

SUMMARY
Advanced transcriptome sequencing has revealed that the majority of eukaryotic genes undergo alternative splicing (AS). Nonetheless, limited effort has been dedicated to investigating the functional relevance of particular splicing events, even those in the key developmental and hormonal regulators. Here we reveal, in the plant model Arabidopsis thaliana, that the PIN7 gene, which encodes a polarly localized transporter for the phytohormone auxin, produces two evolutionarily conserved transcripts. These isoforms PIN7a and PIN7b, differing in a 4 amino acid motif, are present at nearly equal levels in most cells, except some early developing tissues where the expression of PIN7b is moderately prevalent. Both proteins also transport auxin with similar capacity and directionality. However, only PIN7a but not PIN7b cDNA rescues the phenotypes associated with the pin7 knock-out mutation, consistent with their differences in the subcellular trafficking and dynamics at the plasma membrane. Further phenotypic analyses suggested a joint, mutually opposing activity of both isoforms as being required for correct seedling apical hook formation and auxin-mediated tropic responses. These results establish alternative splicing of the PIN family as an evolutionary conserved, functionally relevant mechanism, taking part in the auxin-mediated plant development.
INTRODUCTION
Auxin is an essential phytohormone, which plays a role in nearly all aspects of plant development. To flexibly adapt to rapidly changing environmental cues, directional auxin transport represents a highly dynamic means for triggering downstream morphogenetic processes. PIN FORMED (PIN) auxin efflux carriers are among the key regulators in this respect. Many efforts in the past years uncovered several mechanisms operating transcriptionally or post-translationally on the capacity and directionality of PIN-mediated transport. However, little progress has been made in exploring the contribution of post-transcriptional regulation (Adamowski and Friml, 2015; Hrtyan et al., 2015).
Advances in high throughput sequencing have revealed unexpected complexity within eukaryotic transcriptomes by alternative splicing (AS). Although the majority of AS transcripts may be functionally neutral (Darracq and Adams, 2013; Reddy et al., 2013; Tress et al., 2017; Blencowe, 2017; Mei et al., 2017), several detailed studies have highlighted a plausible role for numerous AS events in physiologically relevant contexts, including those involved in plant developmental and hormonal pathways (Staiger and Brown, 2013; Hrtyan et al., 2015; Shang et al., 2017; Szakonyi and Duque, 2018). Earlier works have described auxin-related defects resulting from the aberrant function of several regulators of AS (Kalyna et al., 2003; Casson et al., 2009; Retzer et al., 2014; Tsugeki et al., 2015; Hrtyan et al., 2015; Bazin et al., 2018). AS changes subcellular localization of the auxin biosynthetic gene YUCCA 4 (Kriechbaumer et al., 2012) and differential splicing of an exitron (Marquez et al., 2015) inside the AUXIN RESPONSE FACTOR 8 results in developmental changes of generative organs (Ghelli et al., 2018). AS of the Major Facilitator Superfamily transporter ZIFL1 interferes with auxin transport, influencing the stability of PINs on the plasma membrane (PM) (Remy et al., 2013). These lines of evidence suggest that AS is an important player in auxin-dependent processes. However, no coherent functional model of any auxin-related AS event has been provided so far.
Here, we characterize AS of the PIN7 gene in Arabidopsis thaliana. PIN7 is, together with PIN3 and PIN4, a member of the PIN3 clade of PIN auxin efflux carriers (Bennett et al., 2014), which are required for a broad range of morphogenetic and tropic processes (Adamowski and Friml, 2015). We reveal that AS influences the dynamics of the PIN7 protein on the PM. We also demonstrate that the coordinated action of both splice variants is required for fine-tuning auxin-mediated tropic responses and during apical hook development.
RESULTS
Arabidopsis PIN7 and PIN4 produce two evolutionarily conserved AS transcripts
Our previous survey (Hrtyan et al., 2015) revealed that several genes involved in auxin-dependent processes undergo AS. Among them, closely related paralogs from the PIN3 clade of auxin transporters, PIN4 and PIN7 (but not PIN3), are regulated by the same type of AS (Petrasek et al., 2006; Bennett et al., 2014; Hrtyan et al., 2015). The resulting transcripts, denoted as a and b, differ in the position of the AS donor site in the first intron (Figure 1A). The differentially spliced region corresponds to a four amino acid motif inside the large internal hydrophilic loop (Ganguly et al., 2014; Nodzyński et al., 2016) of the integral PM transporter (Figure 1A and 1B). We examined the quantities of individual reads spanning the exon junctions in the respective region from the Arabidopsis root tip and in several other available transcriptomes from different tissues and organs (Klepikova et al., 2016; Cheng et al., 2017; Ruzicka et al., 2017) (Figures 1C and Supplemental Figure 1A). We found that both PIN4- and PIN7a and b transcripts are expressed abundantly in all tissues, independently on the data set inspected. Besides these AS events, we also identified a minor PIN4c (Marquez et al., 2012; Hrtyan et al., 2015) splice isoform (but not corresponding PIN7c), which comprised around 3-7% of the PIN4 exon1-exon2 spanning reads (Figures 1A and Supplemental Figure 1A). Other occasionally observed transcripts (also corresponding to the other exon junctions) were not repetitively seen among different RNA-seq data sets. It thereby appears that PIN7 and PIN4 are processed into two and three transcripts, respectively, and that PIN7a and b (or PIN4a and b) are expressed in most of the plant organs at comparable or nearly similar levels.

(A) Scheme of coding regions of the PIN3 clade genes in Arabidopsis thaliana. The alternative donor splice site at the end of the first exon of PIN7 and PIN4, respectively, but not PIN3, results in two transcripts differing in 12 nucleotides. This sequence (orange) corresponds to the protein motif, located in the long internal hydrophilic loop of the transporter. There is also an additional PIN4c transcript present in publicly available transcriptomes.
(B) Amino acid alignment of the region around the 4-amino acid motif changed by AS in the PIN3 clade proteins (boxed in pink) in Arabidopsis thaliana, including the closest PIN paralogs.
(C) Table shows the number of RNA-seq reads spanning the exon1-exon2 junction corresponding to the detected PIN7 transcripts in selected Arabidopsis thaliana tissue sources. Their ratio was calculated as a percentage of total reads mapped to this area as assessed from the genome browser graphic interface.
(D) Protein sequence alignment showing the conservation of AS in the PIN3 clade of auxin transporters among rosids.
(E) Accumulation of [3H]NAA in BY-2 cells following induction of Arabidopsis thaliana G10-90∷XVE≫AtPIN7a-GFP and G10-90∷XVE≫AtPIN7b-GFP cDNAs with 1 μM β-estradiol. [3H]NAA accumulations within the observed period show no difference between both tested isoforms. The values shown in box plots were normalized to the average maximum of the [3H]NAA accumulation rates in the non-induced lines. Middle line shows median, the box corresponds to the 25% and 75% quartiles, the whiskers represent minima and maxima (n = 9).
See also Supplemental Figure 1.
Functionally relevant AS events are commonly evolutionarily conserved (Keren et al., 2010; Reddy et al., 2013), therefore we sought for available validated transcripts to determine whether similar splicing events occur in orthologous PIN3 clade genes in other dicot species (Bennett et al., 2014; O’Leary et al., 2016). We found examples of such mRNAs besides members of the Brassicaceae family, for instance, also in Abrus precatorius (Fabaceae), which documents the conservation of this AS event in rosids, a plant clade which diversified more than 100 million years ago (Li et al., 2019) (Figure 1D). PIN4c did not show any deeper evolutional conservation. Thus, at least some genes of the PIN3 clade are regulated by the same type of AS, across several plant families, which suggests that these AS events may have a relevant biological function.
PIN7a and PIN7b transport auxin with comparable rates in tobacco cells
To test whether both protein isoforms indeed function as auxin transporters, we exemplified this on the expression of the PIN7a and b cDNA variants tagged with GFP, respecting the design of the original PIN7-GFP construct (Blilou et al., 2005), under the control of the β-estradiol-driven promoter in tobacco BY-2 cells (Petrasek et al., 2006; Müller et al., 2019). Following induction of both transgenes, we observed a comparable decrease of radioactive-labeled auxin accumulation inside the BY-2 cells (Figure 1E) and the time course of the auxin accumulation drop appeared to be similar for both constructs (Supplemental Figure 1B), in accord to that of PIN7a cDNA, examined previously (Petrasek et al., 2006). These experiments reveal that PIN7a and PIN7b code for true auxin exporters, which transport auxin at similar rates in tobacco cell cultures.
AS changes subcellular dynamics of PIN proteins
Polarity and dynamic intracellular trafficking are essential functional attributes of PINs (Adamowski and Friml, 2015). We expressed fluorescently-tagged cDNA versions of the respective PIN7 and PIN4 transcripts under their native promoters in Arabidopsis thaliana. Their overall expression patterns resembled those of the PIN7-GFP and PIN4-GFP lines made on the basis of the genomic sequence (Supplemental Figures 2A-2D). At the cellular level, the protein isoform localization did not largely differ from each other in terms of polarity or general subcellular localization in the root tip (Figure 2A). Thus, at a given resolution, it appears that the motif substituted during AS does not change the basic subcellular localization or expression pattern of both PIN7 and PIN4 proteins.

(A) Subcellular localization and polarity of untreated PIN7a-GFP and PIN7b-RFP cDNA-encoded proteins in the primary root meristem cells.
(B) Both PIN7a-GFP and PIN7b-RFP aggregate in the intracellular compartments (BFA bodies) after 30 min of 50 μM BFA treatment, but do not fully co-localize (arrows).
(C) Temporal dynamics of BFA mediated aggregation of PIN7a-GFP and PIN7b-GFP inside cells. The values were determined as a ratio of fluorescence intensities between the cytoplasm and plasma membrane. (*P < 0.05 by ANOVA, n = 12). Data are means ± S. E.
(D) The fluorescence signal from PIN7a-GFP and PIN7b-RFP upon disruption of actin filaments with 20 μM cytochalasin D for 15 min followed by the addition of 50 μM BFA for another 45 min.
(E) PIN7a-GFP and PIN7b-RFP aggregation (arrows) upon disruption of microtubules with 20 μM oryzalin for 15 min, followed by the addition of 50 μM BFA for another 45 min.
(F) Effect of cytochalasin D on the PIN7a-GFP and PIN7b-RFP exit from the BFA bodies upon 45 min pre-treatment with 50 μM BFA and followed by wash-out with 20 μM cytochalasin D for another 90 min.
(G) Effect of oryzalin on the PIN7a-GFP and PIN7b-RFP exit from the BFA bodies upon 45 min pre-treatment with 50 μM BFA and followed by wash-out 20 μM oryzalin.
In (A, B and D-G), the signals of 3 cells in each of 5 root tips were analyzed (n = 15); bars, 10 μm.
See also Supplemental Figure 2.
The anterograde trafficking of proteins towards PM can be effectively blocked by the fungal toxin brefeldin A (BFA). It leads to internal accumulation of the membrane-bound PINs into characteristic BFA bodies (Geldner et al., 2001; Kleine-Vehn et al., 2010). During time-lapse imaging, we observed that while PIN7a-GFP accumulated readily in these intracellular aggregates, PIN7b-RFP formed less pronounced aggregates, co-localizing with PIN7a-GFP incompletely (Figures 2A and 2B; Supplemental Figure 2E). To exclude that the fluorescent tag influences the sensitivity of the PIN7 intracellular trafficking caused by BFA, we compared the response of PIN7a-GFP with additionally generated PIN7b-GFP cDNA expressing lines. We observed that the BFA mediated aggregation of PIN7b-GFP indeed showed a moderate delay, compared with PIN7a-GFP (Figure 2C). These observations suggest that the PIN7 isoforms differ in the speed of their intracellular trafficking pathways or delivery to the PM and the choice of the tag does not appear to interfere significantly with the subcellular dynamics of PIN7.
PIN polarity does not seem to strictly require the cytoskeleton (Glanc et al., 2018), but subcellular PIN trafficking has been proposed to be mediated by two distinct pathways (Geldner et al., 2001; Glanc et al., 2018). The first is dependent on actin filaments (cytochalasin D-sensitive) and occurs in most cells of the root meristem. The second (oryzalin-sensitive) utilizes microtubules and is linked with cytokinesis. Drugs that depolymerize actin filaments (cytochalasin D) and tubulin (oryzalin) (Geldner et al., 2001; Kleine-Vehn et al., 2008b) showed only little effect on the intracellular localization of both PIN7 isoforms when applied alone (Supplemental Figures 2F and S2G). Pretreatment with cytochalasin D prevented the formation of the BFA bodies (Geldner et al., 2001) containing both PIN7 isoforms (Figure 2D). Yet, when we applied oryzalin prior to the BFA treatment, the BFA compartments containing PIN7a-GFP and PIN7b-RFP associated in only very weakly co-localizing structures (Figure 2E).
Next, we tested how the cytoskeleton is involved in the trafficking of both PIN7 isoforms from the BFA bodies to the PM by washing out BFA with cytochalasin D or oryzalin (Geldner et al., 2001). In the presence of cytochalasin D, PIN7a-GFP largely persisted inside the BFA bodies, while the PIN7b-RFP signal was almost absent in these aggregates (Figure 2F). We generally did not see any difference between both isoforms when BFA was washed out with oryzalin (Figure 2G). These data thereby suggest that both PIN7 isoforms use vesicle trafficking pathways that are assisted by a common cytoskeletal scaffold. These pathways differ in their dynamics and the endomembrane components involved and are consistent with previous findings that individual PINs can utilize multiple PM delivery routes (Boutté et al., 2006; Kleine-Vehn et al., 2008b).
AS affects dynamics of PIN7 at the PM but not its ability to relocate in response to tropic stimuli
Complementary to the observations obtained by the pharmacological approach, we tracked intracellular PIN7 dynamics under natural conditions. Auxin transporters of the PIN3 clade change their polar localization on the PM by the reaction to various environmental cues, in particular by switching the light or gravity vectors (Friml et al., 2002; Rakusová et al., 2011; Ding et al., 2011). PIN3 relocation in response to gravity in columella root cells is seen in as little as 2 min, while the relocation of PIN7-GFP requires approximately 30 min to be detected (Friml et al., 2002; Kleine-Vehn et al., 2010; Pernisova et al., 2016; Grones et al., 2018). We examined plants harboring both PIN7a-GFP and PIN7b-RFP cDNAs under short and long gravitropic stimuli. We did not find any difference in relocation dynamics between both isoforms in these experiments (Supplemental Figures 3A-3D). We observed no difference in the polarity change between PIN7a-GFP and PIN7b-RFP in hypocotyl gravitropic (Rakusová et al., 2011; Rakusová et al., 2016) and phototropic (Ding et al., 2011) bending assays (Supplemental Figures 3E-3I; due to limited transparency of hypocotyls, we used lines expressing the cDNAs under strong endodermal SCARECROW (SCR) promoter (Rakusová et al., 2011)). These data indicate that the different subcellular pathways driving both PIN7a-GFP and PIN7b-RFP cargos are not connected with their ability to change polarity in response to tropic stimuli.
Several studies employed the fluorescence recovery after photobleaching (FRAP) analysis to investigate the dynamic turnover of various proteins, including PINs, on PM (Men et al., 2008; Martinière et al., 2012; Langowski et al., 2016). We therefore bleached a region of the PM signal in the root meristem of the PIN7a-GFP and PIN7b-GFP cDNA lines and measured the FRAP in this area. Notably, PIN7a-GFP showed a slower recovery of fluorescence than PIN7b-GFP (Figure 3A). The difference in the recovery speed and also in the overall mobile phase ratio was even more pronounced when we used lines expressing both cDNAs under the stronger SCR promoter (Supplemental Figure 4A); the choice of GFP or RFP tag did not markedly interfere with the fluorescence recovery (Supplemental Figure 4B). To validate our observations further, we generated plants carrying a PIN3∷PIN3Δ-RFP cDNA construct, which lacks the GETK motif corresponding to the four amino acids absent in PIN7b (Figures 1B and 3B). The PIN3Δ-RFP signal showed an incomplete co-localization with the wild type PIN3-GFP variant in the BFA bodies (Supplemental Figures 4C and 4D), and faster recovery on the PM, analogous to that observed for PIN7a and PIN7b (Figure 3B). It therefore appears that the motif altered by AS of PIN7 is required for the regulation of dynamics of individual isoforms within the PM.

(A) The PIN7a-GFP and PIN7b-GFP normalized FRAP, with single-phase exponential fitting curves. PIN7b-GFP shows different fluorescence recovery on the plasma membrane, compared to the longer PIN7a-GFP. The example image of the bleached region in cells is shown on the inset.
(B) The PIN3Δ-RFP and PIN3-GFP normalized FRAP, with single-phase exponential fitting curves. PIN3Δ-RFP, which lacks the 4 amino acid motif to mimic the properties of the PIN7b isoform, shows faster fluorescence recovery, in comparison to the control PIN3-GFP.
Bars, 5 μm. Data are means, error bars are ± S. E. At least 3 membranes in 5-6 root tips were analyzed in each experiment (n ≥ 20).
See also Supplemental Figures 3 and 4.
PIN7 splice isoforms are functionally different, based on the phenotype rescue tests
To explore the role of the PIN7 splice isoforms in plant development, we tested the ability of their fluorescently tagged cDNAs to complement the phenotypes associated with the PIN7 locus. We initially selected the phototropic hypocotyl bending assay, which is highly dependent on the activity of the PIN3 clade proteins (Friml et al., 2002; Willige et al., 2013). We were unable to detect significant defects in the single pin7-2 knockout (Friml et al., 2003) itself, even at a detailed temporal resolution (Supplemental Figure 5A). As weak phenotypes of the pin7-2 loss of function mutants are the result of redundancy with other genes from the PIN3 clade (Friml et al., 2003; Blilou et al., 2005; Willige et al., 2013), we further employed a triple pin3-3 pin4-101 pin7-102 knock out (pin347) as a genetic background, which lacks the phototropic response almost completely (Willige et al., 2013). Here, PIN7a-GFP cDNA was able to rescue the phototropic bending, while the PIN7b-RFP cDNA did not show any effect, regardless of whether the native (Figure 4A) or a strong endodermal SCR promoter (Supplemental Figure 5B), was used. The choice of the tag does not appear to have any effect in these assays, as evidenced by the lines where the fluorophore sequences have been swapped (Figure 4B). Together, these data indicate that the motif changed by AS alters the function of the PIN7 protein in Arabidopsis.

(A) Phototropic bending of the etiolated pin347 seedlings carrying the PIN7a-GFP and PIN7b-RFP constructs.
(B) Phototropic bending of the etiolated pin347 seedlings carrying the PIN7a-RFP and PIN7b-GFP constructs.
(C and D) Quantification of the primary root protoxylem defects (C) and lateral root primordia (D) initiation of the pin347 seedlings harboring the PIN7a-RFP and PIN7b-GFP constructs.
On (A), (B) and (D), the middle line corresponds to median, the box corresponds to the 25% and 75% quartiles, the whiskers correspond to minima and maxima, dots represent single data points. The asterisks indicate significance between respective line and the pin347 mutant control (**P < 0.01, ***P < 0.001 by ANOVA). For each line in each experiment, at least 15 seedlings were analyzed (n ≥ 15).
See also Supplemental Figure 5.
PIN7a, but not PIN7b, cDNA rescues the pin347 phenotypes in the hypocotyl bending test, and PIN3 clade auxin efflux carries have been implicated in numerous instances of auxin-mediated development. We therefore tested whether in some them the role of the particular isoform could be prevalent. These processes include: determining of root protoxylem formation (Bishopp et al., 2011) (Figure 4C), lateral root density (Swarup et al., 2008) (Figure 4D), vertical direction of the root growth (Friml et al., 2002; Kleine-Vehn et al., 2010; Pernisova et al., 2016) (Supplemental Figure 5C), lateral root orthogravitropism (Rosquete et al., 2013) (Supplemental Figure 5D), gravity-induced hypocotyl bending (Rakusová et al., 2011) (Supplemental Figure 5E), number of rosette branches after decapitation (Bennett et al., 2016) (Supplemental Figure 5F) and the overall rosette size (Bennett et al., 2016) (Supplemental Figure 5G). Similar to the results above, the PIN7a-GFP cDNA usually almost completely rescued the tested phenotypes, while the contribution of PIN7b-RFP was smaller or even undetectable. This suggests that the functional roles of both isoforms are common, regardless of the phenotype observed.
A fluorescent reporter for studying the PIN7a and b expression
The levels of PIN7 (and PIN4) AS transcripts seem to be at comparable levels in most organs (Figure 1C and Supplemental Figure 1A). However, this may not describe the actual situation at the resolution of individual cells. To address this, we designed a dual fluorescent reporter, which allows for monitoring the activity of the AS of PIN7 in planta and in situ (Figure 5A). Indeed, in the primary root or in the hypocotyl, we observed generally overlapping expression of both isoforms without any obvious tissue preference (Figure 5B; Supplemental Figure 6A). However, there were several instances in the vegetative tissue, where the ratio of reporter signals appears to be uneven. These include early lateral root primordia (Figure 5C), the mature pericycle adjacent to the phloem area (Supplemental Figure 6B), the stomatal lineage ground cells of the cotyledons (Supplemental Figure 6C), and the concave side of opening apical hook (Figures 5D), where the PIN7b-RFP signal prevailed over that of PIN7a-GFP. In general, these data corroborate the presence of both isoforms in most cells and suggest that they may function in a coordinated manner.
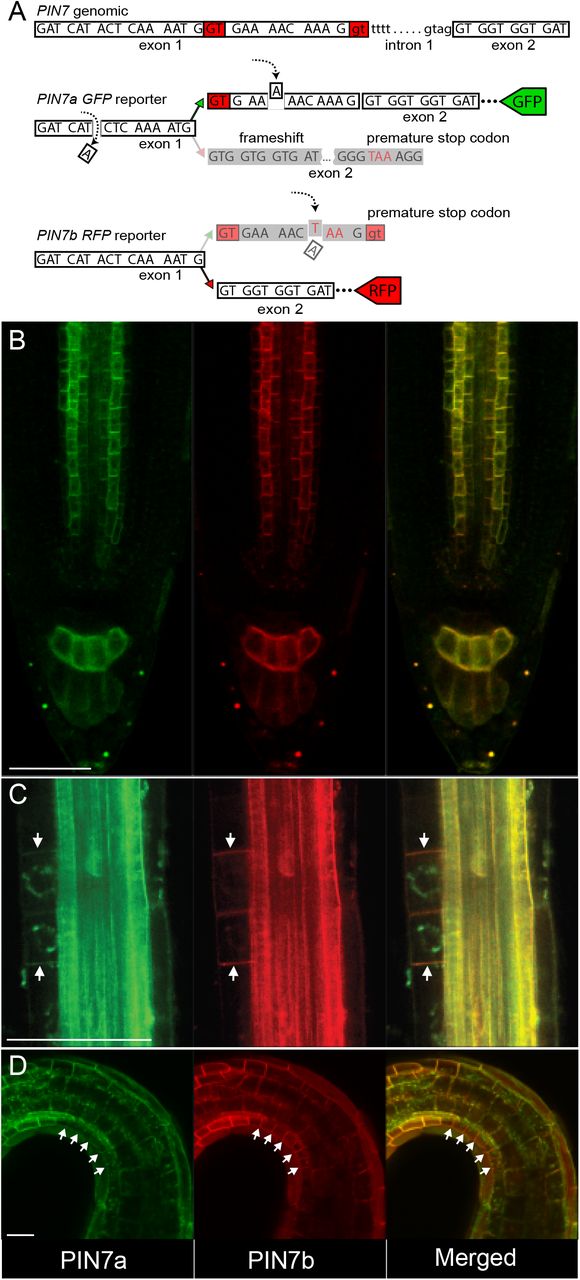
(A) A scheme of the PIN7 splicing reporter. The reporter consists of two constructs: the PIN7a-GFP part contains two point mutations (marked with dashed arrows) and leads to the frameshift and its subsequent restoration when the PIN7a transcript is solely produced. PIN7b-RFP reporter carries a premature stop codon inside the protruding PIN7a region (dashed arrow below).
(B) PIN7a-GFP and PIN7b-RFP expression overlap in the root tip.
(C and D) PIN7b-RFP expression (arrows) PIN7a-GFP in the early lateral root primordia (C) and in the epidermis on the concave side of apical hook (D). The green signal in the perinuclear region of the lateral root primordia is an autofluorescence artifact.
For each tissue, at least 10 plants were analyzed (n ≥ 10). Bars, 50 μm.
See also Supplemental Figure 6.
The combined activity of both PIN7a and PIN7b is required for apical hook formation and tropic responses
The occasional exaggerated response of the pin347 mutants containing the PIN7a-GFP transgene (Supplemental Figure 5E) prompted us to re-design phenotypic tests and adapt our experimental methodology to carefully record the temporal dynamics of the processes linked with the PIN3 clade function. We turned to apical hook development where the time scale measurements of the PIN-mediated development have been well established (Zadnikova et al., 2010). We also analyzed pin4-101 pin7-102 (pin34) mutants and a newly generated pin347 line that carried a combination of both PIN7 cDNA constructs. Similar to gravi- and phototropic experiments, the PIN7b-RFP cDNA generally did not complement the severe pin347 phenotype. Expression of the PIN7a-GFP cDNA in pin347 indeed led to partial rescue of the apical hook formation defects, even surpassing the values observed for pin34. Surprisingly, the simultaneous expression of both PIN7a-GFP and PIN7b-RFP suppressed the dominant effects conferred by the PIN7a-GFP cDNA alone and phenocopied the pin34 mutant (Figure 6A; this effect was not caused by suppressing the expression of PIN7a-GFP by the other transgene, Supplemental Figure 6D). This strongly suggests that both isoforms act in a mutually opposing manner to modulate the processes of apical hook development.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
(A-C) Temporal dynamics of etiolated pin347 seedlings carrying the PIN7a-GFP and PIN7b-RFP transgenes examined (A) during apical hook development, (B) hypocotyl phototropic and (C) gravitropic bending.
Data are means ± S. E. For each data point, the values obtained from 15 vertically grown seedlings were assessed (n = 15).
Apical hook formation is a complex process that involves several bending steps (Zadnikova et al., 2010), and the splicing reporter suggests a slightly different expression pattern of PIN7a and b isoforms during apical hook development (Figure 5D). The hypocotyl phototropic and gravity response includes only a single bending (Rakusová et al., 2011; Rakusová et al., 2016), and the expression of the reporter appears to be uniform in the respective tissue (Supplemental Figure 6A). Tracking dynamic bending of hypocotyls can thus provide a hint to whether one can account for the antagonistic behavior of both isoforms to differential expression or to their different dynamics during subcellular trafficking or at the PM. Similar to apical hook development (Figure 6A), introducing the PIN7b-RFP cDNA did not have any effect on the pin347 phenotype, while the expression of PIN7a-GFP lead to more rapid bending than that observed for the pin34 mutant. Finally, the presence of both PIN7a-GFP and PIN7b-RFP cDNAs in pin347 was reminiscent of the pin34 phenotype in both phototropic and gravity assays (Figures 6B and 6C). We therefore conclude that the shared activity of both PIN7 isoforms, likely conferred by their different trafficking or PM retention properties, but probably not by their differential expression, is required for proper apical hook formation and auxin-mediated tropic responses.
DISCUSSION
In this study, we show that AS diversifies the portfolio of PIN proteins present in Arabidopsis thaliana. At the cellular level, AS does not alter the ability of the PIN7 proteins to transport auxin per se or their overall subcellular localization, but it changes their dynamics on the PM. In general, AS modifies protein properties variably. It can affect protein subcellular localization, ligand binding affinity, enzymatic or transporting activities, protein stability or the presence of covalent post-translational modifications (Stamm et al., 2005; Kelemen et al., 2013). Different covalent modifications alter the subcellular trafficking of most PINs. Phosphorylation sites on serine, threonine or tyrosine residues of various PINs have been identified; their phosphorylation status also changes PIN-mediated tropic responses (Rademacher and Offringa, 2012; Barbosa et al., 2018; Zwiewka et al., 2019). Also PIN ubiquitination (on lysines) and controlled proteolytic degradation act in auxin-mediated processes (Leitner et al., 2012). However, none of the candidate residues required for these modifications is present in the vicinity of the amino acid motif changed by AS. This region, present inside the PIN long hydrophilic loop shows low amino acid conservation and it is perhaps intrinsically disordered (Figure 1C) (Zwiewka et al., 2019). One can therefore speculate whether it may change, perhaps by its length, the ability to assemble internal PIN domains within the long hydrophilic group (Buljan et al., 2013) and modulate their interaction affinity with other factors required for the entry and presence in secretory pathways and/or for PIN7 dynamics at the PM.
We also present genetic evidence that the mutual activity of PIN7a and PIN7b is operational during apical hook formation and tropic responses. We also tested major functional aspects linked with the cellular activity of PIN7 that might be responsible for the observed phenotypes (Adamowski and Friml, 2015). We found the differences between PIN7a and PIN7b at the level of the PM recycling pathways and the stability on the plasma membrane. It was previously demonstrated that the dynamics of PINs on the PM is controlled in principle by two factors: by lateral diffusion in earlier phases and by dynamic recycling from the secretory pathways in longer timelines (Kleine-Vehn et al., 2008a; Kleine-Vehn et al., 2011; Langowski et al., 2016). As the effect of secretion or recycling has been evidenced to be minor within 10 min after photobleaching (Kleine-Vehn et al., 2011; Langowski et al., 2016), it seems that lateral diffusion significantly participates on the differential behavior of PIN7a and PIN7b. In accordance with previous findings (Kleine-Vehn et al., 2011; Feraru et al., 2011; Langowski et al., 2016), slower PIN7a FRAP rates suggest that PIN7 properly functions when only associated with a larger complex or inside stable membrane clusters. According to the earlier published model (Langowski et al., 2016), it therefore seems that PIN7b probably antagonizes PIN7a action by impeding the polar auxin flow provided by PIN7a in these membrane domains (Supplemental Figure 6E).
The mutually antagonistic interaction between two splice isoforms (or a similar coordinated mechanism) has generally been already proposed in Arabidopsis (Szakonyi and Duque, 2018). The seed dormancy regulator DELAY OF GERMINATION 1 (DOG1) is processed into five mRNAs. Only the expression of two or more DOG1 cDNAs under the native promoter rescues the dog1 phenotype by synergistic stabilization of the protein by its multimerization (Nakabayashi et al., 2015). The transcriptional factor HYH (HY5 HOMOLOG) possesses an isoform, which lacks a domain required for proteasomal degradation, which leads to its increased stability and probably works as a semi-dominant splice variant (Sibout et al., 2006; Szakonyi and Duque, 2018). The example of AS of PIN7 is thus one of the first instances in plants where the mutually antagonistic effects of two splice isoforms are observed on a single gene.
EXPERIMENTAL PROCEDURES
Plant material and plant growth conditions
All plant material, except tobacco cell cultures, was in the Arabidopsis thaliana (L.) Heynh., Col-0 ecotype. These mutant and transgenic lines were described previously: PIN3∷PIN3-GFP (Zadnikova et al., 2010), PIN4∷PIN4-GFP, PIN7∷PIN7-GFP (Blilou et al., 2005), pin3-3 pin4-101 pin7-102 (pin347), pin3-3 pin4-101 (pin34) (Willige et al., 2013), pin7-2 (Friml et al., 2003).
For the in vitro cultivation, the seeds were surface-sterilized for 5 h with chlorine gas, plated on 0.5× Murashige & Skoog medium with 1 % sucrose, and then stratified for 2 d at 4°C in darkness. Unless indicated otherwise, the seedlings were grown on vertically oriented plates for 4-6 days under 16 h: 8 h photoperiod, 22: 18°C, light: dark.
The following chemicals were used for treatments: brefeldin A (BFA), cytochalasin D (CytD), oryzalin (Ory), β-estradiol, 2,4-dichlorophenoxyacetic acid (2,4-D), all from Sigma (Sigma-Aldrich, St. Louis, MO, USA). Radioactively labeled auxin accumulation assays were performed with [3H]NAA (naphthalene-1-acetic acid; 20 Ci.mmol−1; American Radiolabeled Chemicals, St. Louis, MO, USA).
DNA manipulations and transgenic work
The genomic PIN7∷PIN7-RFP construct was made by replacing the GFP coding sequence in the original PIN7∷PIN7-GFP construct (Blilou et al., 2005). For creating the PIN7(4)∷PIN7(4)a-GFP and PIN7(4)∷PIN7(4)b-RFP cDNA constructs, the respective cDNAs were cloned into the pDONR221 P5-P2 entry vector (Invitrogen, Life Technologies, Carlsbad, CA., USA) by the Gateway BP reaction (Invitrogen). The XbaI restriction site was introduced at the 1350 bp (for PIN7) or 1341 bp (for PIN4) position of the cDNA coding region for placing the fluorophore tag sequence. In parallel, the PIN7 (or PIN4) promoters were inserted into the pH7WG Gateway vector (Department of Plant Systems Biology, Ghent University, Belgium; Karimi et al., 2002) by the Gibson Assembly kit (New England Biolabs, Ipswich, MA, USA). The tagged PIN7 and PIN4 cDNA entry clones were then recombined with the modified pH7WG destination vector by the Gateway LR reaction (Invitrogen). For the cloning of the PIN7 splicing dual fluorescent reporter, the entry vectors carrying the genomic sequence of PIN7-GFP and PIN7-RFP (Adamowski and Friml, 2015), respectively, were modified by inverse PCR. The resulting constructs were then recombined by the Gateway LR reaction with the PIN7 promoter containing the pH7WG destination vector. The SCR∷PIN7a-GFP and SCR∷PIN7b-RFP constructs were obtained by recombination of the SCR promoter in pDONR221 P1-P5r and PIN7 cDNA entry clones with the pH7WG destination vector by the Multisite Gateway LR reaction. PIN3Δ-RFP cDNA was custom synthesized (Gen9, Ginkgo Bioworks, Boston, MA, USA), cloned into pDONR221 P5-P2 vector and together with the PIN3 promoter pDONR221 P1-P5r construct recombined into the pH7WG vector with Multisite Gateway. The validated binary constructs were transformed into Arabidopsis by floral dipping. For making the estradiol-inducible G10-90∷XVE≫AtPIN7(4)a-GFP, and G10-90∷XVE≫AtPIN7(4)b-GFP constructs, tagged PIN7 or PIN4 cDNA entry clones were recombined with the pMDC7 destination vector (Curtis and Grossniklaus, 2003) by the Gateway LR reaction. Unless stated otherwise, the constructs present in the study were cloned under their natural PIN4 or PIN7 promoter. Primers used in this work are listed in Supplemental Table 1.
For the generation of the stable transgenic lines carrying the cDNA constructs, at least eight independent descendant populations of primary transformants were preselected for the presence of the fluorescent signal. The functional validity of the cDNA constructs was verified in the phototropic bending test, where all candidate lines matched the presented results. Selected lines were used for further phenotypic analysis.
Plant phenotype analysis
The dynamic seedling development was tracked in the custom made dynamic morphogenesis observation chamber equipped with blue and white LED unilateral light sources, infra-red LED back light and a camera for imaging in the infra-red spectra, controlled by the Raspberry Pi3B microcomputer (Raspberry Pi foundation, Cambridge, UK). For the hypocotyl bending assays, the plated seeds were first illuminated for 6 h with white light. The plates were then transferred to the observation chamber for 3-4 d. For the hypocotyl phototropic bending experiments (Friml et al., 2002), the dark-grown seedlings were afterward illuminated for 20 h with unilateral white light and imaged every 20 minutes. For hypocotyl gravitropic bending experiments (Rakusová et al., 2011), the dark-grown seedlings were rotated by 90° clockwise and imaged for 30 h every 60 min. For tracking apical hook development, the seeds were first illuminated for 6 h with white light. They were then transferred to the observation chamber and their development recorded every 4 h for a total 150 - 200 h in infrared spectra. Germination time was set as time 0, when first traces of the main root were observed, individually for each seedling analyzed. Apical hook was determined as an angle between the immobile (non-bending) part of hypocotyl and the distal edge of cotyledons (Zadnikova et al., 2010), using the ImageJ software (Rueden et al., 2017). At least 15 seedlings were analyzed for each line. Each experiment was done at least three times.
For protoxylem defects analysis, 5-d old light-grown seedlings were cleared and analyzed as described previously (Bishopp et al., 2011). For examining lateral root density, 8-d old light-grown seedlings were cleared and observed with a DIC microscope (Dubrovsky et al., 2009). Lateral root density was calculated by dividing the total number of lateral roots and lateral root primordia to the length of the main root as described (Dubrovsky et al., 2009). Vertical Growth Index (VGI), defined as a ratio between main root ordinate and main root length, was quantified on 5-d old seedlings as published previously (Grabov et al., 2005). For measuring the Gravitropic Set-point Angle (GSA), plates with 14-d old light-grown seedlings were scanned and the angles between the vertical axis and five innermost 0.5 mm parts of lateral root were determined as described previously (Roychoudhry et al., 2017). In all cases, 12-20 seedlings were analyzed for each line. Each experiment was done at least three times.
For decapitation experiments, 4-week short-day grown plants were moved into long-day conditions to induce flowering. After the primary bolt reached 10-15 cm, the plant was decapitated. The number of rosette branches was recorded at 7, 10 and 14-d after decapitation (Greb et al., 2003; Waldie and Leyser, 2018). Rosette size was inspected in 18-d light-grown plants prior to documenting. 10 plants were analyzed for each line, the experiment was done three times.
Microscopy
Bright-field microscopy (differential interference contrast, DIC) was conducted on the Olympus BX61 instrument (Olympus, Shinjuku, Tokyo, Japan) equipped with a DP50 camera (Olympus). Routine confocal microscopy was performed on inverted Zeiss Axio Observer.Z1 containing the standard confocal LSM880 and Airyscan modules with 20x/0.8 DIC M27 air, 40x/1.2 W Kor FCS M27 air and 63x/1.40 Oil DIC M27 objectives (Carl Zeiss AG, Jena, Germany). Gravity-induced polarity change experiments were carried out on Zeiss Axio Observer.Z1 with vertically oriented sample position and the 40x/0.75 glycerol objective.
To observe the light-induced polarity change of PIN7a-GFP and PIN7b-RFP, 4-d dark-grown seedlings were irradiated for 4 h with unilateral white light and then imaged with Zeiss Axio Observer.Z1 LSM880 with a vertically oriented sample position, as described (Willige et al., 2013). For analyzing gravity-induced polarity change, 4-d dark-grown seedlings were reoriented by 90° clockwise and imaged 6 and 24 h after rotation, as described (Rakusová et al., 2011).
For BFA treatments, 5-d light-grown seedlings were transferred to liquid 0.5× MS media containing 50 μM BFA. The membrane/cytosol ratio was determined with ImageJ, it was defined as the mean membrane signal intensity divided by the mean fluorescence in the cytosol. For cytoskeleton depolymerizing drug treatments (Geldner et al., 2001), 5-d light-grown seedlings were transferred to liquid 0.5× MS media supplemented with 20 μM of cytochalasin D or 20 μM oryzalin. The co-treatments were done by the direct addition of BFA. For the BFA removal, prior to the addition of the cytoskeleton depolymerizing compounds, seedlings were twice washed out with fresh media and then transferred to that supplemented with the respective cytoskeleton-depolymerizing drug.
Fluorescent recovery after photobleaching (FRAP)
For the FRAP experiments, Zeiss Axio Observer.Z1 equipped with the LSM880 confocal and Airyscan modules and the 40x/1.2 W Kor FCS M27 air objective was used. A rectangular region of interest (ROI) of 40 × 20 pixels was selected on the basal or apical PM of cells inside the vascular cylinder in the primary root meristematic area of 5-d light-growth seedlings. ROI was bleached with the maximum 488 nm laser intensity and fluorescence recovery was documented every 1 or 2 sec for a total 200 sec. Recovery time lapses were analyzed with ImageJ. The Slices Alignment plugin (Tseng et al., 2012) for ImageJ was used for the elimination of cell movement caused by root growth. In parallel, another rectangular ROI (100 × 20 pixels) on the non-bleached cell was selected as a reference. To compensate for the fluorophore bleaching during the recovery period, the data were normalized using equation (Laňková et al., 2016)
 where In is normalized fluorescence intensity, It is the intensity at the specific time point, Imax is the intensity after the initial bleaching, Imax is the intensity before initial bleaching. Itref, Imaxref and Iminref represent the same values for the reference ROI. At least 3 membranes in 4-5 root tips were analyzed in each experiment. Single-phase exponential fitting was applied to the normalized FRAP data (SigmaPlot, Systat Software, Chicago, ILL, US) as described (Sprague and McNally, 2005; Laňková et al., 2016). Recovery half-time is defined as a time required for the fluorescence recovery to reach half of the steady-state fluorescence intensity (Soumpasis, 1983; Sprague and McNally, 2005).
where In is normalized fluorescence intensity, It is the intensity at the specific time point, Imax is the intensity after the initial bleaching, Imax is the intensity before initial bleaching. Itref, Imaxref and Iminref represent the same values for the reference ROI. At least 3 membranes in 4-5 root tips were analyzed in each experiment. Single-phase exponential fitting was applied to the normalized FRAP data (SigmaPlot, Systat Software, Chicago, ILL, US) as described (Sprague and McNally, 2005; Laňková et al., 2016). Recovery half-time is defined as a time required for the fluorescence recovery to reach half of the steady-state fluorescence intensity (Soumpasis, 1983; Sprague and McNally, 2005).
Tobacco cell lines and auxin accumulation assays
Tobacco cell line BY-2 (Nicotiana tabacum L. cv. Bright Yellow 2) was cultivated as described (Müller et al., 2019). BY-2 cells were transformed with the pMDC7 constructs by co-cultivation with Agrobacterium tumefaciens strain GV2260 as earlier described (Petrasek et al., 2006; Müller et al., 2019). The transgene expression was maintained by cultivation on media supplemented with 40 μg.ml−1 hygromycin (Roche) and 100 μg.ml−1 cefotaxime (Sigma). Accumulation assays of radioactively labeled auxin were performed as previously published (Delbarre et al., 1996; Petrasek et al., 2006) with cells cultured for 2 days in β-estradiol or DMSO mock treatments. Presented results are from 3 biological replicates for each representative PIN7a and PIN7b line and were confirmed for each on two other independent genotypes. Independent β-estradiol inductions were done within 3-9 months after the establishment of the cell suspension on lines, which showed comparable levels of the expressed PIN7-GFP signal.
Statistics and sequence analysis
For two groups mean comparison, Student’s t-test was applied. Statistical analysis of multiple groups was performed by one-way ANOVA with subsequent Tukey HSD post-hoc test. All statistical tests, including normality tests, were performed in R-studio IDE (R-studio, Boston, MA, USA). In the box plots, the whisker length was defined as Q ± 1.5 × IQR, where Q is the corresponding quartile and IQR is the interquartile range. For creating the multiple sequence alignments, the protein sequences were aligned using the Clustal Omega algorithm (Sievers et al., 2011) and graphically outlined by Mega-X, using default ClustalX color code (Kumar et al., 2018).
AUTHOR CONTRIBUTIONS
K., M. H., R. F., Z. V., D. R., and J. P. conducted experiments. J. H. enabled making part of this project in his laboratory. S. S. and J. F. provided unpublished material. I. K., J. P., T. B. J., M. J. F., J. F., J. P., and K. R. conceived the research and designed experiments. K. R. and I. K. wrote the manuscript. All authors read and commented on the final version of the manuscript.
DECLARATION OF INTERESTS
The authors declare no competing interests.
ACKNOWLEDGMENT
We thank Claus Schwechheimer for the pin34 and pin347 seeds, Huibin Han for assistance with the hypocotyl imaging, Dmitry Konovalov for help with the evolutional analysis, Karel Müller for the initial qRT-PCR analyses of the tobacco cell lines and Ksenia Timofeyenko and Jozef Mravec for discussions. This work was supported by the Czech Science Foundation (16-26428S) to I. K., M. H. and K. R, and the Ministry of Education, Youth and Sports of the Czech Republic (MEYS, CZ.02.1.01/0.0/0.0/16_019/0000738) to K. R. Imaging Facility of the Institute of Experimental Botany is supported by MEYS (projects LM2018129-Czech BioImaging, ERDF CZ.02.1.01/0.0/0.0/16_013/0001775 and OPPK CZ.2.16/3.1.00/21519).
Footnotes
↵7 Lead Contact
REFERENCES




