

SUMMARY
Neuromodulators promote adaptive behaviors in response to either environmental or internal physiological changes. These responses are often complex and may involve concerted activity changes across circuits that are not physically connected. It is not well understood how neuromodulatory systems act across circuits to elicit complex behavioral responses. Here we show that the C. elegans NLP-12 neuropeptide system shapes responses to food availability by selectively modulating the activity of head and body wall motor neurons. NLP-12 modulation of the head and body wall motor circuits is generated through conditional involvement of alternate GPCR targets. The CKR-1 GPCR is highly expressed in the head motor circuit, and functions to enhance head bending and increase trajectory reorientations during local food searching, primarily through stimulatory actions on SMD head motor neurons. In contrast, NLP-12 activation of CKR-1 and CKR-2 GPCRs regulates body bending under basal conditions, primarily through actions on body wall motor neurons. Thus, locomotor responses to changing environmental conditions emerge from conditional NLP-12 stimulation of head or body wall motor neuron targets.
Introduction
Neuromodulators serve critical roles in altering the functions of neurons to elicit alternate behavior. Disruptions in neuromodulatory transmitter systems are associated with a variety of behavioral and neuropsychiatric conditions, including eating disorders, anxiety, stress and mood disorders, depression, and schizophrenia.1–3 To achieve their effects, neuromodulatory systems may act broadly through projections across many brain regions, or have circuit-specific actions based on the GPCRs involved and their cellular expression. A single neuromodulator may therefore perform vastly different signaling functions across the circuits where it is released. For example, Neuropeptide Y (NPY) coordinates a variety of energy and feeding-related behaviors in mammals through circuit-specific mechanisms. NPY signaling may increase or decrease food intake depending upon the circuit and GPCR targets involved.4,5 Due to the varied actions of neuromodulators across cell types and neural circuits, it has remained challenging to define how specific neuromodulatory systems act in vivo to elicit alternate behavior. Addressing this question in the mammalian brain is further complicated by the often widespread and complex projection patterns of neuromodulatory transmitter systems, and our still growing knowledge of brain connectivity.
The compact neural organization and robust genetics of invertebrate systems such as Caenorhabditis elegans are attractive features for studies of neuromodulatory function. Prior work has shown that C. elegans NLP-12 neuropeptides are key modulatory signals in the control of behavioral adaptations to changing environmental conditions, such as food availability or oxygen abundance.6–8 The NLP-12 system is the closest relative of the mammalian Cholecystokinin (CCK) neuropeptide system and is highly conserved across flies, worms and mammals.9–11 CCK is abundantly expressed in the mammalian brain, however a clear understanding of the regulatory actions of CCK on the circuits where it is expressed is only now beginning to emerge.12–15 Like mammals, the C. elegans genome encodes two putative CCK-responsive G protein-coupled receptors (GPCRs) (CKR-1 and CKR-2), though, to date, direct activation by NLP-12 peptides has only been demonstrated for the CKR-2 GPCR.9–11,16 The experimental tractability of C. elegans, combined with the highly conserved nature of the NLP-12/CCK system, offers a complementary approach for uncovering circuit-level actions underlying neuropeptide modulation, in particular NLP-12/CCK neuropeptide signaling.
Sudden decreases in food availability or environmental oxygen levels each evoke a characteristic behavioral response in C. elegans where animals limit their movement to a restricted area by increasing the frequency of trajectory changes (reorientations), a behavior known as local or area-restricted searching (ARS). ARS responses during food searching in particular are rapid and transient. Trajectory changes increase within a few minutes after food removal, and decrease with prolonged removal from food (>15-20 minutes) as animals transition to global searching (dispersal).6–8,17–20 ARS is a highly conserved adaptive behavior and is evident across diverse animal species.21–27 NLP-12 signaling promotes increases in body bending amplitude and turning during movement,6,7 motor adaptations that are particularly relevant for ARS. Notably, nlp-12 is strongly expressed in only a single neuron, the interneuron DVA6 that has synaptic targets in the motor circuit and elsewhere28. Despite the restricted expression of nlp-12, there remains considerable uncertainty about the cellular targets of NLP-12 peptides and the circuit-level mechanisms by which NLP-12 modulation promotes its behavioral effects.
Here we explore the GPCR and cellular targets involved in NLP-12 neuromodulation of local food searching. Our findings reveal a primary requirement for NLP-12 signaling onto SMD head motor neurons, mediated through the CKR-1 GPCR, for trajectory changes during local searching. In contrast, NLP-12 signaling through both CKR-1 and CKR-2 GPCRs contribute to NLP-12 regulation of basal locomotion, likely through signaling onto body wall motor neurons. Our results suggest a model where NLP-12 signaling acts conditionally through CKR-1 and CKR-2 to coordinate activity changes across head and body wall motor circuits during transitions between basal and adaptive motor states.
Results
NLP-12/CCK induced locomotor responses require functional CKR-1 signaling
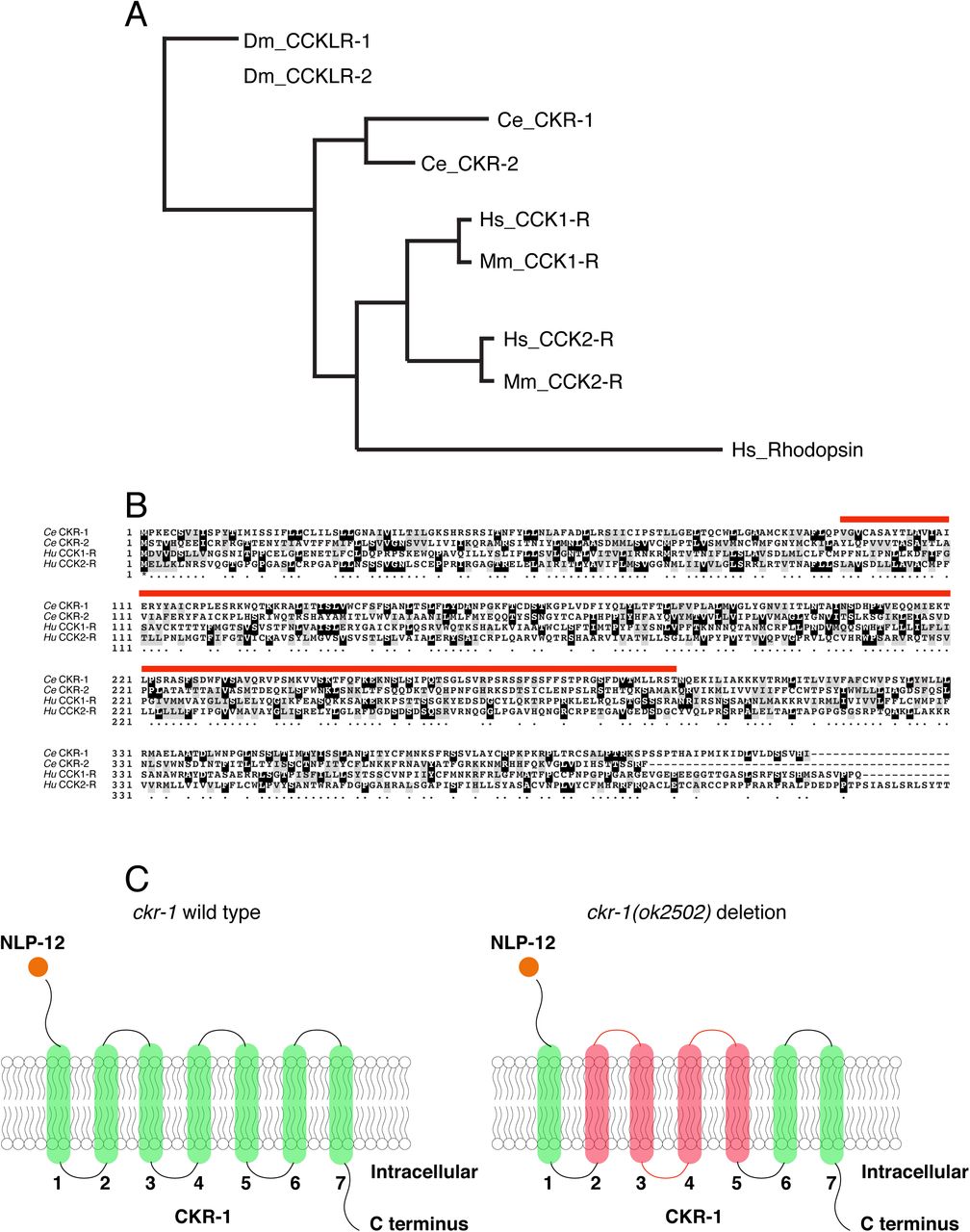
To decipher mechanisms underlying NLP-12 regulation of local food searching, we sought to identify genes required for NLP-12-mediated locomotor changes, in particular the G protein-coupled receptors (GPCRs) responsible for NLP-12 signaling. The C. elegans genome encodes two GPCRs that can be activated by NLP-12, ckr-1 and ckr-2 (Cholecystokinin-like Receptors 1 and 2), with close sequence homology to the mammalian Cholecystokinin receptors CCK-1 and CCK-2 (Fig. S1).9–11 Prior work demonstrated that NLP-12 activates CKR-2 in vitro9. Further genetic studies provided evidence that NLP-12 signaling mediates functional plasticity at cholinergic neuromuscular synapses through CKR-2 modulation of acetylcholine release from motor neurons.6,29,30 Surprisingly however, deletion of ckr-2 does not strongly affect local search behavior.6 As functional roles for the CKR-1 GPCR have not been previously described, we sought to determine whether CKR-1 may be acting either alone or in combination with CKR-2 to direct NLP-12 regulation of local searching. We first isolated a full-length ckr-1 cDNA identical to the predicted ckr-1 sequence. As expected, we found the ckr-1 locus encodes a predicted protein containing 7 transmembrane domains and sharing strong similarity to the CCK-like GPCR family (Fig. S1).
To define potential roles for CKR-1 and CKR-2 in local searching, we took advantage of a strain we had previously generated that stably expresses high levels of the NLP-12 precursor [nlp-12(OE)].6 Overexpression of nlp-12 in this manner elicits exaggerated loopy movement, increased trajectory changes and enhanced body bend amplitude (Fig. 1A, S2A). The average amplitude of bending is increased approximately 3-fold in comparison to wild type (Fig. 1B), and body bends are more broadly distributed over steeper angles (Fig. 1C). These overexpression effects are constitutive, offering experimental advantages for pursuing genetic strategies to identify signaling mechanisms. We investigated the requirement for CKR-1 and CKR-2 in the locomotor changes elicited by nlp-12 overexpression using available strains carrying independent deletions in each of these genes. The ckr-2 deletion (tm3082) has been characterized previously and likely represents a null allele.9,11,29 The ckr-1 deletion (ok2502) removes 1289 base pairs, including exons 3-7 that encode predicted transmembrane domains 2-5 (Fig. S1B-C) and therefore also likely represents a null allele. ckr-1 and ckr-2 single gene deletions each partially reversed the effects of nlp-12 overexpression (Fig. 1B-C, S2A), indicating that both CKR-1 and CKR-2 GPCRs are active under conditions when NLP-12 peptides are present at high levels. Notably, ckr-1 deletion showed slightly greater suppression of nlp-12(OE) phenotypes compared with ckr-2 deletion (Fig. 1B-C). Combined deletion of ckr-1 and ckr-2 completely reversed the locomotor changes produced by NLP-12 overexpression (Fig. 1A-C, S2A), suggesting that the GPCRs may act in a partially redundant manner. Our genetic analysis of nlp-12 overexpression confirms a role for the CKR-2 GPCR in NLP-12-elicited motor adaptations, and importantly, provides first evidence implicating the previously uncharacterized CKR-1 GPCR in NLP-12 modulation of motor activity.

(A) Representative movement trajectories of wild type (black), nlp-12(OE) (red) and nlp-12(OE);ckr-1(lf);ckr-2(lf) (green) animals during forward runs (30s) on NGM agar plates seeded with OP50 bacteria. nlp-12(OE) refers to the transgenic strain (ufIs104) stably expressing high levels of wild type nlp-12 genomic sequence. Note the convoluted nlp-12(OE) movement tracks are restored to wild type by combined ckr-1 and ckr-2 deletion. Scale bar, 1 mm. Asterisks (*) indicate position of worm at start of recording.
(B) Average body bend amplitude (indicated in schematic by blue arrow between orange lines, centroid (green) of worm) for the genotypes as indicated. Bars represent mean ± SEM. In this and subsequent figures, numbers within bars indicate n for each genotype. ****p<0.0001, ***p<0.001, ANOVA with Holms-Sidak post-hoc test.
(C) Frequency distribution of body bending angle (indicated in blue in schematic) for the genotypes indicated. wild type: n= 12, nlp-12(OE): n=10, nlp-12(OE);ckr-1(lf): n=10, nlp-12(OE);ckr-2(lf): n=12, nlp-12(OE);ckr-1(lf);ckr-2(lf): n=12.
(D, E) Concentration-response curves of the mean calcium responses (% activation ± SEM) in CHO cells expressing either CKR-1 (D) or CKR-2 (E) for different concentrations of synthetic peptides NLP-12-1 (solid blue circles) or NLP-12-2 (solid black squares). Solid lines indicate curve fits to the data (n = 6). 95% confidence intervals (nM), CKR-1: NLP-12-1, 1.79-7.07; NLP-12-2, 0.93-3.77 and CKR-2: NLP-12-1, 5.16-12.51; NLP-12-2, 6.43-16.73.
NLP-12 activates CKR-1 with high potency
To obtain direct evidence for NLP-12 activation of CKR-1, we used an in vitro bioluminescence-based approach. CKR-1 was expressed in Chinese hamster ovarian (CHO) cells stably expressing the promiscuous G-protein alpha subunit Gα16 and a bioluminescent calcium indicator, aequorin.31 The NLP-12 precursor gives rise to 2 distinct mature peptides, NLP-12-1 and NLP-12-2. Application of either NLP-12-1 or NLP-12-2 synthetic peptides produced robust calcium responses in cells expressing CKR-1. These responses were concentration-dependent with EC50 values of 3.5 and 1.9 nM for NLP-12-1 and NLP-12-2 peptides, respectively (Fig. 1D). These EC50 values are comparable to those measured for NLP-12 activation of CKR-2 (8.0 nM and 10.2 nM) (Fig. 1E)9, suggesting NLP-12 peptides act with similar potency across CKR-1 and CKR-2 GPCRs. Importantly, none of the other peptides from a library of over 350 synthetic C. elegans peptides elicited activation of CKR-1, nor did the NLP-12 peptides evoke calcium responses in cells transfected with an empty control vector (data not shown), indicating that CKR-1, like CKR-2, is a highly specific receptor for NLP-12.
CKR-1 is a key signaling component for local search behavior
To more deeply investigate roles for CKR-1 and CKR-2 in NLP-12 regulation of movement, we quantified body and head bending during basal locomotion (in the presence of food) using single worm tracking analysis. nlp-12 deletion significantly reduced both body bending and head bending angles in comparison to wild type (Fig. 2A-B). Similarly, single deletions in ckr-1 and ckr-2 each produced significant reductions in body bending, and combined deletion produced effects similar to nlp-12 deletion (Fig. 2A). In contrast, head bending was most strikingly affected by ckr-1 deletion, with ckr-2 deletion producing a more modest reduction (Fig. 2B). The preferential involvement of CKR-1 in head bending suggested the interesting possibility that CKR-1 and CKR-2 GPCRs are differentially utilized to regulate specific aspects of locomotion.

Scatterplots of single worm track analyses of movement (5 minutes) in the presence of food. Body bending (A) and head bending (B) angles (measured angle (blue) is indicated in the schematics) are quantified for the indicated genotypes. Each data point represents the average body or head bend angle for a single animal from analysis of 5 minutes of locomotion. Horizontal red bar indicates mean, shading indicates SEM for wildtype (blue) and mutants (orange). ****p<0.0001 compared to wild type, Wilcoxon rank-sum test. wild type: n=19, nlp-12(ok335): n=16, ckr-1(ok2502): n=16, ckr-2(tm3082): n=16, ckr-1(ok2502);ckr-2(tm3082): n=8.
To explore this possibility further, we investigated whether CKR-1 and CKR-2 GPCRs are differentially involved in local search responses following removal from food. Specifically, we monitored worm movement during a 35-minute period immediately after removal from food and quantified turning behavior during the first (0-5, local searching) and last (30-35, dispersal) five minutes (Fig. 3A-B). Consistent with our previous findings6, we found that deletion of nlp-12 significantly decreased reorientations during the five minutes immediately following removal from food (0-5 minutes). Deletion of ckr-2 produced no appreciable effect on reorientations (Fig. 3C,6); however, single deletion of ckr-1 decreased reorientations to a similar level as observed for nlp-12 deletion (Fig. 3C). Combined deletion of ckr-1 and ckr-2 provided no additional decrease beyond that observed for single ckr-1 deletion (Fig. 3C). In addition, combined deletion of nlp-12 and ckr-1 did not further decrease reorientations compared with either of the single mutants (Fig. 3C). Expression of wild type ckr-1 under the control of native ckr-1 promoter elements (3.5 kb) in ckr-1 mutants restored normal reorientation behavior (Fig. 3C). These findings show that nlp-12 and ckr-1 act in the same genetic pathway and point to a selective requirement for NLP-12 signaling through CKR-1 in regulating trajectory changes during local searching. Deletion of nlp-12 did not produce significant changes in dispersal behavior, but we noted a modest decrease in reorientations during dispersal in ckr-1 mutants (Fig. 3D). This may indicate additional roles for CKR-1 during dispersal. Together, our genetic and behavioral studies implicate CKR-1 and CKR-2 GPCRs as targets of NLP-12 signaling under conditions of overexpression and during basal locomotion. In contrast, we find that NLP-12 modulation of local searching is primarily achieved through CKR-1 activation.
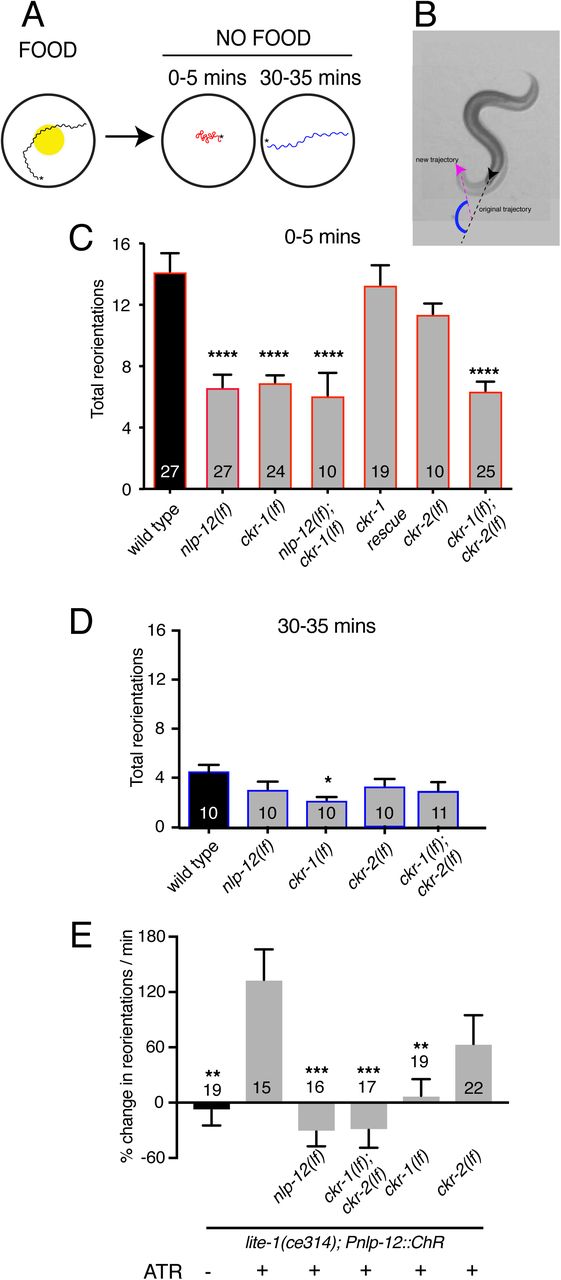
(A) Schematic of the food search assay indicating the time intervals when reorientations were scored. Wild type animals increase reorientations during the first 5 mins (0-5 mins) after removal from food (local search) and reduce reorientations during dispersal (30-35 mins).
(B) Snapshot images overlaid showing worm posture prior to and upon reorientation (5 s between original trajectory (black arrowhead) and new trajectory (pink arrow head). Supplementary angle (blue) between the black and pink dashed lines indicates the change in trajectory. Trajectory changes were scored as reorientations if changes in trajectory were greater than 50°.
(C) Quantification of reorientations during 0-5 minutes following removal from food for the genotypes indicated. Rescue refers to transgenic expression of wild type ckr-1 in ckr-1 mutants. Bars represent mean ± SEM. ****p<0.0001, ANOVA with Holms-Sidak’s post-hoc test. Numbers within bars indicate n for each genotype. See also Figure S2B.
(D) Total number of reorientations during an interval of 30-35 minutes following removal from food for the genotypes as shown. Each bar represents mean ± SEM. *p<0.05, ANOVA with Holms-Sidak’s post-hoc test.
(E) Trajectory changes (reorientations) scored in response to photostimulation of DVA. Percent change in the number of high angle turns elicited during a period of 1 min of blue light exposure compared to prestimulus (no blue light). Bars represent mean ± SEM. ***p<0.001, **p<0.01, *p<0.05 compared to +ATR control. ATR: all-trans retinal.
Acute stimulation of DVA promotes reorientation behavior and requires NLP-12 and CKR-1
We next addressed the question of how neuronal release of NLP-12 promotes area restricted searching. We measured trajectory changes elicited by acute depolarization of the DVA neuron. We used the nlp-12 promoter to drive cell-specific expression of Channelrhodopsin-2 (ChR2)32 in DVA and tracked worm movement during a 1 minute period of blue light (470 nm) photostimulation. We found that animals alter their trajectory of movement (reorientations) more frequently with depolarization of DVA compared to pre-stimulus control (Fig. 3E). Importantly, we did not observe increases in reorientations in the absence of retinal (–ATR) (Fig. 3E), demonstrating that the increase in reorientation frequency was a direct result of DVA depolarization. Depolarization of the DVA neuron in nlp-12 mutants failed to produce a similar enhancement (Fig. 3E), offering support for the idea that reorientations primarily arise due to release of NLP-12 peptides. Single ckr-1 deletion or combined ckr-1 and ckr-2 deletion also abrogated DVA-elicited increases in reorientation behavior while single ckr-2 deletion had no appreciable effect (Fig. 3E), similar to our above findings for area restricted food searching. Our photostimulation experiments provide direct evidence that NLP-12 release from the DVA neuron promotes reorientation behavior, and, in addition, provide evidence for central involvement of NLP-12 signaling through the CKR-1 GPCR in directing trajectory reorientations.
Elevated CKR-1 signaling enhances turning and body bending in an nlp-12 dependent manner
To further define the role of CKR-1, we next asked whether increased CKR-1 signaling would be sufficient to induce local search-like behavior. To address this question, we pursued an overexpression strategy similar to our above approach for nlp-12. We expressed the ckr-1 genomic sequence at high copy levels in wild type animals under native ckr-1 promoter elements. We found that ckr-1 overexpression produced striking increases in turning and large head to tail body bends, qualitatively similar to the effects of nlp-12 overexpression (Fig. 4A, S2A). ckr-1(OE) animals made steep bends during runs of forward movement, with angles approaching 200°, whereas bending angles in wild type rarely exceeded 75° (Fig. 4B). Notably, these high angle bends often produced spontaneous reorientations during forward movement and sometimes elicited sustained coiling. The amplitude of body bends during movement also increased by approximately 3-fold in ckr-1(OE) animals compared to wild type (Fig. 4C). These increases in bending angles and body bend depth were returned to wild type levels by nlp-12 deletion (Fig. 4B-C), demonstrating that NLP-12 peptides are the sole ligands acting on CKR-1 GPCRs to elicit these characteristic changes in movement. Together, our genetic studies define NLP-12/CKR-1 as a novel ligand-GPCR pathway that controls trajectory changes and body bending to produce adaptive behavior.

(A) Representative movement trajectories of wild type (black), ckr-1(OE) (blue) and ckr-1(OE); nlp-12(lf) (green) animals for 30 seconds on NGM agar plates seeded with OP50 bacteria. ckr-1(OE) refers to high copy expression of the wild type ckr-1 genomic locus (ufEx802). Note the increased frequency of high angle turns and convoluted track for ckr-1(OE). These movement phenotypes are reversed by nlp-12 deletion. Scale bar, 1 mm.
(B) Frequency distribution of body bending angles (mean ±SEM) during forward runs (30 s) on plates thinly seeded with OP50 bacteria. wild type: n= 8, ckr-1(OE): n=10, and ckr-1(OE);nlp-12(lf): n=10.
(C) Comparison of the average body bend amplitude for the indicated genotypes. Bars represent mean ± SEM. ****p<0.0001, ANOVA with Holms-Sidak’s post-hoc test. Numbers within bars indicate n for each genotype.
ckr-1 is expressed in many neurons that do not receive direct synaptic inputs from DVA
To identify cells where CKR-1 may act to promote local searching, we examined ckr-1 expression. We generated strains expressing a ckr-1 reporter transgene that included the complete ckr-1 genomic locus and ∼3.5 kb of upstream regulatory sequence SL2 trans-spliced to sequence encoding GFP (green fluorescent protein) or mCherry. We found that ckr-1 is broadly expressed in the nervous system, showing expression in a subset of ventral nerve cord motor neurons, amphid and phasmid sensory neurons, premotor interneurons, and motor neurons in the nerve ring (Fig. 5A-B). We identified many of these neurons, largely from analysis of ckr-1 co-expression with previously characterized reporters (Table S2).

(A) Confocal maximum intensity projection of adult expressing the Pckr-1::ckr-1::SL2::GFP reporter. Note expression in multiple head neurons (white box) and a subset of ventral nerve cord motor neurons (white arrowheads).
(B) Confocal maximum intensity projection of the head region of adult expressing the Pckr-1::ckr-1::SL2::GFP reporter. Scale bar, 10 μm. See Fig. S3 and Table S2 for additional expression information.
(C) Quantification of average body bend amplitudes (mean ± SEM) for ckr-1 overexpression in the indicated cell types. Promoters used for listed cell types: pan-neuronal Prgef-1, muscle Pmyo-3, GABA motor neurons Punc-47, cholinergic ventral cord motor neurons Punc-17β. See Table S3 for details about cellular expression of promoters used for head neurons. ****p<0.0001, ***p<0.001, ANOVA with Holms-Sidak’s post-hoc test. Numbers within bars indicate n for each genotype.
In the ventral nerve cord, we found that ckr-1 is expressed in cholinergic, but not GABAergic, ventral cord motor neurons (Fig. S3A-B, Table S2). Amongst head neurons, the ckr-1 reporter is expressed in GABAergic RMEV, RMED, AVL and RIS neurons, cholinergic SMDV, SMDD and RIV head motor neurons, the interneuron RIG, the serotonergic NSM neuron, and in the interneurons AIA and AIB (Fig. 5B, Table S2). Additional studies using DiI uptake indicated that ckr-1 is also expressed in the amphid sensory neurons ASK and ASI and the phasmid sensory neurons PHA and PHB (Fig. 5, Table S2). Notably, with the exception of the ventral cord cholinergic neurons, the ckr-1 reporter almost exclusively labeled neurons that do not receive direct synaptic input from DVA, suggesting that NLP-12 acts at least partially through extrasynaptic mechanisms.
CKR-1 functions in the SMD head motor neurons to modulate body bending
We next pursued cell-specific ckr-1 overexpression to gain insight into which of the ckr-1-expressing neurons defined above may be primary targets for modulation during local searching (Table S3-S4). We focused our analysis on body bending amplitude because this was the most easily quantifiable aspect of movement to be modified by ckr-1 overexpression. High levels of pan-neuronal ckr-1 expression (rgef-1 promoter) increased body bending amplitude to a similar degree as overexpression using the native promoter (Fig. 5C). In contrast, high levels of ectopic ckr-1 expression in muscles produced no appreciable change, consistent with a primary site of CKR-1 action in neurons (Fig. 5C). Surprisingly, ckr-1 overexpression in cholinergic ventral cord motor neurons (unc-17β promoter) also did not elicit an appreciable change in body bend depth (Fig. 5C). We therefore next targeted the head neurons identified by our ckr-1 reporter, using several different promoters for ckr-1 overexpression in subsets of head neurons (Fig. 5C, Table S3-S4). ckr-1 overexpression using either the odr-2(16) or lgc-55 promoters produced a striking (2.5-fold) increase in body bend depth, comparable with ckr-1 overexpressed under its endogenous promoter. In contrast, ckr-1 overexpression in GABAergic neurons, including RMED and RMEV (using unc-47 promoter), did not produce an appreciable effect. Likewise, ckr-1 overexpression in RIV, RIG, NSM, AIA, AIB or amphid neurons failed to significantly enhance body bend depth. The lgc-55 promoter drives expression in AVB, RMD, SMD and IL1 neurons, as well as neck muscles and a few other head neurons33, while the odr-2(16) promoter primarily labels the RME and SMD head neurons34 (Table S2-S3). The overlapping expression of the odr-2(16) and lgc-55 promoters in SMD neurons suggests that these neurons may be centrally involved. SMD co-labeling by ckr-1::SL2::mCherry and Plad-2::GFP35 provided additional evidence for significant levels of ckr-1 expression in these neurons (Fig. S3C). Intriguingly, we also noted that NLP-12::Venus clusters are concentrated in the nerve ring region of the DVA process (Fig. S5), in the vicinity of SMD processes. In contrast to ckr-1, ckr-2 expression in the SMDs was absent or barely detectable (Fig. S3D)36, pointing towards a specific role for ckr-1 in regulating SMD motor neurons.
The 4 SMDs (dorsal-projecting SMDDL and SMDDR and ventral-projecting SMDVL and SMDVR) are bilateral motor neuron pairs that innervate dorsal and ventral head/neck musculature28, and also form reciprocal connections with one another. They have been previously implicated in directional head bending and steering.37–40 To better define the behavioral effects of SMD modulation, we more closely examined body bending in animals overexpressing ckr-1 under control of the odr-2(16) promoter, and also using a second promoter, flp-22Δ4, that was recently shown to drive selective expression in the SMD neurons41 (Fig. 6A). For both overexpression strains, we observed significant increases in body bending amplitude and bending angle compared to wild type (Fig. 5C, 6B, S2A). These increases were dependent on NLP-12 signaling (Fig. S6) and were similar to those observed for native ckr-1 (Fig. 4, S2A) and nlp-12 overexpression (Fig. 1, S2A). Thus, the actions of CKR-1 in the SMD motor neurons recapitulate many of the behavioral effects of NLP-12 overexpression.
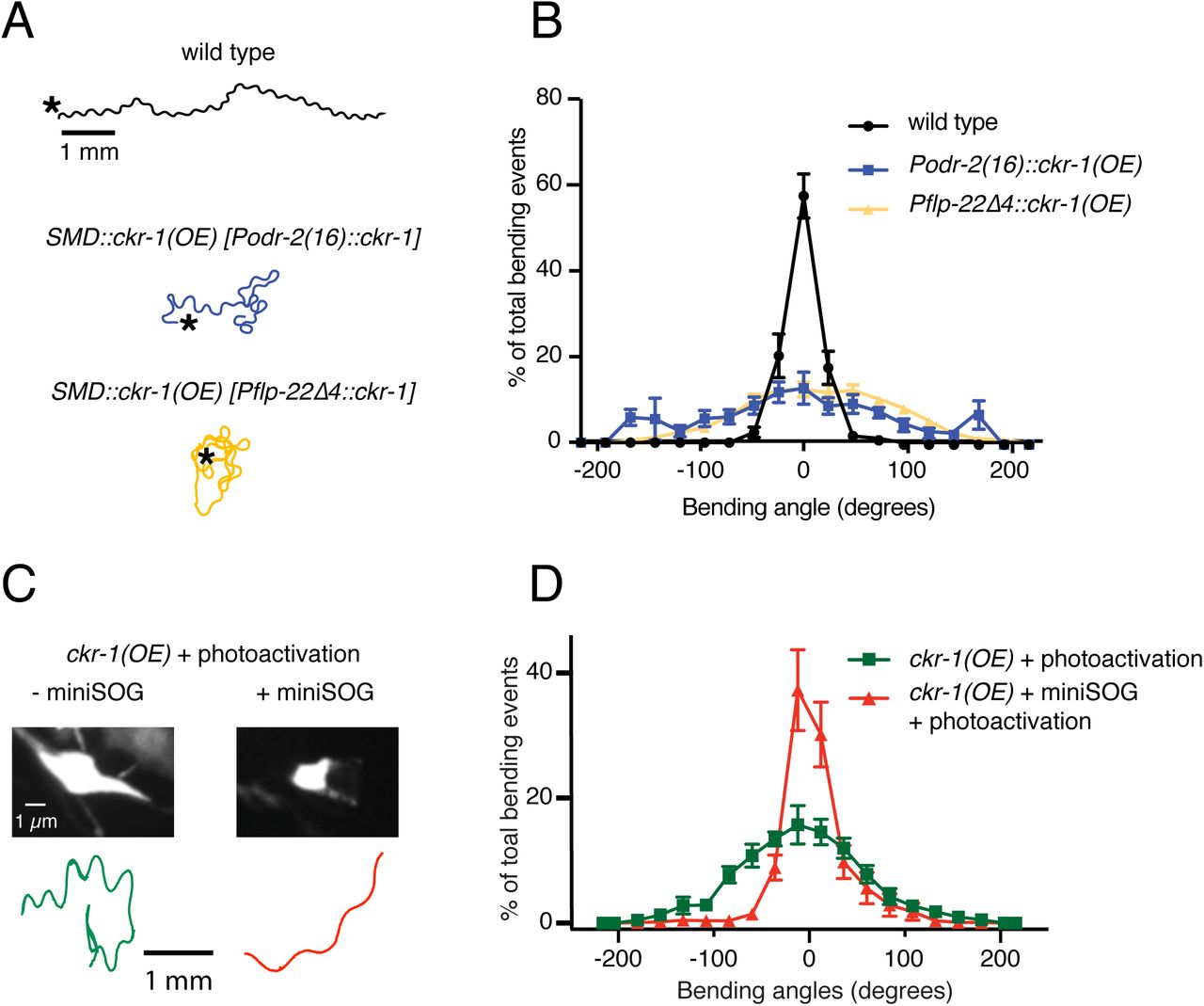
(A) Representative tracks (1 minute) for indicated genotypes. Asterisks indicate position of animal at the beginning of recordings. Note increased reorientations and body bending depth in the tracks with cell-specific ckr-1 overexpression. Scale bar, 1mm.
(B) Average body bending angle distribution (mean ± SEM) for the indicated genotypes. High level expression of ckr-1 in SMDs using the odr-2(16) or flp-22Δ4 promoters increases bending angle. wild type n=9 (black circles), Podr-2(16)::ckr-1(OE): n=8 (blue squares), Pflp-22Δ4::ckr-1(OE): n=11 (orange triangles)
(C) Top, representative fluorescent images of SMD motor neuron in ckr-1(OE) animals without (left) or with (right) miniSOG expression 16 hours following photoactivation. Bottom, representative 30 s track for control ckr-1(OE) (-miniSOG, left) animal or SMD ablated ckr-1(OE) (+miniSOG, right) animal 16 hours after photostimulation. Scale bar, 1 µm.
(D) Average body bending angle distribution (mean ± SEM) for control ckr-1(OE) (green squares, n=8) and SMD ablated ckr-1(OE) (red triangles, n=11) animals. SMD ablation reduces the frequency of large bending angles produced by ckr-1(OE).
To ask if the SMD neurons are required for the locomotor changes produced by ckr-1 overexpression, we expressed the photoactivatable cell ablation agent PH-miniSOG in the SMD neurons (Pflp-22Δ4) of animals overexpressing ckr-1 (native promoter). When activated by blue light (470 nm) PH-miniSOG produces reactive oxygen species and disrupts cellular function.42 Following photoactivation of miniSOG in animals overexpressing ckr-1, we observed striking decreases in bending angles during movement (Fig. 6C-D). We confirmed successful SMD ablation by examining morphological changes in GFP-labeled SMD neurons following photoactivation of miniSOG (Fig. 6C). Expression of miniSOG did not have appreciable effects on the movement of ckr-1(OE) animals under control conditions (without light exposure) (Fig. S7A). In addition, stimulation of control animals without the miniSOG transgene did not appreciably alter body bending (Fig. 6D) or SMD neuron morphology (Fig. S7B). These results indicate that SMD motor neurons are required for the locomotor effects of ckr-1 overexpression, and, importantly, raise the possibility that the SMD neurons are key targets for NLP-12 neuromodulation during local searching in wild type.
NLP-12/CKR-1 excitation of the SMD neurons promotes local searching
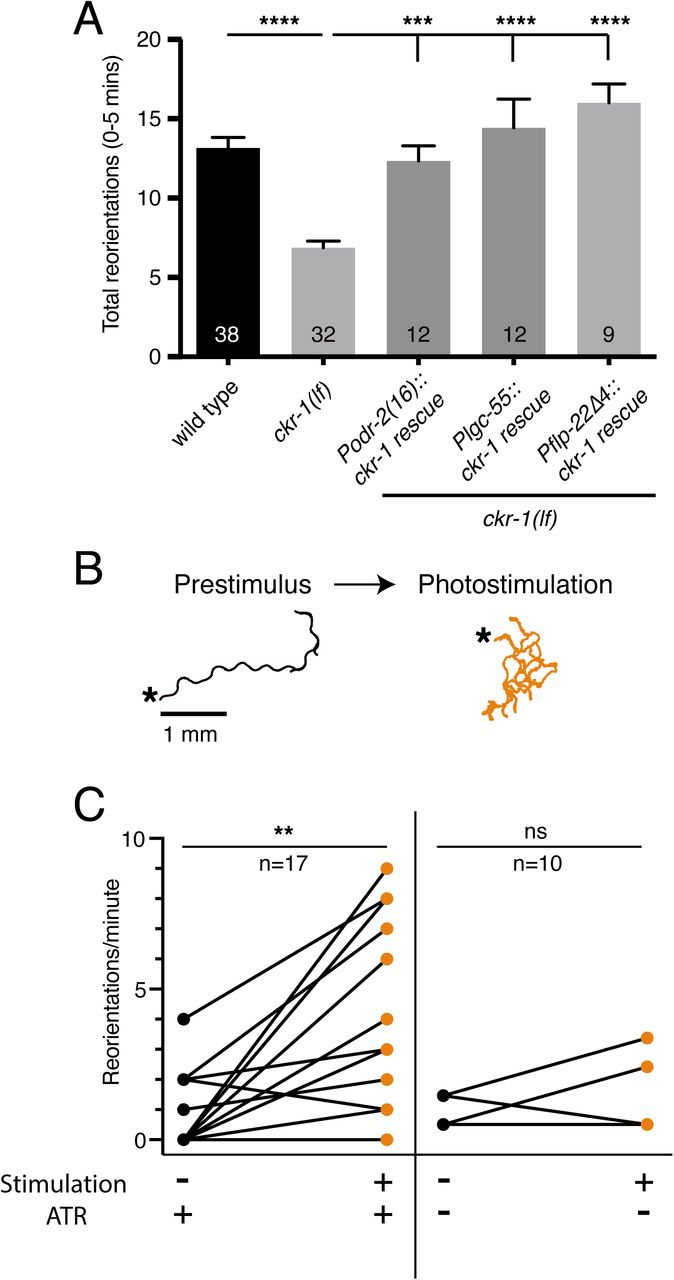
To test this hypothesis, we examined rescue of area restricted searching in ckr-1 mutants using low-level (5-fold lower than used for overexpression) SMD-specific expression of wild type ckr-1. When the ckr-1 transgene was expressed at this concentration in wild type animals, we did not observe appreciable increases in bending depth or angle (Fig. S8). However, low-level SMD-specific ckr-1 expression in ckr-1 mutants restored reorientations during food searching to roughly wild type levels (Fig. 7A). This result indicates that CKR-1 function in the SMD neurons is sufficient to support NLP-12 modulation of local searching.

(A) Total reorientations measured during 0-5 minutes following removal from food for the genotypes indicated. ckr-1 rescue refers to expression of wild type ckr-1 (5 ng/µL) in ckr-1(ok2502) animals using the indicated promoters. Bars represent mean ± SEM. ****p<0.0001, ***p<0.001 ANOVA with Holms-Sidak’s post-hoc test. Numbers within bars indicate n for each genotype.
(B) Representative tracks (1 minute) on thinly seeded NGM agar plates prior to (left) and during photostimulation (right) for transgenic animals expressing Podr-2(16)::Chrimson. Scale bar, 1 mm. Asterisks (*) indicate position of worm at start of recording.
(C) Left, quantification of reorientations for individual animals over 1 minute durations prior to (prestimulus) and during photostimulation (+ATR). Right, quantification of reorientations for individual animals prior to and during photostimulation in control animals (-ATR). Black circles, reorientations during prestimulus. Orange circles, reorientations during photostimulation. **p<0.01, ns not significant. Paired t-test. ATR: all trans retinal.
To investigate how increased SMD activity may impact movement, we photostimulated the SMDs in animals expressing Podr-2(16)::Chrimson.43 Prior to photostimulation, animals demonstrated long forward runs with relatively few changes in trajectory. Following the onset of photostimulation, Chrimson-expressing animals rapidly increased turning and trajectory reorientations (Fig. 7B-C). In contrast, control animals (-Retinal) did not increase trajectory changes during the light stimulation period (Fig. 7C). These results indicate that depolarization of the SMD neurons is sufficient to promote turning and reorientations, similar to our findings for NLP-12/CKR-1 signaling onto the SMD neurons. Taken together, our findings support a model (Fig. 8) where NLP-12/CKR-1 signaling promotes local searching by increasing the activity of SMD head motor neurons.

During basal locomotion, NLP-12 activation of CKR-1 and CKR-2 GPCRs in ventral nerve cord motor neurons regulates body bending. During local searching, NLP-12 acts primarily through CKR-1 in SMD motor neurons to promote increased turning, trajectory changes and enhance body bending. Solid arrows indicate known synaptic connections, dotted arrows indicate extrasynaptic. Sensory neurons (green), head interneurons (orange), and motor neurons (red). Olfactory sensory neurons: AWA, AWB, AWC, ASE.
Discussion
Neuropeptidergic systems have crucial roles in modulating neuronal function to shape alternate behavioral responses, but we have limited knowledge of the circuit-level mechanisms by which these alternate responses are generated. Here, we show that the C. elegans NLP-12 neuropeptide system, closely related to the CCK system in mammals, shapes adaptive behavior through conditional modulation of motor circuits dedicated to control of either head or body wall musculature. We demonstrate that NLP-12 modulation of these circuits occurs through distinct GPCRs, CKR-1 and CKR-2, that primarily act on either head or body wall motor neurons respectively. Under basal conditions, NLP-12 modulation of the body wall motor circuit predominates, influencing the depth of body bends during sinusoidal movement through CKR-1 and CKR-2 GPCRs located on body wall motor neurons. NLP-12 activation of head motor neurons through CKR-1 becomes predominant in the absence of food, promoting reorientations. We propose that changes in food availability reconfigure functional connectivity in the NLP-12 system by differentially engaging GPCRs across the head and body wall motor circuits. Intriguingly, the involvement of 2 GPCRs in nematode NLP-12 signaling is reminiscent of the organization of the CCK system in mammals, which relies on signaling through CCK1 and CCK2 GPCRs. CCK is among the most abundant peptides in the mammalian brain. New details about functional roles for CCK signaling in the brain and the GPCRs involved are continuing to emerge.12–15,44–47 Our findings in worms may point towards similar conditional utilization of specific CCK-responsive GPCRs to coordinate activity across circuits in mammals.
NLP-12 neuropeptides act as key modulators in a range of C. elegans behaviors. Local search responses to varying oxygen levels and decreased food availability both involve NLP-12 signaling.6,7 Additionally, NLP-12 signaling has been implicated in various aspects of proprioceptive signaling and postural control.29,30 However, the mechanisms by which NLP-12 peptides exert their influence over these diverse behavioral responses have remained unclear. Our work addresses these mechanistic questions by defining specific roles for CKR-1 and CKR-2 GPCRs during basal locomotion and area-restricted searching. Area-restricted searching is a complex motor behavior, involving rapid trajectory changes that serve to maintain the animal within a restricted area of their immediate environment.6,7,17,20 Reorientations during searching are produced through high angle forward turns6,48 and reversal-coupled omega turns6,17. We previously demonstrated a requirement for NLP-12 in promoting reorientations during local searching.6 Our analysis here shows that loss of nlp-12 also has modest effects on body posture during normal exploratory movement, indicating additional NLP-12 regulation of motor targets under basal conditions. Intriguingly, the behavioral requirement for NLP-12 is far more apparent during local searching compared with basal locomotion, suggesting enhanced involvement of NLP-12 signaling for performance of local searching. Similar observations about NLP-12 involvement in chemotactic responses to varying oxygen levels suggested a model for graded NLP-12 regulation of movement.7 Based on our observations, we speculate that increased engagement of head motor neurons through CKR-1 activation may be a generalizable mechanism for dynamic NLP-12 regulation of behavior over changing external conditions.
Prior studies had implicated the CKR-2 GPCR in NLP-12 function9,29,30, but roles for CKR-1 had not been previously described. CKR-2 shows slightly broader expression compared with CKR-1, but both GPCRs are expressed across a variety of neuron classes, including many that do not receive direct synaptic inputs from DVA. We noted very little overlap in CKR-1 and CKR-2 expression (Fig. S4), consistent with the idea that the two GPCRs serve distinct roles in modulating behavior. Our genetic analyses and heterologous expression studies firmly establish CKR-1 as a functional target for NLP-12 signaling with an activation profile similar to CKR-2. NLP-12 activation of CKR-2 stimulates neurotransmission through coupling with egl-30 (Gαq) and egl-8 (PLCβ) likely by DAG interaction with the synaptic vesicle priming factor UNC-13.29,30 Given the sequence homology between CKR-1 and CKR-2, it seems likely that CKR-1 also functions to positively regulate neuronal activity through egl-30. In support of this idea, we found that SMD-specific CKR-1 overexpression and SMD neuron photostimulation produced qualitatively similar behavioral effects, an increase in reorientations during movement. The DVA neuron makes a single synapse with SMDVL (Worm wiring). It is unclear whether this single synapse could account for NLP-12 elicited behavioral changes during local searching, or whether additional extrasynaptic signaling to other SMD neurons may also contribute.
SMDs innervate head and neck muscles28,49 and biased activity of dorsal or ventral SMDs is correlated with directional head bending37,38,40,41,50. Turning is preceded by head bending or head swings. Shallow bends promote more gradual turns, while large head swings drive sharp turns.48,51 Our results suggest NLP-12 activation of CKR-1 may modulate functional connectivity between SMD neurons and dorsal and ventral head/neck muscles. According to this model, enhanced transmission from dorsal or ventral SMD synaptic outputs onto head and neck muscles would promote exaggerated head bending, thereby promoting enhanced turning and trajectory changes. CKR-1 activation could also influence SMD proprioceptive functions41 or functional connectivity with other neurons implicated in regulation of head posture and bending, such as RIA37,50 and RME38.
Surprisingly, selective ckr-1 overexpression using the odr-2(16) or flp-22Δ4 promoters increased body bend depth, raising the question of how altered SMD activity might translate into increased body bending. Recent work suggests an interesting functional coupling between the activity of SMD neurons and ventral cord B-type motor neurons.40 B-type motor neurons are suggested to act as a distributed central pattern generator for the propagation of body bends.52,53 CKR-1 activation of SMDs may therefore influence body depth directly by altering body wall motor neuron excitability through a gap junction connection between VB1 and SMDVR or through neuromuscular synapses located in the sublateral processes.
The similar potency of NLP-12 peptides for activating CKR-1 and CKR-2, suggests that differential contributions of these GPCRs during basal locomotion and search responses do not arise differences in their responsiveness to NLP-12 peptides. This raises important questions about how a bias toward CKR-1 modulation of the head motor circuit during local searching may occur. We envision that NLP-12 regulation of the SMD neurons acts in parallel with other neural pathways previously shown to promote reversals during local searching. For example, olfactory information about food availability is conveyed by sensory neurons such as AWC and ASK to premotor interneurons (AIA, AIB, AIY) and ultimately transformed into patterns of motor neuron activity that drive reversals.17,18,50,54 The SMD neurons also receive synaptic information from this circuit (for example, through synaptic connections from the AIB and RIM neurons)28, raising the possibility that a pathway activated by food removal may enhance SMD sensitivity to CKR-1 activation. In this case, SMD neurons may be a site for integration of information encoding reversals and forward reorientations during local searching. A shift to CKR-1 modulation of head neurons during searching could also be triggered by dopaminergic stimulation of DVA. Prior work implicated dopaminergic signaling from PDE neurons in the regulation of NLP-12 and motor responses.6,8 In this case, elevated levels of NLP-12 secretion, perhaps from release sites in the nerve ring region, would be predicted to bias the system towards enhanced activation of the SMD neurons and elicit increased turning. Notably, PDE also regulates an antagonistic peptidergic circuit, mediated by FLP-1 neuropeptides, through inhibitory connections with AVK interneurons8, suggesting the potential for more complex behavioral regulation.
Our studies of the nematode NLP-12 system offer new mechanistic insights into neuropeptide modulation of behavior. Our findings provide a key first step in defining roles for two NLP-12-responsive GPCRs in coordinating motor control across changing conditions. We propose that the NLP-12 system conditionally engages GPCRs expressed in head or body motor neurons to modify specific features of locomotion, most notably reorientations during searching and body bend depth during basal locomotion. Brain CCK has been increasingly implicated as a key regulator in diverse aspects of behavior, including feeding, satiety, memory, nociception and anxiety.14,44,55–58 Thus our studies elucidating mechanisms for NLP-12 regulation of circuit function in the compact nematode nervous system may have important and broadly applicable implications for neuromodulation in more complex systems.
Disclosure Statement
The authors have nothing to declare.
Materials and Methods
Strains
All nematode strains (Table S1) were maintained on OP50 seeded agar nematode growth media (NGM) at room temperature (22–24°C). N2 Bristol strain was used as wild type. Transgenic animals were generated by microinjection into the germ line and transformation monitored by co-injection markers. Multiple independent extrachromosomal lines were obtained for each transgenic strain and data presented from a single representative transgenic line. Stably integrated lines were generated by X-ray integration and outcrossed at least four times to wild type.
Molecular Biology
All plasmids, unless specified, were generated by Gateway cloning (see Supplementary Tables). p-ENTR plasmids were generated for all promoters used (Table S5). The ckr-1 minigene construct (pRB12/pRB13) was generated by cloning the ckr-1 coding sequence (start to stop), with introns 1, 8 and 9. For cell specific overexpression or rescue, the ckr-1 minigene was recombined with entry vectors containing the relevant cell-specific promoters. (Table S3-S4).
Behavioral assays and analyses
All behavioral assays were carried out using staged 1-day adult animals on Bacto-agar NGM agar plates seeded with a thin lawn of OP50 bacteria (50 µL) unless otherwise noted. Video recordings for behavioral analyses were obtained using a Firewire camera (Imaging Source) and ICCapture2.2. Animals were allowed to acclimate for 30 seconds prior to video recording. Locomotor analysis was performed using Wormlab (MBF Bioscience). Body bending angle was measured at the centroid of the animal’s body as the supplementary angle measured between the head, midpoint and tail (Fig. 1C), and represented as binned histogram distributions. Body bend amplitude was quantified as the average centroid displacement over the duration of a locomotion track (Fig. 1B). Kymographs were generated from worm body curvature data using custom MATLAB script (Jeremy Florman and Mark Alkema).
Area restricted search behavior
For quantification of local search behavior, single well-fed animals were transferred to an intermediate unseeded plate. After one minute, animals were repicked without bacteria and transferred to an unseeded behavior assay plate. Digital movies of animals (5 minutes duration) were captured over the first 5 mins (local search) and after 30 mins (dispersal). Videos were analyzed in ImageJ using the angle tool to measure the supplementary angle between the original and new trajectory (Fig. 3B) and reorientations were scored when changes in trajectory were >50°.
Single worm tracking
Single worm tracking was carried out using Worm Tracker 2.59 Animals were allowed to acclimate for 30 seconds prior to tracking. Movement features were extracted from five minutes of continuous locomotion tracking. Worm tracker software version 2.0.3.1, created by Eviatar Yemini and Tadas Jucikas (Schafer lab, MRC, Cambridge, UK), was used to analyze movement.60 Absolute midbody bending (Fig. 2A) and head bending (Fig. 2B) angles were quantified.
SMD ablation
Cell ablation protocol by miniSOG activation was adapted from Xu et. al. 2016.42 MiniSOG activation was achieved by stimulation with repetitive 2 Hz 250 ms blue light pulses for 12 minutes [200mW/cm2, 488 nm 50 W LED (Mightex Systems)]. Experiments were performed on unseeded plates using larval stage 4 ckr-1(OE) animals expressing miniSOG and GFP transgenes under the flp-22Δ4 promoter. Following stimulation, animals were allowed to recover in the dark on NGM OP50 plates for 16 hours prior to behavioral analysis or imaging.
Photostimulation experiments
All-Trans Retinal (ATR) plates were prepared (100 mM stock in Ethanol, final working 2.7 mM in OP50). Plates were stored at 4°C under dark conditions and used within one week. Animals were grown on +ATR OP50 plates in dark and L4 animals were transferred to a fresh +ATR plate prior to the day of experiment. Experiments were performed using one-day adults. For ChR2 photostimulation, experiments were conducted using a fluorescent dissecting microscope (Zeiss stereo Discovery.V12) equipped with a GFP filter set. Behavior was recorded for a 1-minute period prior to photostimulation and during a subsequent 1-minute period during photostimulation. Data are expressed as % change in reorientations across these time intervals. Chrimson photostimulation (26 mW/cm2) experiments were conducted using a 625 nm 50 W LED (Mightex Systems). Animals were video recorded for 1 minute in the absence of light stimulation (prestimulus) and subsequently for 1 minute with light stimulation. Control experiments (-ATR) were performed in the same manner.
Imaging
Fluorescent images were acquired using either BX51WI (Olympus) or Yokogawa (Perkin Elmer) spinning disc confocal microscopes. Data acquisition was performed using Volocity software. Staged one-day adult animals were immobilized using 0.3 M sodium azide on 2% agarose pads. Images were analyzed using ImageJ software.
in vitro GPCR characterization
The GPCR activation assay was performed as previously described.31,61,62 Briefly, CHO-K1 cells stably expressing the luminescent Ca2+ indicator aequorin and the promiscuous Gα16 protein (ES-000-A24 cell line, PerkinElmer) were transiently transfected with ckr-1/pcDNA3.1, ckr-2/pcDNA3.1 or empty pcDNA3.1 vector. Cells were transfected with Lipofectamine LTX and Plus reagent (Invitrogen) at 60–80% confluency and grown overnight at 37°C. After 24 hours, they were shifted to 28°C overnight. On the day of the assay, transfected cells were collected in bovine serum albumin (BSA) medium (DMEM/F12 without phenol red with L-glutamine and 15 mM HEPES, Gibco, supplemented with 0.1% BSA), at a density of 5 million cells per mL, and loaded with 5 µM coelenterazine h (Invitrogen) for 4 hours at room temperature. Compound plates containing synthetic peptides in DMEM/BSA were placed in a MicroBeta LumiJet luminometer (PerkinElmer). After loading, the transfected cells were added at a density of 25,000 cells/well, and luminescence was measured for 30 seconds at a wavelength of 469 nm. After 30 seconds, 0.1% triton X-100 (Merck) was added to lyse the cells, resulting in a maximal Ca2+ response that was measured for 30 seconds. To constitute concentration-response curves of NLP-12 peptides, peptide concentrations ranging from 1 pM to 10 µM were tested in triplicate on two independent days.
Supplementary Figures

(A) Dendrogram (generated using Phylogeny,fr75) showing the predicted relationship between Drosophila (Dm_CCKLR-1/2), C. elegans (Ce_CKR-1/2), mouse (Mm) and human (Hs) CCK1/2-R GPCRs.
(B) Boxshade alignment of C. elegans CKR-1 and CKR-2 with Human CCK-1 and CCK-2 receptors. Black shading indicates identical amino acids, while grey shading indicates similar amino acids. Red bar indicates the amino acids removed by ckr-1(ok2502) deletion.
(C) Schematic representation of CKR-1 GPCR membrane topology and domains affected by the ckr-1(ok2502) deletion (red shading). NLP-12 ligand indicated by solid orange circle.

Representative body curvature kymographs for worm locomotion, basal locomotion on food (A) and area restricted searching (ARS) off food (B), during first minute of locomotion on a thin seeded OP50 assay plate. Head to tail orientation along the horizontal axis in each kymograph is left to right as indicated for wild type. Time is indicated along the vertical axis from 0 to 1 minute. SMD ckr-1(OE) indicated is Pflp-22(Δ4)::ckr-1.

(A) Confocal maximum intensity projections of a segment of the ventral nerve cord of a transgenic animal coexpressing Pckr-1::ckr-1::SL2::GFP and the cholinergic reporter Pacr-2::mCherry. ckr-1 is expressed in the DA and DB motor neurons in the ventral nerve cord. Anterior is to the left in all panels. Scale bar, 10 μm.
(B) Confocal maximum intensity projections of a segment of the ventral nerve cord of a transgenic animal coexpressing Pckr-1::ckr-1::SL2::mCherry and the GABAergic reporter Punc-47::GFP.
(C) Confocal maximum intensity projections of optical sections with SMD fluorescence (GFP) from the head region of a transgenic animal expressing ckr-1::SL2::mCherry (left panel) together with Plad-2::GFP (middle panel). White arrowheads denote the SMD cell bodies in all cases. Note the colocalization of the red and green fluorescence exclusively in the SMD neurons (merge right panel).
(D) Confocal maximum intensity projections of optical sections with SMD fluorescence (mCherry) from the head region of a transgenic animal coexpressing Podr-2(16)::mCherry (left panel), and Pckr-2::GFP (middle panel). Note weak ckr-2 expression in a single SMDD neuron (merge, right panel).

Confocal maximum intensity projections of transgenic worm expressing Pckr-1::ckr-1::SL2::mCherry and Pckr-2::GFP. (A) ckr-1 and ckr-2 expression in the entire worm. Both ckr-1 and ckr-2 are highly expressed in head neurons and ventral nerve cord motor neurons. However, there is very little overlap between expression of ckr-1 and ckr-2. (B) Magnified view of ckr-1 and ckr-2 expression in the head region. (C) Magnified view of ckr-1 and ckr-2 expression in the ventral nerve cord. Scale bar, 10 µm.

Confocal maximum intensity projection of the nerve ring region of a transgenic animal expressing Pnlp-12::NLP-12::Venus. Note the high levels of NLP-12::Venus in the nerve ring. Scale bar, 5 µm.

(A) Representative tracks (30 s) for transgenic animals with high levels of cell-specific ckr-1 expression (Pflp-22Δ4::ckr-1) in wild type (top) or nlp-12 deletion background (bottom). Asterisks indicate position of animals at the beginning of recording. Scale bar, 1 mm.
(B) Average bending angle distribution (mean ± SEM) for SMD-specific ckr-1(OE) in wild type (green squares, n=11) or nlp-12(lf) background (orange circles, n=10).

(A) Average body bending angle distribution (mean ± SEM) for pSMD::ckr-1(OE) animals expressing miniSOG in SMDs (Pflp-22Δ4::miniSOG), but not subjected to photoactivation (control, black diamonds) compared to wild type (blue triangles). n=8 for each group.
(B) Single confocal slices of GFP-labeled SMD neurons, following photoactivation (right) compared to control (-photoactivation, left), in transgenic animals without miniSOG expression. Photoactivation protocol does not alter SMD neuron morphology in the absence of miniSOG expression. Scale bar, 1 µm.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
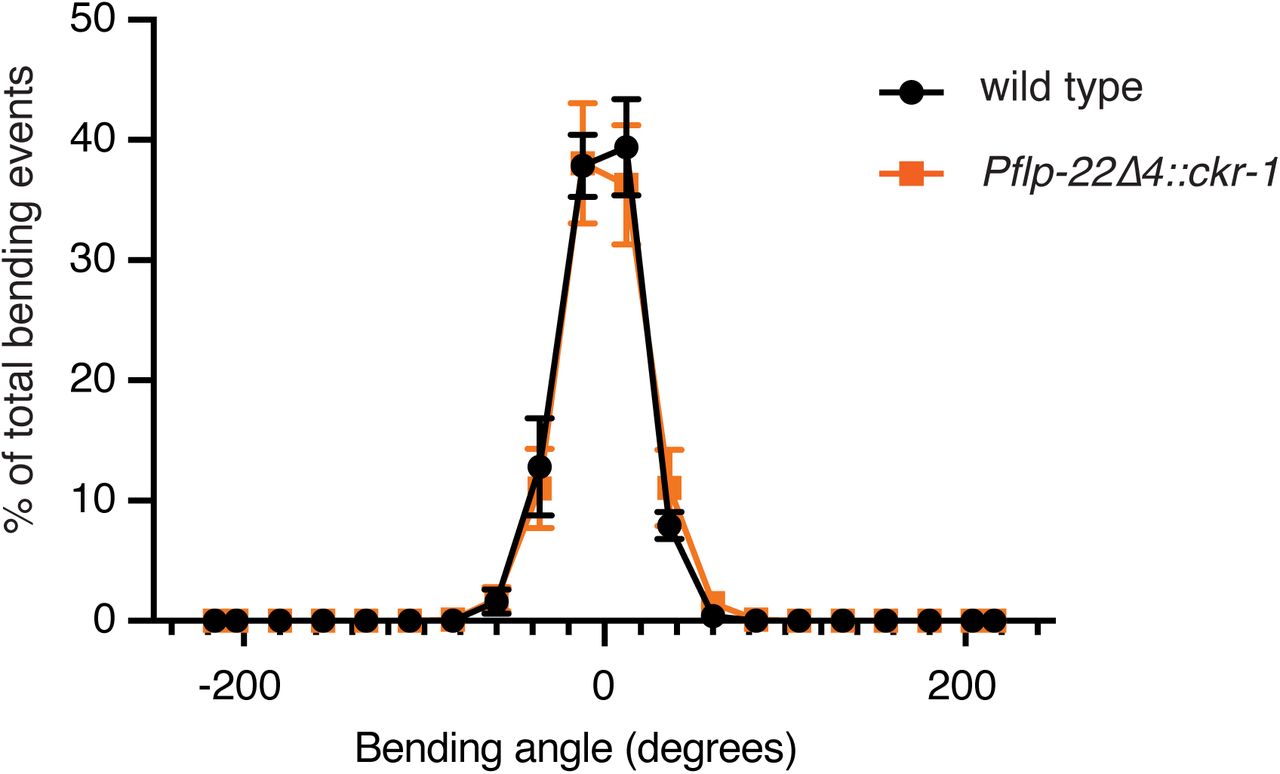
Average body bending angle distribution (mean ± SEM) plotted for wildtype control animals (solid black circles, n=8) and Pflp-22Δ4::ckr-1 (solid orange squares, n=8). Low level (5 ng/µL) cell-specific expression of ckr-1 in SMDs in wild type did not alter body bending.
Supplementary Tables
Strains generated/used in this work.
Cell identification (method of ID, marker and strain indicated for each neuron) to determine ckr-1 expressing neurons. * Indicated strains were crossed into ufIs141 (Pckr-1::ckr-1::SL2::GFP) to generate strains to determine colocalization. # + or – indicates presence or absence of ckr-1 expression in identified neuron.
Promoters used in ckr-1(OE) screen (Fig.5C) indicating expression pattern. Bold neurons indicate ckr-1 expression.
Promoter lengths and primer information for promoters used.
Acknowledgements
We thank the Caenorhabditis Genetics Center, which is funded by the National Institutes of Health National Centre for Research Resources, and the Mitani laboratory (National Bioresource Project) for providing Caenorhabditis elegans strains. We thank Mark Alkema and Jeremy Florman for strains, and help with single worm tracking and analysis, Claire Bénard for strains, Michael Gorczyca and William Joyce for technical support. We thank Francis lab members Devyn Oliver and Kellie Alexander for helpful comments on the manuscript. This work was supported by NIH R21NS093492 (MMF), European Research Council 340318 and Research Foundation Flanders grant G0C0618N (IB).
References




