

Abstract
Social animals benefit from their group-mates, so why do they sometimes kill each other’s offspring? A major barrier to understanding the evolution of infanticide is a lack of data from natural populations. Especially when perpetrated by females, infanticide remains poorly understood, because the increased mating opportunities that explain infanticide by males do not apply in females. Using 30 years of data from several spotted hyena groups, we show that infanticide is a leading source of juvenile mortality, and we describe the circumstances under which it occurs. In all observed cases, killers were adult females, but victims could be of both sexes. Killers only sometimes consumed the victims. Mothers sometimes cared for their deceased offspring, and sometimes consumed the body. Killers tended to be higher-ranking than the mothers of victims, and killers were sometimes aided by kin. Our results are consistent with theory that infanticide by females reflects competition among matrilines.
Introduction
Why do animals kill the offspring of their group members? Infanticide (Hausfater & Hrdy, 1984) has been observed in both lab settings and the wild, and may be committed by either males or females (Agrell et al., 1998; Hrdy, 1979). Infanticide is especially prominent in mammals, where it has been observed in over 100 species, but it has also been documented in birds, fish, and invertebrates (Agrell et al., 1998; Hausfater & Hrdy, 1984; Hrdy, 1979). In species where infanticide represents a common source of infant mortality, infant defense and avoidance of infanticidal individuals may function importantly in the developmental biology and social behavior of both adults and juveniles (Balme & Hunter, 2013; Lowe et al., 2018; Muller & Wrangham, 2002; Packer & Pusey, 1983). However, because infanticide events occur infrequently and take place rapidly, it can be difficult to identify the circumstances under which they occur and to assess their importance. For example, the existence of infanticide by male lions, now a canonical example of infanticide, was hotly debated as recently as the late 1990s (Dagg, 1998; Packer, 2000; Silk & Stanford, 1999).
Infanticide by females is less well understood than infanticide by males, despite the fact that it may be the more widespread and frequent form of infanticide (Clutton-Brock & Huchard, 2013; Digby, 2000). Infanticide by males is usually explained by the sexual selection hypothesis, where fitness advantages accrue to males by killing infants sired by others (Balme & Hunter, 2013; Lukas & Huchard, 2014). Despite being less well studied than infanticide by males, infanticide by females occurs in at least 89 mammalian species and may be motivated by competition with other females, depending on social organization and ecology (Digby, 2000; Lukas & Huchard, 2019). In cooperatively breeding species, infanticide by females is committed primarily by the dominant breeder and serves to increase her number of available helpers by preventing would-be breeders from reproducing (Clutton-Brock et al., 1998; Corbett, 2010; Creel & Creel, 1998; Lukas & Huchard, 2019). In plural breeders with female philopatry, infanticide by females is thought to be more likely to occur in species where dominance hierarchies are influenced by nepotism (i.e., the ranks of daughters are influenced by the ranks of their mothers) (Lukas & Huchard, 2019). Furthermore, it is predicted that in such societies, infanticide should be perpetrated by individuals of high rank against lower ranking groupmates (Clutton-Brock & Huchard, 2013; Lukas & Huchard, 2019; Vullioud et al., 2019).
Lukas & Huchard (2019) provide the most comprehensive view to date on the evolution of infanticide by females, but they identify a lack of observational data as a remaining obstacle. Some species in their study were represented by only a few observations; in fact, the details and context of infanticidal behavior of spotted hyenas (Crocuta crocuta) in their study was inferred from only two observations of infanticide (White, 2005). In this species, infanticide has been posited as a leading source of juvenile mortality, and has been implicated in the evolution of masculinized genitalia in females (Muller & Wrangham, 2002), but reports of the frequency and context of infanticide are lacking.
Here we use three decades of behavioral observations from multiple social groups to provide the first quantitative assessment of the prevalence and context of infanticide in spotted hyenas, a plural breeding species with female philopatry and nepotistic rank inheritance.
Methods
Study Animals
Spotted hyenas are large carnivores found widely across sub-Saharan Africa (Holekamp & Dloniak, 2010). Individuals reside in mixed-sex clans, each of which contains multiple matrilineal kin groups and is structured by a linear dominance hierarchy (Frank, 1986). The dominance hierarchy is maintained by social support from groupmates, especially kin (Smith et al., 2010; Strauss & Holekamp, 2019b; Vullioud et al., 2019), and rank is inherited through a learning process akin to what is found in many cercopithecine primates (Holekamp & Smale, 1991). Spotted hyena societies are characterized by fission-fusion dynamics where individuals associate in subgroups that change composition throughout the day (Smith et al., 2008). Males disperse during the years after reproductive maturity, which occurs at around 24 months of age (Holekamp et al., 2012). Spotted hyenas are polygynandrous and breed year-round. Females give birth to one or two (and rarely, three) cubs in an isolated natal den, where they are maintained for a few weeks before being moved to the clan’s communal den. The communal den may contain up to 31 cubs at any given time (Johnson-Ulrich & Holekamp, 2020), and cubs typically remain in or around the den until they are 9-12 months of age (Holekamp & Dloniak, 2010). These cubs, which belong to several different mothers, are often left unattended during much of the day while the mothers are away. Both mothers and other groupmates visit the communal den regularly, either alone or with clan-mates. Starting at 1-2 months of age, cubs emerge from the den to socialize when their mothers are present and, as they get older, when their mothers are absent.
Study Area
Data presented here were collected from two study areas in Kenya near the Tanzanian border. Most observations come from eight clans in the Masai Mara National Reserve (MMNR, 1510 km2), a savanna ecosystem in southwestern Kenya that is contiguous with the Serengeti National Park in Tanzania and grazed year-round by multiple herbivore species (Holekamp et al., 1997). Data were collected in MMNR from 1988 to 2018 during 149,377 observation sessions; we observed one clan for the entire study period, and seven other clans during subsets of that period. Our second study area was in Amboseli National Park (ANP, 392 km2), which is located in southeastern Kenya; data were collected from two clans in ANP from 2003 to 2005 during 4,651 observation sessions (Watts et al., 2011).
For describing cases of infanticide and their frequency, we use data from all ten clans of spotted hyenas located in both study areas to capture the full breadth of circumstances under which infanticide occurs. For investigations into rank-related patterns of infanticide, we limited the dataset to only our six most well-studied clans of hyenas in the MMNR. We elected to use this restricted dataset because these clans live under similar ecological conditions, because all covariate data were available for these clans, and because these clans account for the majority of our data (75.7% of juvenile mortality from the larger dataset).
Data Collection and Analysis
Data were collected during twice-daily observation periods that took place around dawn and dusk. Observers used vehicles as mobile blinds from which to find and observe hyenas. Scan sampling (Altmann, 1974) was used to collect demographic data. Maternity was determined based on nursing associations and genotyping. Data documenting specific types of social interactions, including observed infanticide events, were collected using all-occurrence sampling (Altmann, 1974). Individual hyenas were identified by their unique spot patterns, and the sexes of juveniles were determined based on the dimorphic morphology of the erect phallus (possible after cubs are 2-3 months old). Juvenile age was determined (to ± 1 week) based on appearance when first seen. Social ranks of adult females were determined based on the outcomes of aggressive interactions as described elsewhere (Strauss, 2019; Strauss & Holekamp, 2019a). All statistical analysis and visualization was done using the statistical software environment, R (R Core Team, 2020).
Juvenile mortality
We assessed various causes of mortality among juveniles less than 1 year of age. In our 30 years of data, the most common sources of mortality for young juveniles were starvation, humans, lions, siblicide, and infanticide (n = 102 cases with known mortality sources). Starvation was identified in cases where juveniles were observed to be becoming progressively gaunter before they vanished, or in cases where dead juveniles were found in an emaciated state. Death by humans was assigned when there was a clear anthropogenic cause of mortality; for example, hit by a car, speared, poisoned, etc. Death by lions was either observed or could be inferred based on the deep, widely spaced puncture wounds typically inflicted during lion attacks. Death by siblicide occurred when a cub prevented its littermate from nursing (Golla et al., 1999; Smale et al., 1999).
We identified 21 cases of infanticide in our dataset, divided into two categories based on our confidence in the cause of death. We identified 17 ‘confident’ cases of infanticide, where we observed a cub being killed by another hyena by having its skull crushed or found a cub dead as a result of having its skull recently crushed (Figure 1). Additionally, we identified four ‘likely’ cases of infanticide, where a dead cub was found but no information on the state of the skull was available (usually because the victim had already been largely consumed), but other common aspects of ‘confident’ cases of infanticide were noted (e.g., mother guarding or grooming the victim, victim consumed by mother or others, victim found at communal den). Finally, cubs dying due to causes other than those listed here were combined into an ‘other’ mortality source.

Adult female spotted hyena carrying a cub recently killed by infanticide. Infanticide was typically achieved by crushing the skull, as seen here. Photo by Kate Yoshida.
To assess the relative contributions of different sources of mortality to overall juvenile mortality, we first considered two categories of juvenile mortality: cases where the mother preceded the cub in death, and cases when she did not. We separated mortality in this way because cubs under 12 months old whose mothers die also perish in almost all cases. These juveniles are still dependent on their mother’s milk and therefore 80.1% of juvenile deaths in this category are a result of starvation. In the remaining 19.9%, although cause of death was not directly starvation (causes of death were humans [n=3], infanticide [n=1], and lions [n=3]), prolonged absence of the mother combined with nutritional stress likely motivated the cubs to engage in abnormal behavior. To account for these cases, we re-classified these known mortality sources to be ‘death of mother,’ and treated this mortality source separately from the others when determining the frequency of different causes of mortality. For juveniles with unknown cause of death whose mothers preceded them in death (n = 38), we inferred their mortality source to be ‘death of mother.’
We then used a Bayesian multinomial model to predict the mortality source for the remaining juveniles with living mothers (n = 66) as a function of their age at death, and used this model to infer the frequency of mortality sources in the sample of juveniles whose cause of death remained unknown. We used posterior samples from this model to generate predictions for the cause of death for each juvenile with an unknown mortality source (n = 566) and the probability of different mortality sources as a function of age at death. This model was initialized with weak, uninformative priors, and three chains were run for 15000 iterations each (7500 warmup); the model was implemented in Stan using the RStan and brms R packages (Burkner, 2017; Stan Development Team, 2018).
Results
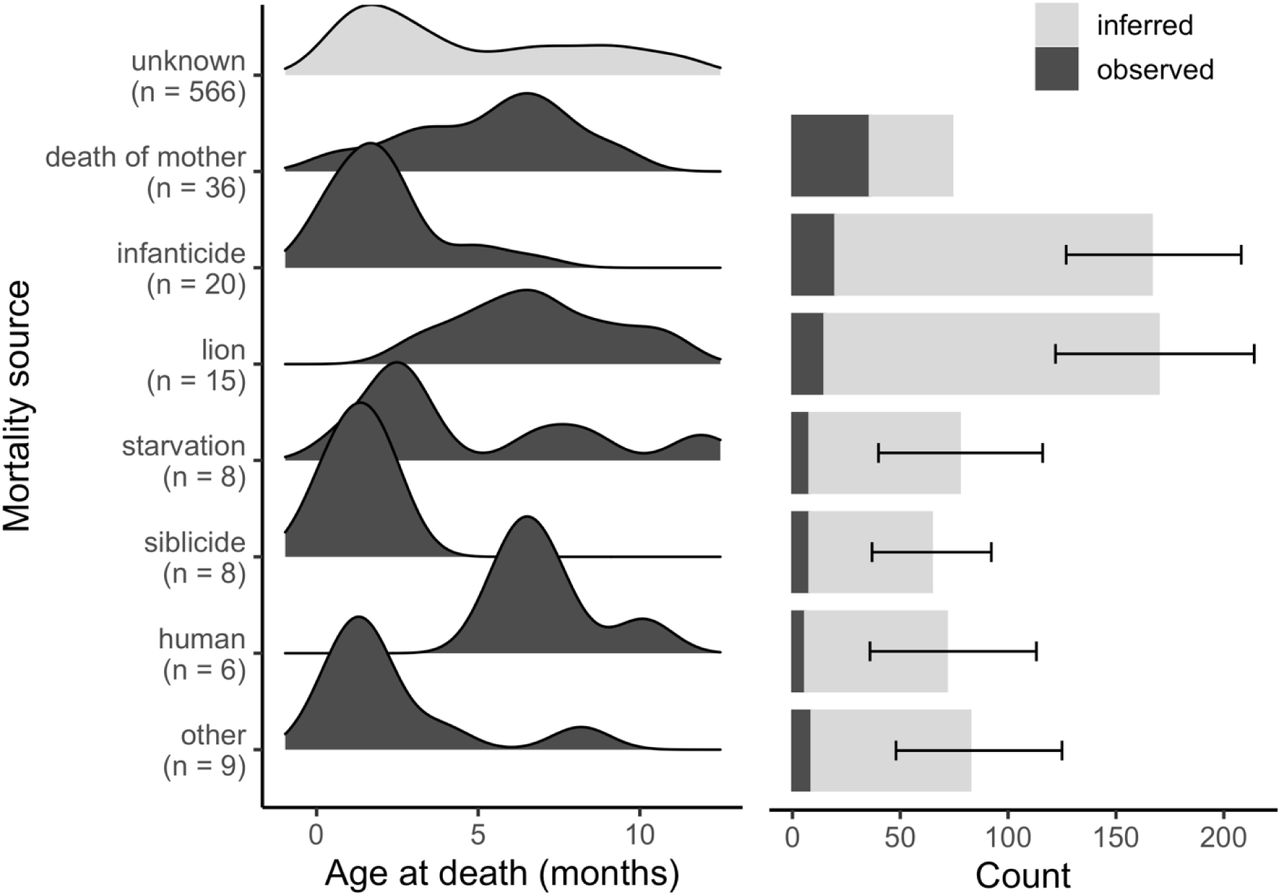
Infanticide was a leading source of mortality for cubs under one year old. Of the 102 cases of death of den-dependent juveniles with a known mortality source (of 706 total juvenile mortality cases), infanticide accounted for 19.6% of mortality (Figure 2, dark bars). This number places infanticide as the second largest mortality source for den-dependent juveniles, ranking below death of mother (35.3%) but above lions (14.7%) (Figure 2, dark bars). After inferring cause of death based on age at death for cases of juvenile mortality with unknown cause (Figure 2, light bars), infanticide and lions were the leading cause of death for juveniles, respectively accounting for a mean of 23.43% (95% prediction interval = [17.14,29.19]) and 23.78% [16.85, 30.46] of juvenile mortality. Cubs who died young were most likely victims of infanticide, whereas cubs who died at ages over 4.2 months were most likely victims of lions (Figure 3). Death of mother, starvation, siblicide, humans, and other mortality sources together accounted for the remaining roughly 50% of total juvenile mortality (Figure 2).

Age distribution of observed causes of juvenile (<1-year-old) mortality in spotted hyenas (left), and frequencies of the 6 leading causes of juvenile mortality (right). Dark bars indicate cases where the mortality source was known (n = 102). Lighter bars indicate inferred mortality sources for cases where the cause of mortality was unknown (n = 604). Error bars indicate the 95% prediction intervals for inferred mortality cases based on a Bayesian multinomial model of mortality source as a function of age. Mortality by ‘death of mother’ was inferred analytically rather than statistically.

Probability of different mortality sources as a function of age at death, as estimated by a Bayesian multinomial model. Juveniles under 4.2 months old are most at risk of death by infanticide, whereas older juveniles are most at risk of death by lions.
In every case of infanticide with a known killer, the act was perpetrated by an adult female (n = 10). All cases of infanticide occurred at a communal den, although in one case the victim was killed by other groupmates as its mother was transferring it from the natal den to the communal den. Attackers sometimes acted alone (n = 7), and other times were aided by groupmates (n = 3). In three cases where infanticide took place while the victim’s mother was present, multiple hyenas displayed aggression against the mother while her offspring was being attacked. In one of these events, the highest-ranking female killed a low-ranking juvenile while her offspring chased away the victim’s mother. In two other cases where the victim’s mother was present, multiple hyenas attacked the mother while the perpetrator killed the cub. In cases where females committed infanticide unaided, they often did so during what appeared to be normal social behavior, and in a few cases prosocial ‘groan’ vocalizations were emitted by the attacker immediately before attacking. Close kinship did not prevent females from committing infanticide: in one case, a female coaxed each of her full sister’s two offspring out of the den by groaning, then killed both cubs (previously reported in (White, 2005)). However, although we did not have full pedigree data available for most killed infants, infanticidal females most often killed juveniles other than those born to their closest relatives. Thus, this prior report of infanticide against kin was not representative of typical patterns of infanticide in this species.
Infanticidal females typically targeted young juveniles (<5 months old), although we did observe two cases of infanticide in older juveniles (Figure 2). Victims with known sexes were evenly split between males (n = 4) and females (n = 5), suggesting that juveniles of both sexes were equally likely to be attacked by conspecifics. An unpaired comparison of attacker and victim ranks revealed that infanticidal adult females were on average higher ranking than the mothers of victims (Welch’s two-sample t-test: t = −3.44, df = 20.99, p = 0.003; Figure 4). Whereas victims of infanticide were of diverse ranks, perpetrators of infanticide where almost exclusively high-ranking females.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Rank distributions of killers and mothers of cubs killed by infanticide. Ranks of infant-killers were on average higher than the ranks of mothers of infants killed by infanticide (Welch’s two-sample t-test: t = −3.44, df = 20.98, p = 0.003).
Victims’ bodies were consumed by one or more hyenas in 11 out of 21 cases; they were sometimes consumed by the killer (n = 3) or the killer’s offspring (n = 3), sometimes by the mother of the dead infant (n = 4), and sometimes by other group-members (n=3; note that these numbers don’t add to 11 because multiple hyenas were often observed consuming infanticide victims). When given the opportunity, mothers sometimes (n = 7) groomed or otherwise cared for their deceased offspring after its death. In 3 cases, observers noted unusual, unprovoked, and distressed-sounding vocalizations emitted by the mother. In 5 cases, the body was either carried away from the den or completely consumed in less than 50 minutes, with one cub being completely consumed in 13 minutes. These records may underestimate the frequency with which victims are consumed because observers collected the victim’s body for biological samples in 6 cases, and halted observations before the fate of the body was determined in 4 cases. However, it is clear from our observations that the remains of victims of infanticide do not persist for long.
Discussion
We found that infanticide is a significant source of mortality among young juvenile spotted hyenas (Figure 2). In support of predictions about infanticide in nepotistic societies (Clutton-Brock & Huchard, 2013; Lukas & Huchard, 2019; Vullioud et al., 2019), infanticide was typically perpetrated by high-ranking females against the offspring of lower-ranked groupmates. This supports the theory that infanticide by females in spotted hyenas may reflect matrilineal competition. Social support has been found to be a significant force for establishing, maintaining, and changing dominance in spotted hyenas (Engh et al., 2000; Strauss & Holekamp, 2019b; Vullioud et al., 2019), and kin provide a significant portion of that support (Smith et al., 2010). Thus, reducing another female’s matriline size via infanticide may serve to prevent potential coalitions of lower-ranked matrilines aspiring to improve their status. Interestingly, we did observe one case of infanticide directly related to escalated aggression between matrilines. Observers arrived at the den to find a recently killed offspring from a high-ranked matriline. Many hyenas were acting highly agitated, and roughly one hour later we observed a coalition of related low-ranking adult females viciously attacking members of the high-ranking matriline. Our observations of infanticide events where the perpetrator’s kin assisted by chasing away the victim’s kin reflects how infanticide might both arise as a function of disparities in social support within groups and serve to reinforce those disparities.
Our findings also indicate the dual use of the communal den as protection against both outside predation sources such as lions, and intraspecific killing via infanticide. The diameter of den holes limits the size of individuals able to enter the den, such that cubs can escape inside when threatened by adults or large predators, which cannot follow cubs into the den hole. This function has been born out in our observations: we have seen larger cubs killed by lions while they attempted escape into the communal den, as well as cubs escaping into the den while lions attempt (and fail) to extract them with their paws (unpublished data). In our observations of infanticide, we occasionally observed perpetrators coaxing cubs out of the den before attacking them, and in one case a targeted juvenile attempted to escape into the den but was caught and killed before reaching safety (Supplemental Videos). Mothers of victims sometimes displayed grooming or other maternal behaviors towards the deceased cub, which may be of interest to those interested in comparative thanatology (Anderson & Anderson, 2016; Carter et al., 2020).
Our results highlight the conflicting forces that characterize the lives of gregarious animals. The prevalence of infanticide highlights risks faced by females choosing to rear their cubs in a social environment. More solitary individuals could choose to keep their cubs at a natal den for several months, and thereby avoid the potential risks of infanticide, but female hyenas rarely choose to do so (White, 2007).
This suggests that the benefits of social integration for cubs raised at communal dens outweigh the costs to females imposed by the risk of conspecific infanticide. However, these conflicting forces may also lead to a tradeoff between appropriate social development and survival if the behavior required for social integration in early-life is associated with infanticide risk.
Finally, our results demonstrate the power of using long-term data to study rare or difficult-to-observe phenomena. Rarity of phenomena can obscure their importance, and ephemeral, high-impact events like infanticide are the types of phenomena that require extensive data collection to permit analysis. The frequency of infanticide relative to other sources of infant mortality suggest that it is a significant feature of spotted hyena biology, which was unclear prior to this study (Muller & Wrangham, 2002). Continued, direct study of this phenomenon has the potential to answer outstanding questions about the function of infanticide in this species and further reveal the way in which it moderates conflicts of interest within complex societies.
Data Availability
Data and code used for this study are available on github (https://github.com/straussed/infanticide).
Author contributions
The data were collected by KEH, BMP, and EDS. Data processing and analysis were done by AKB and EDS. All authors wrote and edited the manuscript.
Acknowledgments
We thank the Kenya Wildlife Service, the Narok County Government, and the Kenyan National Committee on Science, Technology and Innovation, the Naboisho Conservancy, the Mara Conservancy and Brian Heath for permissions to conduct this research. This work was supported by NSF grants OISE1853934 and IOS1755089 to KEH, and by an NSF Graduate Research Fellowship to EDS. This work was also supported in part by NSF Grant OIA 0939454 (Science and Technology Centers) via “BEACON: An NSF Center for the Study of Evolution in Action.” AKB was supported by the MSU Honors College.
Footnotes
References




