

Abstract
Natural plant populations encounter strong pathogen pressure and defense-associated genes are known to be under different selection pressure dependent on the pressure by the pathogens. Here we use wild tomato Solanum chilense populations to investigate natural resistance against Cladosporium fulvum, a well-known pathogenic fungus of domesticated tomatoes. We show that populations of S. chilense differ in resistance against the pathogen. Next, we explored the underlying molecular processes in a species wide-context. Then, focusing on recognition of the two prominent avirulence factors secreted by C. fulvum (Avr4 and Avr9) in central and northern populations of S. chilense we observed high complexity in the cognate homologues of Cladosporium resistance (Hcr9) locus underlying the recognition of these effectors. Presence of canonical genomic regions coding for Cf-4 and Cf-9, two major dominant resistance genes in the Hcr9 locus recognizing Avr4 and Avr9, respectively, does not meet prediction from Avr response phenotypes. We find both genes in varying fractions of the plant populations and we show possible co-existence of two functionally active resistance genes, previously thought to be allelic. Additionally, we observed the complete local absence of recognition of additional Avr proteins of C. fulvum. In the southern populations we attribute this to changes in the coregulatory network. As a result of loss of pathogen pressure or adaptation to extreme climatic conditions. This may ultimately explain the observed pathogen susceptibility in the southern populations. This work puts major gene mediated disease resistance in an ecological context.
- Solanum chilense
- Cladosporium fulvum
- receptor-like proteins (RLPs)
- Cf-9
- tomato
- resistance
Introduction
How plants and their pathogens adapt to one another in their natural habitat is still poorly understood. Some studies highlight the local adaptations in context to plant-pathogen interactions: the wild flax-flax rust pathosystem is such an example, where more resistant wild flax harbored more virulent strains of the rust (Thrall et al., 2002; Thrall & Burdon, 2003). Similar mechanisms of the co-occurrence of virulent strains of the powdery mildew Podosphaera plantaginis and more resistant plants of Plantago lanceolata have also been documented (Laine, 2005; Soubeyrand et al., 2009). Furthermore, complex multi-host and multi- pathogen systems, with clear differences at a regional scale, have been observed in anther smut- fungi-infecting Dianthus hosts in the southern European Alps (Petit et al., 2017). The molecular mechanisms that is thought to drive these interactions is a gene-for-gene interaction (Flor, 1971), where single gene encoded pathogen molecules, now referred to as effectors, are recognized by specific receptors present in resistant host plants, also known as major resistance genes. The need to recognize an invading pathogen and subsequently evade the recognition by the plant and pathogen, respectively, leads to a constant co-evolutionary arms race, often described as the Red Queen dynamics (Van Valen, 1973). These dynamics are thought to have led to the plethora of different defense related genes in plants as we know it today. Wild crop relatives have been used as a gene pool for isolating and introducing genetic resistance against many different fungal pathogens. Genetic diversity of natural populations against microbes have been explored by sequence analysis or by experimental biology often still as independent, though complementary, approaches (Salvaudon et al., 2008), leading to a huge gap in our understanding of local adaptations and their role in shaping the current diversity of such resistance genes in an ecological context.
Tomato leaf mold is caused by the non-obligate biotrophic fungus Cladosporium fulvum syn. Passalora fulva. At the time of infection, C. fulvum enters through stomata into the leaf and colonizes the apoplastic spaces (Stergiopoulos & de Wit, 2009). The fungus secretes various small proteins, also referred to as effectors or avirulence factors (Avrs) when being recognized by a resistant host, into the apoplast with the aim to manipulate the host for its successful colonization of the host tissue. The infection leads to severe yellowing and wilting of the leaves, which ultimately leads to a loss of photosynthetic capacity and thus a loss in yield or reduced reproducible fitness. C. fulvum is a globally occurring pathogen with a clear genetic diversity (Iida et al., 2015). It is thought to originate in the Andean region, where it likely has co-evolved with one or several of wild tomato species that inhabit a mountain range from central Ecuador to northern Peru (de Wit et al., 2012). During evolution, as a result of selection pressure imposed by virulent strains of C. fulvum, several of the wild tomato species have evolved resistance genes whose products mediate recognition of the Avrs secreted by C. fulvum (Joosten & de Wit, 1999). This recognition facilitates host resistance following the gene-for-gene model. Resistance is eventually achieved when Avr-activated defense leads to a hypersensitive response (HR), which includes programmed cell death (PCD). This localized PCD is associated with various additional defense responses such as a massive callose deposition, and prevents C. fulvum from obtaining nutrients from the host, thereby limiting further pathogen ingress and multiplication (Lazarovits & Higgins, 1976).
To date, 24 C. fulvum (Cf) resistance genes have been mapped (Kanwar et al., 1980). Due to the complexity of the Cf genes and their loci, only a small number of Cf genes have been cloned and verified for their functionality. Cf genes are highly repetitive in their leucine-rich repeat encoding parts and functional homologs are often accompanied by non-functional ones, with very little sequence variation between them (Kruijt 2005). Cf-9 from S. pimpinellifolium encodes a cell surface receptor-like protein (RLP) and was the first Cf resistance gene to be cloned (Jones et al., 1994). The Cf-9 gene product recognizes the Avr9 protein of C. fulvum, which is a highly stable, cysteine-knotted peptide of unknown function (Scholtens-Toma & de Wit 1988). The Cf-9 gene belongs to the Hcr9 (homologs of C. fulvum resistance gene Cf-9) gene cluster, which is located on chromosome 1. In addition to Cf-9, Cf-4 is another well-studied Cf gene from the Hcr9 cluster. The Cf-4 gene product recognizes Avr4 of C. fulvum, which is a chitin-binding protein having eight cysteine residues (Joosten et al., 1994; van den Burg et al., 2004), and the Cf-4 gene originated from S. habrochaites (Thomas et al., 1997). Studies on MM-Cf9 which is an Avr9- recognising introgression line of the domesticated tomato, S. lycopersicum cv Money Maker, revealed presence of five Cf-9 homologs, Hcr9-9A to Hcr9-9E, mapped at the short arm of chromosome 1, with Hcr9-9C being the functional Cf-9 gene (Parniske et al., 1997). Similarly, Hcr9-4A to Hcr9-4E is present in the Avr4-recognising MM-Cf4 and Hcr9-4D corresponds to Cf-4 (Thomas et al., 1997). Cf-4 and Cf-9 lie at the same locus on chromosome 1 and are assumed to be mutually exclusive or even allelic. Crossings between recombinant inbred lines carrying Cf-4 and Cf-9 resulted in extreme genetic instability in the offspring (Parniske et al., 1997, Thomas et al., 1997).
Appreciating the important roles of these Cf genes and their assumed role in co-evolution between wild tomato and native C. fulvum, it is surprising that only a few studies have sought to investigate Cf gene diversity. An effectoromics approach was exploited by Mesarich et al. (2017), to identify plants carrying novel Cf genes to be potentially used in plant breeding programs in the future. Studies have shown that in S. pimpinellifolium several putative homologs are present, but their function remains unknown (Caicedo et al., 2004; Caicedo, 2008). Another study identified four variants of Cf-9 (originally isolated from S. pimpinellifolium), from its close relative S. habrochaites and each one variant of Cf-4 from S. habrochaites, S. chilense, S. chmielewskii, S. neorickii and S. arcanum (Kruijt et al. 2005). In-spite of multiple single nucleotide polymorphisms (SNPs) being present in the isolated Cf gene variants, all of them showed the ability to induce an HR after recognition of Avr9 and Avr4, respectively, which led the authors to conclude that Avr4 and Avr9 recognition is conserved among wild tomato. However, a species-wide analysis of different accessions of S. pimpinellifolium revealed intragenic recombination to have occurred between Hcr9-9D and Hcr9-9C/Cf-9. The resulting allele, 9DC, does not co-exist with the original Cf-9 allele in the individual plants. Variant 9DC is the more common allele in the species and the product also recognizes Avr9 (Van der Hoorn et al., 2001), indicating that one likely cannot speak of conservation of Cf alleles sensu stricto. In addition, plants that recognize both Avr4 and Avr9 have not been identified. Detailed knowledge on the relationship between Cf-4 and Cf-9 in other accessions or wild populations of tomato, and on their actual roles in resistance is still lacking.
To perform more detailed studies on resistance provided by Cf genes in an ecological context, we selected S. chilense, one of seventeen wild tomato species, as it covers a wide variety of habitats on the western slopes of the Andes, ranging from Peru to Northern Chile (Nakazato et al., 2010). The species range spreads from the edges of the Atacama Desert, as the southern edge of the range, to relatively wet, high altitude regions (up to 3500 meter above sea level), as well as in very specific coastal regions that experience regular sea fog (resulting in a relatively high humidity) in the most northern part of Chile, as well as in south Peru (Cereceda & Schemenauer 1991; Peralta et al., 2008; Chetelat et al., 2009). This highly varied distribution results in sub- populations of this species that encounter different environmental challenges. Ultimately, these different habitats lead to genetic differentiation within the species. The S. chilense population can be clustered in four groups: north, central, southern highlands and southern lowlands. The southern highlands and southern lowlands populations are derived from the central group (Böndelet al., 2015). This divergence was confirmed by whole genome sequencing, and multiple sequential markovian coalescent simulations revealed that migrations happened from the central group southward 50,000 to 200,000 years ago (Stam et al., 2019b). The strong differentiation between habitats leads to clearly observable adaptations. Southern populations respond faster to drought (Fischer et al., 2013) whereas high altitude populations are more cold-tolerant (Nosenko et al., 2016). In addition to adaptations to abiotic factors, these habitats are expected to be home to different biotic stressors at different intensity levels, including various pathogen species, which is anticipated to result in genetic variation in pathogen defense-associated genes. Indeed, differences in resistance properties of the various S. chilense populations against three pathogens, Alternaria solani, Phytophthora infestans and a Fusarium spp, are observed (Stam et al., 2017). Moreover, large genetic variation has been observed within S. chilense populations in another resistance-associated gene family; the nucleotide-binding leucine-rich repeat (NLR) resistance genes. These genes show clear presence-absence-variation when compared to related tomato species (Stam et al., 2019a), and an in-depth resequencing study, covering the whole species range of S. chilense, shows evidence that the selection pressure imposed on the individual NLRs differs for each of the populations (Stam et al., 2019b).
Little data exists showing biological interactions between C. fulvum and wild tomato species. In this study we investigated the interaction of the fungal pathogen C. fulvum with the wild tomato species S. chilense throughout the geographical range of the species. We show that S. chilense plants from different locations first of all show differences in their resistance to the fungus, including complete loss of resistance in some populations. By investigating the well characterized genes of the Hcr9 locus, we furthermore place major gene mediated immunity in an ecological context.
Materials and methods
Plants and fungal material
Seeds of the fifteen accessions (populations) of S. chilense used in our studies were originally obtained from the C. M. Rick Tomato Genetics Resource Center of the University of California, Davis (TGRC UC-Davis) (http://tgrc.ucdavis.edu/). The selected accessions were LA1958, LA1963, LA2747, LA2931, LA2932, LA2750, LA2959, LA3111, LA3784, LA3786, LA4107, LA4117A, LA4118 and LA4330. Each Accession number represents a random collection of seeds from a wild population and is propagated by TGRC in a way to maintain maximum genetic diversity. Böndel et al (2015) have shown that this seed multiplication has negligible effect on the genetic diversity of the accessions. Hence, each accession can be considered to truly represent the diversity in the wild plant populations. Introgression lines S. lycopersicum cv Moneymaker (MM) of Cf-9, Cf-4 and Cf-5 were generated by Tigchelaar et al. (1984) and S. pimpinellifolium (LP12), in which the 9DC was identified, and which was used as a control in all our assays were provided by TGRC UC-Davis. Per S. chilense accession 8-17 plants were grown. Plants were grown in a controlled glasshouse, with a minimum daytime temperature of 24°C and 16 hours of light and 8 hours of dark conditions. Adult plants were cut back at bi- weekly intervals to assure the presence of mature, fully developed, yet not senescent, leaves during each repetition of the experiment.
A race 5 strain of C. fulvum was maintained on 1/2 potato dextrose agar (PDA) medium and incubated at 16°C in the dark.
Visualization of infection phenotype and quantification of C. fulvum biomass from tomato leaves
Spray inoculation of C. fulvum at a concentration 20,000 conidia/ml was performed on 3-week old plant cuttings of population LA3111, LA4330, MM-Cf-9 and MM-Cf-5 plants; water- inoculated plants served as a negative control in the experiment. Plants were maintained at 24°C and 16 hours of light and 8 hours of dark, with 100% humidity for the first two days, after which 80% humidity was maintained throughout the experiment.
Photographs of leaflets of inoculated plants were collected at 14 days post inoculation (dpi) and Microscopy was performed from 7dpi to 19dpi on bleached leaves following staining in acetic acid (25%) 1:9 + ink (K□nigsblau, Pelikan, 4001) then washed in water and analysed under brightfield microscope (Zeiss, Imager Z1m).
For quantification, leaflets of inoculated plants were collected at 14 days post inoculation (dpi) and DNA was isolated using the protocol published by Yan et al. (2008). The infection loads or approximate amounts of C. fulvum DNA present in inoculated leaves were quantified by qPCR using the DNA-binding fluorophore Maxima SYBR Green Master Mix (2X) with ROX solution (Thermo Scientific). All qPCR reactions were performed on an AriaMx Real-Time PCR system (Agilent Technologies, Waldbronn, Germany). PCR were performed with the primer pairs RS158 (5’-GTCTCCGGCTGAGCAGTT-3’)/ ITS4 (5’-TCCTCCGCTTATTGATATGC-3’) and RS001 (5’-GCCTACCATGAGCAGCTTTC-3’)/ RS002 (5’-CAATGCGTGAGAAGACCTCA- 3’), annealing to the ITS region and elongation factor 1 alpha, respectively, present in the C. fulvum DNA. Templates used for this experiment were genomic DNA isolated from the leaves inoculated with the pathogen and water. Amplifications using both primer pairs for each sample were done on the same plate and in the run. The experiment was performed on three plants of MM-Cf-9 and MM-Cf-5 each and five plants of LA3111 and LA4330 each. All the plants samples well evaluated in three technical replicates and Cq differences higher than one among technical replicates was not considered in the final evaluation. The Cq values and the linear equations from the sensitivity graphs were used to calculate the quantities of pathogen and plant DNA in each sample (Figure S1).
Apoplastic washing fluid (AF), Avr9 and Avr4 infiltration assay
AF containing the complete set of C. fulvum Avrs, except for Avr5, was obtained by isolating AF from leaves of MM-Cf-5 plants colonized by race 5 of C. fulvum, at 10 to 14 days after inoculation. Furthermore, a preparation of Avr9 concentrated from AF by acetone precipitation, leaving the Avr9 peptide in the supernatant, and Avr4 produced in the yeast Pichia pastoris, were employed.
Using a 1 ml syringe (Braun, Omnifix) without a needle, the AF, Avr9 and Avr4 preparations were infiltrated from the lower side of fully expanded leaves of the different populations. Experiments were performed in three independent biological replicates. Infiltrations were done in fully expanded leaves of the same adult plant, with two technical replicates per plant. Readings were performed between 2 to7 days post infiltration. For Avr9 infiltration, an HR was typically observed within 1-4 days, whereas the AF-and Avr4-triggered HR were observed later. Readings were not performed later than 7 days after infiltration.
Presence of Cf-9-, 9DC-and Cf-4-specific regions in different S. chilense populations
Screening of S. chilense populations for the presence of gene-specific regions of Cf-9, 9DC and Cf-4 was performed through PCR amplification using the gene-specific primers CS5-CS1 and DS1-CS1 for Cf-9 and 9DC, respectively (Van der Hoorn et al., 2001). Primer pair PSK047 (5’- ACGACAGAAGAACTC-3’)/ PSK050 (5’-GATGGAATTGGTCCTT-3’), was designed to amplify the canonical Cf-4 domain (Fig. S2). DNA was isolated using standard CTAB extraction, using the same samples as used in a previous study (Stam et al., 2016; Stam et al., 2019b). PCR on gDNA was performed using Promega Green GoTaq® Polymerase and the products were analyzed by 1% agarose gel electrophoresis. As a PCR control elongation factor 1 alpha, amplified with the primer pair 5’-GTCCCCATCTCTGGTTTTGA-3’/ 5’- GGGTCATCTTTGGAGTTGGA-3’, was included and MM-Cf-9 and MM-Cf-4, and LP12 served as positive and negative controls, respectively.
Semi-quantitative evaluation of the expression of a Cf-9 homolog in the southern population LA4330
We evaluated the transcript levels of the Cf-9 homolog in LA4330 at eight hours after Avr9 or water infiltration. RNA from the infiltrated leaves was extracted using the RNeasy plant mini kit (Qiagen) and cDNA was synthesized using the QuantiTect reverse transcription kit (Qiagen). Amplification of Cf-9 the canonical region of the Cf-9 homolog was performed with primer pair binding on the start and end of the ORF, PSK009 (5’-ATGGATTGTGTAAAACTTGTATTCCT -3’) /PSK010 (5’-CTAATATCTTTTCTTGTGCTTTTTCA -3’), and the product was visualized on 1% agarose gel.
Whole genome sequence analyses for Cf co-receptors in S. chilense
We extracted the sequences for four important components of the Cf protein signaling complex, for which we chose SUPPRESSOR OF BIR1 (SORBIR1), SOMATIC EMBRYOGENESIS RECEPTOR-KINASE 3A (SERK3a) and AVR9/CF-9-INDUCED KINASE 1 (ACIK1) (Wu, 2020), as well as for an additional potential co-receptor of the BAK1 signaling complex, BIR2 (Halter et al., 2014) from NCBI. To identify the genomic sequences encoding these co-receptors and regulators of Cf proteins in S. chilense, we performed a BLAST search against the S. chilense reference genome (Stam et al., 2019b). Each SOBIR1, SEKR3a and ACIK1 yielded one unequivocal best target sequence in the reference genome and visual inspection after alignment confirmed that the S. chilense homologs are in all cases similar to the reference gene of S. lycopersicum. BIR2 yielded two good hits, of which we dubbed the second one BIR2b. In order to look for polymorphisms in three plants, one from LA3111, and one each from southern populations LA2932 and LA4330 (Stam, et al. 2019a), we obtained the genomic coordinates of the genes of interest from our reference genome and called the SNPs in the region of interest using samtools mpileup and bcftools call (-mv -Oz) (Li et al., 2009; Li, 2011). We removed low quality indels and the resulting vcf files were tabulated using tabix, to allow the consensus sequences for the genomic regions to be extracted using bcftools consensus. Finally, we used gffread from gffutils (Pertea and Pertea 2020) to obtain the correct coding sequence for each of the plants. All multiple sequence alignments were made and inspected using aliview (Larsson, 2014).
Statistical testing
ANOVA, with post hoc Tukey honest significant difference test was performed using R command Tukey HSD (anova) to test the statistical significance in pathogen load detected in the inoculated plants via qPCR.
Results
S. chilense populations show differences in resistance against C. fulvum
To confirm that C. fulvum is capable of successfully infecting S. chilense and to test whether S. chilense populations show differences in their resistance spectrum to C. fulvum, we spray- inoculated randomly selected individual plants of S. chilense from geographically distinct populations LA3111 (central) and LA4330 (southern highlands), with a conidial suspension of C. fulvum race 5. Upon visual inspection and microscopic evaluation, we found that plants of population LA3111 show similar phenotypes as our resistant control plant MM-Cf-9, a recombinant inbred line of S. lycopersicum cv. MoneyMaker with introgressed Cf-9, thus being resistant to C. fulvum race 5 as this race does produce Avr9. By contrast, LA4330 shows a phenotype similar to our susceptible control MM-Cf-5, a Cf-5 introgression line of S. lycopersicum of which resistance is circumvented due to a loss of the Avr5 gene by race 5 (Mesarich et al., 2014) (Figure 1a-b).

Tomato leaves inoculated with 2×104 conidia/ml. a) Left to right: inoculated leaves harvested at 14 dpi of MM-Cf-9, MM-Cf-5 and S. chilense population LA3111 and LA4330. b) Microscopic pictures of bleached leaves (scale bar = 20µm), after staining with ink (mycelium of pathogen indicated with red arrows). c) Quantification of C. fulvum DNA load in pg/ng of plant DNA after 14 dpi of inoculation in 3 plants per MM control and 5 plants per S. chilense population. All the plants were evaluated in three technical replicates and each data point shows pathogen load per technical replicate.
Next we performed quantification of C. fulvum DNA load and found significant differences in pathogen DNA contents between the tested plants (p < 0.001, ANOVA). Differences can be seen between the resistant and susceptible control plants (p = 0.05, TukeyHSD) and LA4330 showed a significantly higher (p < 0.001, TukeyHSD) presence of C. fulvum DNA than LA3111 (Figure 1c). For this we tested five individual plants from each S. chilense population and three plants per control. Taken together these findings show that C. fulvum is able to infect S. chilense plants and that the various populations behave differently when exposed to C. fulvum.
Southern S. chilense populations do not recognize C. fulvum effectors
Visible HR upon effector infiltration is an efficient and reliable proxy to test for the resistance properties of tomato plants on a large scale. To test the geographical variation in C. fulvum race 5 resistance, we performed an infiltration assay with apoplastic fluid (AF) of infected susceptible tomato plants, which is sufficient to trigger an HR in plants carrying matching Cf genes. As for the inoculation a race 5 was used, the AF is anticipated to contain all secreted C. fulvum effectors, except for Avr5. We infiltrated the AF at two sites in the leaves of 155 individuals, representing 15 different populations (8-17 individuals for each population). Interestingly, we did not observe any HR-associated recognition of an effector present in the AF by any of the 6 southern populations that we tested, e.g. those belonging to the southern highlands and southern coastal groups as described by Böndel (2015) and Stam et al. (2019b) (Figure 2). Populations from the northern and central regions do show recognition of at least one of the effectors present in the AF, as HR development was taking place. Yet, we did not observe this recognition to take place in all tested plants of the population. Rather, we observed differences in recognition capacities within the population, with some plants being able to respond with an HR, and some not. Populations from central regions showed recognition of components present in the AF, ranging from 10-100% of the plants tested in the different populations (Table S1). Two northern populations tested showed AF mix recognition for 75% and 80% of the plants tested (Table S1). All of our infiltration experiments were performed at least three times, in leaves from the same perennial plants and at the same location.
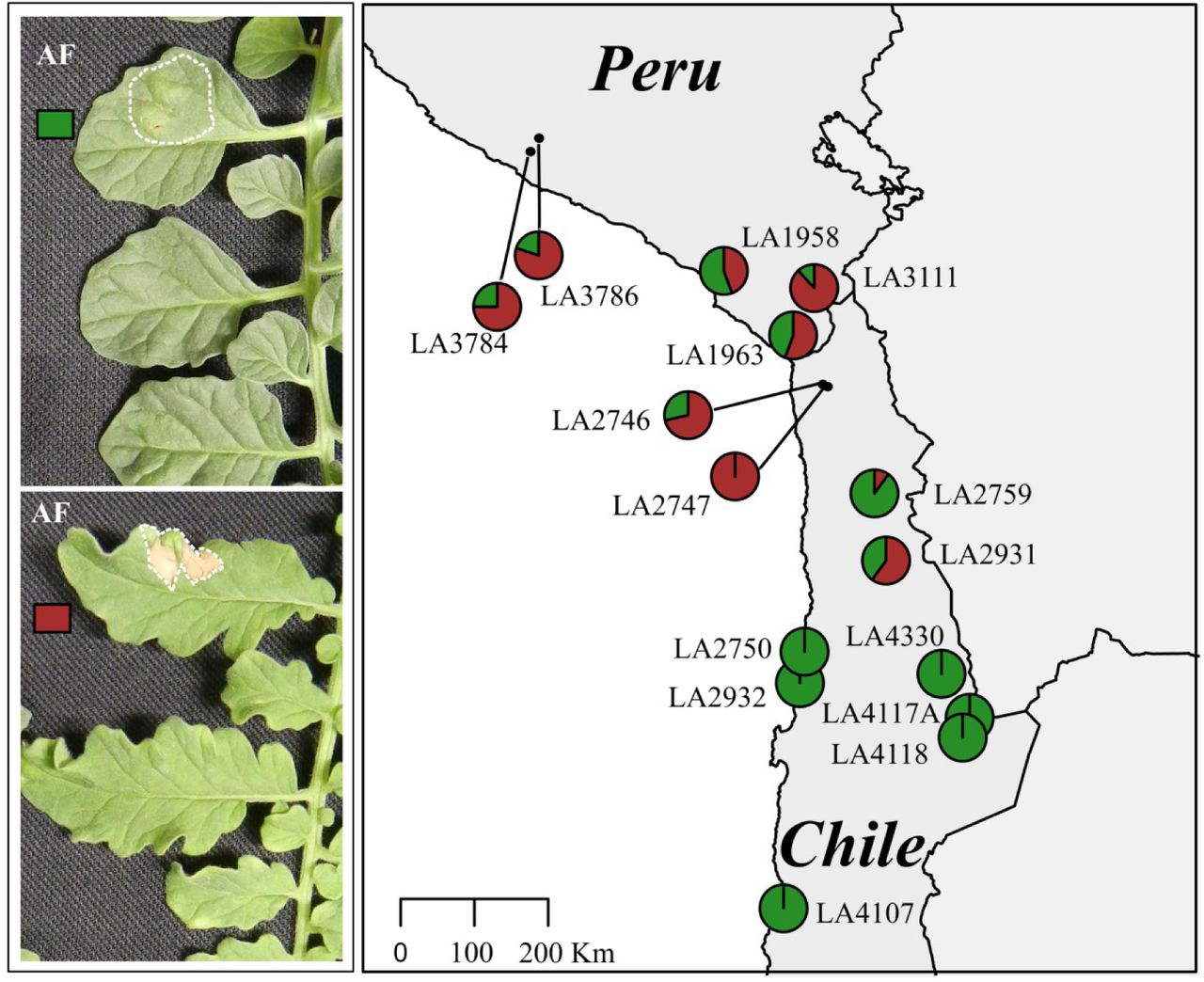
Infiltration of AF in populations of S. chilense. Leaf of an individual plant with no recognition of components present in the infiltrated AF (top left) and leaf of an individual plant recognizing at least one component present in the AF (bottom left). The infiltrated areas are indicated with white dotted lines. The map on the right shows the geographical distribution of AF component perception in S. chilense. The map shows the geographically distinct populations of S. chilense and their response to infiltration with AF Each pie chart indicates one population with 8 to 17 individuals. The brick red fraction represents plants that respond to C. fulvum AF, whereas green indicates the fraction of non-responding plants.
Differential Avr9 and Avr4 recognition patterns are present in S. chilense
Recognition of Avr9 and Avr4 has been hypothesized to be an important conserved feature in wild tomato species in order to maintain C. fulvum resistance (Kruijt et al., 2005). To understand the role of the recognition of these Avrs in more detail, we performed additional infiltration assays, but now with the individual effectors. In order to test whether S. chilense plants are able to specifically recognize Avr9, we infiltrated the plants with a preparation enriched for Avr9. Populations from the central region showed a very large variation in the ability to recognize Avr9, with the capacity to recognize Avr9 ranging from 10-80% of the plants that were tested in individual populations (Table S1). The two northern populations that were tested showed Avr9 recognition for 20% and 37.5% of the plants tested (Figure 3).

Geographical distribution of Avr9 and Avr4 perception in S. chilense. The map shows populations of S. chilense and their response to infiltration with Avr9 and Avr4 (purple when there is an HR-associated response to both), Avr9 (blue when there is an HR-associated response) and Avr4 (red when there is an HR-associated response). White sectors indicate plants showing no recognition of either Avr9 or Avr4. The infiltrated areas are indicated with red dotted lines. Each pie chart indicates one population with 8 to 17 individuals.
We also tested the ability of S. chilense to recognize Avr4 in a similar fashion, using Pichia pastoris-produced Avr4. In the two northern populations we observed that 12.5% and 30% of the plants showed the capacity to recognize Avr4, whereas in the central populations 0-33% of the plants showed recognition. Interestingly, contrary to previous reports that Cf-4 and Cf-9 are mutually exclusive, we also found plants that were able to recognize both Avr4 and Avr9 (Figure 2 and Table S1).
Presence of canonical Cf-9-, 9DC- and Cf-4-specific region in different S. chilense populations does not correlate with their recognition properties
It has been shown that in S. pimpinellifolium either Cf-9 or 9DC are responsible for Avr9 recognition (Van der Hoorn et al., 2001). Cf-9 or 9DC gene sequences in S. chilense have not been reported to date. Putative full length Cf-9 or 9DC genes cannot be found in the currently available reference genome sequence, possibly due to misassemblies of the complex LRR regions (Stam et al., 2019a). Thus, to investigate which gene is responsible for Avr9 recognition in S. chilense we looked into the presence of Cf-9 and/or 9DC, using gene-specific primer sequences that were used before to identify these genes (Van der Hoorn et al., 2001).
We performed Cf-9 gene-specific PCR amplification on genomic DNA isolated from nine S. chilense populations (9 plants per population), using the previously published primers CS5-CS1, amplifying a 379bp specific region of Cf-9 (Van der Hoorn et al., 2001). Cf-9 and Cf-4 introgression lines of S. lycopersicum served as positive and negative control, respectively. The Cf-9-specific sequence was amplified from all plants, although with different efficiency from all plants from the phenotyping assay. However, there was no association between presence or abundance of the amplicon and the response to Avr9 (Figure 4a).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Amplification of the canonical Cf-9, 9DC and Cf-4 region in geographically distinct populations of S. chilense. a) The CS5/CS1 primer pair was used to amplify a fragment of Cf-9 (378 bp), the DS1/CS1 primer pair for 9DC (507bp) and the PSK047/PSK050 primer pair for Cf-4 (728bp). As a PCR control part of the coding region of elongation factor 1 alpha (EF-1α) was amplified using the primer pair RS003/RS004 (400 bp). MM-Cf-9 and MM-CF-4, and LP12 (9DC) were used as controls (Figure S2). + or – are indicative of Avr9 and Avr4 responsiveness upon infiltration of these effectors (Figure 2). b) Map shows the populations used in the analysis.
In a similar way, we tested the presence of 9DC using the gene-specific primers DS1-CS1 (product size 507bp). LP12 and MM-Cf-9 served as positive and negative control, respectively. We found complete absence of the 9DC canonical region in our populations (Figure 4a). Lastly, we also evaluated the presence of Cf-4, using newly designed primers (PSK047-PSK050) that fall over the intron that defines the difference between Cf-4 and Cf-9 (Figure S2) and amplify a 786bp product that is only present in Cf-4. We found that this Cf-4-specific region is present in a few plants belonging to the central population, suggesting that unlike in other Solanum. spp., Cf-4 and Cf-9 are not mutually exclusive in S. chilense (Figure 4). In addition, the presence of bands corresponding to the canonical Cf-4 region in plants non-responsive to Avr4 and the absence in responders, suggests that other or new recombinant gene products with differences in their function exist in the different populations.
Cf-9 is expressed in a southern population
To test whether failed Cf-9(-like) gene expression is responsible for the complete loss of Avr9- triggered HR in the southern populations; we extracted RNA from 10 individuals from the LA4330 population. We used a primer pair PSK009-PSK010 that should allow us to amplify cDNA originating from Cf-9 orthologs and performed semi quantitative RT-PCR on plants infiltrated with water and Avr9 (8 hours after infiltration), assuming that Avr9 recognition might upregulate Cf-9(-like) gene expression.
Without treatment, we found a transcript expressed in all tested S. chilense plants, as well as in our MM-Cf-9 control plant (Figure S4), indicating that steady state Cf-9 gene expression is not affected in the southern S. chilense populations.
As expected, in the MM-Cf-9 plant we see a stronger band at 8 hours after treatment, indicative of the upregulation and positive feedback induced by Cf-9, a trait that is not visible in the southern S. chilense population (Figure S4).
Loss of resistance might be due to mutations in Cf-coreceptors the southern populations
Seeing that steady state expression levels of Cf-9 are not affected in the southern populations and complete loss of resistance is unlikely to result from detrimental mutations on all individual Cf genes. We hypothesize that the general loss of Cf responses likely results from changes in Cf regulatory genes, rather than mutations in the individual Cf genes themselves. Several core regulators or co-receptors are known to regulate the Cf protein function.
To test this hypothesis, we examined genome sequence data that are available for three plants, which are from representative populations of the central, southern coastal and southern highlands region (LA3111, LA2932 and LA4330 respectively) (Stam et al., 2019a). We extracted and aligned the genomic sequences of the co-receptors SERK3a (also known as BRI1-ASSOCIATED KINASE 1, BAK1) and SOBIR1, the adaptor ACIK1, required for regulation of the complex, as well as another regulatory co-receptor of the BAK1-containing complex, BIR2 and its homolog BIR2b (Table S2). In all cases, the extracted sequence data show polymorphisms between the reference genome of S. lycopersicum Heinz1706 and the Avr4 and Avr9 responsive plant from LA3111. In-frame indels were found for BIR2b between its sequence in S. lycopersicum and those in the three S. chilense populations, yet, the three S. chilense populations show similar, complete sequences. Several unique polymorphisms exist in the southern populations, e.g. occurring in the plants from LA2932 and LA4330, but not in LA3111 from the central region or in the S. lycopersicum reference genome sequence, resulting in up to 19 non-synonymous amino acid changes for SERK3a in the plant from LA4330 (Table 1). In ACIK1, both the LA2932 and the LA4330 plant have unique indels not present in the S. lycopersicum reference genome sequence and in the LA3111 plant (Table 1). These results suggest that there are various possible amino acid changes these receptors, co-receptors and adaptors that could be responsible for the observed loss of resistance.
Number of non-synonymous SNPs or indels observed in the open reading frame of genes encoding co-receptors or adaptors of Cf genes in the two southern populations LA2932 and LA4330.
Discussion
Natural plant populations are hypothesized to maintain a certain durable resistance against naturally co-occurring pathogens, yet little is known about the dynamics and the underlying genomic diversity in these populations throughout a species range, which result in the maintenance or loss of durable resistances.
C. fulvum is likely a natural pathogen of wild tomato species and known functional Cf resistance genes have been isolated from several wild tomato species. Yet, the physiological interaction between C. fulvum and wild tomato has not been documented. We show that C. fulvum race 5 is able to infect S. chilense. The formed intercellular hyphae resemble those observed in cultivated tomato, meaning that also infection in nature will lead to severe reduction in photosynthetic potential and thus will lead to loss of host fitness. We observed, both by the naked eye, as well as using microscopy and staining, that after inoculation with C. fulvum plants from a S. chilense population from the southern edge of the species range (LA4330) show higher susceptibility when compared to plants from a population from the central part of the range (LA3111). These findings were confirmed via quantification of fungal DNA. Thus, we show a compatible interaction between C. fulvum and wild tomato species, but also clear differences in resistance between the host populations.
The interaction between C. fulvum and tomato has been proven to be governed by gene-for-gene interactions, in which secreted Avrs from C. fulvum are recognized by corresponding RLP product of Cf gene from tomato (Joosten & de Wit, 1999; Stergiopoulos & de Wit, 2007). We tested recognition of such Avrs in different populations of S. chilense from different geographical locations with diverse climatic conditions, to understand whether the observed resistance and susceptibility within S. chilense populations to C. fulvum follows a specific geographical pattern. We phenotyped fifteen populations of S. chilense covering the whole species range by infiltrating an apoplastic extract potentially containing all Avrs except for Avr5, which is able to elicit immune responses that result in an HR. In northern and central populations, around 70% of the plants recognized at least one Avr present in this extract. Populations from the southern highlands and the southern coastal genotype groups showed no Avr recognition. These results point at two important conclusions. First of all, Avr recognition in the northern and central regions is not as conserved as previously hypothesized and secondly, Avr recognition appears to have been completely lost at the southern edge of the species range.
Previous reports suggest that Avr9 and Avr4 are the main factors in recognition of C. fulvum and this recognition is conserved throughout all wild tomato species (Kruijt et al., 2005). We observe Avr9 recognition to be present in 29% of the tested plants, whereas 11% of the plants recognized Avr4. These findings are somewhat contrasting with earlier results. Kruijt et al. (2005) tested only a small number of plants per population and did not report on the actual amounts of responding plants. In line with our results, they showed Avr4 recognition in 16 out of 20 tested populations and comparison of the accession numbers and linkage to their geographical origin, confirmed that only four of the populations tested by Kruijt et al. belonged to the southern genotype groups. Surprisingly, Avr9 recognition was not observed in all 20 accessions of S. chilense tested by Kruijt et al. (2005), although similar Avr9 have been used. These differences in results could be explained by the fact that all previous reports used young seedlings to allow quick screening of the plants. In our assays we use fully mature (over 1-year-old) adult plants, and we performed repeated infiltrations of the same plants. In addition, we observed differences in the strength of the HR upon Avr9 infiltration in young and fully developed mature leaves (Figure S3). Our finding therefore poses the first evidence of Avr9 recognition in S. chilense and suggests that detailed testing of other species under different conditions might yield novel interesting results. In about 6% of the tested plants we now show dual recognition of both Avr4 and Avr9, which has not been shown earlier.
It has been shown that Avr9 can be recognized by two Cf-9 homologs, referred to as Cf-9/Hcr9- 9C and the recombinant 9DC. For both of them, allelic variants with only a few nonsynonymous mutations are known. We evaluated the presence of known canonical domains that define Cf-9 and 9DC and found that the tested plants in all our populations do have the Cf-9 domain but are lacking the 9DC domain. The sole presence of the Cf-9 domain is an interesting contrast with the findings of Van der Hoorn et al. (2001), who showed the presence of 9DC to be predominant in another wild tomato species, S. pimpinellifolium, and a complete loss of Cf-9 in southern populations of that spoecies. Note that the southernmost S. pimpinellifolium populations are geographically relatively close to the most northernly S. chilense populations that were analyzed in our current study, yet they populate clearly different ecological niches (Peralta et al., 2008).
Likewise, we also evaluated the presence of the Cf-4 canonical region in our populations. The canonical Cf-4 domain is present in some individuals, but this does not correlate with our phenotypic data on the development of an HR upon infiltration with the Avr4 protein. There can be various reasons for this observation. First, the annealing sites of the Cf-4 primers used might carry crucial SNPs in S. chilense, resulting in failure of the PCR no detectable bands. Second, the targeted region might be missing in the gene coding for the Avr4-responsive Cf protein, which would not be surprising as similar mechanisms of recombination events, resulting in the generation of a new gene of which the encoded protein has retained its recognition specificity, have been reported earlier for other Hcr9s, albeit on a phylogenetic rather than population scale (Van der Hoorn et al., 2001). Moreover, a study on Cf-2 identified 26 different homologs in S. pimpinellifolium populations and revealed possible presence/absence variation of Cf-2 among individuals (Caicedo & Schaal, 2004). Presence of canonical regions of Cf-4 in the individual with Cf-9 canonical region confirms the dual presence of previous though allelic gene which can be possibly due to heterozygosity at the locus.
Recombination and gene conversion have been shown to play a major role in gene family evolution for RLP family as well as other resistance gene families (Paniske et al., 1997; Mondragon-Palomino & Gaut, 2005; Mondragon-Palomino et al., 2017). Recently, it has been reported that such gene conversions, or micro recombinations, of NLR genes can also be observed between accessions of another wild Solanum sp., leading to an alternative mechanism to maintain different functional alleles (Witek et al., 2020). We conclude that also the Cf gene family is likely not conserved sensu stricto, and hypothesize that a large number of possibly functional alleles are formed and maintained through intragenic micro recombinations, not just on a phylogenetic scale, but also between or even within populations of the same species.
The loss of Avr recognition in southern populations could be explained by the possibility that these plants recognize different Avrs which are absent in C. fulvum race 5. Yet, our findings show that northern and central populations of S. chilense do possess resistance against C. fulvum race 5. Our apoplastic extract contains at least 70 secreted effectors, which are all potential Avrs (Mesarich et al., 2017). The chance of losing recognition for all of these effectors by mutations in the matching receptors is highly unlikely, thus hinting at general differences in the regulation of Avr recognition. Semi quantitative RT PCR experiments showed that Cf-9-like genes appear to be expressed in both Avr9-recognizing and non-recognizing plants. Single point mutations in genes encoding essential co-receptors, like SOBIR1 or BAK1 have been shown to lead to loss in ability to induce HR when expressed in a heterologous system (Bi et al., 2016) or Arabidopsis thaliana (Albert et al., 2019). Thus accumulation of deleterious mutations in such coreceptors or contrarily, accumulation of beneficial mutations related to, for example drought adaptation, that generate a trade of in resistance, are possible. BAK1/SERK3a is a co-regulator, in not only defense responses, but the receptor-like kinase also plays a role in general cell regulatory processes and is drought-responsive (Schwessinger et al., 2011). Genomic data for non- responding plants from the southern populations revealed several nonsynonymous mutations, as well as indels in multiple RLP/Cf co-receptor-encoding genes, when comparing them to either the sequence from the LA3111 reference plant, or the S. lycopersicum reference Heinz 1706, thus suggesting that changes in downstream regulation mechanisms could have caused the loss of Avr recognition in the south. Follow-up studies should determine which of the many altered genes are causal. Since the co-receptors have multiple functions apart from Cf protein signaling, such experiments might simultaneously shed light on how the different components of the signaling network interact.
Loss of resistance over time at the population level is rather poorly understood, with some theories possibly explaining this mechanism (Koskella, 2018). For instance, evolutionary loss of resistance in certain populations might happen as a result of genetic drift, e.g. after a severe bottle neck. However, if these populations would encounter any pathogen pressure afterwards, this would be detrimental. Loss of resistance can be the outcome of random processes that are triggered to take place due to loss of selection pressure, but this loss can also be the result of accelerated evolution due to the, much debated, assumed fitness costs of carrying obsolete resistance genes (Tian et al., 2003; Sheldon & Verhulst, 1996). The role of distribution of C. fulvum in shaping the evolutionary distribution of Cf-2 homologs has been proposed by Caicedo (2008).
Giraud et al. (2017) point out that parasites need to develop a local adaptation to their environment beyond their host. Loss of resistance, as observed in our system, strongly suggests that a mechanism of ecological feedback is taking place, where the ecology of the population becomes a driving force to lose resistance or to maintain it. Therefore, the absence of the C. fulvum pathogen in the southern locations, due to extremely dry climatic conditions, which are not suitable for infection by C. fulvum, would be a plausible explanation. Interestingly, higher susceptibility to other pathogens also requiring relatively high humidity for successful infection, like Phytophthora infestans and Alternaria sp., was already described for southern populations of S. chilense (Stam et al., 2017). Seeing that some co-receptors potentially play a role in abiotic stress responses, a third possibility would be that the loss of resistance comes as a tradeoff for environmental adaptation. Yet, this is likely still intrinsically coupled to a decreased pathogen pressure.
In conclusion, we show that Hcr9 locus in S. chilense is much more complex than was thought before (Parniske et al., 1997; Van der Hoorn et al., 2001; Kruijt et al., 2004; 2005). This might in part be due to the stronger niche differentiation, or the larger heterozygosity of S. chilense specifically (Moyle, 2008). However, it is also likely that there is a lot of undiscovered diversity in the Cf gene family present in other Solanum species. Furthermore, we provide an example of the loss of resistance in a wild tomato species at the edge of its geographical distribution, possibly explained by changes in the underlying immune receptor complexes.
Overall this study provides a step forward in terms of placing major gene-mediated molecular defense mechanisms in an ecological context.
Conflicts of interests
The authors declare that no competing interests exist
Author contributions
Conceptualization: RS, RH, PSK and MJ; Investigation: PSK, MSS, GZ and DS; Contribution of materials: MJ; Funding acquisition: RS; Writing: PSK and RS. All authors reviewed and approved the manuscript.
Acknowledgements
We like to thank Liza Keitel and Lina Muñoz for help with the experiments, Sabine Zuber, Bärbel Breulmann and Anneliese Keil for maintaining the S. chilense populations and members of SBF924 for fruitful discussions and useful feedback.
Footnotes
Funding: This work was funded through the German Science Foundation (DFG), SFB924
References




