

Abstract
Empirical studies on the promiscuous species of Drosophila revealed that the laboratory evolution of resistance to a certain type of environmental stress can impact the ability of the organism to resist other kinds of stresses. The mechanisms of resistance to a particular stress are specialized and costly, then, mechanisms of resistance to other stresses can be negatively affected. However, it is also possible that at least a part of the stress resistance mechanisms is generic. With this premise we aimed to understand whether increased resistance to a cold stress can increase resistance to other types of stresses.
To address this issue, we used populations of Drosophila melanogaster (D. melanogaster) that have been selected for 57-71 generations for increased resistance to cold shock (−5°C for one hour). We subjected the selected (FSB; selected for cold shock resistance, derived from BRB population) and control FCB; cold shock control, derived from BRB population) populations to a variety of environmental stresses such as cold shock, heat shock, starvation, desiccation and bacterial infection. We found that the compared to FCB populations, FSB populations had higher resistance to heat stress in terms of adult survivorship and mating ability post cold or heat shock. Desiccation resistance was observed higher in FSB females compared to FCB females but no such difference was found in males. We observed that FSB populations had lower starvation resistance relative to FCB populations. There was no difference between FSB and FCB populations in their ability to survive post bacterial infection. Our findings suggest that resistance to heat stress and desiccation (in females) are positively correlated with increased resistance to cold shock. However, resistance to starvation was negatively correlated with increased resistance to cold shock.
1. Introduction
Empirical evolution of resistance to an environmental stress may confer an advantage or a disadvantage with respect to resistance to other types of environmental stresses. A large number of earlier studies have suggested that resistance to multiple stresses (such as desiccation and starvation, high and low temperature, starvation and cold temperature) might be correlated (Nghiem et al., 2000, Hoffmann et al., 2003 and Bubliy and Loeschcke, 2005). Other studies have investigated, if there are certain universal mechanisms that allow organisms to concurrently increase resistance to multiple environmental stresses (reviewed in Hoffmann and Parsons, 1991). Multiple studies have documented increased cross-tolerance in insects indicating that either the underlying mechanisms of resistance to these stresses are common or that there are strong genetic correlations between resistance traits. Such positive correlation has been found between resistance to cold and desiccation as well as between resistance to heat and desiccation (Bayley et al., 2001, Wu et al., 2002, Phelan et al., 2003, Bubliy and Loeschcke, 2005). Cross-tolerance with respect to high and low temperature stress has also been explored. For example, exposure to mild desiccation can increase cold tolerance in the springtail, Folsomia candida (Bayley et al., 2001), house flies subjected to anoxic conditions at 27°C show greater tolerance when exposed to −7°C (Coulson and Bale, 1992). Positively correlated responses in stress resistance traits have also been observed in laboratory selection studies. For example, Bubliy and Loeschcke (2005) observed increased cold stress resistance in lines selected for resistance to heat stress or desiccation stress. They also observed increased desiccation resistance in lines selected for resistance to heat knock down. Chill-coma recovery, cold resistance and desiccation are known to be positively correlated (Sinclair et al., 2007 and Anderson et al., 2005).
Alternatively, mechanisms required to adapt to a specific type of stress might conflict with mechanisms required to adapt with other kinds of stress, thereby leading to trade-offs across stress resistance traits (Kellett et al., 2005 and Overgaard et al., 2006). For example, Hoffmann et al. (2005) showed that flies selected for starvation resistance have decreased resistance to cold, whereas those selected for increased cold resistance show decreased starvation resistance. Quite often though, the relationship between resistances to various stresses seems to be complex. Bubliy and Loeschcke (2005) found a positive correlation between resistance to cold stress and desiccation. However, Sinclair et al. (2007), found no change in cold resistance in populations selected for desiccation resistance. Though it has been suggested that resistance to the multiple stresses that insects commonly encounter in nature (such as temperature extremes, desiccation etc) should be positively correlated, the evidence for such correlations is variable (Ring and Danks, 1994).
In this study, our major goal was to assess whether increased resistance to cold shock leads to correlated response with other environmental stresses such as resistance to desiccation, starvation, heat shock, and pathogenic challenge. Our study consisted of 10 populations of D. melanogaster (5 selected populations and 5 control populations), and experiments were conducted over 57-71 generations of selection.
We specifically addressed the following questions:
Does egg viability and reproductive behavior post heat shock evolve as a correlated response to increased resistance to cold shock?
Does adult survival under starvation, desiccation, heat shock, cold shock, and resistance to pathogenic challenge evolve in the selected populations of D. melanogaster?
2. Materials and Methods
2.1 Stock populations
2.1.1 Blue Ridge Base line population
Maintenance and derivation of Blue Ridge Base line populations have been thoroughly described in our previous study (Singh et al., 2015).
2.1.2 Derivation and maintenance of selected and control populations
Derivation and maintenance of selected (FSB; selected for cold shock resistance (non-lethal cold shock of −5°C for one hour), derived from BRB population) and control (FCB; cold shock control, derived from BRB population) populations have been explained in detailed in Singh et al. (2015).
2.2 Experimental protocol
2.2.1 Standardization of flies
To control over the non-genetic parental effects for selected and control populations (Rose 1984 and Crill et al., 1996, Singh et al. 2015, Singh et al. 2016a), all the populations (FSB1-5 and FCB 1-5) were passed through one generation common laboratory rearing conditions as described below, before any experimental egg collection. During this generation, the FSB populations were not imposed selection pressure (−5°C for one hour). This process known as ‘standardization’ and the flies maintained in this manner are known as ‘standardized flies’. In order to standardize, eggs were cultured from each of the FSB and FCB stock populations. Eggs from each of FSB and FCB population were transferred into culture vials containing standard banana-yeast-jaggery (here-to-forth referred to as “standard food”) at a density of 70 eggs per vial. For each of the FSB and FCB populations, 20 such vials were set up. These vials were incubated at standard laboratory conditions (25°C temperature, 50-60% RH, 12:12hours cycle from day to night on standard food). On the 12th day after egg collection, the flies from a specific population were shifted into a Plexiglas cage provided with standard food. In order to collect experimental eggs, on the 13th day after egg collection, a fresh standard food plate was given and the flies were allowed to oviposit for 6 hours. Adults emerging from these vials (i.e., the progeny of the standardized flies) were used for experimental assays. All the experiments done in the present study were conducted over 57 to 71 generations of selection.
2.2.2 Cold shock
Flies were subjected to cold shock as explained by Singh et al., (2015). Briefly, on 12th day after egg collection (2-3 days post eclosion), flies were transferred to clean dry glass vials (25mm diameter × 90mm height) at a density of 50 individuals per vial (in mixed sex groups or single sex groups as per the assay’s requirements). The cotton plug was pushed deep into the vial such that the flies were allowed to stay in the bottom of the vial (one third of the total volume of the vial which is roughly 25mm diameter × 30mm length of the vial). The vials were then shifted in ice-salt-water slurry maintained at −5°C and kept for one hour. Post cold shock, the flies were then immediately transferred to Plexiglas cages (14cm length × 16cm width × 13cm height) at a density of 100 pairs (100 males and 100 females) per cage. The cage was provided with a Petri plate having standard food.
2.2.3 Heat shock
Flies were handled in a similar way as described above for the cold shock treatment with the exception that experimental flies were subjected to 37.5°C in water bath for one hour (instead of being exposed to −5°C). After heat shock, flies were immediately transferred into Plexiglas cage provided with standard food plate.
2.2.4 No shock
In this treatment flies were also handled in an identical way as described in the cold shock treatment (see above) with the an exception that vials containing flies were placed in a water bath maintained at 25°C temperature for one hour. Following this, the flies were immediately transferred into Plexiglas cage provided with food plate.
2.3 Experiment 1: Effect of heat or cold stress on the mating ability and egg viability
In one of our previous studies we observed that both mating and egg viability are influenced by cold shock. We found that populations of D. melanogaster selected against cold shock had higher egg viability and mating frequency relative to their control populations (Singh et al., 2015). In this experimental setting, we wanted to investigate whether egg viability and mating frequency are also correlated with heat shock like with cold shock. After 60 generations of selection, experimental flies were raised followed by one generation of standardization as described above. Twelve vials of fixed density of 70eggs/vial were established for each FSB and FCB populations from the respective standardized populations. On 12th day (by the time almost all flies had emerged and mated) after egg collection, 4 vials were randomly assigned to one of the following three treatments.
Cold-shock: Both males and females from a given population were imposed to cold shock (as described above in cold shock treatment) and following this, flies were immediately transferred into a Plexiglas cage at a density of 100 mating pairs per cage.
Heat-shock: Both males and females from each FSB and FCB populations were subjected to (heat shock as describe above) and after that flies were quickly transferred to the Plexiglas cage at a density of 100 mating pairs per cage.
No-shock: Both males and females from each of FSB and FCB populations were subjected to a temperature of 25°C for one hour (as described above) and subsequently transferred to a Plexiglas cage at a density of 100 mating pairs per Plexiglas cage.
2.3.1: Assayed the egg hatchability at two points
0-hour post cold shock/heat shock/no shock and
24 hours post cold shock/heat shock/no shock.
We selected these two time points because of the following reason; first, egg viability measurement at 0 hours post shock would demonstrate the immediate impact of treatment. Second, in their normal maintenance cycle eggs are collected from the flies 24 hours after cold shock to start the next generation and therefore it is crucial time point to the fitness of the flies. To measure the egg viability, 0 hours or 24 hours post cold shock/ heat shock/ no shock, a fresh standard food plate was kept in the cage for flies to lay eggs for 6 hours. A group of 200 eggs were transferred to a Petri plate containing 1.2% agar from the standard food plate. Following this, these plates were incubated at standard laboratory conditions as described above for 48 hours, after which, the numbers of hatched eggs were counted as a measurement of the egg viability.
2.3.2: Assayed the mating ability
We monitored the total number of mating for all the three treatments. Once the flies were transferred to Plexiglas cages, we observed the cages every half an hour and recorded the total number of mating pairs. We followed the protocol of monitoring mating pair as we described previously (Singh et al., 2015). We tracked mating pairs every 30 minutes intervals until 36 hours post treatment (cold shock/heat shock/no shock). We then summed the number of mating pairs across all the observations for a given cage to obtain an estimate of the total number of mating. The total number of mating pairs per cage was used as the unit of analysis.
2.4 Experiment 2: Effect of heat or cold stress on adult survivorship
We wanted to check whether adult survivorship has changed in the selected populations (FSB) after 63 generations of selection. We also wanted to probe whether the population selected for increased resistance to cold shock could show cross-tolerance to other stress i.e. heat shock. We already knew from previous studies that both cold shock and heat shock influence adult survivorship (Tucic, 1979, Chen and Walker, 1993, Rohmer et al., 2004 and MacMillan et al., 2009). We collected eggs to generate experimental flies after 63 generations of selection following one generation of standardization. Twenty five vials (70eggs/vial) were set up for each FSB and FCB population. Virgin males and females were collected on the 9-10th days post egg collection from the peak of eclosion using mild CO2 anesthesia and were housed separately in single sex vials at density 10 flies/vial. On the 12th days post egg collection 50 flies were transferred into an empty glass vial and cotton plug was pushed deep into the bottom one third of vial to allow flies to stay in a restricted space. Followed by vials were randomly assigned one of the following treatments:
Cold shock: Vials containing female flies were exposed to −5°C in ice-water-salt slurry for one hour (as described above). Male flies were handled identical manner as described above except that they were exposed to −5.6°C (we used −5.6°C to get at least 50% mortality post cold shock) instead of −5°C for one hour in ice-water-salt slurry.
Heat shock: Both male and female flies were handled similar ways as described above for heat shock treatment in the previous experiment except that the temperature was different in the following manner because male were more susceptible at higher temperature:
Vials containing male flies were exposed to 39°C temperature for 1 hour in water-bath.
Vials containing female flies were subjected to 39.2°C temperature for 1 hour in water-bath.
Three replicate Plexiglas cages of 100 flies per cage density were set up for each treatment, population, block and sex. We measured adult survivorship at 24 hours post stress. We selected this time point because 24 hours post cold shock is the time that eggs are collected from the flies to start the next generation in their normal maintenance cycle and it is hence directly relevant to the fitness of the flies. Twenty four hours post cold shock, dead flies (if any) were aspirated out of the cage and counted. Mean percentage mortality of each cage was used as the unit of analysis.
2.5 Experiment3: Starvation Resistance
Sex specific starvation resistance assay was carried out after 57 generations of selection. Experimental eggs were collected from standardized flies and reared them at density of 70eggs/vials in standard food at standard laboratory condition as aforementioned. Assay was carried out using the method described in Kwan et al. (2008) with minor modifications. Ten vials were established for each FSB and FCB populations. Virgin flies were collected on 9th and 10th days during peak of eclosion, employing mild CO2 anesthesia. Males and females were held separately at a density of 10 flies per vial containing fresh standard food. On the 12th days, flies were transferred from food vials to 1.24% agar vials (Kwan et al., 2008). For these assay, seven replicate vials containing 10 flies were set up for each sex and population (FSB and FCB). Flies were transferred into a fresh agar vial (1.24%) every alternate day until the last fly in a given vial died. Mortality was recorded every four hours. Mean time of mortality was computed for each vial and was used as the unit of analysis.
2.6 Experiment 4: Desiccation Resistance
Sex-specific desiccation resistance assay was performed for each of the FSB (1-5) and FCB (1-5) populations. After 57 generations of selection, experimental flies were raised from standardization flies of FSB and FCB populations. Ten vials containing eggs at a density of 70eggs/vial were set up for each population. On 9-10th day post egg collection, virgin flies were collected using light CO2 anesthesia and were dispensed in vials provisioned with standard food at a density of 10 virgin males or females in a vial. On 12th day, flies were transferred from food vials to food-less glass vials containing ~6g of silica gel (desiccant). The flies were separated from the silica gel by a thin layer of cotton. The open end of each vial was sealed with Parafilm (Kwan et al., 2008). Seven replicate vials were set up at a density of 10 flies per vial for each population. Mortality was monitored every half an hour until the last fly died. Mean time to death was computed for each vial and was used as the unit of analysis.
2.6 Experiment 5: Resistance to a bacterial infection
We investigated whether flies selected for resistance to cold stress have also evolved resistance to bacterial infection as a correlated response after 70 generations of selection. To raise the experimental flies, eggs were collected from standardized flies with fix density 70eggs/vial containing 6 ml of standard food. Five vials were set up for each population. On 9-10th day post egg collection, virgin males and females were sorted from the peak of eclosion using light CO2 anesthesia at very young stage (approximately 4 hours post eclosion) and housed in vials provisioned with 2ml of standard food at a density of 10 individuals per vial. On day 12 post egg collection, flies of known age (2-3 days old as adult), population regime and sex (see below for details) were lightly anaesthetized using CO2. Fifty five to sixty flies of each sex for each population were infected by pricking the thorax with a Minutien pin (0.1 mm, fine Science Tools, Foster City, CA, USA) dipped in the bacterial slurry (Staphylococcus succinus subsp. succinus strain PK-1 is a natural pathogen of D. melanogaster, which we isolated from wild captured Drosophila (Singh et al., 2016b)) of OD600nm2. For sham infection, the pin was dipped in 10mM MgSO4 prior to pricking the lateral thorax of the flies. The number of dead flies in each vial was tracked at every 3 hrs intervals till 30 hours post infection. After this period, vials were observed every hour till 80 hrs post infection. Proportion of flies that survived the infection was calculated for each population and was used as the unit of analysis.
2.7 Statistical analysis
2.7.1 Experiment 1: Effect of heat or cold stress on the mating ability and egg viability
Egg viability data from Experiment 1 was analyzed using four-factor mixed model analysis of variance (ANOVA) with selection regime (FSB vs. FCB), treatment (Cold shock/ no shock/ heat shock) and period (0 hour vs. 24hours) as fixed factors crossed with block (1-5) as random factor. All multiple comparisons were performed employing Tukey’s HSD. Mating number data from Experiment 1 was analyzed using three factor mixed model ANOVA with selection regime (FSB vs. FCB) and treatment (Cold shock vs. no shock/ heat shock) as fixed factors crossed with block (1-5) as random factor. All multiple comparisons were performed using Tukey’s HSD.
2.7.2 Experiment 2, 3, 4 and 5
Survivorship post heat or cold stress, starvation resistance, desiccation resistance, mortality post bacterial infection data from Experiment 2, 3, 4 and 5 respectively were analyzed using two-factor mixed model ANOVA treating selection regime (FSB vs. FCB) as a fixed factor crossed with random block (1-5). We also analyzed the mortality post bacterial infection data from experiment 5 using Kaplan-Meier method. All the analyses were done at p = <0.05 level of significance using Statistica (for Windows, version 10, StatSoft).
3. Results
3.1 Experiment 1: Egg viability and mating ability post heat shock or cold shock
Our findings indicate that egg viability evolves in response to selection. We found significant main effect of selection and treatment on the egg viability (Table 1). We also found a two-way interaction between selection (FCB and FSB) and treatment (Cold shock/heat shock/no shock) (Table 1). Multiple comparisons employing Tukey’s HSD suggested that egg viability in no shocked treatment was more than 90% and there was no significant difference between FCB and FSB populations (Figure 1). Cold shock or heat shock treatment significantly reduced egg viability (Figure 1). At 0th hour post cold shock, egg viability was found to be very low (approximately 2-3%) and post heat shock egg viability was also very low which is about 5-10%. However, difference between FSB and FCB population was not significant (Table 1). Multiple comparisons using Tukey’s HSD suggested that FSB population had greater egg viability when compared to FCB population 24 hours after cold shock (~41%) or heat shock (~7%) (Figure 1, Table 1).
Summary of results of a four-factor mixed model ANOVA on egg viability with selection regime (FSB and FCB), period (0 hour and 24 hours) and Treatment (Cold Shock, Heat Shock, No Shock) as the fixed factors crossed with blocks (1-5) as random factor. p-values in bold are statistically significant.

We measured egg viability at 0 and 24 hours post heat/cold shock. Open bars represent FSB and closed bars represent FCB populations. Viability of eggs from No-shock treatment was high with no difference between FCB and FSB populations. At 0 hours post cold shock, viability of eggs from the cold-shock and heat-shock treatment was very low and not different between FCB and FSB populations. However, 24 hours post cold shock, egg viability improved and the FSB populations had significantly higher egg viability than the FCB populations.
We observed significant main effect of selection and treatment on the number of mating pairs. We also found a statistically significant two way selection × treatment interaction (Table 2). Multiple comparisons using Tukey’s HSD indicated that flies subjected to cold shock treatment show nearly twice as many mating pairs when compared to flies subjected to heat shock or no shock treatment (Figure 2). However, in case of neither shock treatment FSB populations had about 7% more mating pairs compared to FCB populations (Figure 2).
Summary of the results of a three-factor mixed model ANOVA on mating number with selection regime (FSB and FCB) and treatment (cold shock, heat shock or no shock) as fixed factors crossed with blocks (1-5) as random factor. For mating number the sum of all observed matings until 36 hours post treatment for each population was used as the unit of analysis. p-values in bold are statistically significant.
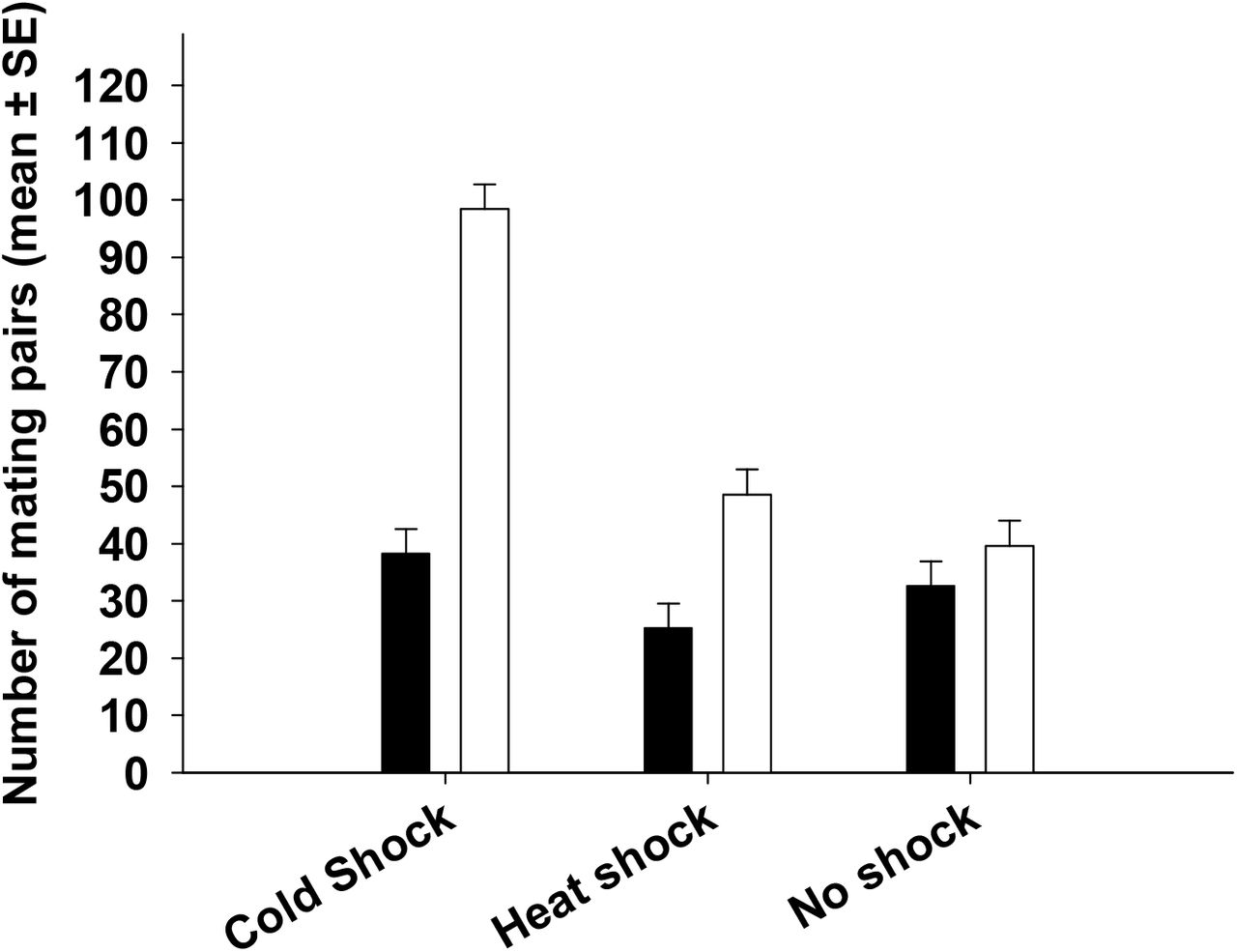
We assayed mating frequency post heat or cold shock (0-36 hours). Open bars represent FSB and closed bars represent FCB populations. The number of mating pairs observed in FSB flies from cold-shock and heat shock treatment was significantly higher relative to FCB populations.
3.2 Experiment 2: Mortality post cold or heat shock
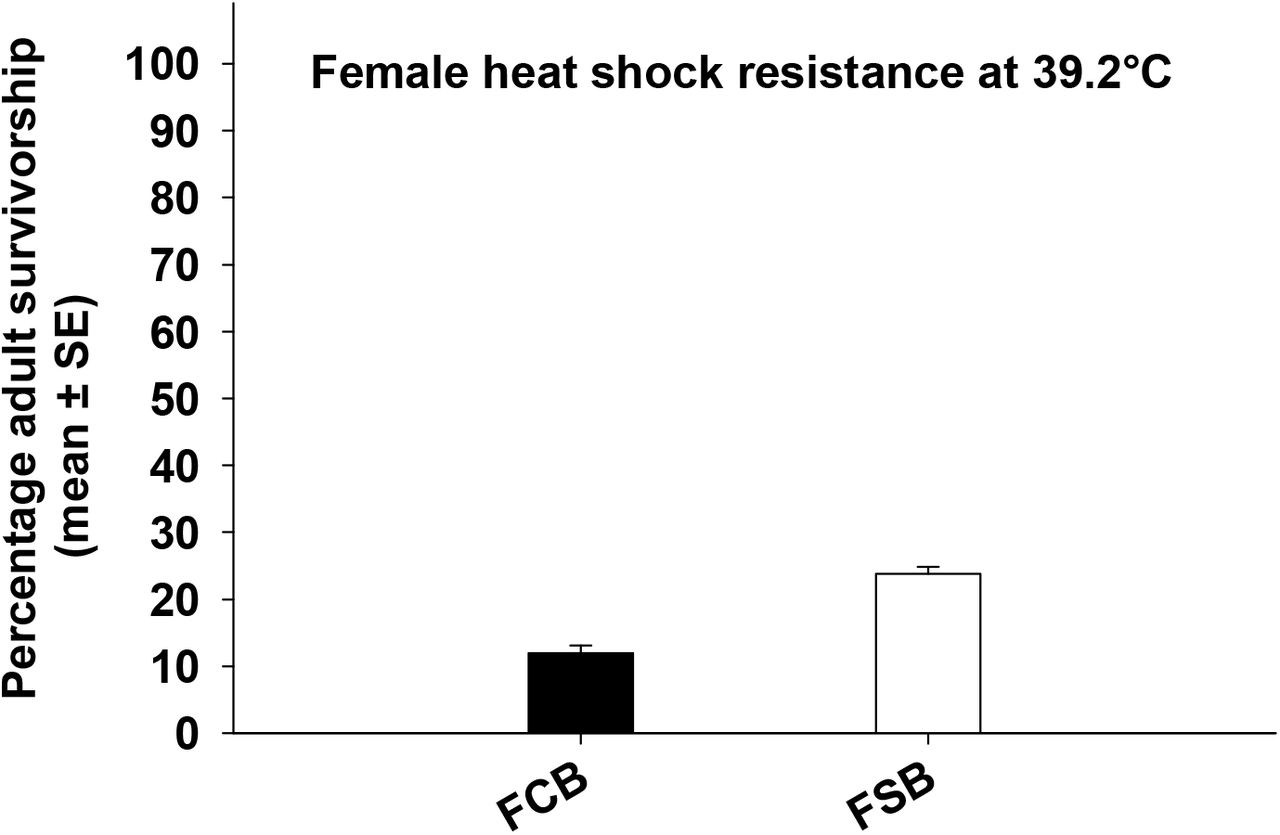
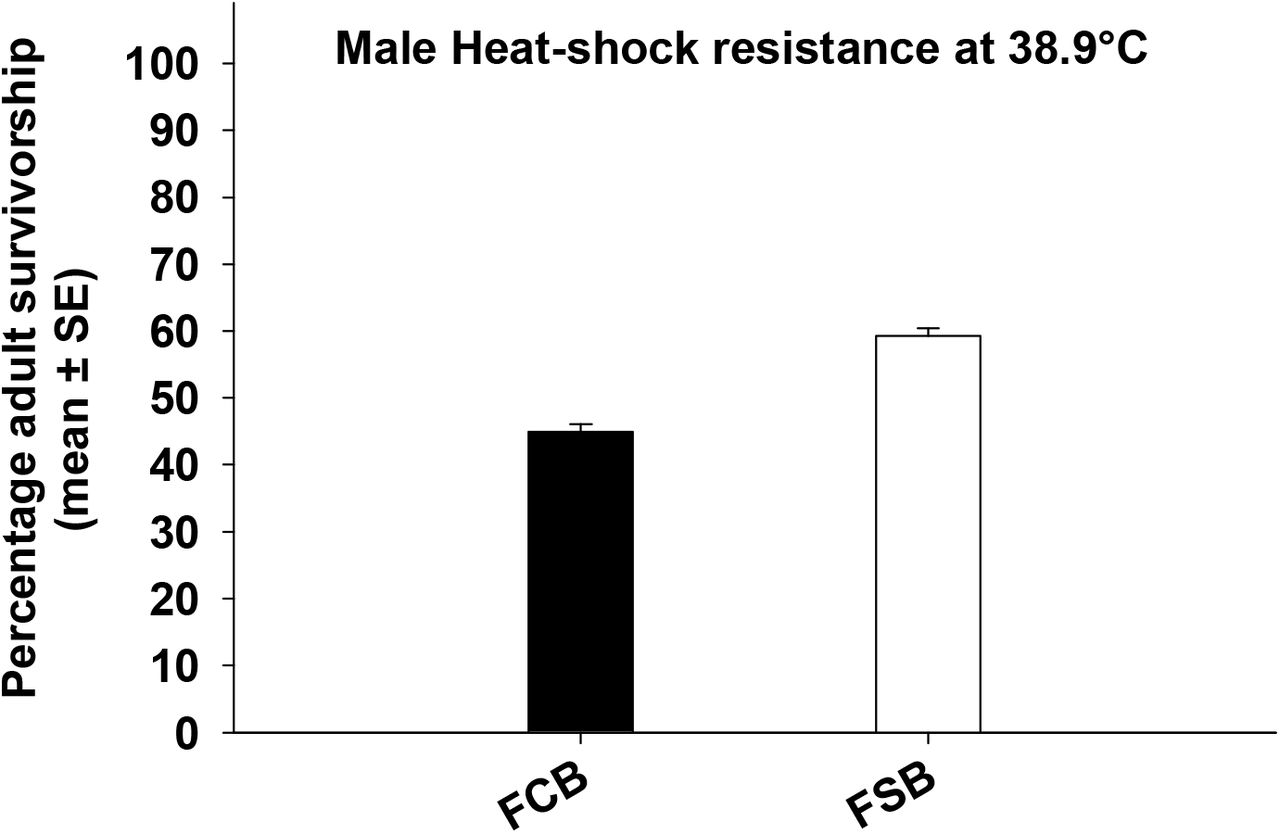
We quantified the effect of selection on virgin male and female mortality post cold/heat shock and observed a significant effect of selection on male and female mortality post cold shock (Table 33a and 3b, Figure 3a and 3b). In case of males, 24 hours post cold shock FSB populations had about 35% lower mortality compared to FCB populations (Figure 3a). In case of females, 24 hours after receiving cold shock, FSB populations had approximately 29% lower mortality than FCB population (Figure 3b). These results indicate that the flies from FSB population have evolved to significantly lower mortality relative to FCB population.

FSB populations had significantly higher survivorship compared to FCB populations.

FSB populations had higher survivorship relative to FCB populations.
Twenty-four hours post heat shock in males, we found significant effect of selection and block on male mortality (Table 3c). A significant effect of selection suggested that FSB populations had lower mortality (about 15%) compared to FCB populations (Figure 3c). For females, we found a significant effect of selection on female mortality post heat shock (Table 3d). A significant effect of selection indicated that FSB population had approximately 11% lower mortality compared to FCB population (Figure 3d).
Summary of the results of two-factor mixed model ANOVA on mortality in male (a) and in female (b) post cold shock and on mortality in male (c) and in female (d) post heat shock with selection regime (FSB and FCB) as the fixed factor crossed with block (1-5) as random factor. p-values in bold are statistically significant.

FSB populations had higher survivorship relative to FCB populations.
FSB populations had higher survivorship relative to FCB populations.
3.3 Experiment 3: Evolution of starvation resistance
We found that starvation resistance was negatively correlated with resistance to cold stress. Starvation resistance was significantly lower in the FSB populations compared to FCB populations (Table 4a and 4b, Figure 4a and 4b). We also observed a significant main effect of selection and block on starvation resistance in males (Table 4a) and in females (Table 4b). Compared to FCB populations, resistance to starvation (mean time to death) in FSB males is lower by approximately 15 hours and in FSB females by about 12 hours (Figure 4a and b).
Summary of the results from two-factor mixed model ANOVA on resistance to starvation in Males (a) and Females (b) with selection regime (FSB and FCB) as the fixed factor crossed with random blocks (1-5). Mean time to death in hours for each vial was used as the unit of analysis. p-values in bold are statistically significant.
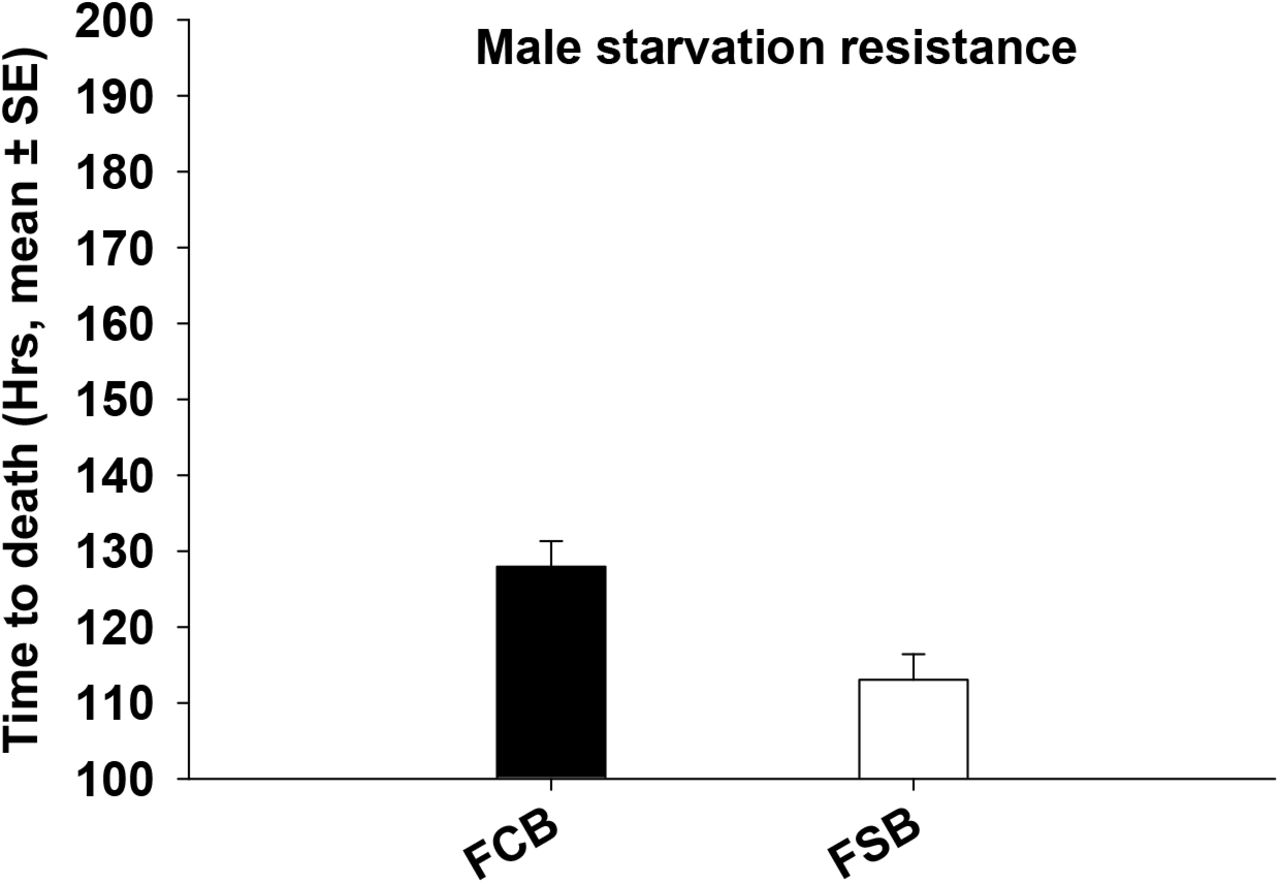
FSB populations had lower starvation resistance relative to FCB populations.

FSB populations had lower starvation resistance relative to FCB populations.
3.4 Experiment 4: Desiccation resistance
We found significant main effect of selection on female desiccation resistance (Table 5b). FSB females have higher resistance to desiccation (mean time to death) by about one hour and ten minutes as compared to FCB populations (Figure 5b). However, we did not observe an effect of block or any two-way interaction (i.e. selection × block). In case of males, we did not find any effect of selection or block on desiccation resistance (Table 5a).
Summary of results from a two-factor mixed model ANOVA on resistance to desiccation in Males (a) and Females (b) using Selection regime (FCB and FSB) as fixed factor crossed with random Block (1-5). Mean time to death in hours for each vial was used as the unit of analysis. p-values in bold are statistically significant.

We did not find any significant main effect of selection on mean time to death.

We found significant main effect of selection on desiccation resistance, indicating that FSB populations had higher desiccation resistance relative to FCB populations.
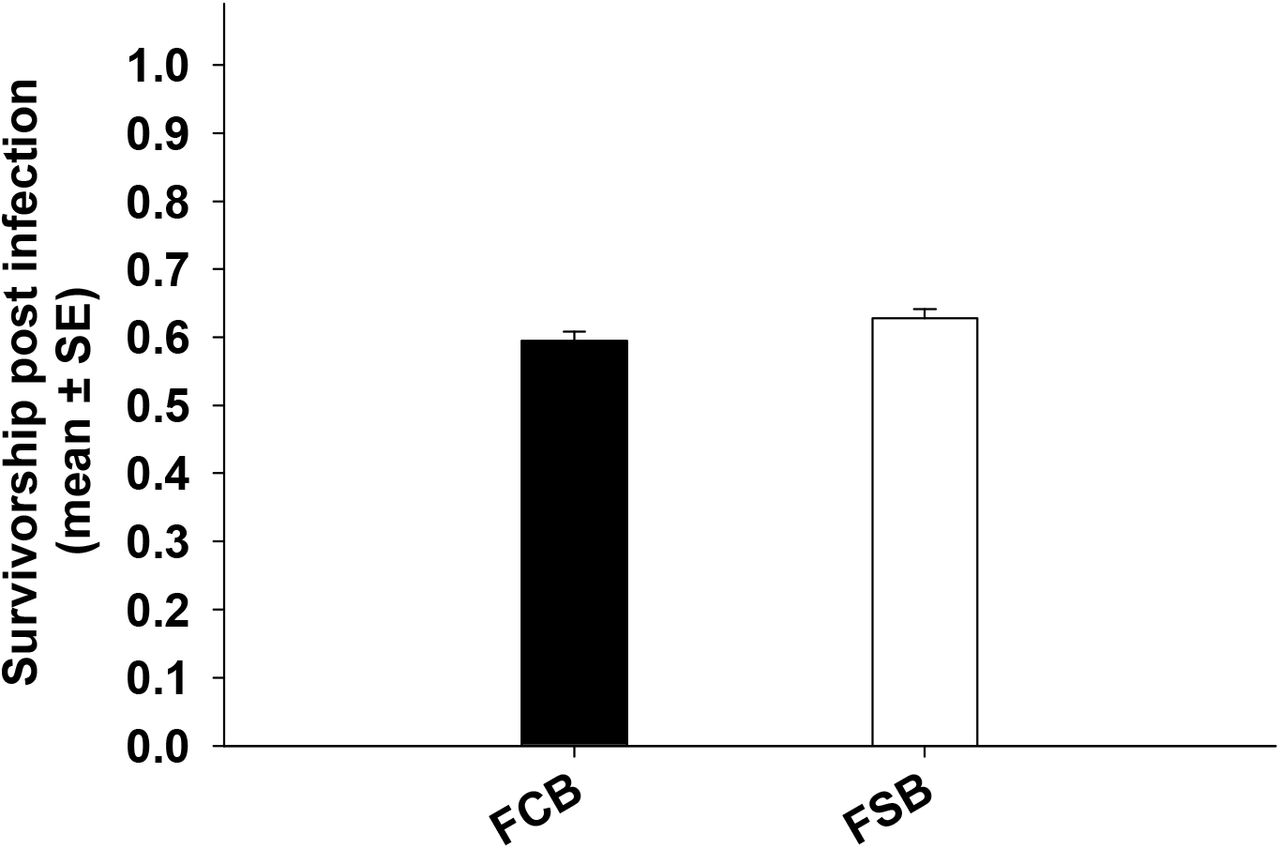
3.5 Experiment 5: Resistance to bacterial infection
We did not find any significant main effect of selection on male or female survivorship post pathogenic challenge (Table 6a and 6b). Survivorship in case of males in both FSB and FCB population is about 58-63% (Figure 6a) and in case of female survivorship was about 62-65% (Figure 6b). The results were similar even when data were analyzed using Kaplan-Meier method.
Summary of results of a two-factor mixed model ANOVA on proportion of survivorship post bacterial infection in males (a) and females (b) with selection regime (FSB and FCB) as the fixed factor crossed with blocks (1-5) as random factor. p-values in bold are statistically significant.

We did not noticed significant difference between FSB and FCB males survivorship post infection.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
We did not observe any significant difference between FSB and FCB females survivorship post infection.
4. Discussion
In this study, our aims was to explore the cross-tolerance in the lines that have evolved for the early recovery in context of reproductive traits such as egg viability, mating frequency, male mating ability, mating latency, sperm offence ability and progeny production (Singh et al., 2015 and Singh et al., 2016a) post cold stress. In our present experimental evolution study, we measured resistance to starvation, desiccation, heat shock and cold shock, resistance to challenge with natural pathogen (Staphylococcus succinus subsp. succinus strain PK-1) D. melanogaster in the populations selected for resistance to cold shock (Singh et al., 2016b). We found higher mating frequency and adult survivorship in the selected populations relative to their control populations after the subjection of heat shock or cold shock. Egg viability has increased in the selected populations post cold shock relative to control populations as we have seen in our previous study (Singh et al., 2015). Desiccation resistance has increased in females of selected populations indicating that selection for single type of environmental stress leads to improve resistance towards other stress also. However, in case of starvation resistance, we found that the selected populations had lower starvation resistance relative to control populations suggesting that increased resistance to cold shock is negatively correlated with starvation resistance. We discuss each of these observations below in more detail.
At 0 hour post cold shock, we found approximately 95-97-5% reduction in egg viability. This could be because below zero temperatures cause sperm mortality in male seminal vesicle, female seminal receptacle and spermathecae (Lefevre and Jonson, 1962 and Novitski and Rush, 1949). This result is in line with several other studies that have observed reduced egg viability and sterility in insects upon exposure to extremes of temperature (Arbogast, 1981, Coulson and Bale, 1992, and Singh et al., 2015). However, we found greater egg viability in the FSB populations compared to FCB populations 24 hours after the subjection cold shock as we have found previously (Singh et al., 2015). There are a number of possible explanations for increased egg viability at 24 hours post cold shock: (a) The selected populations could be better at protecting their stored sperm/eggs from damage caused by heat or cold shock. For instance, Collett and Jarman (2001) have shown that D. pseudoobscura females can store the sperm up to 6 months during cold environment. These stored sperm can be used to fertilize ova in warm environment. However, in our previous study we have shown that D. melanogaster females from the selected populations relative to their control populations does not store the fertilized egg or sperm we had measured this by assessing the egg viability at different time points after subjection of the cold shock to the mated females, post cold shock females were not allowed to accessing of males (Singh et al., 2015).
(b) The selected populations mate more after heat or cold shock to enhance egg viability. A number of studies documented that high and low temperature have an impact on the mating behavior (Schnebel and Grossfield, 1984, Chakir et al., 2002, David et al., 2005, Dolgin et al., 2006, David, 2008 and Zhang et al., 2013, Singh et al 2015, 2016). However, very few studies have addressed the effect of cold shock on mating behavior (Singh et al., 2015, Singh et al., 2016a and Singh et al., 2016b). In FSB populations, the frequency of mating has increased post heat or cold shock compared to FCB populations. Hence, it is likely that increased mating post heator cold shock is largely responsible for increase in egg viability. While the pattern of increased mating correlated with increased egg viability post cold shock had been observed in our previous study (Singh et al., 2015), it is interesting that this pattern is seen even under heat stress. This finding indicates that probably some of the mechanisms underlying resistance to heat and cold stress might be common (such as expression of heat shock proteins). This also forms an example of positive correlation between resistances to two stressors.
We found that FSB populations have lower mortality relative to FCB populations over 24 hours post cold shock. It is important to note that during regular maintenance regime, adult mortality due to cold shock is negligible. Our results indicate that FSB population have evolved the ability to withstand cold temperatures in terms of reduced adult mortality along with their ability to maintain higher egg viability after shock at a temperature of −5°C. Multiple laboratory selection studies show increased adult survivorship as a correlated response to selection for cold tolerance (Anderson et al., 2005, MacMillan et al., 2009, Tucic, 1979 and Chen and Walker, 1993). In our previous study (Singh et al., 2015), we found that mortality post cold shock was negligible. However, in the present study, mortality post cold shock is substantial. These results seem quite contradictory. There are several possible explanations. First, the populations have evolved for first, number of generations between these two experiments. Second, in the current study, the flies were virgins when subjected to cold shock whereas in the previous study, the flies had already mated by the time they were subjected to cold shock. Third, in the present study, the flies were moved into a fresh food vial soon after eclosion while in the previous study, the flies remained in the culture vials (with old, spent food) for two days after eclosion. We did a small experiment (data not shown) to differentiate between possibilities two and three. We used a factorial combination of mating status and food type to dissect out the effects. The experimental design was as follows-
The results from this experiment indicate that flies maintained on new food soon after eclosion have higher mortality than flies maintained on old food soon after eclosion.
More interestingly, the FSB populations also showed lower mortality post heat shock compared to FCB populations. The present literature depicts some disagreement with regards to cross-resistance between cold and heat stress (reviewed in Hoffmann et al., 2003). Anderson et al. (2005) and MacMillan et al. (2009) did not find correlated increase in heat shock resistance in populations of D. melanogaster selected for faster chill coma recovery or freeze resistance respectively. Our results are in agreement with those of Kristensen et al. (2007) who show that cold selected lines of D. melanogaster were more heat tolerant and vice versa. Previous studies in Drosophila along latitudinal clines suggest that there is a trade-off between heat and cold tolerance (Hoffmann et al., 2002). Our results suggest that heat and cold tolerance might be positively correlated in D. melanogaster. There could be multiple explanations for the superior survivorship of FSB populations post cold shock. (a) Chen and Walker (1994) report that cold selected lines have higher glycogen and total proteins relative to controls lines. Insects are known to store various sugars in order to tolerate cold temperatures (Ring and Danks, 1994, 1998). It is possible that the FSB populations have similarly altered resource storage in terms of carbohydrates, proteins or lipids. (b) Several studies have shown that there are several heat shock proteins that are expressed both during heat and cold stress. It is quite possible that at least some of these genes are expressed at a higher level in our populations. However, these genes are certainly not among the set that we analyzed for expression differences in our other experiment (Singh et al. Unpublished data).
Starvation resistance has decreased in populations selected for increased resistance to cold shock relative to control populations. Our findings are similar to those of MacMillan et al. (2009) and Anderson et al. (2005) who found lower starvation resistance in populations of D. melanogaster selected for increased resistance to cold shock. Interestingly, Bubliy and Loeschcke (2005) found decreased cold stress tolerance in populations of D. melanogaster selected for increased starvation resistance. Thus, across multiple studies, the correlation between starvation resistance and cold stress tolerance seems to be robust.
We found that desiccation resistance increased in females of the selected populations. Our findings are in line with results from other studies (Bubliy and Loeschcke, 2005 and Sinclair et. al., 2009,) which show that increased resistance to cold shock may lead to increased desiccation resistance as a correlated response. However, populations selected for desiccation resistance do not show increased cold tolerance (Sinclair et al., 2007). There is at least one common factor between cold and desiccation resistance that might explain their correlated evolution. Glycogen is known to act as cryoprotectant (Ramløv and Lee, 2000 and Holmstrup et al., 2002). Chippindale et al., (1998) showed that selection for increased desiccation resistance leads to increased glycogen content. Thus, increases in glycogen through selection on cold shock resistance could in principle lead to evolution of increased desiccation resistance. Other possible explanation to increased desiccation resistance could be body size in female, in our other related experiment we found that females from selected populations had higher body weight relative to females from control populations (data not shown). However, such increase, if any, is likely to be sex specific since we found no change in the desiccation resistance of FSB and FCB males.
In insects, cold stress can cause somatic injury to the gut and malphigian tubules. This can open up a way for the gut flora to enter the haemocoel and thereby cause an infection (Yi and Lee, 2003, MacMillan and Sinclair, 2011 and reviewed in Sinclair et al., 2013). Therefore, in our selected populations, immune activity can potentially evolve. However, we did not find any difference between FSB and FCB populations in their immunity against S. succinus subsp. Succinus strain PK-1. One possibility is that the immune response is elicited only in response to the gut flora. In Drosophila, evolution against a pathogen can be fairly specific and the host might not have increased immunity against other pathogens (Roxstrom◻Lindquist et al., 2004, Pham et al., 2007 and Mikonranta et al., 2014). Thus, in the present assay, where we use Staphylococcus succinus subsp. Succinus PK-1 as the pathogen, the appropriate immune response might not have been elicited.
5. Conclusions
In this experimental evolution study, we explored the cross-tolerance in the population of D. melanogaster selected for increase resistance to cold stress, we found that cold shock resistance was positively correlated with heat shock resistance, negatively correlated with starvation resistance and not correlated with pathogen resistance. More interestingly, cold shock was positively correlated with desiccation resistance only in the females. Thus, genetic correlations across traits, at least to some extent seem to be independent of each other and might even be sex-specific.
All authors have declared no conflicts of interest.
Acknowledgement
We are thankful to Daniel Promislow for kindly providing us the iso-female lines. We thank to Indian Institute of Science Education and Research Mohali for providing financial support for this work. Karan Singh thanks to Indian Institute of Science Education and Research Mohali, for financial support in the form Senior Research Fellowship. We also thank to Manas Arun Samant, Megha Treesa Tom, Karan Bhatt and Prakhar Gahlot for laboratory assistance.
Footnotes
Previous address: Indian Institute of Science Education and Research Mohali, Knowledge City, Sector 81, SAS Nagar, PO Manauli, Punjab 140306, India, Ph: 91 172 2240266, 2240124, Fax: 91 172 2240266, 2240124, E-mail: karan.iiser{at}gmail.com
References




