

Abstract
Background Morbidity and mortality from COVID-19 caused by novel coronavirus SARS-CoV-2 is accelerating worldwide and novel clinical presentations of COVID-19 are often reported. The range of human cells and tissues targeted by SARS-CoV-2, its potential receptors and associated regulating factors are still largely unknown. The aim of our study was to analyze the expression of known and potential SARS-CoV-2 receptors and related molecules in the extensive collection of primary human cells and tissues from healthy subjects of different age and from patients with risk factors and known comorbidities of COVID-19.
Methods We performed RNA sequencing and explored available RNA-Seq databases to study gene expression and co-expression of ACE2, CD147 (BSG), CD26 (DPP4) and their direct and indirect molecular partners in primary human bronchial epithelial cells, bronchial and skin biopsies, bronchoalveolar lavage fluid, whole blood, peripheral blood mononuclear cells (PBMCs), monocytes, neutrophils, DCs, NK cells, ILC1, ILC2, ILC3, CD4+ and CD8+ T cells, B cells and plasmablasts. We analyzed the material from healthy children and adults, and from adults in relation to their disease or COVID-19 risk factor status.
Results ACE2 and TMPRSS2 were coexpressed at the epithelial sites of the lung and skin, whereas CD147 (BSG), cyclophilins (PPIA and PPIB), CD26 (DPP4) and related molecules were expressed in both, epithelium and in immune cells. We also observed a distinct age-related expression profile of these genes in the PBMCs and T cells from healthy children and adults. Asthma, COPD, hypertension, smoking, obesity, and male gender status generally led to the higher expression of ACE2- and CD147-related genes in the bronchial biopsy, BAL or blood. Additionally, CD147-related genes correlated positively with age and BMI. Interestingly, we also observed higher expression of ACE2- and CD147-related genes in the lesional skin of patients with atopic dermatitis.
Conclusions Our data suggest different receptor repertoire potentially involved in the SARS-CoV-2 infection at the epithelial barriers and in the immune cells. Altered expression of these receptors related with age, gender, obesity and smoking, as well as with the disease status might contribute to COVID-19 morbidity and severity patterns.
Introduction
A novel coronavirus SARS-CoV-2 leading to COVID-19 was identified for the first time in December 2019 and as of today has infected already more than 4.3 million and killed more than 290 000 people worldwide (as of 13th of May 2020)1. SARS-CoV-2 virus has very high genome sequence similarity to two other human coronaviruses SARS-CoV and MERS-CoV2. Therefore, it is highly possible that SARS-CoV-2 uses similar approaches to cell entry and replication in various cells and tissues. In fact, it is already known that SARS-CoV-2 uses the same receptor ACE2 to enter the cells via its structural spike glycoprotein (S), yet with much higher affinity, which might translate to the massive SARS-CoV-2 spread as compared to SARS-CoV3. Host transmembrane protease serine 2 (TMPRSS2; encoded by TMPRSS2) cleaves spike protein into two subunits, which is a necessary step for the virus fusion to cellular membranes and entry the cell3. Recent structural studies revealed that this process can be potentially inhibited by B0AT1 (S6A19; encoded by SLC6A19), an amino acid transporter, presence of which may block the access of TMPRSS2 to the cleavage site on ACE24 (Table S1).
The expression of ACE2 (encoded by ACE2) is very ubiquitous in the lung, heart, kidney and intestine, but it is rarely expressed in immune cells5,6. However, immune cells can be potentially infected by SARS-CoV-2, as in case of MERS-CoV and SARS-CoV7,8. Thus, it is highly possible that there are other receptors for virus entry in different cell types. Indeed, similarly with what has been shown in SARS-CoV, another receptor - CD147, called also basigin (encoded by BSG), has been recently shown to act as a receptor for SARS-CoV-2 in T cell lines and in cell lines of epithelial origin9,10. Interestingly, the same receptor is a putative receptor not only for SARS-CoV, but also for HIV-1 and measles, as well as is a receptor for malaria entry to erythrocytes11–14. An anti-CD147 antibody that blocks infection with SARS-CoV-2 in vitro and its humanized drug (meplazumab) has been already used in a clinical trial in patients with COVID-19 pneumonia. Meplazumab seemed to facilitate viral clearance, return to normal levels of lymphocyte count and decrease in CRP9,15. The percentage of improvement in patients with severe and critical COVID-19 presentations seemed higher in weekly meplazumab treatment compared to patients on the conventional treatment15.
CD147 is a transmembrane receptor interacting with several extracellular and intracellular partners forming a transmembrane supramolecular complex16. A group of extracellular molecules can bind and activate CD147 are cyclophilins A and B (encoded by PPIA and PPIB), acute phase protein S100A9, E-selectin (encoded by SELE) and platelet glycoprotein VI (encoded by GP6)17–20. Additionally, CD147 has three Asn glycosylation sites, to which high mannose-type and complex-type glycans might bind21. Spike protein of SARS-CoV-2 is highly glycosylated22, which increases the chance of binding to the cells. It is possible that the virus can only attach to the cell surface and induce different cellular programs, leading to cell overactivation, exhaustion and death, especially in the severe phase of the disease, where cytokine storm and lymphopenia is commonly reported23.
Cyclophilins A and B have been shown to interact with non-structural protein 1 (nsp1) of SARS-CoV intracellularly24,25. They can be incorporated to the viral capsid and released, which further enables virus binding to CD147 and subsequent infection of CD147-expressing cells. Cyclophilins play a critical role in the replication process of HIV-1, HCV and many other viruses12,26. Cyclosporine A, a strong immunosuppressive agent acts mainly via binding to cellular cyclophilins. In the same mechanism, cyclosporine A suppresses replication of various coronaviruses27. Thus, cyclophilins were identified as targets for pan-coronavirus inhibitors28. Nevertheless, it is still not known if cyclophilins can interact with SARS-CoV-2.
Intracellular and transmembrane partners of CD147 are equally important in the infection process of HIV, measles and SARS-CoV. It has been shown that truncation of cytoplasmic tail of CD147 prevents HIV infection29. Therefore, it is possible that these molecules can take part and regulate the process of SARS-CoV-2 entry through CD147 in a comparable way as TMPRSS2 and SLC6A19 for ACE2. Main transmembrane partners of CD147 with direct binding sites to CD147 are monocarboxylate transporters 1-4 (MCT1-MCT4; encoded by: SLC16A1, SLC16A7, SLC16A8, SLC16A3), main glucose transporter, GLUT-1 (encoded by SLC2A1) and CD44 (encoded by CD44), an important receptor for hyaluronan-main component of extracellular matrix30–32. CD147 also interacts intracellularly with integrins α3β1 and α6β1 (encoded by ITGA3, ITGA6, ITGB1) indirectly via CD98 (SLC7A5), CD43 (SPN), MCT4 and galectin-3 (LGALS3)33–36. In addition, CD147 suppresses two important protein complexes by direct interaction: NOD2 (NOD2), an important innate immunity component and gamma-secretase complex, responsible for cleavage of beta-amyloid precursor from plasma membranes, encoded by PSEN1, NCSTN, APH1A, APH1B, PSENEN37,38. Due to the interactions with so many molecules, CD147 plays a crucial role in energy metabolism of the cell, motility, recruitment and activation, although the final outcome of its stimulation depends on the cell type and co-expression of the other molecules. However, the co-expression of these molecules in different cell types and across various human tissues is not known.
Importantly, in T cells CD147 suppresses T-cell receptor-dependent activation mainly by inhibition of nuclear factor of activated T-cells (NFATs)39,40. The NFAT family consists of five proteins: NF-ATc1 (encoded by NFATC1), NF-ATc2 (encoded by NFATC2), NF-ATc3 (encoded by NFATC3), NF-ATc4 (encoded by NFATC4) and NF-AT5 (encoded by NFAT5)41,42. NFAT complex is a major transcriptional regulator in naive T cells and differentiated effector T cells, dependent on calcium/PLCγ/calmodulin/calcineurin signaling41,42. It is also crucial in regulation of T cell anergy and in differentiation and function of T regulatory (Treg) cells42. CD147 has been reported as a marker of human Treg cells with highly suppressive activity43. NFAT signaling is also important in other cell types such as DCs, mast cells and B cells41. Since cyclophilins are regulators of NFAT activation, the extracellular binding of the virus through cyclophilin-CD147 complex, as well as intracellular interactions of cyclophilins with the virus proteins might be important in COVID-19 development.
CD26 (encoded by DPP4), has emerged recently as a potential receptor for SARS-CoV-2, due to the fact that it is a main cellular entry for MERS-CoV44,45. Recent structural studies predict that SARS-CoV-2 spike protein directly interact with CD26 on the host cells22. CD26 is a cell surface glycoprotein involved in T-cell receptor-mediated T-cell activation and proliferation. It is highly expressed in CD4 and CD8 T cells, and in lower quantities also in NK cells and DCs. As an additional function, it acts as a serine exopeptidase, cleaving peptides of various chemokines, growth factors and peptide hormones. Interestingly, it is also involved in extracellular matrix cleavage. It has been shown in human cells that several polymorphisms in CD26 gene reduce entry of MERS into the host cells46. It remains to be functionally determined if this protein can be a functional receptor for SARS-CoV-2.
The aim of our study was to analyze the gene expression of ACE2, CD147, cyclophilins, CD26 and other SARS-CoV-2-related molecules in the broad range of primary human innate and adaptive immune cells and tissues, based on our own next generation sequencing data and public databases from different human cell types and across the diseases which are known to predispose to COVID-19.
Material and Methods
Study subjects, samples and study description
We analyzed gene expression of SARS-CoV-2 receptors and related molecules’ (Table S1) in a broad range of tissues and immune cells from the human RNA-seq databases generated by our ex vivo and in vitro approaches in the Swiss Institute of Asthma and Allergy Research (SIAF), by our collaborators or from the Gene Expression Omnibus (http://www.ncbi.nlm.nih.gov/). Baseline gene expression was evaluated in vitro in air liquid interface (ALI) - differentiated human primary bronchial epithelial cells (HBECs) (n=10). Direct ex vivo analyses of non-diseased human tissues included bronchial biopsies (n=5), bronchoalveolar lavage (BAL) cells (n=5) and skin biopsies (n=6). The other ex vivo investigated samples included whole blood (n=5), neutrophils (n=4), classical monocytes (n=4), plasmacytoid dendritic cells (pDCs) (n=4), innate lymphoid cells (ILC) 1, 2 and 3 (n=6 for each), natural killer cells (NK) (n=4), naïve CD4+ T cells (n=4), terminal effector CD4+ T cells (n=2), naïve CD8+ T cells (n=4), effector memory CD8+ T cells (n=4), naïve B cells (n=4) and plasmablasts (n=4) (for sorting markers please see Table S2).
We further investigated different expression patterns of ACE-2-, CD147- and CD26-related genes in the context of potential COVID-19 risk factors, namely age, gender, smoking status, diagnosis of asthma, chronic obstructive pulmonary disease (COPD), hypertension, obesity, atopic dermatitis (AD) (Table S3). Briefly, we performed in-depth curated analysis of HBECs (Control = 5, Asthma = 6, COPD =5), bronchial biopsies (Control = 16, Asthma = 22, COPD = 3, Non-obese = 20, Obese = 21, Normotension = 32, Hypertension = 9, Non-smoker = 19, Smoker = 21, Female = 14, Mal e= 27), BAL fluid (Control = 16, Asthma = 22, COPD = 2, Non-obese = 19 Obese = 21, Normotension = 31, Hypertension = 9, Non-smoker = 19, Smoker = 20, Female = 14, Male = 26), whole blood (Control = 17, Asthma = 21, COPD = 3, Non-obese = 20, Obese = 21, Normotension = 32, Hypertension = 9, Non-smoker = 19, Smoker = 21, Female = 14, Male = 27). In addition, we analyzed gene expression in PBMCs from infants and young children at age of 5-17 or 12-36 months (n=21 and 14, respectively), older children and adolescents at age of 4-16 years (n=16) and adults at age of 16-67 years (n=19), naïve CD4+ T cells from children at age 12 months (n=18) and adults at age 20-35 years (n=4) and skin biopsies (control: n=6, atopic dermatitis: non lesional sites n=11, lesional sites n=11). All studies were accompanied by the relevant ethical permissions, given by the appropriate Institutional Review Board. Each control and diseased subject gave informed consent. Total RNA from evaluated cells and tissues was extracted and transcriptome was analyzed with use of RNA-seq approaches.
Data processing
Signatures of ACE2-, CD147- and CD26-related genes were curated from GSEA and MSigDB Database (Broad Institute, Massachusetts Institute of Technology, and Regents of the University of California) and from literature. Full sets of analyzed genes are described in the Table S1. Genes of interest were extracted from all data sets. RNA-seq data were processed with inhouse workflow available at https://github.com/uzh/ezRun. Significance threshold for differentially expressed genes was set to p < 0.05 and was calculated for the entire gene lists in each project. All calculations between different conditions were done using the edgeR R package47. Spearman correlation coefficient was calculated using Hmisc R package, with the threshold for significance set to α = 0.05. Correlation plots were done using Python’s Seaborn library. Co-expression heatmaps as well as correlation heatmaps were done using the corrplot R package
Detailed description of each study is included in the Supplementary Methods and the Table S3.
Results
ACE2 and related molecules are exclusively expressed at the barrier sites, whereas CD147, cyclophilins, CD26 and their interaction partners are ubiquitously expressed in immune cells and in epithelium
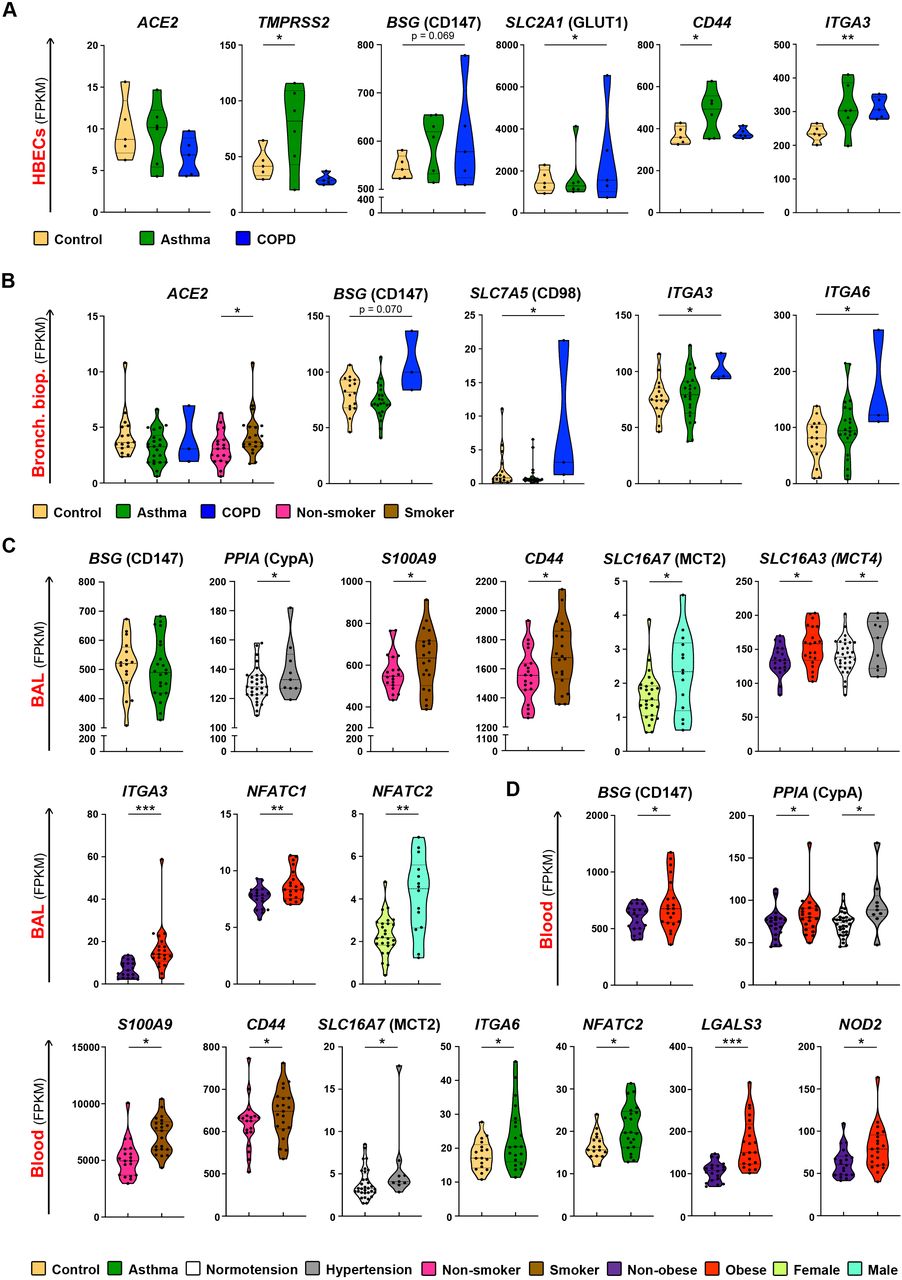
First, we analyzed the baseline expression and co-expression of ACE2-, CD147- and CD26-related genes in several immune cell types from control, non-diseased adult individuals. In agreement with other recent reports, we observed the expression of ACE2 mainly in epithelial tissues, such as human bronchial epithelial cells and bronchial and skin biopsies (Figure 1A and S1)6,48. ACE2 was more abundant in the lung, when compared with the skin barrier sides. In bronchial biopsies ACE2 was co-expressed with TMPRSS2 (Figure S2). Also, in the skin TMPRSS2 was very prominent, while we did not find SLC6A19 in any of the analyzed cells and tissues (Figure 1B and S1).
None of the analyzed immune cells expressed ACE2 (Figure 1A), TMPRSS2 (Figure 1B) or SLC6A19 (Figures S1-3), either in the local lung environment (BAL cells) or in the blood.

Expression of A) ACE2, B) TMPRSS2, C) BSG, D) PPIA, E) PPIB, F) S100A9, G) SLC16A7, H) SLC2A1, I) CD44, J) ITGB1, K) NFATC3, L) DPP4 genes in in vitro Air Liquid Interface (ALI) – differentiated human primary bronchial epithelial cells (n=10) and in ex vivo human primary bronchial biopsies (n=5), bronchoalveolar fluid cells (n=5), whole blood (n=5), neutrophils (n=4), classical monocytes (n=4), plasmocytoid dendritic cells (n=4), group 1, 2 and 3 innate lymphoid cells (n=6 per group), natural-killer cells (n=4), naïve CD4+ T cells (n=4), terminal effector CD4+ T cells (n=2), naïve CD8+ T cells (n=4), effector memory CD8+ T cells (n=4), naïve B cells (n=4), plasmablasts (n=4) and skin biopsies (n=6) from healthy adults. Data obtained from in vitro approaches are highlighted in red. Names of the proteins encoded by analysed genes are stated in the brackets. HBECS, human bronchial epithelial cells; Bronch. biop., bronchial biopsy; BAL, bronchoalveolar fluid cells; Class. monocytes, classical monocytes; pDCs, plasmocytoid dendritic cells; ILC1, group 1 innate lymphoid cells; ILC2, group 2 innate lymphoid cells, ILC3, group 3 innate lymphoid cells; NK, natural killer cells; naïve CD4+, naïve CD4+ T cells; term. Eff. CD4+, terminal effector CD4+ T cells; naïve CD8+, naïve CD8+ T cells, eff. mem. CD8+, effector memory CD8+ T cells.
In contrast to ACE2, we observed a prominent expression of CD147 (BSG) in epithelial and innate and adaptive immune cells in the lung (BAL), in the skin and in circulation (whole blood, neutrophils, classical monocytes, pDCs, ex vivo sorted ILC, NK, naïve CD4+ T cells, terminal effector CD4+ T cells, naïve CD8+ T cells, effector memory CD8+ T cells, naïve B cells and plasmablasts) (Figure 1C, Figures S1-S3). Interestingly, ACE2 and CD147 (BSG) are not co-expressed in bronchial biopsy (Figure S2). High expression of CD147 (BSG) in primary human bronchial epithelial cells suggests that these cells having more than one receptor for SARS-CoV-2 entry could be infected in various conditions. Granulocytes and macrophages are the main source of CD147 (BSG) expression49, which may explain high CD147 (BSG) expression in the whole blood and BAL observed in our data. Presence of CD147 (BSG) on innate and adaptive memory immune cells may indicate the importance of this receptor in systemic inflammatory response and development of immunological memory.
Since it is not clear yet how exactly the process of infection via CD147 (BSG) takes place in case of SARS-CoV-2, we examined the presence and co-expression of extracellular partners of CD147 (BSG), which are implicated in the infection with other viruses using CD147 (BSG) for cell entry, including SARS-CoV, HIV-1, measles and others11–13. All investigated cell populations showed high expression of cyclophilin A and B (PPIA and PPIB) (Figure 1D and E, Figure S1). Interestingly, cyclophilin A was highly expressed in all ILC. In addition, cyclophilin B was highly expressed in pDCS, ILC and plasmablasts. S100A9 was highly expressed not only in bronchial epithelium, but also in the whole blood, neutrophils, classical monocytes and, surprisingly, in naïve B cells (Figure 1F and Figure S1).
Next, we examined transmembrane and intracellular partners of CD147 (BSG), because its cytoplasmic tail, bound to many transmembrane partners, is essential in the entry of other viruses29. We observed that monocarboxylate transporters, such as SLC16A7 (MCT2) and SLC2A1 (GLUT1) are highly expressed not only at the barrier sites, but also in ILC (MCT2), pDCs, NK cells, T cells and B cells (MCT2, GLUT1) (Figure 1G and H). CD44 is highly expressed in majority of cells, with the exceptionally high expression in BAL, classical monocytes, naïve CD4+ T cells, terminal effector CD4+ T cells, naïve CD8+ T cells and effector memory CD8+ T cells (Figure 1I). ITGB1 (Integrin β-1) and other integrins are highly expressed in airway epithelial cells, but also other cells expressed them (Figure 1J and Figure S1). NF-ATc1-3 (NFATC1, NFATC2, NFATC3) were expressed predominantly in CD4+ and CD8+ T and naïve B cells, whereas expression of NFAT4 and NFAT5 was higher in the lung and skin (Figure 1K and Figure S1).
Finally, we looked at the expression of CD26 (DPP4). Notably, it was expressed highly in all ILC, and in naïve CD4+ and CD8+ T cells, as well as in pDCs and effector memory CD8+ T cells (Figure 1L and Figure S1).
Distinct expression profiles of ACE2-, CD147-, and CD26-related genes in PBMCs and T cells of children, adolescents and adults
Clinical evidence from COVID-19 patients worldwide clearly demonstrate an association between disease severity and morbidity with age1. In China, less than 1% of the SARS-CoV-2 positive cases were in children younger than 10 years of age50,51. Therefore, we analyzed expression patterns of ACE2-, CD147-, and CD26-related genes in peripheral blood mononuclear cells (PBMCs) from infants, young children, adolescents and adults. While these comparisons can be limited due to the different distribution of immune subsets in the blood of children and adults52 and different ethnicity of children (African American from South Africa and Tanzania) compared to adolescents and adults (Caucasians), we observed that there are different patterns of expression for the majority of analyzed genes (Figure 2A). We observed that CD147 (BSG) was highly expressed in PBMCs of children, whereas its expression was lower in adolescents and in adults. Similarly, to CD147 (BSG), cyclophilins B (PPIB), S100A9, SLC3A2 (CD98), SLC16A1 (MCT1), NFATC4 were less expressed in adults. On the other hand, SLC16A3 (MCT4), SLC16A7 (MCT2), NFATC2, NFAT5, NFATC1, NFATC3, integrins (ITGB1) were expressed on higher level in adults. Older children and adolescents showed intermediary gene expression between children and adults.

A) Expression of ACE2-, CD147-, NFAT- and CD-26-related genes in the primary human PBMCs in healthy children aged 5-17 months (n=21), 12-36 month (n=14), 4-16 years (n=16) and healthy adults aged 16-67 years (n=19). B) Expression of ACE2-, CD147-, NFAT- and CD-26-related genes in the primary human naïve CD4+ T cells from 12 months old healthy children (n=18) and 20-35 years old healthy adults (n=4). All heatmaps display normalized gene expression across the groups (rows normalization). Color-coding represents gene families related with ACE2 (orange), CD147 (green), NF-ATs (purple) and CD26 (yellow). MO, months old, YO, years old, PBMCs, peripheral blood mononuclear cells.
Next, we compared ACE2-, CD147- and CD26-related gene expression profile in the purified naïve CD4+ T cells between young children and adults (Figure 2B). Low expression of ACE2-related genes was observed in both children and adults. Several CD147-related genes showed higher expression in adults, including CD44, certain MCTs, SLC3A2 (CD98), SLC2A1 (GLUT1), NFATC1 and NFATC3. Also, CD26 (DPP4) was expressed at a higher level in adults. In contrast, other CD147-related genes were expressed at lower levels in adults, such as cyclophilins A and B (PPIA and PPIB), S100A9, SLC7A5 (CD98), LGALS3, integrins (ITGA6, ITGA3) and NFATC2.
Asthma, COPD, hypertension, smoking, obesity and gender show different expression profiles of ACE2-, CD147-, and CD26-related genes
We analyzed expression of ACE2-, CD147-and CD26-related genes from different cells and tissues in various comorbidities and risk factors, which have been shown, or are suspected to predispose to SARS-CoV-2 infection and/or COVID-19 progression. Upper and lower airways are the initial entry of SARS-CoV-2, thus we first analyzed gene expression in the HBECs and the bronchial biopsies from patients with asthma or COPD as compared to the non-diseased controls. In our cohorts, we did not see any significant difference in ACE2 expression in HBECs or bronchial biopsy between control, asthma and COPD patients (Figure 3A and B). However, in bronchial biopsies ACE2 expression was higher in smokers (Figure 3B). In HBECs, we observed higher expression of TMPRSS2 in asthma (Figure 3A), raising the possibility that in asthmatic airways the cleavage of spike protein of SARS-CoV-2 might be more efficient. We also observed a trend of increased expression of CD147 (BSG) in HBECs (Figure 3A) and in bronchial biopsies (Figure 3B) from COPD patients. Additionally, glucose transporter GLUT1 (SLC2A1), integrin α-3 (ITGA3) and galectin-3 (LGALS3) were higher expressed in HBECs, whereas SLC7A5 (CD98), integrins α-3 (ITGA3) and α-6 (ITGA6) were higher expressed in the bronchial biopsies of COPD patients. CD44 and APOD were higher expressed in asthma (Figure 3A, B and Figures S4, S5). In bronchial biopsies, ACE2-, CD147- and CD26-related genes showed similar cluster of co-expression in control and asthma (Figure S2 and S6). Interestingly, ACE2 co-expressed with PPIB and NME1 in asthma, which was not found in controls (Figure S6). Taken together, airway epithelium in asthma and COPD showed a gene signature that potentially can facilitate SARS-CoV-2 entry and enhance internalization after receptor binding.

A) Differential expression of ACE2, TMPRSS2, BSG, SLC2A1, CD44 and ITGA3 genes in in vitro Air Liquid Interface (ALI) – differentiated human primary bronchial epithelial cells from non-diseased controls (n=5), asthma (n=6) and COPD (n=5) patients. B) Differential expression of ACE2, BSG, SLC7A5, ITGA3, ITGA6 genes in bronchial biopsies from non-diseased controls (n=16), patients with asthma (n=22) and COPD (n=3), or in comparison of smokers (n=21) with non-smoking individuals (n=19). C) Differential expression of BSG, PPIA, S100A9, CD44, SLC16A7, SLC16A3, ITGA3, NFATC1, NFATC2 genes in the bronchoalveolar fluid (BAL) from the control individuals (n=16), patients with asthma (n=22) and COPD (n=2), or in comparison of hypertensive (n=9) with normotensive (n=31) individuals; smokers (n=20) with non-smokers (n=19); obese (n=21) with non-obese (n=19); and males (n=26) with females (n=14). D) Differential expression of BSG, PPIA, S100A9, CD44, SLC16A7, ITGA6, NFATC2, LGALS3 and NOD2 genes in the whole blood of non-diseased controls (n=17), patients with asthma (n=21) and COPD (n=3), or in comparison of hypertensive (n=9) with normotensive (n=32) individuals; smokers (n=21), with non-smokers (n=19); obese (n=21) with non-obese individuals (n=20); and males (n=27) with females (n=14). Names of the proteins encoded by analyzed genes are stated in the brackets. *p < 0.05, **p < 0.01, ***p < 0.001, ****p< 0.0001. HBECS, human bronchial epithelial cells; Bronch. biop., bronchial biopsy; BAL, bronchoalveolar fluid cells; COPD, chronic obstructive pulmonary disease.
Next, we analyzed BAL and whole blood gene expression in patients with asthma and controls (Figure 3 C, D and Figure S7, S8). Unfortunately, we have access to the limited number of BAL samples from COPD patients. In BAL, which reflects local lung immune microenvironment, and in blood, reflecting systemic immune responses, CD147 (BSG) was expressed equally high in asthma patients and in controls (Figure 3C, Figure S7). In blood, patients with asthma had also higher expression of integrin ITGA6 and NFATC2, as compared to controls (Figure 3D). There was a greater abundance in the cluster of co-expressed genes in asthma in BAL and especially in blood (Figure S9, S10). In asthma patients, BSG (CD147) showed co-expression with SLC7A5 (CD98) in blood, DPP4 (CD26) also co-expressed with CD44 and ITGA6 (Figure S10).
Next, we analyzed the ACE2-, CD147- and CD26-related gene expression in our controls, asthma and COPD patients according to the additional clinical features, which have been reported as a risk factor comorbidity for COVID-19, such as hypertension, smoking, gender and obesity23,53. We did not see any major differences in the gene expression in the bronchial biopsies based on these features, except higher ACE2 expression in smokers (Figure 3B, Figure S5). However, we noted several important differences in the BAL and in the whole blood (Figure 3 C, D and Figure S7, S8). Subjects with hypertension had increased expression of cyclophilin A in both the BAL and blood, as well as upregulated expression of MCT4 (SLC16A3), APH1A and PSENEN in the BAL and MCT2 (SLC16A7) in the blood (Figure 3 C, D and Figure S7, S8). In case of smoking, expression of S100A9 and CD44 was elevated in both BAL (Figure 3C, Figure S7) and blood (Figure 3D, Figure S8). Obesity was an important factor leading to the significant changes in the BAL and blood. We observed that MCT4 (SLC16A3), integrin ITGA3, NFATC1 and PSEN1 were more expressed in the BAL of obese individuals (Figure 3C, Figure S7), whereas CD147 (BSG), PPIA, LGALS3 and NOD2 were more expressed in their blood (Figure 3D). Finally, regarding gender in our cohort, MCT2 (SLC16A7), CD98 (SLC7A5), NFATC2 were higher expressed in the BAL of male subjects (Figure 3C, Figure S7).
Expression of CD147-related genes correlates with BMI and age in the BAL and blood
Since obesity and age were important variables, we additionally correlated these two variables with the expression of ACE2-, CD147- and CD26-related genes in the bronchial biopsy, BAL and blood of our adult non-diseased and diseased cohorts. We did not find any significant correlations between BMI and age and the receptor-related gene expression in the bronchial biopsies. However, we noted that the expression of SLC16A3 (MCT4), ITGA3, LGALS3 in BAL positively correlated with BMI (Figure 4A-C) and the expression of CD44 positively correlated with the age of the subjects (Figure 4D). Interestingly, whole blood expression of CD147 (BSG), PPIA, S100A9, CD44 and LGALS3 correlated positively with the BMI, whereas SLC16A3 positively correlated with age (Figure 4 E-J). In summary, these findings suggest that higher BMI and older age lead to higher expression of CD147-related genes on immune cells, but not on the barrier cells, which potentially can influence the development and the course of COVID-19.

Correlation of A) SLC16A3 expression and BMI, B) ITGA3 expression and BMI, C) LGALS3 expression and BMI, and D) CD44 expression and age in the bronchoalveolar fluid (BAL). Correlation of E) BSG expression and BMI, F) PPIA expression and BMI, G) S100A9 expression and BMI, H) CD44 expression and BMI, I) LGALS3 expression and BMI, J) SLC16A3 and age in the whole blood. Spearman correlation coefficient (r) was calculated, with the threshold of significance set to p = 0.05. Names of the proteins encoded by analyzed genes are stated in the brackets. BAL, bronchoalveolar fluid cells; BMI, body-mass index.
Eczema lesional skin in patients with atopic dermatitis present unique ACE2- and CD147-related gene expression profile
As we found a high expression of ACE2-, CD147- and CD26-related genes in the skin biopsies of healthy subjects, and since COVID-19-related skin lesions have been recently reported54, we also analyzed the biopsies of healthy and atopic dermatitis (AD) skin. Interestingly, in the lesional skin as compared to non-lesional skin of the same subjects, many CD147-related genes showed higher expression, including cyclophilins (PPIA and PPIB), S100A9, CD44, MCTs, SLC7A5 (CD98) and integrins (Figure 5).

A) Expression of ACE2, TMPRSS2, BSG, PPIA, PPIB, S100A9, CD44, SLC16A1, SLC16A3, SLC7A5, SLC3A2, SLC2A1, ITGA3, ITGA6, NFATC3, JUP, NME1, NOD2, SDC1 and DPP4 genes in the skin of healthy controls (n=6) and in the lesional (n=11), and non-lesional (n=11) skin of atopic dermatitis patients. Names of the proteins encoded by analyzed genes are stated in the margins. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001.
Discussion
COVID-19 pandemic is developing at such a pace that extraordinary actions are being initiated to learn quickly about its biology, transmission and potential means of prevention and treatment. In this spirit, we first performed an extensive literature and database search and curated a list of proven and potential receptors for SARS-CoV-2 and interaction partners, whose expression in different tissues and cells might be involved in the course of immunological response in COVID-19 (Figure 6)55. Next, we analyzed these genes in the broad spectrum of cell types and tissues in healthy controls to evaluate the level of their expression and their co-expression profiles, as well as evaluated their expression in healthy children, adolescents and adults. While interesting associations were observed to be age-dependent, these findings must be further explored and repeated due to the possibility of batch-specific systematic variations in the gene expression values between in-house and public datasets. Despite our efforts in applying identical data analysis workflow for the datasets produced by our group and those available in public repositories, different sequencing facilities and experiment protocols can lead to altered gene expression values, especially for genes with extreme gene length and G/C content. Finally, we analyzed gene expression in adults with known and potential COVID-19 comorbidities and risk factors such as COPD, asthma, hypertension, obesity, smoking, male gender and AD. These conditions can be directly compared, as they were performed internally in the context of same project.
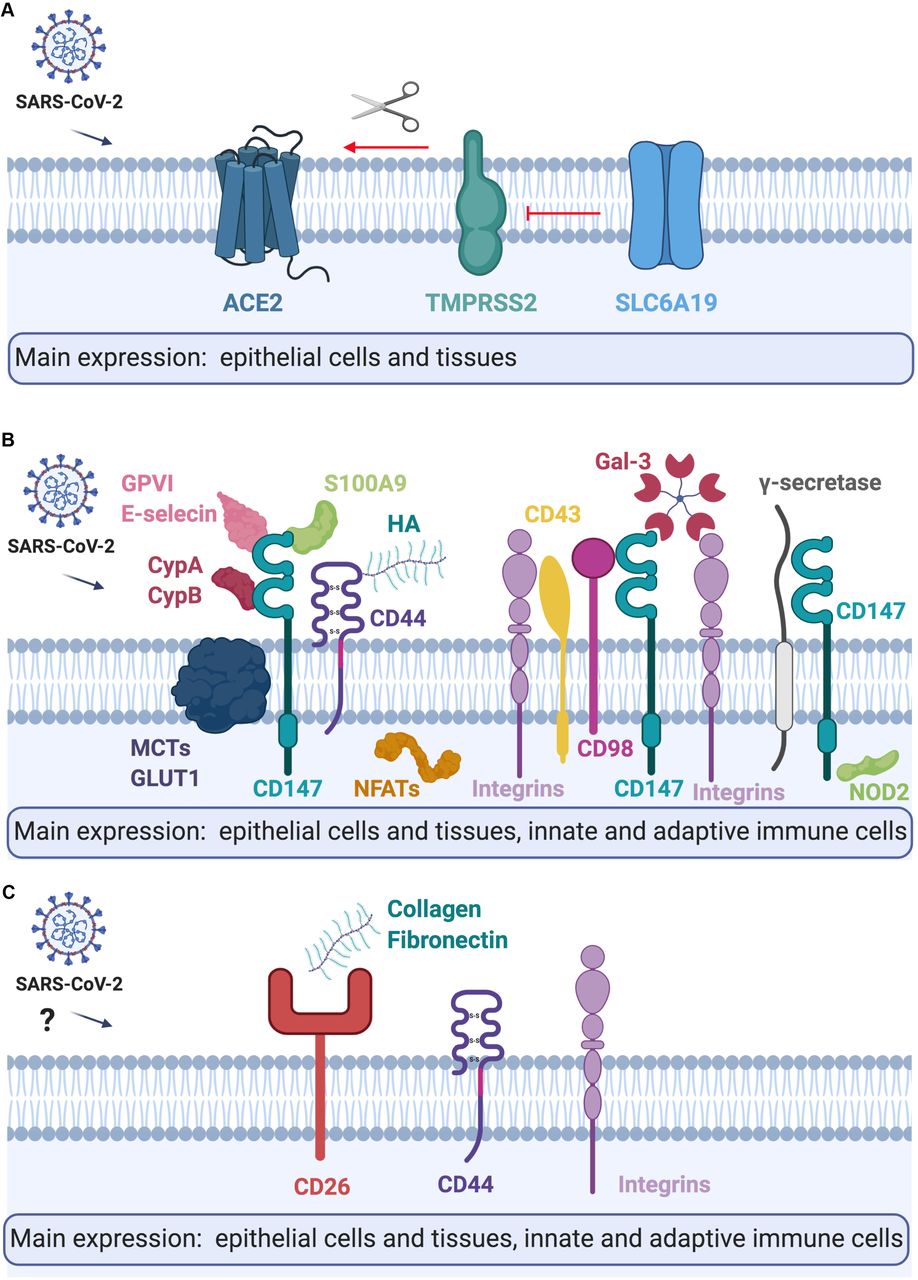
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Summary of the tissue and cellular expression, and models of A) ACE2, B) CD147, C) CD26 and their interaction partners. Please refer to the text for further details. CypA, Cyclophilin A; CypB, Cyclophilin B; HA, Hyaluronic acid; Gal-3, Galectin 3; MCTs, Monocarboxylate transporters. Figure created with BioRender.com.
ACE2 is a receptor for SARS-CoV-23, whereas TMPRSS2 is a transmembrane host protease, which cleaves the viral spike protein thus facilitating virus fusion to the cellular membranes process (S)3,56. SLC6A19, physiologically a neutral amino acid transporter, potentially can block the access of TMPRSS2 to ACE2 and subsequently reduce active infection4,6. Our results, in agreement with recent reports6,57,58, indicate that airway epithelium has high ACE2 and TMPRSS2 co-expression, and no expression of protective SLC6A19 and thus might be highly susceptible for SARS-CoV-2 infection. In addition, it has been shown recently that ACE2 is highly expressed in naso- and oropharynx, which are the sites of active SARS-CoV-2 replication and a main source of infectious particles6,57,58. Interestingly, we observed high expression of ACE2 and TMPRSS2 in bronchial airway epithelium: in HBECs ALI cultures in vitro, as well as in vivo in humans in bronchial biopsies, indicating that they can be as important in initiation and progression of COVID-19, as recently shown for type II pneumocytes58. We have not observed the expression of SLC6A19 in any of the analyzed cells and tissues, yet it is known to be expressed is intestine59. Interestingly, in contrast to Leung and colleagues60, we have not seen higher expression of ACE2 in HBECs or bronchial biopsy of patients with COPD, but our sample size was very limited, and potentially a bronchial biopsy site was different, which might be the reason of this inconsistency. However, consistently with this report61, we observed higher expression of ACE2 in the bronchial biopsy of current smokers, which might indicate that current smoking status, might be a stronger factor of increased ACE2 expression in airway epithelium than COPD per se. On the other hand, ACE2 and the angiotensin II type 2 receptor (AT2) were reported to protect from severe acute lung injury in mice62, therefore an importance of ACE2 in SARS-CoV-2 infection and COVID-19 progression should be further explored. In addition, in HBECs of patients with asthma, we observed higher expression of TMPRSS2, which increases the possibility of SARS-CoV-2 cleavage in asthmatic bronchi. Even though initially asthma was not reported to be a significant comorbidity for COVID-19, more observations from Europe and the US seem to show otherwise63. It might be related to the heterogeneity of asthma endotypes type 2 asthma (allergic) and non-type 2 (non-allergic). Allergen exposure, allergic sensitization and high IgE lead to lower ACE2 expression in the nasal and bronchial epithelium of asthma patients64. Thus, it is possible that ACE2 expression in the airways of allergic patients, even if slightly protective in terms of infection with SARS-CoV-2, might also predispose to faster progression of COVID-19 to more severe forms, especially in case of higher TMPRSS2 expression.
We found that immune cells do not express ACE2, TMPRSS2 or SLC6A19, which has also been observed by others2,5 Importantly though, we found that CD147 and its extracellular agonists and transmembrane partners are highly expressed in innate and adaptive immune cells in the lungs and in the periphery, suggesting that they should be further investigated in SARS-CoV-2 spread and COVID-19 pathology9. Here, we report high expression of CD147 in both, epithelial tissues and innate and adaptive immune cells. Potentially, epithelial cells, macrophages, monocytes, ILCs, NK cells, T cells and B cells can be infected in the lungs or can carry SARS-CoV-2 from infected epithelial cells via CD147 and participate in the local and systemic spread of the virus, and in exaggerated immune response65.
We found here that CD147 is slightly higher expressed in the HBECs and bronchial biopsy of COPD patients, as well as it is higher expressed in the blood of obese individuals. Obesity is one of the main comorbidities reported in patients with severe COVID-1963, and since CD147 expression in the whole blood correlates positively with BMI in our cohort, it certainly needs further attention in relation to COVID-19. Importantly, CD147 expression is upregulated by high glucose concentrations66, which might reflect its correlation with obesity, and potentially also with diabetes, another very important COVID-19 comorbidity. Additionally, extracellular ligands and transmembrane partners of CD147 participate in the progress of other viral infections12 in variety of mechanisms. Viruses integrate cyclophilin A and cyclophilin B in their virions and further can bind and either infect cells via 147 or activate CD147 signaling11–13. We found that expression of both cyclophilins (PPIA and PPIB) is increased in subjects with hypertension and obesity and PPIA correlated positively with BMI in the whole blood, which is confirmed in other studies67. In addition, nsp1 protein of SARS-CoV binds to CD147 via cyclophilins and reduces interferon responses25 in infected cells. It also inhibits NF-ATs translocation activation in T cells, leading to suppression of immune responses25,68. Both mechanisms might be more potent in obese patients, especially with hypertension, due to higher expression of both cyclophilins on the periphery. Also, the other extracellular ligand of CD147-S100A9 correlated positively with BMI. CD147 transmembrane partners such as LGALS3, MCT4 (SLC16A3), MCT2 (SLC16A7), ITGA3 and NFATC1 were elevated in the blood or BAL of obese or hypertensive individuals, as well as the expression of LGALS3, S100A9 and CD44 correlated positively with BMI. Importantly in individuals who are current smokers S100A9 and CD44 are elevated in the BAL and in the blood. MCTs regulate the transport of lactate, pyruvate and ketone bodies across the cell membrane69. Galectin-3 (LGALS3) via CD147 signaling is responsible for disrupting cell-cell contact at the epithelial barriers70. CD44 in acute inflammation triggered by hyaluronic acid lead to the cell activation and release of many proinflammatory cytokines whereas in the longer terms induce lung fibrosis71,72. Interestingly, it has been shown that patients who recovered from SARS-CoV have spike protein-specific memory cytotoxic T cells, which express CD4473. Thus, our results suggest a possible role of CD147 and its extracellular and intracellular ligands in COVID-19 pathogenesis via regulation of cell metabolism, motility and activation, especially in patients with comorbidities. A clinical trial with anti-CD147 in COVID-19 patients seems to confirm its pleiotropic role in SARS-CoV-2 infection, but it requires further research to investigate the involved mechanisms and molecules.
Finally, expression of ACE2/TMPRSS2, CD147- and CD26-related genes in the skin biopsies highlights the potential roles of these molecules in various viral diseases. Recent studies reported that many COVID-19 cases developed nonpruritic, erythematous rashes, urticaria or varicella-like lesions54,74, but so far it is not known if they are a place of viral replication or just a local reaction to systemic infection. Similarly, not much is yet known about the potential connections between AD and the course of COVID-1975. Yet, severe and untreated AD is a known risk factor for disseminated viral skin disease76. Therefore, our observations indicating higher expression of cyclophilins, CD44, S100A9, integrins, MCTs and other CD147-related genes require further studies about their role in the course of SARS-CoV-2 infection in healthy subjects and in AD patients.
CD26 (DPP4) is another receptor important in coronavirus infection, described in MERS-CoV, potentially recognizing SARS-CoV-221,77. Similar to CD147, CD26 was expressed in all investigated cell types, except B cells. ILCs showed highest expression of DPP4 from all analyzed cells, which is in agreement with recent findings78. However, not much is known about the variable expression of CD26 among ILC1, ILC2 and ILC3. Potentially, a high level of CD26 on the surface of ILCs in the lung may be another mechanism of systemic spread of the virus.
The differential profile of ACE2-, CD147- and CD26-related gene expression in healthy subjects of different ages need to be interpreted with caution due to the distinct origins of these populations, as well as potential experimental bias (even if we performed normalization strategies, similar bioinformatic pipelines and cautious assessment of raw data quality, prior to analysis). Nevertheless, our observations are largely in agreement with other recent findings correlating gene expression in the whole blood and PBMCs with age79. Having this in mind, it is intriguing to observe that expression of NOD2, some MCTs, integrins and NFATs seemed to increase with age, whereas the expression of genes coding γ-secretase complex is decreasing with age in PBMCs. CD44 and MCT4 also correlated with age in BAL and blood, respectively. Moreover, some of the differences between children and adults, such as higher expression of CD26, CD44, GLUT1 (SLC2A1) and specific NFATs in adults were also observed in naïve purified CD4+ T cells. This requires further investigation, but differences in expression pattern may be related to the striking differences in the morbidity of SARS-CoV-2 between children and adults80.
References
- 1.↵
- 2.↵
- 3.↵
- 4.↵
- 5.↵
- 6.↵
- 7.↵
- 8.↵
- 9.↵
- 10.↵
- 11.↵
- 12.↵
- 13.↵
- 14.↵
- 15.↵
- 16.↵
- 17.↵
- 18.
- 19.
- 20.↵
- 21.↵
- 22.↵
- 23.↵
- 24.↵
- 25.↵
- 26.↵
- 27.↵
- 28.↵
- 29.↵
- 30.↵
- 31.
- 32.↵
- 33.↵
- 34.
- 35.
- 36.↵
- 37.↵
- 38.↵
- 39.↵
- 40.↵
- 41.↵
- 42.↵
- 43.↵
- 44.↵
- 45.↵
- 46.↵
- 47.↵
- 48.↵
- 49.↵
- 50.↵
- 51.↵
- 52.↵
- 53.↵
- 54.↵
- 55.↵
- 56.↵
- 57.↵
- 58.↵
- 59.↵
- 60.↵
- 61.↵
- 62.↵
- 63.↵
- 64.↵
- 65.↵
- 66.↵
- 67.↵
- 68.↵
- 69.↵
- 70.↵
- 71.↵
- 72.↵
- 73.↵
- 74.↵
- 75.↵
- 76.↵
- 77.↵
- 78.↵
- 79.↵
- 80.↵




