplemental Table and Figure Legends.
Supplemental Table 1. The phosphorylation sites of bacterial type III effectors identified by mass spectrometry. Related to Figure 1. Data were collected from the references in the table. Red indicates the phosphorylated residue.
Supplemental Table 2. Primers used in this study. Related to Materials and Methods.
Supplemental Table 3. Isolation list for PRM. Related to Figure 2. Modified and unmodified peptide sequences used to quantify phosphorylation by PRM are noted. CID = collision induced dissociation, m/z = mass to charge, z = charge state.
Supplemental Figure 1. Amino acid conservation surrounding known effector phosphorylation sites. Related to Figure 1.
WebLogo alignment of the effector phosphorylation sites shown in Table S1. Red arrows point the phosphorylated residues. Amino acid size correlates with degree of conservation. The sequence logo was created by the WebLogo 3 (Crooks et al., 2004).
Supplemental Figure 2. Yeast-two hybrid screening of interactions between thePseudomonas syringae effector AvrPtoB and SnRK or CDPK members. Related to Figure 1.
Yeast two-hybrid (Y2H) assay of AvrPtoB and SnRK-CDPKs in the Matchmaker GaL4 system. AvrPtoB was expressed from the pGADT7 vector, while SnRKs and C-terminal deletions of CPKs (CPKs△C) were expressed from the pGBKT7 vector in Saccharomyces cerevisea. The pGBKT7 empty vector (EV) was included as a negative control. Blue colonies on SD -Leu/-Trp/-His/X-α-gal media indicate protein-protein interactions. AD = activation domain vector pGADT7, BD = binding domain vector pGBKT7.
Supplemental Figure 3. Protein expression ofSnRKs, CPKs, and AvrPtoB used in the Y2H assay. Related to Figure 1.
Yeast proteins were extracted and SnRK and CPK proteins were detected by anti-myc immunoblot and AvrPtoB was detected by anti-HA immunoblot. Expression of the positive (Pto) and negative (mCherry) controls used in Figure 1 are shown.
Supplemental Figure 4. AvrPtoB T111 and S217 phosphorylation are not lower in the snrk2.8 knockout compared to wild-type Col-0. Related to Figure 2.
AvrPtoB-GFP was expressed in Col-0 and snrk2.8 protoplasts, total proteins were subjected to anti-GFP IP followed by tryptic digestion. Phosphorylated peptides were detected by LC-MS/MS with the parallel reaction monitoring method. The peptide phosphorylation intensity of T111 (A) and the phosphorylation ratio of S217 (B) were determined using Skyline software. Data are means ± SE of three biological replicates (separate transfections). Asterisks indicate significant differences (Student’s t-test, **p<0.01).
Supplemental Figure 5. Conservation of phosphorylated residues in different AvrPtoB homologs. Related to Figure 2.
The AvrPtoB phosphorylated residues identified by mass spectrometry were analyzed for their conservation across 27 AvrPtoB homologs from P. syringae. The translated sequence of AvrPtoB was used in BLAST searches against the NCBI nr database on January 5th, 2020 (Pseudomonas syringae group, taxid: 136849; default parameters otherwise) to identify homologs. Hits were filtered based on a minimum 65% sequence identity, a minimum 85% query coverage and an E-value of 0. Sequences were downloaded and aligned using Genious alignment (default parameters).The percentage of identity is shown by gradient white to black (white means less than 60% similar, gray means 60-80% similar, dark gray means 80-100%, and black means 100% similar). AvrPtoB phosphorylation sites identified in vivo are shown with arrows.
Supplemental Figure 6. AvrPtoB S258 is required for virulence in tomato andArabidopsis. Related to Figure 3.
(A) Bacterial populations in the susceptible tomato genotype Rio Grande 76S five days post-inoculation with P. syringae pv. tomato DC3000ΔavrPtoΔavrPtoB (−/−) variants. DC3000 −/− carrying empty vector (EV) or plasmids expressing wild-type AvrPtoB, AvrPtoBS258A, AvrPtoB1-307, and AvrPtoB1-307S258A were dip inoculated on tomato at a concentration of 2 × 108 CFU mL−1 with 0.005% Silwet. LOG CFU/cm2 = log colony-forming units per cm2 of leaf tissue. Data are means ± SD (n ≥ 6 plants). Different letters indicate significant differences (one-way ANOVA, Tukey’s test, p < 0.05).
(B) Bacterial populations in Arabidopsis Col-0 three days post-inoculation with DC3000 −/− variants. DC3000 −/− variants as described in (A) were syringe infiltrated in Col-0 at a concentration of 2 × 105 CFU mL−1. Data are means ± SD (n = 4 plants for day 0, n = 6 plants for day 3). Different letters indicate significant differences (two-way ANOVA, Tukey’s test, p < 0.05).
(C) Expression of AvrPtoB and its derivatives from DC3000 −/−. Bacteria were grown in minimal media at 18 ℃. Anti-HA western blotting was performed to detect the expression of AvrPtoB in bacterial pellets.
Acknowledgements
The authors thank Sheng Luan (University of California, Berkeley) for providing clones of different SnRK members and Michelle Salemi (University of California, Davis) for assistance in mass spectrometry analyses.
References




