

Abstract
Peripersonal space is a multisensory representation of the space near body parts facilitating interactions with the close environment. Studies on non-human and human primates converge in showing that peripersonal space (PPS) is a body-part-centred representation that guides actions. Because of these characteristics, growing confusion conflates peripersonal and arm-reaching space (ARS) that is the space one’s arm can reach. Despite neuroanatomical evidence favors their distinction, whether PPS and ARS tap into different spatial representations remains poorly understood. Here, in five experiments we found that PPS differs from ARS in male and female human participants (N = 120), as evidenced both by their performance and the modeling of their multisensory facilitation. We mapped multisensory facilitation in detecting touches at the hand, placed in different locations radially within ARS. Results showed that 1) PPS is smaller than ARS; 2) multivariate modeling of spatial patterns of multisensory facilitation predicts well the position of the participants’ hand within ARS; 3) multisensory facilitation maps shift according to changes of hand position, revealing hand-centred coding of PPS, but not ARS; and 4) cross-correlation analyses highlight isomorphic multisensory facilitation maps across hand positions, suggesting their functional similarity to the receptive fields of monkeys’ multisensory neurons. In sharp contrast, ARS mapping produced undistinguishable patterns across hand positions, cross-validating the conclusion that PPS and ARS are distinct. These findings call for a refinement of theoretical models of PPS and ARS, which are relevant in constructs as diverse as action and self representation, (social) interpersonal distance, brain-machine interfaces and neuroprosthetics.
Significance Statement The peripersonal space (PPS) is a multisensory interface allowing us to interact with objects in the space close to our body-parts, playing a fundamental role for the defense of the body and for the motor control of actions. Recent research has conflated PPS with the arm-reaching space (ARS), that is whole space reachable by the arm. However, there is actually no evidence supporting this equivalence and the anatomical and functional differences between PPS and ARS have been largely overlooked. In this paper, we frame the theoretical issue in depth, validate a novel methodological paradigm across five experiments, and eventually report robust and cross-validated evidence for the distinction between PPS and ARS, supported by both multivariate and univariate analyses.
Introduction
Rizzolatti’s seminal study in primates described multisensory neurons in fronto-parietal regions, representing stimuli in the space close to the body, called peripersonal space (PPS) (Graziano & Gross, 1993; Rizzolatti et al., 1981b, 1981a). These neurons display a tactile receptive field (RF) on a specific body part (e.g., the hand) and a visual RF anchored to it and protruding over a limited sector (~5 to 30 cm) of space (Graziano & Gross, 1993; Rizzolatti et al., 1981a, 1981b). Neuroimaging results in humans are in line with these electrophysiological findings: the ventral (VIP) and anterior intraparietal sulcus (aIPS), the ventral (PMv) and dorsal premotor cortices (PMd) and the putamen integrate visual, tactile and proprioceptive signals, allowing for a body-part-centred representation of space (Brozzoli et al., 2011, 2012; Gentile et al., 2011, 2013; Makin et al., 2007; Serino et al., 2011). Behaviorally, visual stimuli more strongly modulate responses to touches of the hand when presented near compared to far from it (Farnè et al., 2005; Làdavas & Farnè, 2004; Serino et al., 2015; Spence et al., 2004). This advantage subserves defensive (de Haan et al., 2016; Graziano & Cooke, 2006) and acquisitive aims (Brozzoli et al., 2009, 2010, 2014; Patané et al., 2019; Vignemont & Iannetti, 2014).
As a multisensory interface guiding interactions in the surrounding environment, PPS shares some characteristics with the arm-reaching space (ARS), the space reachable by extending the arm without leaning (Coello et al., 2008). In humans, tasks usually adopted to study ARS require judging the reachability of a target stimulus or performing reachability movements, revealing a constant overestimation of this portion of space (Carello et al., 1989; Coello et al., 2008; Coello & Iwanow, 2006). Despite their anatomo-functional differences (Desmurget et al., 1999; Filimon, 2010; Lara et al., 2018; Pitzalis et al., 2013), some research on PPS in humans grew apart from the original electrophysiological findings and conflated ARS and PPS, defining PPS as ARS, or testing PPS using reachability tasks (Coello et al., 2008; Iachini et al., 2014; Vieira et al., 2020). In stark contrast with their alleged identity, multisensory stimuli within ARS and close to the hand do activate neural areas typically associated with PPS, whereas the same stimuli within ARS, but far from the hand, do not (Brozzoli et al., 2012; Graziano et al., 1994).
In five experiments, we tested the hypothesis that PPS and ARS are distinct spatial representations. In Experiments I and II, we tested the multisensory facilitation in a speeded tactile detection task, considering it as a proxy of PPS extent. To probe the ARS extent, from Experiments III to V, we adopted a reachability judgement task (Bourgeois & Coello, 2012) to compute the point of subjective equality (PSE), the individual estimation of the perceived extent of ARS. Across experiments, we manipulated the vision (visible or not, see Làdavas et al., 2000) and position (close or distant) of the participants’ hand. If PPS and ARS are equal, we should observe similar multisensory facilitation from all visual stimuli falling within ARS, but not those outside, independently of hand position. Therefore, the two positions of the hand should elicit undistinguishable spatial patterns of multisensory facilitation, up to the ARS limit, either objectively (the 6th position) or subjectively measured (PSE). Conversely, if they are different we should observe multisensory facilitation only from visual stimuli near the hand but not from those far from it, irrespective of their reachability: the hand positions would elicit specific and distinguishable spatial patterns of multisensory facilitation. Our results show that hand position was robustly classified from the distribution of multisensory facilitation in Experiment I, but not in Experiment V. Thus, the proximity of the visual stimuli to the hand–not their reachability– predicts the increase in multisensory facilitation. Cross-correlation and univariate analyses further demonstrated that visual boosting of touch is hand-centred following changes in hand position. Taken together, these results converge in showing that PPS and ARS are not superimposable.
Methods
Participants
Overall, 120 participants (56 females; mean age = 25.12 years, range: 18-41) took part in a total of 5 behavioural experiments. Twenty-seven subjects (13 females; mean age = 26.12, range: 20-34; mean arm length = 79.41 ± 5.83 cm, measured from the acromion to the tip of the right middle finger) participated in Experiment I; twenty-one subjects (9 females; mean age = 25.71, range: 19-43; mean arm length = 75.98 ± 4.56 cm) in Experiment II; twenty-five subjects (13 females; mean age = 26.17, range: 20-33; mean arm length = 79.37 ± 6.07 cm) in Experiment III; twenty-four subjects (14 females; mean age = 23.75, range: 18-38; mean arm length = 79.28 ± 5.10 cm) in Experiment IV; and twenty-five (16 females; mean age = 24.44, range: 18-41; mean arm length = 78.46 ± 7.26 cm) in Experiment V. All the experiments adopted a within-subject design.
All participants reported normal or corrected-to-normal vision, normal tactile sensitivity and no history of psychiatric disorders. The participants gave their informed consent before taking part in the study, which was approved by the local ethics committee (Comité d’Evaluation de l’Ethique de l’Inserm, IRB 00003888) and was in line with the Declaration of Helsinki. The participants were remunerated 15 euros per experiment.
Stimuli and Apparatus
Visual stimuli were identical for all the experiments. We used a projector (Panasonic PT-LM1E_C) to present 2D grey (RGB = 32, 32, 32) circles in 10 different positions, ranging from the space near to the space far from the body. Positions were calibrated based on the participant’s arm length, ensuring that the sixth one corresponded to the objective limit of reachability for each participant. To verify this, before the experiment, the participants had to put the chin on a chinrest (with a fixed height of 30 cm) and place their right hand as far as possible on the table, keeping the eyes closed and the shoulders parallel to the side of the table. After the measure, the arm was removed from the table by the experimenter, preventing the participant from seeing the objective reachable limit. Starting from the sixth position, 4 visual stimuli were projected beyond the reachable limit, and 5 were projected closer to the participant’s body, 8 cm rightward with respect to the body’s sagittal axis; the distance between each visual stimulus centre was 9 cm. The diameter of the light dot (3 cm at a distance of 57 cm from the eye) was corrected for retinal size with the formula:
 where 3 cm is the diameter of the stimulus, 57 cm is the distance from the eye at which 1° of the visual field corresponds to 1 cm and x is the distance of the centre of the stimulus from the point at 57 cm, measured on the table.
where 3 cm is the diameter of the stimulus, 57 cm is the distance from the eye at which 1° of the visual field corresponds to 1 cm and x is the distance of the centre of the stimulus from the point at 57 cm, measured on the table.
The visual stimulus duration (500 ms) was the same in all experiments, as was the fixation cross (2.5 cm) that was projected along the body’s sagittal axis (see Fig. 1).
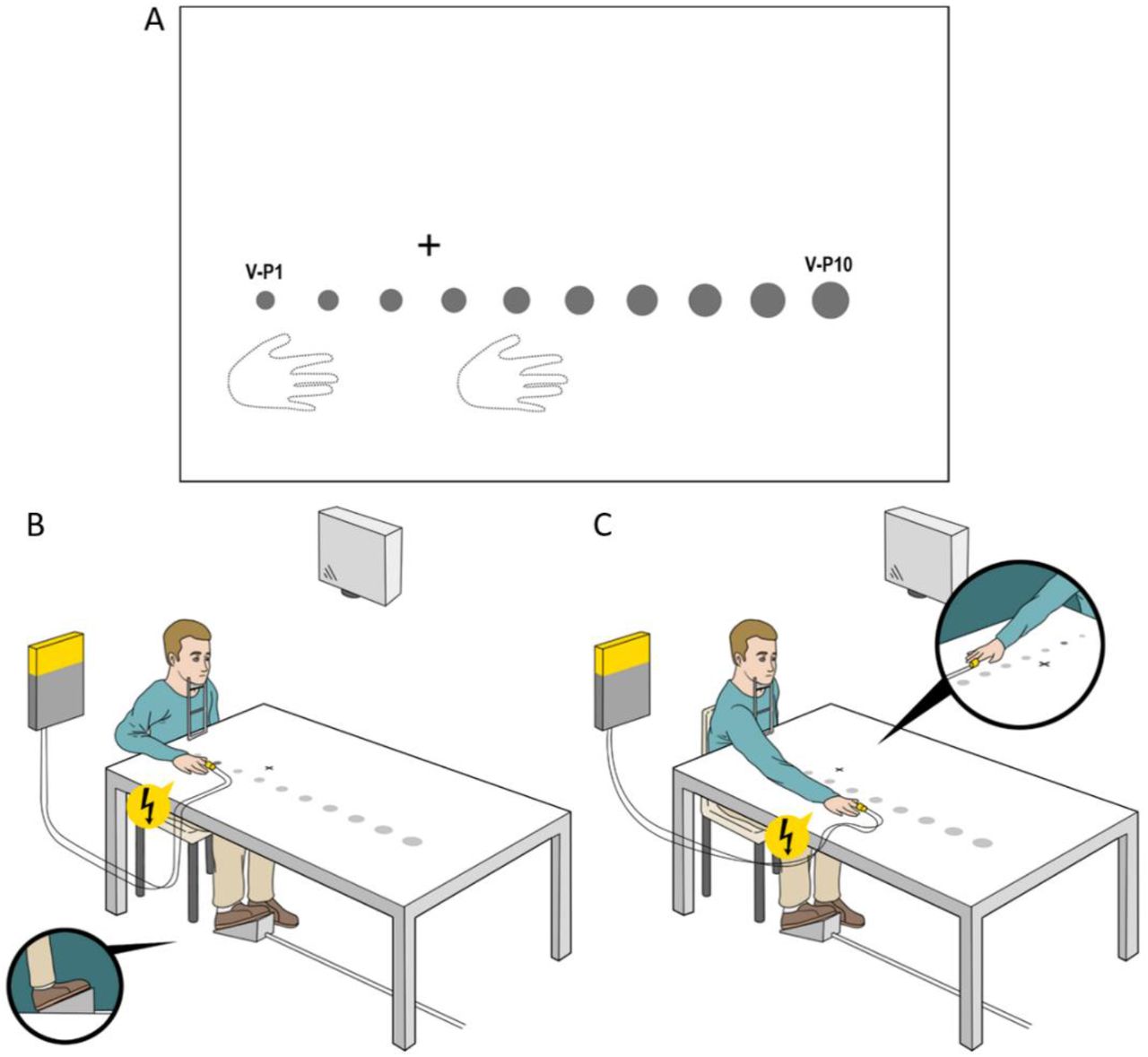
a) Positions of right hand (Experiment III was the only one in which the right hand was not visible, i.e., under the table), fixation cross, and visual stimuli. b) and c) represent the close hand (b) and the distant hand condition (c). In all experiments, the visual stimuli (here displayed as grey circles) were projected one at a time, in one of the ten possible positions (from V-P1 to V-P10), corrected for retinal size (a-c). Tactile stimuli, when present, were applied on the right index finger, synchronous with the eventual visual one. Globally, we adopted two conditions of unisensory stimulation (only tactile or visual stimulation) and a multisensory condition (visuo-tactile stimulation). To these, we added catch trials (nor visual nor tactile stimuli presented) to monitor participant’s compliance.
Tactile stimuli were brief electrocutaneous stimulations (100 μs, 400 mV) delivered to the right index finger via a constant current stimulator (DS7A, DigiTimer, UK) through a pair of electrodes (1.5*1.9 cm, Neuroline, Ambu, Denmark). In Experiments I, II and V, the intensity of the tactile stimulation was selected through an ascending and a descending staircase procedure, incrementing and decrementing, respectively, the intensity of the stimulation to find the minimum intensity at which the participant could detect the tactile stimulus 100% of the time over 10 consecutive stimulations. To ensure high accuracy, the intensity was further increased by 10% before the first and third experimental blocks. Overall, Experiments I, II and V required four blocks each (structure of the blocks described in Design & Procedure).
Design & Procedure
After measurement of the subject’s arm length, in Experiments I, II and V, we applied the tactile electrodes to the right index finger. Depending on the experiment, the participants were informed about the experimental demands. In Experiment I, the participants were asked to perform a tactile detection task; thus, a tactile stimulation on their right index finger was applied, accompanied or not by a 2D light dot projected in one of the 10 possible visual positions (hereafter named V-P1, V-P2… V-P10 from the closest to the farthest one). The participants rested with their head on the chinrest and eyes on the fixation cross. The right hand was placed on the table 16 cm rightward from the body’s sagittal axis, with the tip of the medium finger in correspondence to V-P2 (hereafter close hand) or V-P6 (hereafter distant hand). For each hand posture, our experimental design included 16 visuo-tactile stimulations (hereafter VT) per visual position (160 VT trials; 69% of the total trials) and 16 unimodal tactile trials (T trials; 7%). Moreover, we included 4 unimodal visual trials per position (V trials; 17%) and 16 trials with no stimulation at all (N trials; 7%) as catch trials. These catch trials were included to avoid automatic motor responses connected to a frequently repeated target. In each trial, the participants had to focus on the tactile stimulus and respond as fast as possible via a single pedal using the tip of the right foot. Globally, we had 76% of trials in which the tactile target was presented. In the VT trials, the visual stimulus was always presented synchronous with the tactile stimulus. The false alarm rate was very low (< 0.5%). The participants with accuracy lower than 2 standard deviations from the group mean were discarded (4 participants).
To complete the experiment, the participants had to perform two blocks with the close hand (116 randomized trials per block) and two with the distant hand (116 randomized trials per block). The order of hand postures was counterbalanced across participants.
In the reachability judgement task (Experiments III, IV and V), the 2D light dot projected in one of the 10 possible visual positions was the target stimulus. In Experiment V, we took advantage of the same settings as those used in Experiment I and applied the same tactile stimulation to the index of the right hand, which was placed in either the close or distant position. However, in this reachability judgement task, the tactile stimulus was irrelevant, and the visual stimulus represented the target. When the visual stimulus appeared, they were asked to judge whether it was reachable by using the left or the right pedal (order counterbalanced across subjects). Overall, 160 randomized V and 160 randomized VT trials were presented for each position of the hand. These 320 trials per hand position were administered in two blocks in a randomized order; the order of hand postures was counterbalanced across participants.
The procedure and design for Experiments III and IV are reported in more detail in Supplementary Notes 3 and 4.
Analyses
Preliminary analyses in all experiments were conducted on the accuracy of the performance. In Experiment I, although supra-threshold tactile stimuli were adopted, 4 participants reported accuracy more than 2 SD lower than the mean of the sample. However, these low performances were not attributable to any specific condition. Thus, those participants were discarded from further analyses. Globally, we observed high accuracy in Experiment I (> 90%) with a very low rate of false alarms (< 2%). Similarly, the participants’ performance in the reachability tasks was very accurate in Experiments III, IV and V (> 99%), and no differences emerged between conditions.
To have a direct index of the proportion of multisensory facilitation over the unimodal tactile condition in Experiment I, we calculated the multisensory gain as follows:

TM was the average RT for unimodal tactile stimuli, and VT was the raw RT for a multisensory visuo-tactile stimulus. Larger MG values corresponded to greater facilitation (namely, larger benefits for visuo-tactile with respect to tactile-only conditions). This measure is more rigorous than an absolute delta, as it allows correction of the RTs considering the subject-specific speed for each visual position and for each position of the hand. Nonetheless, for the sake of comparison to previous studies, we also calculated and analysed the delta RT. These analyses are reported in Supplementary Note 1. Averaging MG values per hand position and visual position, we obtained two vectors of 10 MG values (from V-P1 to V-P10) for each participant: one for the close hand and one for the distant hand, enabling us to mapping locations where multisensory stimulation facilitated touch perception with respect to unisensory stimulation. With a multivariate SVM approach (Vapnick, 1995), we tested whether a data-driven classifier could reliably predict the position of the hand from the spatial patterns of the MG. In particular, the classifier adopted a leave-one-out strategy: the SVM was trained on (N – 1) participants and tested on the two vectors excluded from the training, making use of a linear kernel. The overall accuracy of our classification was calculated as the sum of the correct predictions for both hand positions divided by the total number of predictions (46).
To study the multisensory facilitation more locally, we subsequently subjected the MG values to FDR-corrected (Benjamini & Hochberg, 1995) comparisons versus zero to observe in which position the multisensory facilitation resulted significantly higher levels than the baseline. Then, we performed a Hand (close vs. distant) * Position (V-P1 to V-P10) within-subject ANOVA to investigate the extent and position of the facilitation with different hand positions.
To compare the shape of the multisensory facilitation obtained for each of the two hand positions with each other, first, we tested which function better fit the spatial patterns, and second, we employed a cross-correlation strategy to prove the isomorphism of the two shapes. To this aim, MG values for both hands were fitted to sigmoidal and normal curves, both limited to two parameters, to compare them. In Table 1, we report the formulas adopted to fit the curves with the “Curve Fitting” toolbox in MATLAB (version R2016b, MathWorks, USA). Regarding the sigmoidal curve, we considered a good fit only the case in which data fitted the sigmoid with a descending slope, as already reported in the literature (Canzoneri et al., 2012; Serino et al., 2015), indicating a facilitation close to the body that fades away with increased distance.
Formulas adopted to fit the sigmoidal and normal curves to reachability judgements in Experiments III, IV and V. The same formulas were adopted to fit the curves for the multisensory gain values in Experiment I. In both cases, X represents one of the 10 experimental positions (from P1 to P10).
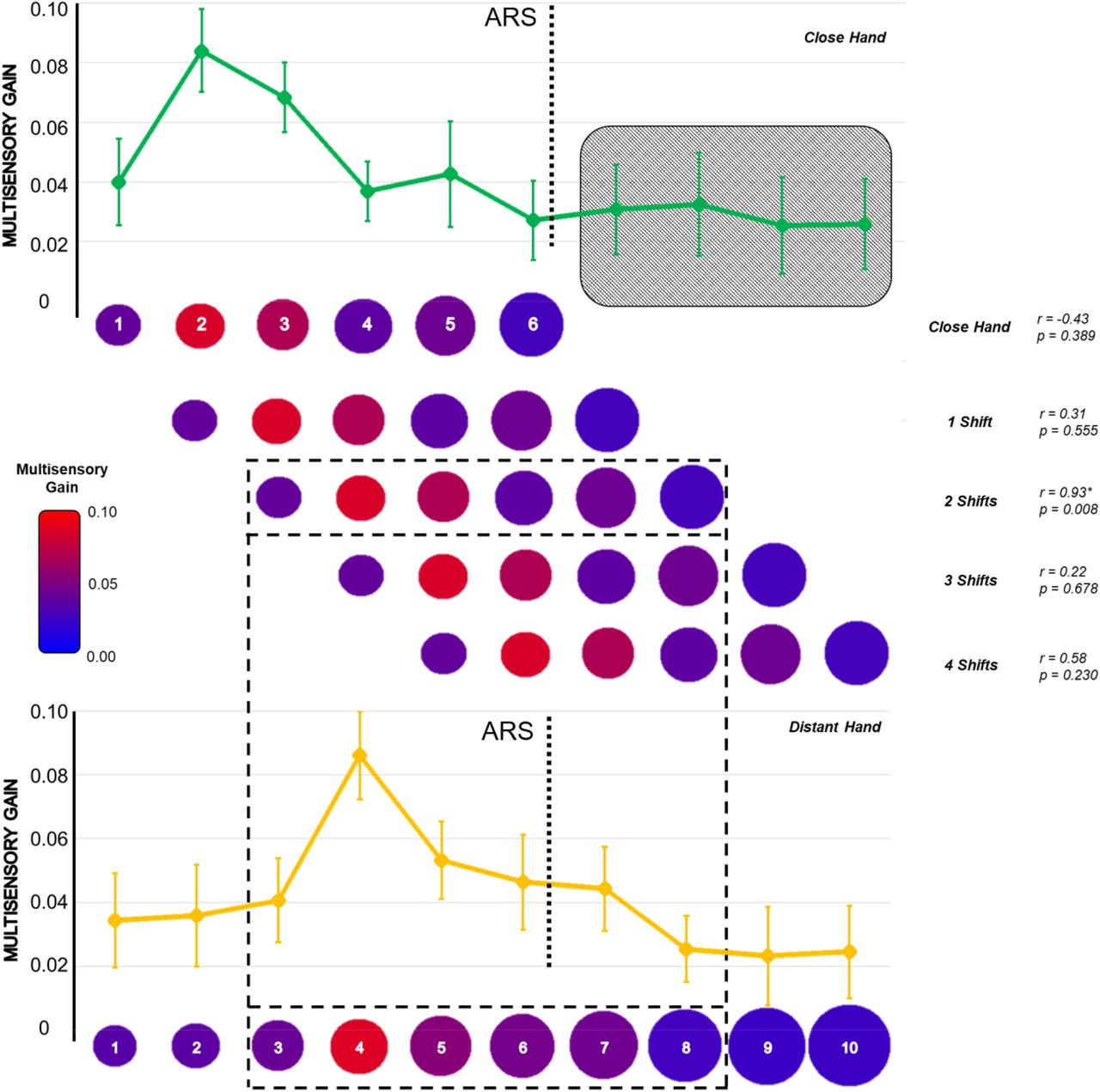
Last, we performed a cross-correlation analysis on MG values to evaluate the isomorphism of the facilitation curve for both hands. The hypothesis was that distally shifting (i.e., towards the distant hand position) the pattern of facilitation obtained with the hand close, we should obtain higher correlations due to overlap in the two curves. We considered the pattern of averaged MG values for all the reachable positions (from V-P1 to V-P6) with the close hand, and we correlated it to the pattern of six averaged MG values obtained with the hand placed far. The first correlation did not reveal any shift: we thus correlated the pattern V-P1 to V-P6 for the close hand position with the pattern V-P1 to V-P6 of the distant hand position. Subsequently, we distally shifted the pattern of the close hand one position at once, obtaining four new values of correlation: with the last shift, we correlated the pattern V-P1 to V-P6 for the close hand with the pattern V-P5 to V-P10 of the distant one.
By showing that PPS and ARS are not superimposable, we cannot yet exclude that the reachability judgement task, meant to measure ARS, might still capture some of the PPS features. Therefore, we performed three experiments adopting this task, though with the same settings of Experiment I. In Experiment III, the hand was not visible, whereas in Experiment IV, the hand was visible and placed close or distant (see Supplementary Notes 3 and 4 for details). However, in both experiments we only adopted a unisensory visual stimulation.
We thus performed Experiment V, wherein the remaining main feature that differed between the experiments was also equated. Namely, we made the ARS task multisensory by asking participants to perform the same reachability judgements while task-irrelevant tactile stimuli were concurrently presented with the visual stimulus. As such, Experiment V allowed us to assess whether the multisensory nature of the stimulation (in addition to having the hand visible and in the same positions as Experiment I) could either induce hand-centred facilitation in the reachability task performance and/or change the extent of the reachability limit. Similar to Experiment I, we calculated the MG to observe the proportion of multisensory facilitation over the unisensory visual baseline (see Fig. 4). We then tested the classifier on the MG patterns and performed the same procedures already described for Experiment I on delta RTs and MG (see Supplementary Notes 3 and 4 for details about Experiments III and IV). The percentage of “reachable” responses per position was calculated by dividing the number of “reachable” answers by the total number of trials per position (16). The percentages obtained were then fitted to sigmoidal and normal curves, taking advantage of the same formulas adopted in Experiment I. We fitted the curves separating hand positions and type of stimulation (unimodal visual versus multisensory visuo-tactile). Hand (close vs. distant) * Stimulation (visual vs. visuo-tactile) * Model (Gaussian vs. Sigmoid) ANOVA on RMSE values assessed which model best fitted the data, both at the individual and group level. As mentioned above, the best-fitting model for these data was the sigmoidal curve. Thus, we investigated the point of subjective equality (PSE, namely, the inflection point of the sigmoidal curve) and slope values by subjecting them to two separate within-subject ANOVAs with Hand (close vs. distant) and Stimulation (visual vs. visuo-tactile) as factors.
Results
Peripersonal Space: not a matter of reachability
First we tested whether a data-driven classifier could reliably predict the position of the hand from the spatial patterns of the multisensory facilitation. To this aim, we calculated the Multisensory Gain (MG) for each distance of the visual stimulation, thus obtaining a vector covering both reachable (up to the 6th position) and unreachable space, per each hand position. We then trained and tested a classifier over the two vectors for each participant, with a leave-one-out strategy. The multivariate classifier was able to predict the two positions of the hand of the test participants with an accuracy of 0.72 (33/46 correct classifications), with no bias for one hand position over the other (16/23 for the close hand and 17/23 for the distant hand). The accuracy of the classification was significantly higher than the chance level, as assessed by a binomial test (two-tailed p = 0.005). It is therefore possible to infer the position in which the hand was placed by looking at the participant’s spatial patterns of MG values. Hence, different patterns of multisensory facilitation were associated with different positions of the hand within the ARS.
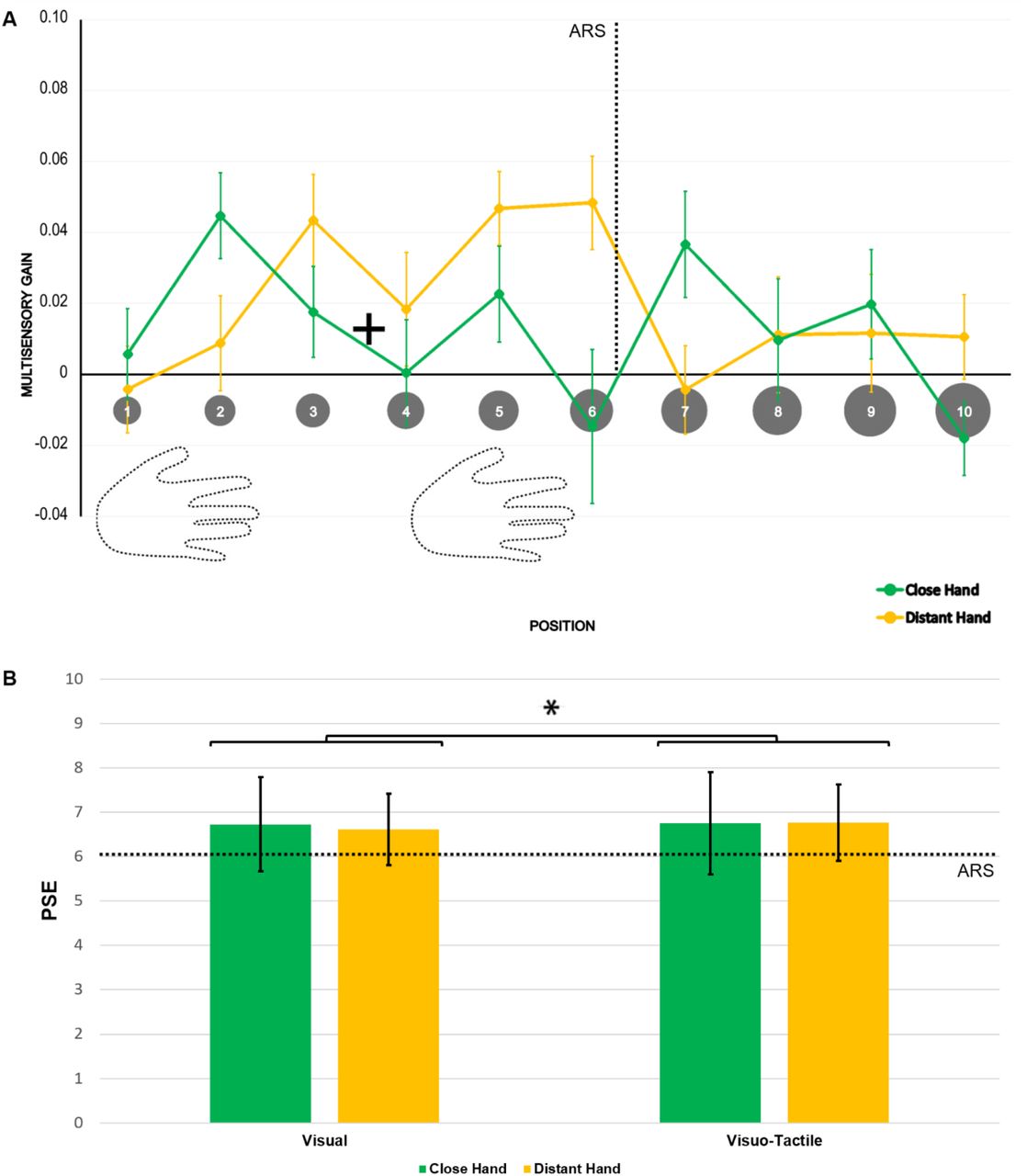
To provide a finer-grained mapping of our MG patterns (see Fig. 2A), we ran a V-Position (V-P1 to V-P10) * Hand (close or distant) repeated measures ANOVA. The main effect of V-Position was significant (F(5.73, 125.99) = 3.02, p = 0.010, η2p = 0.12). More interestingly, this effect was modulated by the position of the hand, as indicated by the significant two-way interaction Hand*V-Position (F(6.27, 137.87) = 2.92, p = 0.009, η2p = 0.12).

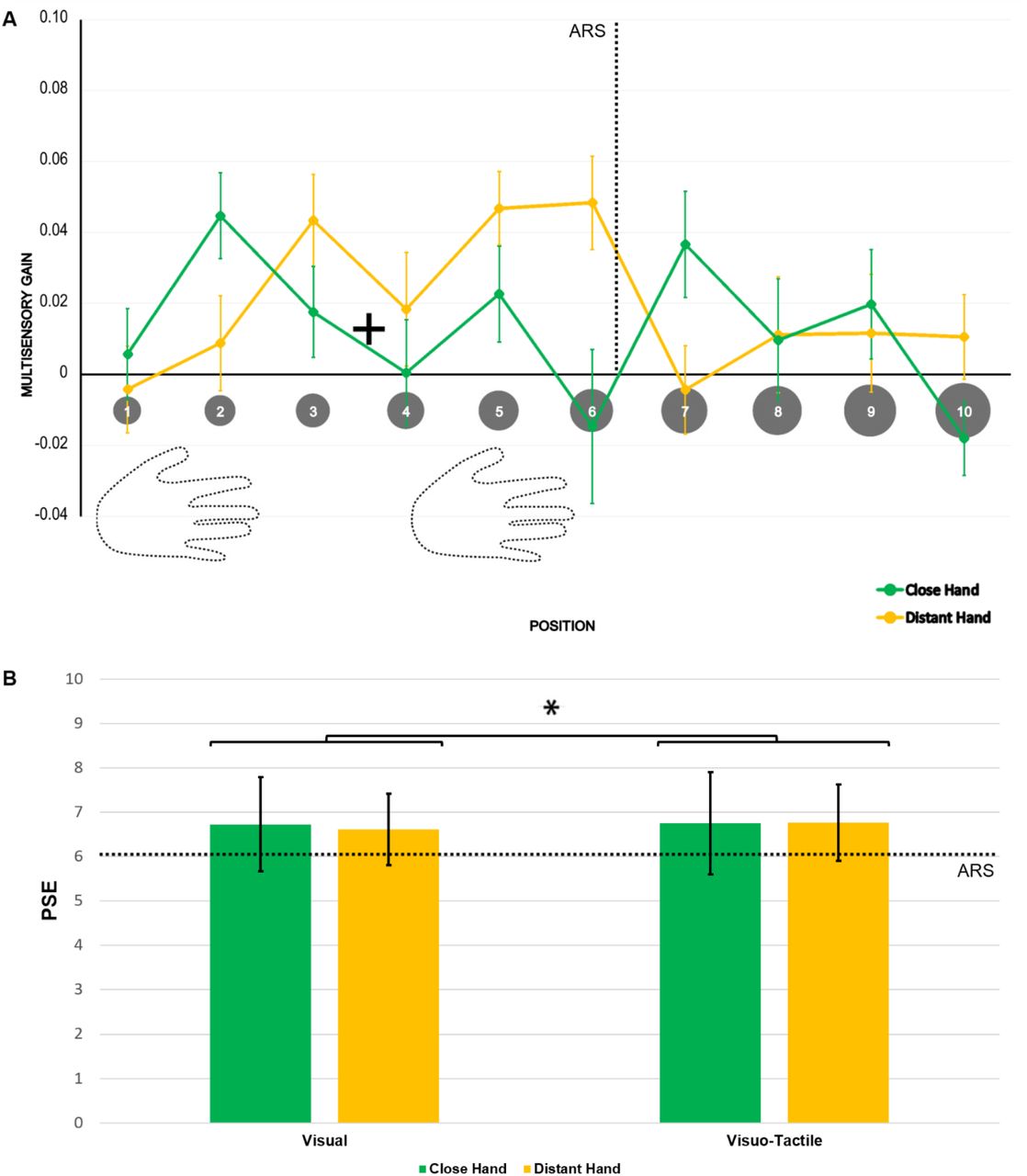
a) Multisensory gain (MG) values along the 10 visual positions, ranging from near to far space, for the distant (yellow) and the close (green) hand conditions. Higher values of MG represent stronger facilitation in terms of RT in the multisensory condition than in the unisensory tactile baseline (by definition, MG = 0). Error bars represent the standard error of the mean. Asterisks represent a significant difference (p < 0.05, corrected). b) and c) Number of trials reporting MG values greater than zero (unisensory tactile baseline) along the 10 visual positions, ranging from near to far space, for the close (b) and the distant (c) hand conditions.
Tukey-corrected multiple t-test comparisons revealed faster responses in V-P2 than in all the positions from V-P6 to V-P10 when the hand was close (all ps < 0.05); when the hand was distant, responses were significantly faster in V-P4 than in V-P1, V-P2, V-P8, V-P9 and V-P10 (all ps < 0.05). Critically, the MG was larger in V-P2 when the hand was close than when it was distant from the body (p = 0.043). This pattern was fully reversed in V-P4, where the MG was larger when the hand was distant than when it was close to the body (p = 0.039). No other differences were significant.
To identify where multisensory facilitation was significant at the single position level, we ran a series of t-tests on the MG values vs. 0 (i.e., absence of visuo-tactile facilitation over touches delivered alone), corrected for false discovery rate.64 When the hand was close, the MG significantly differed from 0 in V-P1, V-P2, V-P3 and V-P4 (all ps < 0.05). In contrast, when the hand was distant, the MG was larger in V-P1, V-P3, V-P4, V-P5, V-P6 and V-P7 (all ps < 0.05). Figure 2 additionally reports the number of trials reporting MG values greater than 0 with the hand close (2B) and distant (2C). In line with the results obtained by the repeated measure ANOVA and the t-tests, the density peak shifted in a coherent manner with the position of the hand within ARS. Similar results were obtained by analysing the delta of raw RTs for both the repeated measure ANOVA and the t-tests (see Supplementary Note 1 and Supplementary Fig. 1). We ran Experiment II to check for consistency of these findings and fully replicated these results in an additional group of naïve participants (see Supplementary Note 2 and Supplementary Fig. 2). Therefore, these findings converge to indicate that multisensory facilitation occurs in different locations, according to changes in hand position. This is evidence for the hand-centred nature of this facilitation. From this, one would expect 1) the facilitation to be maximal in correspondence with hand location and to decay with distance from the hand and 2) the bell-shaped pattern of facilitation to follow the hand when the hand changes position.
To test the first prediction, we modelled our data to a Gaussian curve. To test the alternative hypothesis, namely, that multisensory facilitation spreads all over the ARS to decay when approaching the reachable limit, we compared the Gaussian to a sigmoid function fitting (Canzoneri et al., 2012; Serino et al., 2015). The sigmoidal curve could fit the data for a limited number of participants, both for the distant (5 out of 23 subjects, 21.7%) and for the close (9 out of 23 subjects, 39.1%) hand conditions. However, fitting the Gaussian curve to the same data accommodated convergence problems for a higher number of participants: the fitting converged for 14 participants (59.9%) in the distant hand condition and for 15 participants (65.2%) in the close hand condition. Chi-squared tests with Yates’ continuity correction (Yates, 1934) on percentages of fitting cases for both distant and close hand positions confirmed that the Gaussian model fit a larger proportion of the subjects than the sigmoid model (see Supplementary Note 1). These results suggested that the facilitation was maximal in correspondence with hand location and decayed with increasing distance, following a bell-shaped pattern.
To test the second prediction that the bell-shaped facilitation should shift following the hand, we estimated the position of the peak of the Gaussian curve in each hand position. As predicted, the facilitation peak shifted according to the hand position: with the hand close, it fell between V-P2 and V-P3 (2.34 ± 1.51); with the hand distant, the peak fell between V-P4 and V-P5 (4.15 ± 1.28). We then performed a cross-correlation analysis testing whether the curves reported for the close and distant hand positions overlapped when considered in absolute terms. We reasoned that shifting the position of the hand–within the reachable space–should bring to an isomorphic facilitation around the new hand position. This, in turn, would imply the maximum correlation between MG values to emerge when the close-hand curve shifts distally (i.e., towards the distant-hand position curve). We considered the first six values of MG with the close hand (from V-P1 to V-P6, i.e., the reachable positions) and correlated this distribution with six values of the MG for the distant hand, proceeding as illustrated in Figure 3. As predicted, we found the maximum correlation (r = 0.93 p = 0.008) when shifting the close hand distally by two positions. No other correlations were significant (all ps > 0.20).
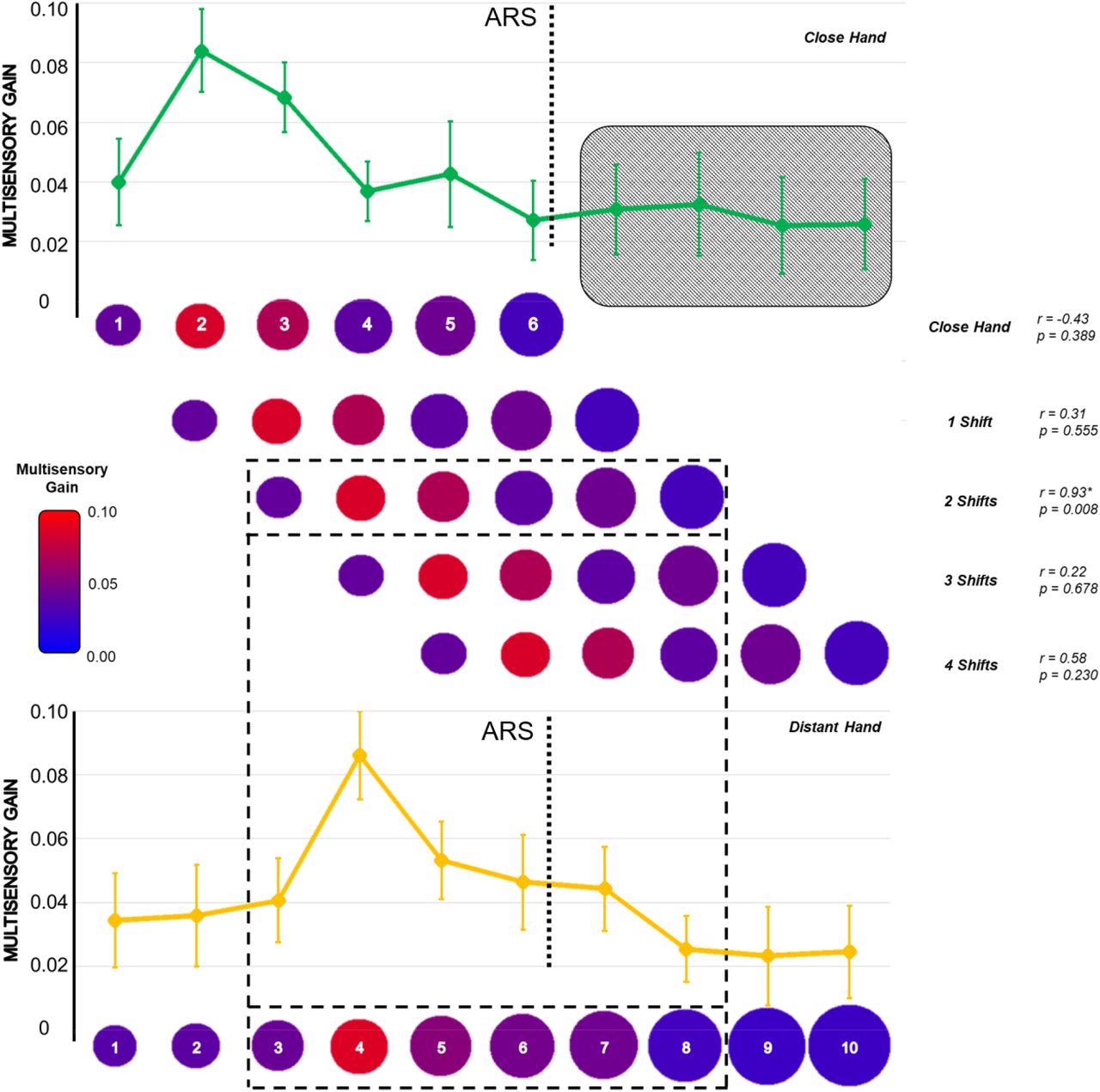
Cross-correlation analysis of distally shifting the pattern of MG values for all reachable positions with the hand close. Red colors represent higher MG values. Values of Pearson’s r and p values are reported for all the correlations performed. The black grid highlights the only significant correlation (p < 0.05).
Arm Reachable Space: not a matter of hand position
By showing that PPS and ARS are not superimposable, we cannot yet exclude that the reachability judgement task, meant to measure ARS, might still capture some of the PPS features. Therefore, we performed three experiments adopting this task, though with the same settings of Experiment I. The results of Experiments III and IV replicated well-established findings about the extent of the ARS, including the overestimation of the reachable limit (Bourgeois & Coello, 2012; Carello et al., 1989; Rochat & Wraga, 1997). Importantly, however, they failed to show any similarity with PPS, either in terms of absolute extent (ARS is larger than PPS) or position-dependent modulation (PPS is hand-centred, whereas ARS is not).
In Experiment V, taking advantage of a multisensory stimulation, we could obtain for each participant two vectors of MG values, from V-P1 to V-P10, as in Experiment I, and we could leverage a similar data-driven classifier to discriminate the close from the distant hand. However, in this case, the accuracy of the prediction was lower than that obtained in Experiment I (0.36, 18/50 correct classifications) and not significantly different from chance level, as assessed by a binomial test (two-tailed p = 0.065). Thus, in Experiment V, the multivariate and data-driven approach was unable to distinguish between the two positions of the hand within ARS.
Moreover, the Hand (close vs. distant)* Position (V1 to V-P10) within-subject ANOVA on the MG did not highlight any significant main effect (Hand: F(1, 24) = 0.83, p = 0.371; Position: F(6.31, 151.56) = 1.20, p = 0.307) or interaction (Hand*Position: F(5.35, 128.5) = 1.82, p = 0.108). However, the significant intercept (F(1, 24) = 9.80, p = 0.005, η2p = 0.29) confirmed the presence of a general facilitation produced by the multisensory stimulation with respect to the unisensory stimulation. Through multiple comparisons corrected for false discovery rate (Benjamini & Hochberg, 1995), we observed that none of the positions presented an MG significantly different from 0 (namely, no facilitation; all ps > 0.05) when the hand was close. When the hand was distant, V-P3, V-P5 and V-P6 differed from 0 (all ps < 0.05). Similar results were obtained by analysing the delta between the RTs to unisensory and multisensory stimulations (see Supplementary Note 5).
We then assessed whether the reachable/not reachable responses provided similar evidence for a difference between PPS and ARS as the RTs. To this aim, reachability judgements were fitted to sigmoidal and Gaussian curves. Within-subject ANOVA on the RMSE of these models was performed with a Model (sigmoidal vs. Gaussian) * Stimulation (visual vs. visuo-tactile) * Hand (close vs. distant) design. A significant main effect of Model emerged (F(1, 24) = 220.11, p < 0.001, η2p = 0.90). In particular, the sigmoidal curve reported the best fit, irrespective of stimulation type and hand position. For each of these variables, we estimated the coefficients of the sigmoid, obtaining the PSE and the curve slope. Through a Hand (close vs. distant) * Stimulation (visual vs. visuo-tactile) within-subject ANOVA on PSE values, we observed a main effect of stimulation type (F(1, 24) = 4.425, p = 0.046, η2p = 0.16). In particular, the mean PSE was closer to the body in the unimodal visual condition (mean ± SE = 6.67 ± 0.17) than in the multisensory visuo-tactile condition (mean ± SE = 6.76 ± 0.18). The main effect of Hand (F(1, 24) = 0.07, p = 0.794) and its interaction with Stimulation (F(1, 24) = 3.453, p = 0.075) were not significant. Last, we performed a Hand (close vs. distant) * Stimulation (visual vs. visuo-tactile) within-subject ANOVA on slope values. Neither main effects (Hand: F(1, 24) = 1.75, p = 0.199; Stimulation: F(1,24) = 0.35, p = 0.558) nor the interaction (Hand*Stimulation: F(1,24) = 0.27, p = 0.610) were significant (Fig. 4).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
A) Multisensory gain (MG) values along the 10 positions, ranging from near to far space, for the close (green) and distant (yellow) hand conditions. Higher values of MG represent a stronger facilitation in terms of RT with respect to the unimodal visual baseline (by definition, MG = 0). Error bars represent the standard error of the mean. No significant differences between hand postures emerged. B) PSE values calculated for both unimodal visual and multisensory visuo-tactile conditions for both hands. Error bars represent the standard error of the mean. Asterisks indicate a significant difference between unisensory and multisensory conditions (p < 0.05).
Discussion
The main aim of this study was to test the hypothesis that PPS and ARS are not superimposable representations. Our hypothesis was based on their anatomical-functional differences (Andersen & Buneo, 2002; Brozzoli et al., 2012; Filimon, 2010; Lara et al., 2018). PPS includes a small portion of space centred on specific body parts (Brozzoli et al., 2011, 2012; Graziano & Gross, 1993; Rizzolatti et al., 1981a, 1981b), whereas ARS would mainly depend on the arm length (Carello et al., 1989; Rochat & Wraga, 1997). Therefore, we predicted that different hand positions within ARS should result in distinguishable patterns of multisensory facilitation in a visuo-tactile PPS task. In particular, different positions of the hand within ARS should result in distinguishable patterns of multisensory facilitation in a visuo-tactile PPS task. Conversely, if PPS and ARS were superimposable, the patterns of multisensory facilitation should be undistinguishable across hand positions. Our findings clearly point against the PPS-ARS identity, whether contrasted in terms of spatial extent, by using their respective gold-standard paradigms, or in terms of pattern of multisensory facilitation. Indeed, our novel visuo-tactile paradigm allowed an unprecedented fine-grained mapping of the multisensory facilitation, revealing its limited spatial extension. The key finding is the fact that two different hand locations within ARS could be decoded from the spatial patterns of multisensory facilitation.
One could still argue that reachability tasks might capture some features of PPS. In this respect, we also validated a novel visuo-tactile version of the reachability judgement task, confirming previous findings on the extension and overestimation of ARS (Bourgeois & Coello, 2012; Carello et al., 1989). Then, the comparison of these novel multisensory paradigms resulted in two main advances arguing against the PPS-ARS identity.
First, we revealed that multisensory facilitation depends on hand position, peaking in correspondence with its location and decaying with distance from it. Thus, the extent of PPS is smaller than the ARS either objectively or subjectively measured. Were they superimposable, we should have observed faster RTs for all the reachable positions of visual stimulation. In fact, both the classifier and the location-specific differences clearly indicated that different spatial patterns of multisensory facilitation emerge for the close and distant hand positions, despite being both within the limits of ARS.
Second, when the comparison was best matched for methodological differences, our findings clearly indicated that ARS is not hand-centred, whereas PPS is. In Experiment V, adopting a task specifically designed to assess the ARS, the only significant effect was a general multisensory facilitation. Remarkably, the topography of this multisensory boosting spread over the 10 positions tested, suggesting that it is not specific to the reachable space. In sum, we show that 1) PPS does not cover the entire ARS, 2) ARS is not hand-centred, and 3) ARS is not susceptible to multisensory stimulation; these last two are critical features of PPS.
These findings are consistent with what one would predict from the differences in the neural networks underlying these spatial representations. Studies on non-human primates requiring reaching movements performed with the upper limb found activations involving M1, PMv and PMd, parietal areas V6A and 5 and the parietal reach region (Andersen & Buneo, 2002; Buneo et al., 2016; Caminiti et al., 1990, 1991; Georgopoulos et al., 1982; Kalaska et al., 1983; Marconi et al., 2001; Mushiake et al., 1997; Pesaran et al., 2006; Scherberger & Andersen, 2007). In humans, tasks adopted to study ARS require judging the reachability of a target stimulus (Carello et al., 1989; Coello et al., 2008; Coello & Iwanow, 2006; Rochat & Wraga, 1997) or performing reaching movements (Battaglia-Mayer et al., 2000; Caminiti et al., 1990, 1991; Gallivan et al., 2009), involving components of motor intention, motor imagery, and motor performance. Brain activations underlying these tasks encompass M1, PMd, supplementary motor area, posterior parietal cortex and V6A as well as the anterior and medial IPS (Coello et al., 2008; Connolly et al., 2003; Desmurget et al., 1999; Lara et al., 2018; Monaco et al., 2011; Pitzalis et al., 2013; see Filimon, 2010 for review). Therefore, despite some overlap in their respective fronto-parietal circuitry, PPS and ARS networks do involve specific and distinct neuroanatomical regions (Andersen & Buneo, 2002; Brozzoli et al., 2012; Coello et al., 2008; Gentile et al., 2011, 2013; Kertzman et al., 1997; Lara et al., 2018), in keeping with the behavioural differences we reported here.
To date, behavioural evidence for hand-centred coding of PPS in humans comes from a neuropsychological single case study (di Pellegrino et al., 1997). Here, we considerably widen this notion by showing that hand-centred coding is a physiological feature of PPS in neurotypical humans. Neural multisensory processing of visuo-tactile stimuli in close body proximity may account for this hand-centred effect (Graziano & Gross, 1993; Rizzolatti et al., 1981a, 1981b). Crucially, the facilitation follows the hand: observed in close positions when the hand is close but only in far(ther) positions when the hand is distant. Because both hand positions fell within the ARS limit, this finding speaks strongly against the equivalence of PPS and ARS.
The amount of multisensory facilitation observed in Experiment I for the position closest to the trunk (V-P1, thus clearly within ARS) is remarkable in two respects: first, it is lower than that observed in correspondence to the hand-close PPS peak (between V-P2 and V-P3) and, second, it is comparable to that obtained for all the out-of-reach positions (V-P7 to V-P10), irrespectively of hand distance. Even more intriguingly, when the hand was distant, multisensory facilitation peaked farther (between V-P4 and V-P5) but was still present up to a much farther position (V-P7), which is both physically and subjectively beyond the reachable limit. Thus, compared to the ARS, the PPS extent is generally smaller, but it can cover a portion of space beyond the ARS when the arm is fully stretched and the hand is at its boundary. These additional findings converge in indicating that PPS and ARS are different spatial representations.
To the best of our knowledge, our study provides the first mapping of the hand-centred space in healthy humans, allowing to track the multisensory facilitation "shifts" following hand position shifts. Notably, this facilitation does not progressively decrease from the body to the far space, and can hardly accommodate a sigmoidal fitting (Canzoneri et al., 2012; Serino et al., 2015).
Instead, here we report that the fit of a Gaussian curve is more appropriate, filling the gap between human and non-human primates studies (Graziano et al., 1997).
At odds with some previous studies employing auditory or visual looming stimuli (Canzoneri et al., 2012; Finisguerra et al., 2015; Noel et al., 2015; Serino et al., 2015), we used ‘static’ stimuli briefly and synchronously flashed with tactile ones to avoid the risk of inflating the estimates of multisensory facilitation. In fact, looming stimuli with predictable arrival times induce foreperiod effects that, though not solely responsible for the boosting of touch, may lead to overestimations of the magnitude of the facilitation (Hobeika et al., 2020; Holmes et al., 2020; Kandula et al., 2017).
The three experiments assessing ARS with the reachability judgement task provided complementary evidence in support of our predictions. The participants slightly overestimated their limit of reachability, thus replicating a robust finding (Bootsma et al., 1992; Carello et al., 1989; Coello & Iwanow, 2006). Overestimation may relate to everyday experience when reachability judgments depend on the representation of action potential, which includes torso flexion and shoulders rotation (Rochat & Wraga, 1997). Interestingly, we add that overestimation is not modulated by hand vision (Experiments III and IV, see Supplementary Notes 3 and 4), and is independent of the position of the hand (Experiments IV and V). Moreover, in the reaching task, the general speed up of performance following multisensory with respect to unisensory stimuli, was not modulated as a function of stimulus reachability or closeness to the hand. In summary, contrary to the results of PPS mapping (Experiment I), there was no sign of hand-centred facilitation in the ARS mapping (Experiment V).
In conclusion, this study is the first to provide a clear distinction between PPS and ARS. The discrepancies concern both their spatial extent and their features and have important implications regarding the fallacy of conflating PPS and ARS. The precise evaluation of the PPS is crucial if we consider that several researchers exploit the body-part-centred nature of such a multisensory interface as an empirical entrance to the study of the bodily self (Blanke et al., 2015; Makin et al., 2008). Moreover, our results may have further implications concerning the relationships between PPS and interpersonal space, generally defined as the space that people maintain with others during social interactions. Several studies have investigated this relationship using reachability tasks and drew conclusions about the characteristics and extension of the interpersonal space (Cartaud et al., 2018; Iachini et al., 2014). The present findings make clear that the use of reachability tasks does not warrant any conclusion regarding PPS or the relationship between interpersonal and peripersonal space. They point instead to the need for investigation of the potential interactions that exist between PPS and ARS to better tune rehabilitative protocols or brain machine interface algorithms for the sensorimotor control of prosthetic arms, for which multisensory integration at multiple levels appears as a crucial factor (Makin et al., 2017; Suminski et al., 2009, 2010).
Author Contributions
C.B., A.F., A.Z. and I.P. conceived and designed the study. R.S. and E.K. developed script and devices for stimuli presentation. A.Z. performed experiments. E.B. and A.Z. analyzed data. All authors wrote and revised the manuscript.
Acknowledgements
This work was supported by the Labex/Idex (ANR-11-LABX-0042) and by grants from the James S. McDonnell Foundation, the ANR-16-CE28-0015-01 and ANR-10-IBHU-0003 to A.F. E.B. was supported by the European Union's Horizon 2020 research and innovation programme (Marie Curie Actions) under grant agreement MSCA-IF-2016-746154; a grant from MIUR (Departments of Excellence DM 11/05/2017 n. 262) to the Department of General Psychology, University of Padova. C.B. was supported by a grant from the Swedish Research Council (2015-01717) and ANR-JC (ANR-16-CE28-0008-01).
Footnotes
Conflict of Interest: The authors declare no competing financial interests.
References




