

Abstract
The increase in activity of the two-pore potassium-leak channel Kcnk5b maintains allometric juvenile growth of adult zebrafish appendages. However, it remains unknown how this channel maintains allometric growth and how its bioelectric activity is regulated to scale these anatomical structures. We show the activation of Kcnk5b is sufficient to activate several development programs, including two morphogen pathways involved in tissue formation, shh and wnt signaling. We provide in vivo transplantation evidence that the activation of developmental programs is cell autonomous and Kcnk5b can induce the expression of these developmental programs in cultured mammalian cell lines. We also demonstrate how post-translational modification of serine 345 in Kcnk5b by calcineurin regulates channel activity and controls these developmental programs to scale the fin. Thus, we show how an endogenous bioelectric program is regulated and promotes coordinated developmental signaling to generate and scale a vertebrate appendage.
Introduction
Tissue scaling involves the coordinated control of developmental programs, since anatomical structures consist of different tissues that form in a coordinated manner and grow proportionally with each other and with the body. While there are several developmental signals known to regulate cell proliferation and tissue formation, mechanisms that concomitantly activate several developmental signals to synchronize the growth of multi-tissue appendages and organs in a manner that is coordinated with body proportions remain poorly defined.
There is growing evidence that several biological phenomena involved in tissue generation and growth are influenced by electrophysiological changes in “non-excitable” cells (Sundelacruz et al., 2009). Several cell behaviors are affected by the addition of electric currents (McCaig et al., 2005): cell migration, cell proliferation, cell differentiation, gene transcription and consequently tissue formation are all altered by the application of an exogenous current (Baer and Colello, 2016; Bartel et al., 1989; Blackiston et al., 2009; Borgens et al., 1977; Geremia et al., 2007; Sundelacruz et al., 2009; Yasuda, 1974; Zhao et al., 2002). The culmination of these findings have led to the hypothesis that bioelectrical fields exist that have higher order organizational non-cell-autonomous properties in the developmental of anatomical structures [For review, see (Levin, 2014; Messerli and Graham, 2011)].
As a regulator of membrane potential, K+ conductance is an important component of the electrophysiological properties of cells. Evidence that illustrates the importance of K+ conductance in tissue formation comes from studies in which disruption of inward rectifying K+ channels of the Kir2 family can cause cranial facial defects, abnormal number of digits and reduced digit size (Andersen et al., 1971; Canun et al., 1999; Sansone et al., 1997; Tawil et al., 1994; Yoon et al., 2006a; Yoon et al., 2006b; Zaritsky et al., 2000). A striking finding concerning the coordinated control of cell behavior is the formation of eye structures by overexpressing different ion channels that alter membrane potential in early Xenopus embryos (Pai et al., 2012): overexpression and activation of a glycine-gated chloride channel in cells that form the eye interferes with eye formation, while overexpression of a dominant-negative K+-ATP channel simulates ectopic eye formation even in unexpected locations on the body (Pai et al., 2012). These findings illustrate that changes in the membrane potential of cells can have significant impacts on the development of anatomical structures. However, how electrophysiological information associates with the multiple necessary signals that control formation and/or growth of multi-tissue structures remains unclear.
The development of body structures not only involves forming tissues, it also involves coordinating the growth of each contributing tissue cell. To form organs that correctly scale with the body, each tissue grows either isometrically (grows with the same rate as the body) or allometrically (disproportionally grows in relation to the growth of the body). The zebrafish mutants another long fin (alf), long fin (lof) and schleier (schl) display continued allometric growth of each appendage from the juvenile stage into the adult stage (Lanni et al., 2019; Perathoner et al., 2014; Stewart et al., 2020). The dominant allometric growth phenotype of alf is due to mutations in the transmembrane pore region of kcnk5b (Perathoner et al., 2014), encoding a two-pore K+-leak channel that regulates membrane potential by outward flow of K+ from the cell (Goldstein et al., 2001). The dominant phenotype of lof is linked to elevated expression of a voltage-gated potassium channel Kcnh2a (Stewart et al., 2020). The phenotype of schl is due to dominant-negative mutations in scc4 (Lanni et al., 2019), encoding a K+-Cl- cotransporter that regulates intracellular K+ levels in a chloride-dependent manner (Marcoux et al., 2017). alf, lof and schl demonstrate the importance of K+ conductance and ultimately of electrophysiological signals for the correct body-to-appendage proportions. Despite the connection between K+ conductance and the proportional growth of the fins, it remains unclear how K+-mediated signal translates into coordinated growth of the fish appendage and how any K+ channel is regulated to scale tissue.
We show that activity of the single two-pore K+-leak channel Kcnk5b is sufficient to induce the activation of two important morphogen pathways (Shh and β-catenin-dependent Wnt) not only in the adult fin but also in the larva. Our data also indicates that this induction is cell autonomous, indicating that increases in membrane potential caused by Kcnk5b regulate growth through the regulation of these developmental pathways. Furthermore, we show that Kcnk5b has similar inductive effects in mammalian cells and that this phenomenon can be induced by other two-pore Kcnk channels, supporting the conclusion that the developmental programs are regulated by the same electrophysiological change induced by potassium leak. Lastly, we show how post-translational modification of Kcnk5b at Serine345 by calcineurin regulates its electrophysiological activity and consequently the scaling of zebrafish fins. Thus, we describe an endogenous cell-autonomous mechanism through which electrophysiological signals induce and coordinate morphogen and growth factor signals to mediate the scaling of an anatomical structure.
Results
Kcnk5b is sufficient to induce several developmental gene programs in different cell types in adult and larva
Mutations in the two-pore K+-leak channel Kcnk5b that increase its activity lead to enhanced growth of the zebrafish appendages (Perathoner et al., 2014). While this finding implicates the importance of bioelectric signaling in appendage scaling, it remains unknown how the activity of a single K+ channel is integrated with the developmental controls that generate new appendage tissues. Growth of any appendage involves the coordinated activation of specific morphogen and growth factor pathways: Shh, β-catenin-dependent Wnt, Bmp, Fgf and Retinoic acid. Therefore, to begin to determine how this channel is involved in the coordinated growth of the entire fin, we generated transgenic zebrafish that expresses kcnk5b under the control of conditionally inducible promoter (heat-shock promoter) to temporally activate this channel in adult fins. After a single ten-minute heat-shock pulse of the Tg[hsp70:kcnk5b-GFP] transgene, we observed significant activation of shh and lef1 (β-catenin-dependent Wnt) (Fig. 1Aa), as well as an increase in aldh1a2 (retinoic acid) and msxb (Bmp) (Fig. 1Ab) within 6 hours by qRT-PCR. The elevated expression of shh and lef1 continued 12 hours after the single pulse (Fig. 1Ba), while the other genes returned to control values or were down regulated (Fig. 1Bb). The transgenic expression of the channel emerged as a lattice pattern indicating that Kcnk5b-GFP was localized on cell membranes (Suppl. Fig. 1A-C). The induction of these developmental genes is linked to kcnk5b expression, since their up-regulation coincided with the temporal expression of the kcnk5b-GFP transgene (Suppl. Fig. 1D,E), and all genes were down-regulated by 24 hours after the single heat-shock pulse (Suppl. Fig. 1F). When we maintained chronic expression of the transgene by heat shocking the caudal fin for 10 minutes once per day for 3 days, we observed expression of lef1, shh, aldh1a2 as well as pea3 over controls (Fig. 1C). Together, these data show that Kcnk5b is sufficient to induce certain developmental programs as though it were a part of a signaling mechanism for tissue generation.

(A) qRT-PCR results of shh and lef1 expression (a) and aldh1α2, pea3 and msxb (b) from caudal fins of 6-month-old zebrafish comparing before (-HS) and 6 hours after (+HS) heat shock induction of the Tg[hsp70:kcnk5b-GFP] transgene in the caudal fins. (B) qRT-PCR results of shh and lef1 expression (a) and aldh1α2, pea3 and msxb (b) from caudal fins of 6-month-old zebrafish comparing before and 12-hours after heat-shock induction of the Tg[hsp70:kcnk5b-GFP] transgene in the caudal fins. (C) qRT-PCR results for several genes in the caudal fin from daily heat-shock pulse of Tg[hsp70:knck5b-GFP] over 3 days. (D) in situ hybridization experiments on fins show expression of shh (a,b) and lef1 (c,d) before (a,c) and after (b,d) heat-shock induction of kcnk5b-GFP. (E) Cross sections through fin rays show expression of shh (a,b) and lef1 (c,d) before (a,c) and after (b,d) heat-shock induction of kcnk5b-GFP. (F) Representative images of Western blots show expression of Shh, Lef1 and β-catenin before and 3 days after 10-minute daily heat-shock induction of kcnk5b-GFP in the fin. (G) Graphed measurements results of Western blots. (H) Immunohistochemistry stainings for (Kcnk5b-GFP) GFP (a,b: green), β-catenin (c,d: red) and DAPI (e,f: blue) of fin cross sections of transgenic kcnk5b-GFP animals without heat shock (a,c,e,g,i) or after heat shock (b,d,f,h,j). White boxes in g and h show location of magnified panels of i and j. Overlapping DAPI and β-catenin staining indicated by white arrows. (I) Graphed measurements of DAPI stained nuclei containing staining of β-catenin. (J) Graphed measurements of β-catenin fluorescence intensity of stained sections. Scale bars are 50 µm (D), 1 mm (E), 10 µm (H). The data for each experiment represent an N of 3 or more, which each N having 2 or more replicates. Student’s T-test used for the tests of significance between indicated experimental groups.
To examine the spatial expression of the two genes most responsive to Kcnk5b, we performed in situ hybridization experiments for shh and lef1. We observed the localization of shh and lef1 in the distal tip of the fin (Fig. 1Da-d) were growth normally occurs, and cross sections through the fins showed that these genes are expressed in overlapping patterns in the epidermal/dermal tissues (Fig. 1Ea-d). We also assessed increases in Shh and Lef1 protein levels after the heat-shock induction of kcnk5b-GFP (Fig. 1F,G). Because Lef1 conveys β-catenin-dependent Wnt signaling by acting as transcriptional platform for β-catenin, we examined protein expression of β-catenin and observed no significant differences in its overall levels (Fig. 1F,G). However, when we examined β-catenin protein distribution in the fin tissues by immunohistochemistry staining, we observed an increased number of nuclei with β-catenin co-staining in the dermal and mesenchyme tissues of the fin (Fig. 1H,I) despite no significant differences in the measured β-catenin-associated fluorescence intensities (Fig. 1J). Together, these results indicate that increased expression of kcnk5b is sufficient to activate the developmental signaling programs involved in generation of new tissue. They also suggest a direct relationship between K+ conductance and the transcriptional regulation of at least two important morphogen pathways.
To test whether increasing the activity of Kcnk5b has the same transcriptional effect on these developmental pathways in another in vivo context, we induced the expression of kcnk5b in the zebrafish larva. We observed that lef1 and shh expression increased, while the other genes representing the other pathways did not (Fig. 2A). We confirmed the increase in β-catenin-dependent Wnt signaling by crossing the heat-shock-inducible transgenic Tg[hsp70:knck5b-GFP] line with the β-catenin-dependent Wnt transgenic reporter line Tg[7xTCFsamois:mCherry]. While the double-transgenic fish Tg[hsp70:kcnk5b-GFP; 7xTCFsamois:mCherry] displayed limited expression of mCherry before heat-shock induction of the kcnk5b-GFP transgene (Fig. 2Ba,b,g), after heat shock, double-transgenic fish showed a broad increase in reporter mCherry expression (Fig. 2Bd,e,g). From histological cross sections of double-transgenic larva, we observed that mCherry was upregulated broadly in the body of the animal (Fig. 2C). Furthermore, we observe an increase in the proportional growth of the dorsal-to-ventral dimensions of the caudal finfold form induction of kcnk5b-GFP by one daily 10-minute pulse for 3 days (Fig. 2D), showing a functional effect on growth by Kcnk5b. Together, these results indicate that the increased Kcnk5b activity is sufficient to promote shh and β-catenin-dependent Wnt signaling in several different tissue types.

(A) Comparison of gene expression from zebrafish larva harboring Tg[hsp70:knck5b-GFP] before and 6 hours after heat-shock (HS) induction. (B) Double transgenic fish harboring Tg[hsp70:kcn5b-GFP] and Tg[7xTCFsamois:mCherry] either before heat shock (a-c) or 12 hours after heat shock (d-f). (Bg) Measurements of mCherry intensity levels of non-transgenic (non-tg), kcnk5b-transgenic (kcnk5b) fish and transgenic fish harboring the β-catenin-dependent Wnt 7xTCFsamois:mCherry reporter (7xTCF) before and 12 hours after heat shock. (C) Cross sections through the trunks of non-transgenic (a-d) and single- (e-h) and double-transgenic (i-l) fish lines after heat shock. (D) Measurements from dorsal to ventral of the caudal finfold of Tg[7xTCFsamois:mCherry] and Tg[7xTCFsamois:mCherry]X Tg[hsp70:kcnk5b] sibling larva after heat shock. Scale bars are 100µm (B) and 20µm (C). The data for each experiment represent an N of 3 or more, which each N having 3 or more replicates. Student’s T-test used for all test of significance between the indicated experimental groups.
Activation of β-catenin-dependent Wnt reporter by Kcnk5b is cell autonomous in several different tissues
Previous work implicates bioelectric intercellular communication as a mechanism for how bioelectricity can influence tissue growth (McLaughlin and Levin, 2018), and changes in K+ channel activity have been shown to regulate different cell behaviors in a non-cell-autonomous manner (Morokuma et al., 2008; Pai et al., 2015). The broad activation of the β-catenin-dependent Wnt reporter in several tissues (Fig. 2) and Kcnk5b’s ability to scale all the tissues of the fin appendages suggest that Kcnk5b acts via non-cell autonomous communication among cells. To determine whether the observed Kcnk5b-mediated induction of gene expression is due to intercellular communication (e.g., through extracellular ligands such as Wnt) or due to cell autonomous activation of transcription, we transplanted cells from Tg[hsp70:kcnk5b-GFP; 7xTCFsamois:mCherry] transgenic embryos into embryos harboring only the Tg[7xTCFsamois:mCherry] transgene and then raised mosaic embryos as larva (Fig. 3A). Analyses of the mosaic larva showed the reported developmental expression of the β-catenin-dependent Wnt reporter before heat shock (Fig. 3Ba,e,i,m)(Moro et al., 2012). However, after heat-shock induction of the Tg[hsp70:kcnk5b-GFP] transgene (Fig. 3Bb,f,j,n), we observed ectopic activation of 7xTCFsamois:mCherry reporter in all chimeric embryos. GFP-mCherry-positive cells appeared in tissues in the head (Fig. 3Bb,c,d), in skeleton surrounding the eye (Fig. 3Bf,g,h), in trunk muscles (Fig. 3Bj,k,l) and in skin (Fig. 3Bn,o,p). From closer inspection, we observed co-expression in neurons in the head (Fig. 3Ca-c), in the ectodermal bones of the skull (Fig. 3Cd-f), mandible bone and cartilage (Fig. 3Cg-i), mesenchyme surrounding the otic vesicle (Fig. 3Cj-l), epithelial cells in the finfold (Fig 3Co-r) and individual striated muscle cells of the trunk (Fig. 3Cr-t). We counted the number of GFP and mCherry positive cells in the different tissues and observed that all Kcnk5b-GFP-positive cells were mCherry positive (Fig. 3C). Moreover, in all tissues, the ectopic mCherry expression was always limited to the Kcnk5b-positive cells (Fig. 3B-D). Together, these data support two conclusions: one, the activation of the β-catenin-dependent reporter by Kcnk5b is cell autonomous; and two, Kcnk5b is able to promote the expression of the Wnt reporter in diverse tissue types.

(A) Diagram of transplantation procedure and possible cell-autonomous and non-cell-autonomous outcomes on the expression of the 7xTCFsamois:mCherry reporter after heat-shock induction of the kcnk5b-GFP transgene. (B) Transplantation experiments of donor cells from double transgenic fish harboring Tg[hsp70:kcnk5b-GFP] and Tg[7xTCFsamois:mCherry] into host embryos harboring only the Tg[7xTCFsamois:mCherry]. The head (a) eye (e) trunk and finfold (m) of mosaic larva before heat shock induction of kcnk5b-GFP expression. Head (b-d), eye (f-h), trunk (j-l) and finfold (n-p) 24 hours after heat shock. (C) Bright field images of the head (a,d), jaw area (g) border tissue of otic vesicle (j) and trunk (o,r); kcnk5b-GFP expression (b,e,h,k,p,s) and 7xTCFsamois: mCherry expression (c,f,i,l,q,t). (D) Total number of positive cells counted in the tissues of all mosaic larva for all mCherry-positive cells (open red circle), all kcnk5b-GFP-positive cells (open green triangle) and all double positive cells (open blue squares).
As a K+-leak channel, Kcnk5b’s activity should decreases intracellular K+ levels. We performed Fluorescence Lifetime Microscopy (FLIM) analysis with an established genetic sensor for K+ to measure intracellular K+ levels (Shen et al., 2019). This sensor uses the FRET potential between two fluorophores that are joined by a K+-binding linker. Changes in FRET due to K+ binding results in changes in the fluorescence lifetime of the fluorophores, which allows for the assessment of intracellular K+ levels. Transfection of the channel in Human Embryonic Kidney HEK293T cells (Suppl. Fig. 3A-L) resulted in significant increase in CFP fluorescence lifetime due to decreased FRET of the sensor compared to control transfected cells (Fig. 4Aa-c,g), which indicated reduced intracellular K+ levels in the cells that express Kcnk5b (Fig. 4Ad-g). Additional higher resolution assessments along the lateral borders of cells showed similar increases in CFP fluorescence lifetime along the plasma membrane, indicating expected reduction of K+ levels at the cell membrane by active Kcnk5b (Suppl. Fig. 3M-O).

(A) FRET-FLIM images after measuring the life time of CFP of the K+ FRET reporter KIRIN (Shen et al., 2019). The color images indicate the differences in CFP fluorescence lifetime of the K+ FRET reporter KIRIN in HEK293T (HEK) cells. Assigned rainbow of colors in the delineated cytoplasm depict the range of numeric values of nanoseconds (ns) of the detected fluorescent lifetime for CFP. Red represents longer lifetime values. Blue represents shorter lifetime values, and the other colors represent intermediary lifetime values. (a) Composite image of all lifetime values of the KIRIN K+ reporter in control cells transfected with mCherry. (b) Image of low lifetime values in a control cell. (c) Image of high lifetime values in a control cell. (d) Composite image of all lifetime values of the KIRIN K+ reporter in cells expressing kcnk5b-mCherry. (e) Image of low lifetime values in cells expressing kcnk5b-mCherry. (f) Image of high lifetime values in cells expressing kcnk5b-mCherry. (g) Compared to GFP-transfected HEK cells, cells transfected with kcnk5b-mCherry show an increase in CFP lifetime due to reduction in intracellular K+. (Ba) qRT-PCR for SHH and LEF1 in HEK cells. (Bb) qRT-PCR for ALDH1a2, PEA3 and MSX1 in HEK cells. (C) qRT-PCR for indicated genes in HEK cells expressing GFP, kcnk9-GFP or kcnk10-GFP 24 hours after transfection. (D) qRT-PCR results in HeLa cells expressing either GFP or kcnk5b-GFP 24 hours after transfection. (E,F) qRT-PCR results in N2A cells expressing either GFP or kcnk5b-GFP 24 hours after transfection. (G) qRT-PCR results in Mcf7 cells expressing either GFP or kcnk5b-GFP 24 hours after transfection. Student’s T-test was used for tests of significance and the levels of significance are indicated between the experimental groups.
To test whether activity kcnk5b promotes the gene expression profile in mammalian cells that we observed in the zebrafish, we established stable HEK293T (HEK) cells lines that either express GFP or zebrafish kcnk5b-GFP. From qRT-PCR analyses comparing HEK cells expressing either GFP or kcnk5b-GFP, we observed an increase in SHH and PEA3 expression (Fig. 4Ba) and the down-regulation of LEF1, ALDH1a2 and MSX1 (Fig. 4Bb). To determine whether this transcriptional response is specific to Kcnk5b or is a general response to two-pore K+-leak channels, we transfected cells with one of two K+-leak channels Kcnk9 and Kcnk10 (Suppl. Fig 4A,B). Transfection of HEK cells with these two other channels resulted in the same transcriptional profile as Kcnk5b (Fig. 4C), indicating that the transcriptional response to Kcnk5b is a response to the electrophysiological changes associated with intracellular K+ leak.
The difference between the transcriptional responses of the zebrafish adult, larva and HEK cells indicates that different cell types will have different responses to Kcnk5b electrophysiological activity. Therefore, we examined the transcriptional responses to Kcnk5b in other mammalian cell lines. In HeLa cells, Kcnk5b induced PEA3 and LEF (Fig. 4D). From the N2A (neural carcinoma) cell line, we observed the increase of ALDH1a2 (Fig. 4E) but decreases in SHH, LEF1 and PEA3 (Fig. 4F). In the MYF7 epithelial carcinoma cell line, Kcnk5b induced ALDH1a2, PEA3 and MSX1 (Fig. 4G) We propose that the variability in genes that are transcribed may explain why the solitary change in the activity of this channel in all cells of the fin leads to the variable transcriptional responses needed to promote coordinated growth of a multi-tissue anatomical structure. These results also reveal that Kcnk channel activity is sufficient to induce the transcription of different developmental pathways in different mammalian cells types and that K+ conductance-mediated induction of developmental transcription is shared among vertebrate species.
Calcineurin regulates Kcnk5b channel activity and Kcnk5b-mediated gene transcription
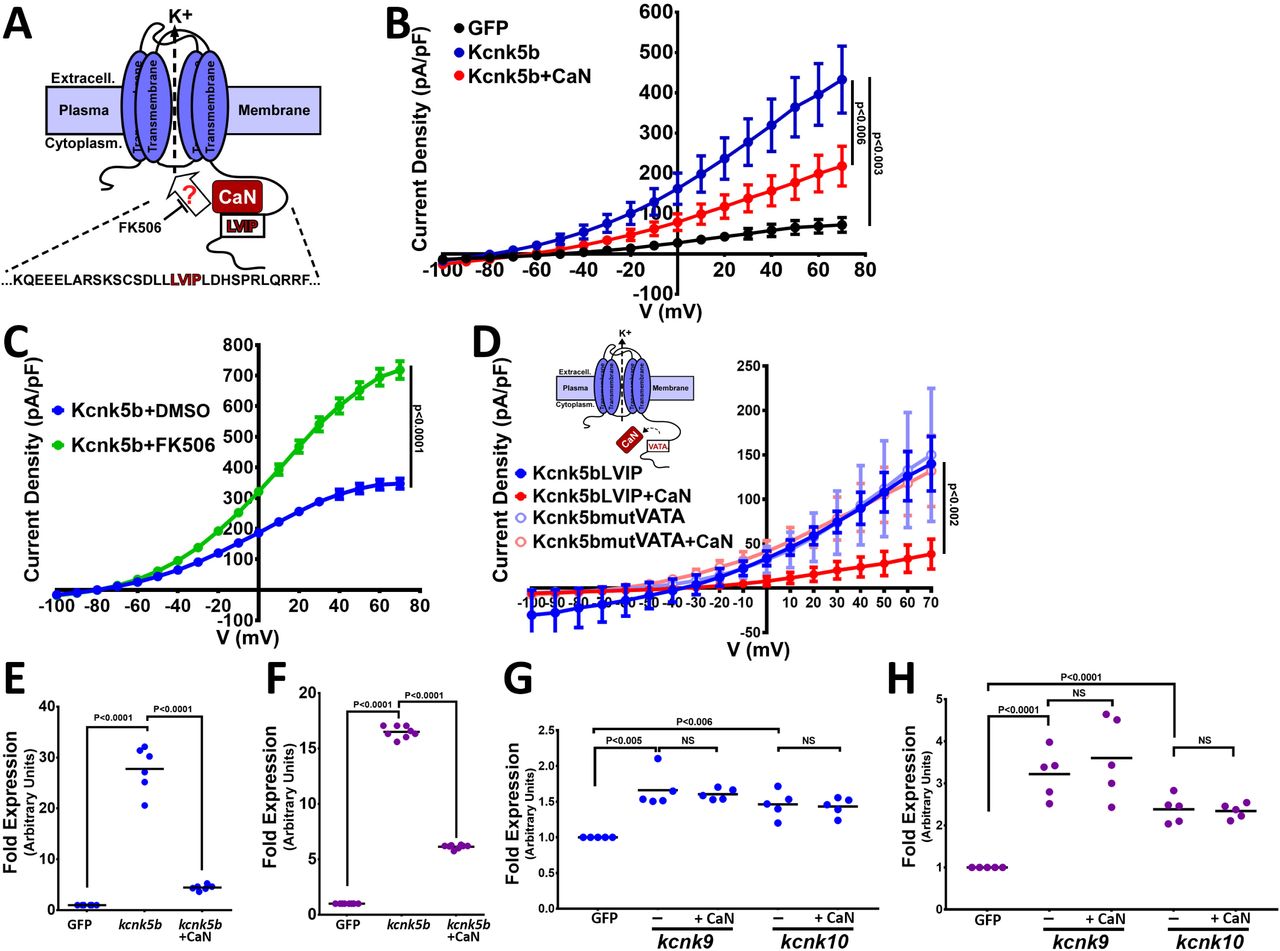
We previously showed that the phosphatase calcineurin acts as a molecular switch between isometric and allometric proportional growth of the zebrafish fins (Kujawski et al., 2014). The similarities in the phenotypes produced by calcineurin inhibition and by the mutations in Knck5b that enhance Kcnk5b channel activity suggest a direct functional relationship between them (Kujawski et al., 2014; Perathoner et al., 2014). Based on whole-cell patch-clamp experiments, the mutations in kcnk5b that maintain allometric growth of the fins also increased K+ conductance at the plasma membrane (Perathoner et al., 2014). Therefore, we hypothesized that calcineurin inhibition will increase in vivo K+ conductance and promote allometric growth of the zebrafish fins (Fig. 5A). To test whether calcineurin alters the channel activity of Kcnk5b, we examined whether the activity of Kcnk5b is altered by calcineurin. Comparison of whole-cell patch-clamp measurements using HEK cells showed that Kcnk5b expression increases current density due to K+ leak from the cells (Fig. 5B), as we observed from FRET-FLIM intracellular K+ measurements (Fig. 4A). However, cells co-expressing both knck5b and calcineurin decreased the K+ conductance of the cells compared to cells expressing kcnk5b alone (Fig. 5B). We then tested whether inhibition of the endogenous calcineurin activity in the HEK cells by the calcineurin inhibitor FK506 affects Kcnk5b channel activity. We found that FK506 treatment of cells expressing kcnk5b resulted in a significant increase in K+ current compared with DMSO-treated kcnk5b-expressing cells (Fig. 5C). These results show that changes in calcineurin activity alter Kcnk5b channel activity in a manner that is constant with the enhanced fin growth induced by calcineurin inhibition and the increased channel activity of the kcnk5b zebrafish mutants, which indicates a functional interaction between calcineurin and Kcnk5b.

(A) Diagram of hypothetical interaction between Calcineurin (CaN) and Kcnk5b at a consensus calcineurin binding site (LVIP) in Kcnk5b. (B) Whole-cell patch clamp of HEK293T (HEK) cells expressing the indicated zebrafish proteins: Calcineurin-mCherry and Kcnk5b-GFP. (C) Whole-cell patch-clamp results of cells expressing zebrafish Kcnk5b-GFP and treated either with DMSO or the calcineurin inhibitor FK506. (D) Diagram shows mutant Kcnk5b with altered amino acids at putative calcineurin binding site and graph of the Patch-clamp results of the wild-type zebrafish Kcnk5b channel (Kcnk5bLVIP) or mutant Kcnk5b (Kcnk5bmutVATA) lacking the putative calcineurin binding site. Each construct is expressed either with or without calcineurin (CaN). (E) qRT-PCR for SHH in HEK cell lines stably expressing either GFP or Kcnk5b-GFP and transfection with calcineurin-mCherry (CaN). (F) qRT-PCR for PEA3 in HEK cell lines stably expressing either GFP or Kcnk5b-GFP and transfection with calcineurin-mCherry (CaN). (G) qRT-PCR of SHH expression HEK cells after transfection either with GFP, kcnk9-GFP or kcnk10-GFP with or without calcineurin. (H) qRT-PCR of PEA3 expression HEK cells after transfection either with GFP, kcnk9-GFP or kcnk10-GFP with or without calcineurin. The electrophysiology measurements (B-D) are averages with SEM. (E-G) Graph panels show averages. The data for each experiment represent an N of 3 or more, which each N having 2 or more replicates. Student’s T test was used to determine the indicated significance (P) values.
Calcineurin interacts with its substrates at particular amino acid sequence sites (Grigoriu et al., 2013). Our analysis of the amino acid sequence in the C-terminal cytoplasmic tail of Kcnk5b suggests a functional calcineurin binding site (LVIP) is present (Fig. 5A, red letters). To test for functional interaction at this site, we mutated the amino acid sequence (Fig. 5D, red letters) and assessed how the mutation affected the ability of calcineurin to regulate the channel. Compared to the decrease in activity of the wild-type channel after co-transfection with calcineurin, co-transfection of the Kcnk5b mutant lacking the calcineurin binding site with calcineurin (Kcnk5bmut+CaN) showed that the mutation made the channel resistant to calcineurin-mediated inhibition (Fig. 5D). The resistance of the Kcnk5bmut to calcineurin indicated that the repression of channel activity on Kcnk5b by calcineurin is due to the interaction of these to proteins at the LVIP site.
The regulation of Kcnk5b by calcineurin suggests that changes in calcineurin activity will have an effect on the Kcnk5b-dependent gene expression. To assess whether the activation of SHH by Knck5b can be altered by calcineurin, we compared the expression of SHH between HEK cells stably expressing GFP and HEK cells stably expressing the Kcnk5b channel as well as between HEK cells stably expressing the channel after transfection with calcineurin. We observed that compared to channel expression alone, co-expression of calcineurin decreased the Kcnk5b-mediated induction of SHH (Fig. 5E) and PEA3 (Fig. 5F). To determine whether calcineurin effect on Kcnk5b-medidated SHH expression is specific to Kcnk5b, we transfected HEK cells with Kcnk9 or Kcnk10. Both Kcnk9 and Kcnk10 lack identifiable calcineurin binding sites (Suppl. Fig. 4A,B), and we observed that unlike the effect on Kcnk5b, calcineurin had no effect on the induction of SHH (Fig. 5G) or PEA3 (Fig. 5H) by Kcnk9 or by Kcnk10, indicating that calcineurin’s regulation of the electrophysiological induction of SHH and PEA3 transcription is specific to Kcnk5b.
Calcineurin regulates Kcnk5b through S345 to scale the fin
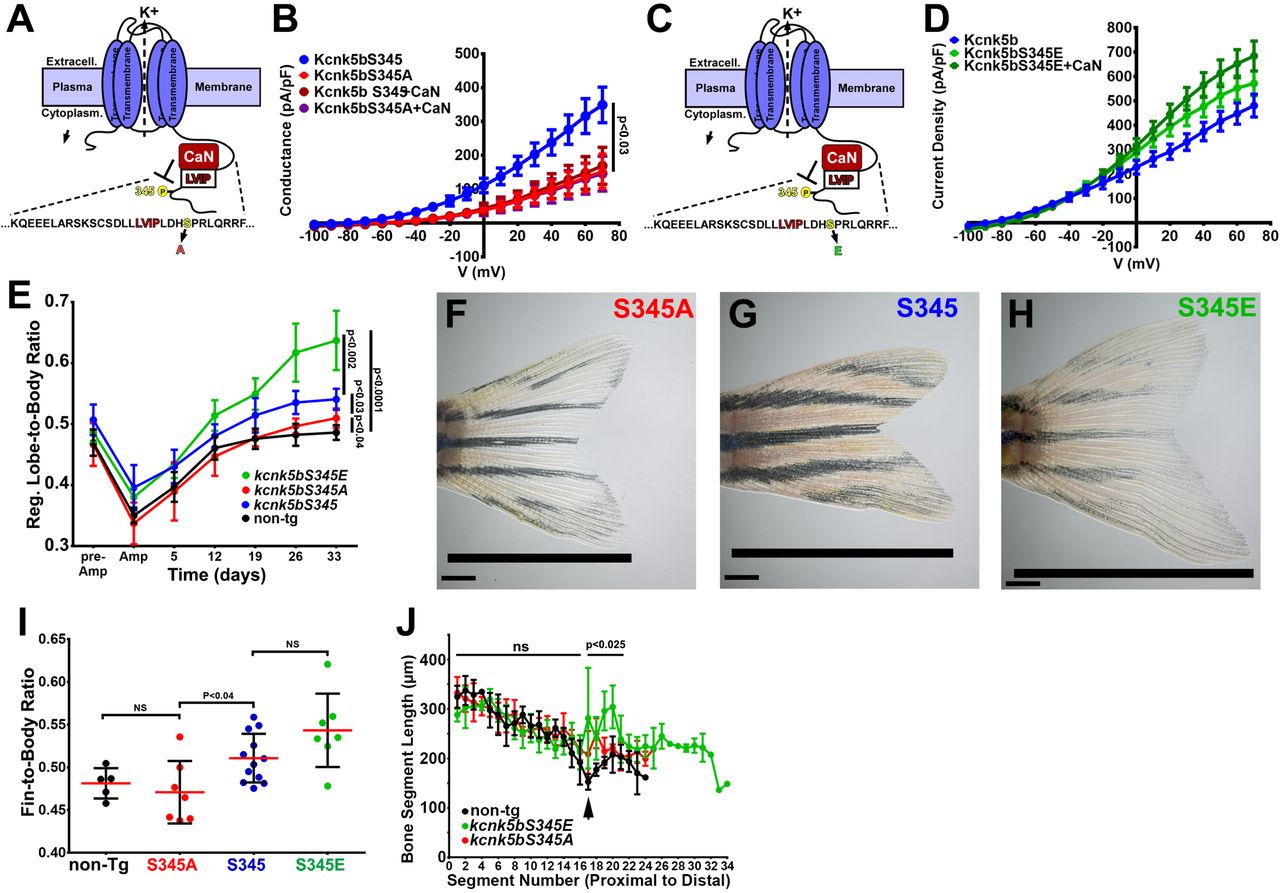
As a phosphatase, calcineurin should regulate Kcnk5b by dephosphorylating the channel at specific serine or threonine residues. A specific serine in the C-terminal tail represented a typical consensus serine-proline (Ser345-Pro346) phosphorylation site for calcineurin (Fig. 6A). Therefore, we hypothesized that calcineurin inhibits the activity the Kcnk5b channel by dephosphorylating this serine. We tested whether rendering the Kcnk5b channel unphosphorylatable at this serine by alanine substitution (S345A) would decrease the channel’s activity. Whole-cell patch-clamp experiments of the kcnk5bS345A showed a significant decrease in K+ conductance of the channel compared to the wild-type (kcnk5bS345) control (Fig. 6B). To assess the specificity of the reduction effect for this serine, we also systematically substituted adjacent serines with alanines and subsequently measured channel activity (Suppl. Fig. 5A). While kcnk5bS345A showed reduction in activity, the substitution of other serines did not (Fig. 6C, Suppl. Fig. 5A-D).

(A) Diagram of Kcnk5b channel showing proposed Serine345Proline346 calcineurin dephosphorylation site adjacent the calcineurin-interaction site (LVIP). Mutation of S345 to alanine (A) mimics dephosphorylation. (B) Whole-cell patch-clamp results of HEK239T (HEK) cells transfected with zebrafish wild-type channel (Kcnk5bS345) or the dephospho-mimic mutant (KcnkS345A) either with or without calcineurin (CaN). (C) Diagram of serine (S) to glutamic acid (E) substitution to mimic phosphorylation of Kcnk5b. (D) Whole-cell patch-clamp measurements for wild-type Kcnk5b and mutant Kcnk5b harboring a Serine345 to glutamic acid either with or without calcineurin (CaN). (E) Graph displays different growth rates of the regenerating caudal fin lobes of the indicated transgenic fish lines. Body length of each fish was used to standardize the fin length measurements (fin-to-body ratio). (F) Caudal fin of Tg[hsp70:kcnkbS345A] transgenic fish after regeneration of ventral lobe. (G) Caudal fin of Tg[hsp70:kcnk5bS345] transgenic fish after regeneration of ventral lobe. (H) Caudal fin of Tg[hsp70:kcnk5bS345E] transgenic fish after regeneration of ventral lobe. (I) Graph of fin-to-body ratios of the unamputated lobes of the indicated transgenic fish lines at 33 days of the same fish as in (F). (J) Graph of the measured bone segment lengths of the unamputated caudal fin lobes from non-transgenic, Tg[hsp70:kcnk5bS345EA], and Tg[hsp70:kcnk5bS345E] fish. The measurements of each segment extend from the proximal base of the fin (segment 0) to distal tip of the lobe (segment 34). p<0.025 value is between Tg[hsp70:kcnk5bS345E] and non-Tg fish. The data for each experiment represent an N of 3 or more, which each N having 3 or more replicates. The electrophysiology measurements (panels B,D) are represented as averages with SEM. Significance values shown in the graphs were measured by students t-tests (E, I, J) are represented as averages and SD. The scale bars equal 2 mm (F-H).
To determine whether the activity of the Kcnk5b channel is associated with the phosphorylation state of this serine, we exchanged the serine for a glutamic acid to mimic serine phosphorylation (kcnk5bS345E) (Fig. 6C). Expression of this mutant displayed elevated K+ conductance compared to knck5bS345 wildtype channel (Fig. 6D). Moreover, the kcnk5bS345E mutant was resistant to calcineurin-mediated inhibition (Fig. 6D), while substitution of other serines with glutamic acid not only had no effect on channel activity, calcineurin could still regulated the channel (Suppl. Fig. 5A,E,F). Together, these results indicate that S345 is the important post-translational regulatory serine involved in calcineurin-mediated regulation of Kcnk5b activity.
To determine whether there is a functional relationship between fin scaling and S345-mediated Kcnk5b channel activity, we placed the cDNA of each channel version (wild-type kcnk5bS345, kcnk5bS345E or kcnk5bS345A) under the control of the heat-shock inducible hsp70 promoter to generate conditionally inducible transgenes for in vivo expression in the fish. We induced the expression of each transgene by heat shocking the caudal fins once daily and subsequently measured the length of the fin in relation to the length of the body (fin-to-body ratio). After 12 days of the heat-shock regimen, we noticed differences between the rates of the regenerating caudal fins lobes of the different transgenic lines (Fig. 6E) after standardizing the length of each fin to the length of the body (Suppl. Fig. 5G). By assessing regenerating lobe-to-body measurements over time, we observed that Tg[hsp70:kcnk5bS345E] fish maintained the highest rates of allometric regenerative growth, while Tg[hsp70:kcnk5bS345A] displayed the lowest growth rates of the transgenic lines (Fig. 6E). There was no significant change in the rates of growth of the bodies (Suppl. Fig. 5G). We also assess the final proportional size of unamputated fin lobes between the different transgenic lines, and we observed a linear relationship between the proportional length of the unamputated lobes and the phosphorylation status of the channels: the S345A dephosphorylation mimic displayed the smallest growth proportions (Fig. 6I), the S345E phosphorylation mimic displayed the largest growth proportions (Fig. 6I), and the average value of the wild-type regulatable version of the channel was between the highly active S345E and marginally active S345A mutants (Fig. 6I). In addition to allometric growth, the genomic kcnk5b mutant another long fin (alf) also displays disruption of the normal bone segmentation pattern (Perathoner et al., 2014). To determine the extent that the different serine mutants phenocopy the segmentation defect of alf, we measured each bone segment from the base to the distal tip of the fin of the regenerating lobes heat-shocked fish, and we observed that the extent of segmentation defects was directly related to the activity of the channel: overexpression of kcnk5bS345E displayed a greater increase in segment length than overexpression of kcnk5bS345A (Fig. 6J). These results directly link post-translational regulation of channel activity with the degree of allometric growth and bone segmentation of the fin. Our ability to control the rate of growth and bone segmentation by mimicking a specific post-translational modification that can be mediated by calcineurin and that correspondingly determines the level of Kcnk5b activity supports the conclusion that calcineurin regulation of Kcnk5b is an in vivo electrophysiological mechanism through which controlling the potassium conductance of cells scales a vertebrate appendage.
Discussion
Anatomical structures consist of a combination of different tissue types that develop and grow in a coordinated manner. Recent discoveries show that K+ channels regulate the scaling of fish appendages, but it is still unclear how this electrophysiological signal controls several diverse developmental phenomena within this anatomical structure to achieve coordinated developmental growth. Our results reveal that this in vivo electrical signal to induce multiple important developmental programs, namely Shh and Lef1-mediated Wnt signaling, in the fish fins and larva to scale a vertebrate appendage.
Two-pore K+-leak channels such as Kcnk5b allow K+ to cross the membrane to establish an electrochemical equilibrium, this activity directly affects the membrane potential of the cell (Goldstein et al., 2001). Normally, the concentration of K+ is higher on the cytoplasmic side of the plasma membrane due to continual active transport of K+ into the cell by the ATP-dependent Na+/K+ pumps (Shattock et al., 2015). As a leak channel, opening of Kcnk5b causes a flow of K+ out of the cell, which hyperpolarizes the membrane potential (Goldstein et al., 2001). The finding that mutations that increase Kcnk5b channel activity maintaining allometric growth (Perathoner et al., 2014) argue that such changes in membrane potential promote disproportional growth. Our findings that conditional induction of Kcnk channel activity is sufficient to induce morphogen pathways (Fig. 1,2,4) in different in vivo and in vitro contexts furthers these original findings by demonstrating transcriptional control of developmental programs by different two-pore K+-leak channels.
In addition to K+-leak channels, cells regulate intracellular K+ through different channels and exchangers. Inward rectifying K+ channels allow K+ to enter the cell along the ion’s electrochemical gradient. Exchangers will exchange K+ with different substrates (e.g., Na+) to facilitate the entry or removal of K+ based on the concentration gradient of K+ and the exchanged substrate. Previous findings show the importance of the inward rectifying K+ channel Kir2 for cranial-facial and digit defects in humans (Andersen et al., 1971; Canun et al., 1999; Sansone et al., 1997; Tawil et al., 1994; Yoon et al., 2006a; Yoon et al., 2006b). Knockout of the mouse Kir2 channels results in similar head and digit defects (Zaritsky et al., 2000), and dominant-negative inhibition of the Drosophila Kir2 leads to wing appendage defects that are analogous to the human and mouse appendage defects (Dahal et al., 2012). While the mammalian phenotypes remain unexplained, the defects in the Drosophila wings have been linked to reduced Dpp (BMP) signaling (Dahal et al., 2012), suggesting that intracellular K+ homeostasis is important for BMP signaling.
Removal of an ATP-sensitive K+ channel in the early Xenopus embryo disrupts eye formation, while ectopic expression of this channel will produce ectopic eyes in the head and in locations that were not considered to be competent for producing eyes (Pai et al., 2012). The ability to ectopically generate eyes was linked to electrophysiological hyperpolarization of the cells and the activation of Pax6-eyeless gene (Pai et al., 2012), a master regulator for eye development (Chow et al., 1999; Halder et al., 1995). In planaria, shortly after wounding, membrane depolarization acts as an early anterior signal that is sufficient (even when induced on the posterior side) to promote the consequent formation of all the anterior structures of the planarian head by inducing notum expression, which inhibits β-catenin-dependent Wnt signal transduction (Durant et al., 2019). These discoveries show that electrophysiological changes are important signals in the formation and growth of anatomical structures.
Our findings help explain how electrophysiological changes in cells can lead to broader tissue organizing phenomena by showing the inductive effect that increasing K+ conductance can have on a broad number of developmental pathways, which is important for coordinating the organized formation of tissues and organs. Furthermore, the effect of the activity of this channel is broader than the traditional mechanisms of growth factor/morphogen signaling pathways, because it is not confined to specific signal transduction mechanisms; rather, it has variable broad effects, that is activation of several developmental signals in the adult fin (Fig. 1A) and the larva (Fig. 2A,B). We propose that the competence to activate different developmental pathways by electrophysiological changes is because the responding cells are either primed to activate them or the pathways are already active. It will be important to find out how this electrophysiological signal coordinates the activity of these developmental signals. In this regard, only few factors are known that regulate shh and lef1 transcription. Thus, our finding that an electrophysiological mechanism is involved not only provides a new understanding of how electrophysiology acts as an inductive signal, it also may lead to the discovery of molecular mechanisms that control the expression of these mediators of important morphogen signals.
The scaling activity of Kcnk5b includes all the tissues of the entire appendages of the fish (Perathoner et al., 2014). Previous findings implicate broader intercellular electrophysiological gradients as a mechanism for tissue growth (Adams and Levin, 2013). Electrophysiological measurements of animal tissues show that electric fields are generated and are important in vivo (Borges et al., 1979; Jenkins et al., 1996; McGinnis and Jr., 1986), which suggests the existence of in vivo bioelectric information that regulates physiological phenomena. However, from our transplantation experiments, we observe that the activation of the growth program is cell autonomous (Fig. 3). Consequently, the question arises about how the activity of this K+-leak channel relates to a broad, coordinated phenotype of scaling the several tissues of the fin. An answer is that the autonomous transcriptional program includes morphogens. What is unclear is whether a limited number of cells in the fin control the growth and organizing information so that Kcnk5b only needs to act on a limited number of cell types, or whether Kcnk5b regulates proportional growth at multiple levels and that the cell autonomous transcriptional response that we observe is one outcome of a combination of intracellular and intercellular responses induced by Kcnk5b.
Changes in membrane potential from alterations in K+ conductance are also associated with the progression through the cell cycle (Blackiston et al., 2009; Urrego et al., 2014), because K+ channel activity increases at specific cell cycle phases (Urrego et al., 2014), and inhibition of K+ channel activity leads to cell cycle arrest in many different tissue cell types (Blackiston et al., 2009). It is possible that this phenomenon explains part of Kcnk5b’s ability to promote allometric growth. We do not yet know whether other phenomena linked to the activity of mammalian Kcnk5 [influence cell tonicity (Niemeyer et al., 2010), metabolic acidosis and alkalinization (Warth et al., 2004), CO2/O2 chemosensing in retrotrapezoid nucleus neurons (Flores et al., 2011) and apoptosis in lymphocytes and neurons (Göb et al., 2015; Nam et al., 2011)] are involved in appendage scaling.
We previously showed that calcineurin inhibition shifts isometric growth to allometric growth (Kujawski et al., 2014). Subsequently, Daane et al. showed that this effect is reversible in that removal of calcineurin inhibitors restores isometric growth (Daane et al., 2018). Together, these data implicate calcineurin as a molecular switch governing isometric versus allometric growth control. Our findings provide a mechanism for how this switch acts to scale the fish appendages by directly regulating the activity of Kcnk5b through the dephosphorylation and phosphorylation of a specific serine (Fig. 7). The ability to mimic or block calcineurin regulation of this K+-leak channel (Fig. 5), whose activity levels directly translate into the extent of allometric growth (Fig. 6), defines how calcineurin inhibition expands clonal populations during fin regeneration (Tornini et al., 2016). However, as we observed from both calcineurin inhibition (Kujawski et al., 2014) and from conditionally inducing Kcnk5b activity (Fig. 5), the induced allometric growth of the entire fin is more than expanding clonal populations, since the outcome is not tumorigenesis. Instead, the growth is coordinated among all the tissues (Fig. 6G-I) (Kujawski et al., 2014; Perathoner et al., 2014), and our finding that Kcnk5b activates several developmental pathways (Figs. 1,2,4) argues that calcineurin activity acting on Kcnk5b regulates more than cell cycle progression.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Kcnk5b activation results in reduced cytoplasmic K+, which is sufficient to induce the transcription of the shh ligand and lef1 transcription factor (β-catenin-dependent Wnt signaling) as well as components of other developmental pathways to induce coordinated allometric growth of the tissues of the fish fin appendage. Scaling information from the body or local tissues in the fin activate calcineurin so that it dephosphorylates Kcnk5b on S345 to reduce its K+-channel activity, which results in isometric growth of the fin.
An important next step is to learn how the calcineurin-Kcnk5b circuit is integrated into the broader mechanisms that scale the appendages. Calcineurin is a Ca2+-dependent enzyme which suggests that intracellular Ca2+ is involved in scaling information. Ca2+ is a broad second messenger that can activate several downstream Ca2+-dependent enzymes, so broad changes in its subcellular levels likely have multiple effects. It remains unclear whether there is a specific intracellular distribution pattern that leads to calcineurin-mediated control of scaling. It is also possible that Ca2+-mediated activation of calcineurin—and consequent restoration of isometric growth—is so dominant that other Ca2+-mediated activities have little effect.
Two mechanisms that regulate proportional growth of organs are vitamin D and Hippo signaling. Increasing vitamin D signaling enhance the growth of the entire body, including the fins (Han et al., 2019). We propose that vitamin D is a systemic body signal that ultimately leads to the increase in Kcnk5b signaling. It is also possible that this hormone acts independently of Kcnk5b. In Drosophila, the Hippo pathway regulates brain size and size of the imaginal discs (Poon et al., 2016; Rogulja et al., 2008). Mice overexpressing a nuclear version of the Hippo-signaling component Yap1 in the adult liver develop significantly enlarged livers (Camargo et al., 2007; Dong et al., 2007). The Hippo signal transduction pathway consists of several core components that can be regulated by different factors at plasma membrane and within the cell (Yu and Guan, 2013), so there are several possible nodes of interaction between of Kcnk5b and Hippo cascade. It is also possible the Hippo-mediated transcription regulates kcnk5b expression or channel activity.
Connexin43 also regulates proportional growth of the fins, since mutations that reduce the intercellular connectivity of connexin43 produce adult fins are half the size as the fins of wild-type siblings (Hoptak-Solga et al., 2007; Iovine et al., 2005). The connective nature of these intercellular junction proteins indicate that direct communication between intracellular compartments of tissue cells is an important component of the scaling mechanism of the fins. Our observation that Kcnk5b cell-autonomously activates the 7xTCFsamois-mCherry reporter (Fig. 3) indicates that it is not due to intracellular transfer of K+. It is still unclear whether the disruption of intracellular trafficking of other ions (such as Na+ or Ca2+) or of other factors is responsible for the connexin43’s effect on scaling.
Kcnk5b’s ability to activate the β-catenin-dependent Wnt reporter cell autonomously in different tissue types supports the conclusion that K+ conductance has the potential to regulate developmental transcription in a broad range of tissues (Fig. 3B-D). The observation that neuronal cells in the brain and myocytes in the trunk muscle respond similarly to non-excitable cells elsewhere in the body suggests that even cells that harbor action potentials use K+ conductance to regulate gene expression. Whether this mechanism contributes to the scaling of other organs or how other electrophysiological mechanisms that control membrane potential can have the same effect needs to be explored.
In conclusion, we show how a specific electrophysiological mechanism activates important morphogen pathways to scale tissues in different in vivo contexts. We propose the observed diversity in morphogen and growth factor expression to Kcnk5b activity explains why the increased activity of Kcnk5b produces the diverse transcriptional response in the different tissues associated with the observed coordinated outgrowth of the entire fin. Also, we show how changes in phosphorylation of S345 in the cytoplasmic C-terminus is regulated by calcineurin to directly control electrophysiological activity of the channel to scale the fin. Thus, we offer an in vivo paradigm in which membrane potential acts as potent regulator of coordinated developmental signaling and that this is how the two-pore K+-leak channel Kcnk5b is able to scale the fish fin appendages.
Competing Interests
The authors declare that there are no competing interests.
Author contributions
C.Y., T.S., E.A.A.A., S.W., S.C., X.Y. Conducted experiments
M.W. Discussions, help with experimental design and analysis of electrophysiology data
A.E.A, K.G. Discussions, advice and financial support
C.A. Concept of the paper, designed experiments, conducted experiments, wrote the paper
Material and Methods
Cloning
Constructs were designed either using standard restriction enzyme or by homologous recombination methods. kcnk5b cDNA was isolated by RT-PCR from regenerating adult fin cDNA library and cloned into MCS region of pcDNA6-myc-6xHIS-tag plasmid (Invitrogen) or pBluescript harboring the hsp70 zebrafish promoter and GFP coding sequence surrounded by 2 miniTol2 sites. Mutagenesis of the Serine345 codon of Kcnk5b was performed using QuikChange Mutagenesis kit (Agilent).
Zebrafish Husbandry
AB strain fish were raised in 10L tanks with constantly flowing water, 26°C standard light-dark cycle (Brand et al., 2002) in either a Schwarz (DFG-Center for Regeneration, TU Dresden) or HaiSheng aquarium (ShanghaiTech University) systems. Transgenic lines were created by injecting 300 µg of each construct together with mRNA of Tol2 transposase (Balciunas et al., 2006). Fish embryos and larva were raised in 1xE3 medium (5mM NaCl, 0.17mM KCl, 0.33mM CaCl2, 0.33mM MgSO4, 10−5% Methylene Blue) until 10-12dpf, then transferred to aquarium water tanks to grow. Transgenic lines established by screening for GFP expression after heat shock. Experiments used male and female fish equally. Fish experiments were compliant to the general animal welfare guidelines and protocols approved by legally authorized animal welfare committees (Technische Universität Dresden, Landesdirektion Dresden, and ShanghaiTech University, ShanghaiTech Animal Welfare Committee).
Heat shock induction of transgenes
Parents of heat-shock-driven transgenic lines were either outcrossed to same-strain wild-type fish or to fish harboring Tg[7xTCFsamois:mCherry] transgene. Progeny were collected in 1xE3 and raised at 28°C. Carriers were confirmed positive for their respective transgenes by a single heat shock at 37°C for 1 hour. For embryo experiments, heat shock was at 12hpf in 37°C E3 medium for 30 min. For larva the heat shock was in 37°C E3 medium for 30 min. at 2 dpf. For adult fin, 6 month-old fish underwent a daily heat-chock regimen: first, sedated in 0.04% tricane in aquarium water, then placed in conical tubes containing 0.04% tricane in aquarium water to allow continued gill movement in oxygenated water and allow the caudal fins to be exposed to 37°C water for 7 min. After heat shocking the caudal fin, the fish were returned to flowing aquarium water and monitored daily.
Immunohistochemistry
After 4 hours, larvae were euthanized in 1% Mesab, fixated in 4%PFA/1xPBS and embedded in 1% agarose (CryoStar NX50, Thermofisher) before cryosectioning. 10 μm sections were mounted on glass slides (Brand, Thermofisher) and dried. The tissue freezing medium (Leica) was removed in ddH2O for 10min. Sections were permeabilized in 0.1%Triton-X for 5min and incubated in 1% BSA/1xPBS/0.1%Tween-20 (PBST) at room temperature for 30min. Sections were incubated in a mouse-anti-GFP (Invitrogen, MA5-15349), rabbit-anti-mCherry antibody (Invitrogen, P5-34974) solution (1:2000 dilution) in PBST at 4°C overnight. The primary antibody solution was replaced by a goat-anti-mouse-GFP (Abcam, ab150113), goat-anti-rabbit-mCherry secondary antibody (Abcam, ab 150078) solution (1:2000 dilution) in PBST and incubated at RT in the dark for 60 min. The secondary antibody solution was then replaced with DAPI solution (Roche, 10236276) in the dark at room temperature for 5 min. DAPI were washed away in 1xPBS. Coverslips were mounted with a 40% glycerol solution and sealed with nail polish. The sections were visualized using an LSM710 confocal (Zeiss) with ZENBlue software (Zeiss). Images were processed with Fiji software.
Cell Transplantations
Transgenic fish lines [hsp70:kcnk5b-GFP] and [7xTCFsamois:mCherry] were inbred for 20-30 min before embryos were collected in E3. The parents were confirmed to be positive for the respective transgene to ensure the highest number of embryos positive for the transgene. The embryos were left to develop in E3 for 3-3.5 hours at 28°C until the blastula stage. Embryos were then placed in agarose ridges for easy access under a Zeiss stereomicroscope and cells from the [hsp70:kcnk5b-GFP] embryos were isolated by air suction via a glass needle mounted on a Precision Instruments piston. The [hsp70:kcnk5b-GFP] cells were then transplanted into the [7xTCFsamois:mCherry] embryos. The transplants were carefully moved from the agarose into clean 90mm dishes with fresh E3 (+/- 20-25 per dish) and incubated at 28°C. Embryos hatched about 3 days after transplantation. They were directly heat shocked for 1 hour at 37°C. Four hours after heat shock, larvae were embedded into 1% low melting agarose (Sigma-Aldrich-Aldrich, A9414-250G) supplemented with Mesab in 35mm glass bottom confocal dishes (Cellvis: D35-20-1-N) and turned to their side. Visualized with a LSM710 confocal argon laser microscope (Zeiss) with ZENBlue software (Zeiss).
In situ hybridization
mRNA probes were made from RT-PCR products isolated from 2 dpf zebrafish embryos. The primer sequences for generating the probes are F-shha:5’-TGCGGCTTTTGACGAGAG TGC-3’R-shha: 5’-GGTAATACGACTCACTATAGGG TTTCCCGCGCTGTCTGCCG-3’ F-lef1 : 5’-GAGTTGGACAGATGACCCCTCCTC-3’; R-lef1:5’-GGTAATACGACTCA CTATAGGGGCAGACCATCCTGGGTAAAG-3’. in vitro transcription reagents are from Promega. Isolated fish fins were surgically isolated and incubated in 4% PFA in 1xPBS at 4°C overnight with gentle rocking. Samples were subsequently washed 5 times in 1xPBS and then dehydrated by incubation for 15 min in a graded series of increasing methanol/1xPBS solutions (25%, 50%, 75%, 3x 100%) on ice. Fins were then incubated in 100% methanol for ≥2hrs at -20°C. Samples were then rehydrated using the reversed dehydration series of methanol/1xPBS solutions). Samples were then incubated more than 4x in 1xPBS to remove all methanol, and subsequently incubated in 10µM Proteinase K for 10 min at RT. Samples were then incubated 20 min. in 4% PFA/1xPBS to inactivate the Proteinase K. Samples were incubated in 1xPBS 6x 10 mins to remove the PFA, then incubated in pre-hybridization buffer for 2 hours at 65C. Samples were subsequently incubated in the hybridization solution containing 200ng/ml of each mRNA probe ≥ 14 hrs at 65°C. Samples then were washed with successive wash steps to remove unbound probe and prepare for antibody incubation: twice 2xSSC/50% deionized formalin at 65°C, twice 2xSSC/25% deionized formalin at 65°C, 2xSSC at 65°C, twice 0.2xSSC at RT, 6 times 1xPBST (1xPBS with Tween-20), once in blocking solution [2% Bovine albumin (Sigma-Aldrich, A3294-100G), 2% Sheep Serum (Meilunbio, M134510)] at RT for 4 hours. Samples were incubated with Anti-digoxigenin-AP Fab Fragment (Sigma-Aldrich, 11093274910) in blocking solution ≥14 hrs at 4°C. Samples were then washed 6 times with 1xPBST, subsequently incubated in [0.1 M Tris-HCl, pH 9.5, 0.1 M NaCl, 0.05 M MgCl2] 3 times for 30 min, and then in Nitro Blue Tetrazolium (Sigma-Aldrich, N6639-1G) and 5-bromo-4-chloro-3-indolyl phosphate (Sigma-Aldrich, 136149) in [0.1 M Tris-HCl, pH 9.5, 0.1 M NaCl, 0.05 M MgCl2] at RT ≥8hrs. Samples images under Stemi508 stereoscope (Zeiss) with Axiocam ERc5s digital color camera (Zeiss) and Zen2.3 software (Zeiss).
Immunohistochemistry
Zebrafish fins or larvae euthanized in 1% Mesab were fixed in 4%PFA/1xPBS and embedded in 1% agarose or tissue freezing medium (Leica, 14020108926) on dry ice. (CryoStar NX50, Thermofisher) before cryosectioning. 10 μm sections were mounted on glass slides (Titan, 02036398) and dried. The tissue freezing medium (Leica) was removed in ddH2O for 10min. Sections were permeabilized in 0.1%Triton-X for 5min and incubated in 1% BSA/1xPBS/0.1%Tween-20 (PBST) at room temperature for 30min. Sections were incubated in a mouse-anti-GFP (Invitrogen, MA5-15349), (1:400) Anti-β-catenin (Cell Signaling, 9562L), rabbit-anti-mCherry antibody (Invitrogen, P5-34974) solution (1:2000 dilution) in PBST at 4°C overnight (>12 hours). The primary antibody solution was replaced by a goat-anti-mouse-GFP (Abcam, ab150113), goat-anti-rabbit-mCherry secondary antibody (Abcam, ab 150078) solution (1:2000 dilution) in PBST and incubated at RT in the dark for 60 min. The secondary antibody solution was then replaced with DAPI solution (Roche, 10236276) in the dark at room temperature for 5 min. DAPI were washed away in 1xPBST 3x 5-min incubations at RT. Coverslips (Titan, 02036401) were mounted with a 40% glycerol solution and sealed with nail polish. The sections were visualized using an LSM710 upright scanning confocal (Zeiss) or a LSM880 inverted scanning confocal (Zeiss) with ZENBlue software (Zeiss). Images were processed with Fiji software.
β-catenin nuclear analysis on IHC cross sections of fins
Using Fiji ImageJ, multiple nuclei of cells in IHC fin cross section image of control fins were manually measured, and a mean mRFP fluorescence intensity value was calculated. This mean value was used as the baseline for assessing nuclear β-catenin levels. β-catenin nuclear values for all the nuclei in each cross section were assessed with ImageJ by splitting the combined images of β-catenin and DAPI and using DAPI to have Fiji ImageJ define and select all nuclei in the image. The nuclear β-catenin levels were determined in selected nuclei by intensity analysis in Fiji ImageJ, which provided a numeric value for the β-catenin channel in all the nuclei of each section.
qRT-PCR
The cDNA that was used for qRT-PCR was extracted from the HEK293T cells and zebrafish. The mRNA was isolated using Tri-Reagent (CWBio, 03917). Then 1µg mRNA was used for the reverse transcription to cDNA using 4x gDNA wiper Mix, 5x HiScript III qRT SuperMix (Vazyme, L/N 7E350C9). qRT-PCR was performed using 2x ChamQ Universal SYBR qPCR Master Mix (Vazyme, L/N TE342F9) with QuantStudio3 machine (Thermofisher). The cycle procedure was at 50.0°C for 2 min, 95.0°C for 10 min in the stage 1; 95.0°C for 15 s, 60.0°C for 20s for 40 routine in the stage2; 95.0°C for 15s, 60.0°C for 1 min, 95.0°C for 15s in the Melt Curve. The data was analyzed in the ΔΔCt method.
Cell Culture
All cell culture lines were incubated at 37°C, 5% CO2, 95% humidity in incubators (Thermofisher, FORMA STERI-CYCLE i160). Cell were split to 50% density and transfected with Lipofectamine™ (Invitrogen, 11668-019) 12 hours later. Expression for the transfected constructs was evaluated by expression of fluorescent marker.
FRET-FLIM detection and analysis
Hek293T cells were transfected with 1µg of pcDNA-kcnk5b-GFP and 1µg of the pcDNA6-Kirin-FRET sensor (Shen et al., 2019). Fluorescence lifetime imaging measurements were made by photon counting the fluorescence emission of CFP using a 2-photon-confocal Hyperscope (Scientifica, UK) and PMT-hybrid 40 MOD 5 photon detectors (Picoquant, Germany). Counted photon emissions were calculated and analyzed using SymPhoTime 64, version 2.4 (Picoquant, Germany).
Electrophysiology
Transfected HEK293T cells were seeded on glass coverslips (Fisher Brand), and incubated in cell culture medium at 37°C, 5% CO2, 95% relative humidity for 4-6 hours. The seeded coverslips were transferred into Tyrode’s solution (138 mM NaCl, 4 mM KCl, 2 mM CaCl2,1 mM MgCl2, 0.33 mM NaH2PO4, 10 mM Glucose, 10 mM HEPES). Cells were assessed in the ruptured-patch whole-cell configuration of the patch-clamp technique using and EPC9 or EPC10 amplifier (HEKA) with borosilicate glass pipettes (Sutter Instruments) with 3-6 MΩ resistance when filled with pipette solution (130 mM glutamic acid, 10 mM KCl, 4 mM MgCl2, 10 mM HEPES, 2 mM ATP, pH to 7.2). After gigaseal formation, cells were voltage-clamped at -80 mV. Potassium conductance was elicited by test pulses from -100 mV to 70 mV (in 10mV increments) of 600 ms duration at a cycle length of 10s. The resulting tracings were converted into itx files by the ABF Software (ABF Software, Inc.) and then analyzed using Clampfit™ Software (Molecular Devices). Currents were measured at the end of the test pulses.
Acknowledgements
This work was supported by funding from ShanghaiTech University, the Deutsche Forschungsgemeinschaft Grant AN 797/4-1. We also wish to thank C. Bökel for scientific discussions.
References




