

ABSTRACT
Plant long noncoding RNAs (lncRNAs) have emerged as important regulators of chromatin dynamics, impacting on transcriptional programs leading to different developmental outputs. The lncRNA AUXIN REGULATED PROMOTER LOOP (APOLO) directly recognizes multiple independent loci across the Arabidopsis genome and modulates their three-dimensional chromatin conformation, leading to transcriptional shifts. Here, we show that APOLO recognizes the locus encoding the root hair (RH) master regulator ROOT HAIR DEFECTIVE 6 (RHD6) and controls RHD6 transcriptional activity leading to cold-enhanced RH elongation through the consequent activation of the transcription factor gene RHD6-like RSL4. Furthermore, we demonstrate that APOLO interacts with the transcription factor WRKY42 and modulates its binding to the RHD6 promoter. WRKY42 is required for the activation of RHD6 by low temperatures and WRKY42 deregulation impairs cold-induced RH expansion. Collectively, our results indicate that a novel ribonucleoprotein complex involving APOLO and WRKY42 forms a regulatory hub which activates RHD6 by shaping its epigenetic environment and integrates signals governing RH growth and development.
SUMMARY The lncRNA APOLO directly regulates the transcription of the root hair-master gene RHD6. In response to cold, APOLO is induced and it decoys the H3K27me3-binding protein LHP1 away from RHD6. In addition, APOLO modulates the binding of the transcription factor WRKY42 to the RHD6 promoter at low temperatures.
INTRODUCTION
Root hairs (RHs) are single cell projections developed from specialized epidermal trichoblast cells able to increase their size several hundred times in a polar manner to reach and promote the uptake of water-soluble nutrients, interact with soil microorganisms and support anchor to the plant. The specification of epidermal cells into RHs is a complex process whose underlying mechanisms are partially understood. In Arabidopsis thaliana, RH cell fate is controlled by a developmental program involving a complex of transcription factors (TFs) promoting the expression of the homeodomain protein GLABRA 2 (GL2) (Ryu et al., 2005; Song et al., 2011; Schiefelbein et al., 2014; Balcerowicz et al., 2015). GL2 blocks RH development by inhibiting the transcription of the master regulator ROOT HAIR DEFECTIVE 6 (RHD6) (Lin et al., 2015). In the cells that differentiate into RHs (known as trichoblasts), a second TF complex suppresses GL2 expression (Schiefelbein et al., 2014), forcing the cells to enter the RH cell fate program via the concomitant activation of RHD6 along with downstream TFs (Menand et al., 2007; Pires et al., 2013). Briefly, RHD6 together with its homolog RSL1 (ROOT HAIR DEFECTIVE 6 LIKE 1) induce the expression of TFs from the bHLH family, including RSL2 (ROOT HAIR DEFECTIVE 6 LIKE 2) and RSL4 (ROOT HAIR DEFECTIVE 6 LIKE 4), ultimately triggering the differentiation of the RHs and their subsequent polarized tip-growth (Karas et al., 2009; Yi et al., 2010; Bruex et al., 2012). In addition, it was proposed that RSL4 controls the expression of a small subset of nearly 125 genes (Won et al., 2009; Yi et al., 2010; Datta et al., 2015; Vijayakumar et al., 2016), including several cell wall extensins (EXTs) (Ringli, 2010; Velasquez et al., 2011) sufficient to promote RH growth (Hwang et al., 2017).
RH expansion is regulated both by cell-intrinsic factors (e.g. endogenous phytohormones such as auxin) and external environmental signals (e.g. phosphate (Pi) availability in the soil) (Mangano et al., 2017; Bhosale et al., 2018). Pi starvation is one of the key environmental factors promoting rapid RH growth (Yi et al., 2010; Datta et al., 2015; Vijayakumar et al., 2016). In Arabidopsis, it triggers RSL4 expression via an enhanced auxin production, activating downstream effector genes mediating cell growth (Yi et al., 2010; Datta et al., 2015; Mangano et al., 2017; Marzol et al., 2017; Bhosale et al., 2018). Accordingly, several auxin-related TFs have been implicated in Pi-starvation signaling in roots, including WRKY proteins that control the expression of the Pi transporter families Pi-permease PHO1 and PHOSPHATE TRANSPORTER (PHT) (Devaiah et al., 2007; Chen et al., 2009; Wang et al., 2014; Su et al., 2015). Under Pi-sufficient conditions, WRKY6 and WRKY42 bind to W-boxes of the PHO1 promoter and suppress its expression. During Pi starvation, WRKY42 is degraded by the 26S proteasome pathway, resulting in the activation of PHO1 transcription (Chen et al., 2009; Su et al., 2015). In addition, WRKY42 functions as a positive regulator of PHT1;1, by binding to its promoter under Pi-sufficient condition (Su et al., 2015). Overall, WRKY42 is part of the components activating root early-responses to Pi starvation, although its role in controlling RH growth remains unexplored.
In recent years, plant long noncoding RNAs (lncRNAs) have emerged as important regulators of gene expression, and several among them, have been functionally linked to Pi homeostasis. For instance, the lncRNA INDUCED BY PHOSPHATE STARVATION 1 (IPS1) can sequester the Pi starvation-induced microRNA miR-399, attenuating miR-399-mediated repression of PHO2, a gene encoding an E3 ligase affecting Pi uptake (Franco-Zorrilla et al., 2007). In addition, the cis-natural antisense (cis-NAT) transcript PHO1;2, induced under Pi deficiency, was shown to promote the translation of the PHO1;2 mRNA involved in Pi loading into the xylem. The expression of this cis-NAT is associated with the transport of the sense–antisense RNA pair toward the polysomes (Jabnoune et al., 2013). More recently, it was shown that the lncRNA AUXIN REGULATED PROMOTER LOOP (APOLO) recognizes multiple spatially independent genes by sequence complementarity and DNA-RNA duplex formation, known as R-loops. Upon recognition, APOLO shapes the three-dimensional (3D) conformation of its target regions by decoying the Poly-comb Repressive Complex 1 (PRC1) component LIKE HETEROCHROMATIN PROTEIN 1 (LHP1), thereby regulating their transcription (Ariel et al., 2014; Ariel et al., 2020).
Here, we show that the lncRNA APOLO directly regulates a subset of genes involved in RH development, including the master regulator of RH initiation RHD6. APOLO activates RHD6 transcription by modulating the formation of a local chromatin loop encompassing its promoter region, an epigenetic regulatory mechanism likely involving PRC1 and PRC2 components. Furthermore, we found that APOLO interacts with the TF WRKY42, forming a new hub that regulates RHD6 to induce RH growth in response to low temperatures. RHD6-mediated induction of RH expansion likely occurs through the transcriptional activation of the TF-encoding gene RSL4, which emerged as a key factor in the response to cold.
RESULTS
APOLO regulates root hair cell elongation in response to low temperatures
Based on Chromatin Isolation by RNA Purification (ChIRP-Seq) performed in wild-type (WT, Col-0) plants, it was previously reported that the lncRNA APOLO recognizes a subset of independent loci en-riched in categories related to cell wall composition and organization (Ariel et al., 2020). A closer look at APOLO bona fide targets allowed us to identify seventeen genes involved in RH growth and expansion (Supplementary Table 1), a process dependent on cell wall remodeling molecules, including EXTs and EXT-related proteins (Ringli, 2010; Lamport et al., 2011; Velasquez et al., 2011; Velasquez et al., 2015; Marzol et al., 2018). Interestingly, according to single-cell RNA-seq datasets (Zhang et al., 2019) APOLO transcripts are enriched in RH cells (Supplementary Figure 1A). Notably, sixteen APOLO direct targets were upregulated upon APOLO over-expression (Supplementary Table 2). Furthermore, 52 additional RH-related genes were upregulated in 35S:APOLO seedlings (Supplementary Table 2) (Ariel et al., 2020). Among them, the RH central TFs RHD6 (as a direct target), RSL2 and RSL4 (as indirectly regulated) were induced upon APOLO over-expression.
It was reported that the APOLO locus is targeted by the RNA-polymerase Pol V and silenced by RNA-directed DNA Methylation (RdDM,(Ariel et al., 2014)). A search in a small RNA-Seq performed in WT roots subjected to different temperature treatments (Gyula et al., 2018) revealed that RdDM-related 24nt siRNA accumulation over the APOLO locus is less abundant at low temperatures (Supplementary Figure 1B), suggesting that APOLO transcription is regulated by cold. Accordingly, we found that APOLO transcriptional accumulation increases in roots after 24h at 10°C (Figure 1A). An analysis of the promoter activity of the 5.2kb region upstream APOLO (Ariel et al., 2020) directing the expression of a GFP reporter gene, additionally revealed a higher transcriptional activity at low temperatures in the RHs (Figure 1B). Strikingly, we observed that two RNAi-APOLO (repression of approximately 90%; Ariel et al., 2014) and 35S:APOLO independent lines (over expression of 30-fold and 60-fold, respectively; Ariel et al., 2020) exhibit a basal increase of RH length at 22°C, and uncovered a strong induction of RH elongation in WT and RNAi-APOLO at 10°C, in contrast to 35S:APOLO lines (Figure 1C). Accordingly, RHD6 is induced in response to cold in WT roots, whereas RNAi-APOLO and 35S:APOLO roots display higher RHD6 basal levels than the WT (Figure 1D). Notably, RHD6 transcript levels are not further induced by cold in 35S:APOLO roots (Figure 1D), in agreement with the RH phenotype (Figure 1C). Collectively, our findings suggest that APOLO participates in the induction of cold-mediated RH elongation and a deregulation of APOLO transcript levels can impact RH growth.

A. APOLO transcript levels measured by RT-qPCR in roots of 10-day-old plants grown at 22 °C treated or not for 24h at 10 °C.
B. The APOLO promoter region (full intergenic region between PID and APOLO) is active in root hairs at 22 °C and 10 °C, using the GFP reporter gene. Images are representative of the lowest and highest GFP signal detected in the observation of several roots. The quantification of the fluorescence is shown in the right panel.
C. Quantification of RH length of 8-day-old Col-0, RNAi-APOLO and 35S:APOLO plants, respectively, at 22 °C and 10 °C. Each point is the mean ±error of the length of the 10 longest RHs identified in the root elongation zone in a single root. Representative images of each genotype are shown below the graph. The scale bas represent 500μm. Asterisks indicate significant differences between Col-0 and the corresponding genotype at the same temperature or significant differences between temperature treatments on the same genotype at the same temperature (one way ANOVA followed by a Tukey-Kramer test; “*”<0.05, “**”<0.01, “***”<0.001). NS stands for no statistically significant difference.
D. RHD6 transcript levels measured by RT-qPCR in roots of Col-0 plants vs. two independent RNAi-APOLO lines and two independent 35S:APOLO lines grown at 22 °C treated or not for 24h at 10 °C.
In A and D, the error bars represent the SD of 3 biological replicates. The asterisks indicate that the differences are significant (t test p<0.05). Values are normalized using the constant housekeeping transcript PP2A.
Previous studies pointed out a key role of RHD6 (together with RSL1) in RH development, which is mediated by RSL4 and RSL2 as downstream regulators of RH cell elongation (Menand et al., 2007; Pires et al., 2013). Notably, RHD6, RSL2 and RSL4 transcript levels were upregulated in 35S:APOLO seedlings (Supplementary Table 2; (Ariel et al., 2020)), although only RHD6 was identified as a direct target of APOLO (Ariel et al, 2020). In agreement with RHD6 transcriptional behavior (Figure 1D), RSL2 and RSL4 basal transcript levels are higher in RNAi-APOLO and 35S:APOLO compared to WT. Interestingly, RSL2 and RSL4 are induced by cold in WT and RNAi-APOLO, but not in 35S:APOLO roots (Supplementary Figure 2A), suggesting that low temperatures can activate these two genes still in the absence of APOLO and bypassing RHD6 for RH expansion (Figure 1C). Thus, we assessed if these TFs were also controlling the promotion of RH growth by low temperatures. To this end, we tested how rhd6/rsl1/rsl4 and rsl2, rsl4 and double mutant plants rsl2/rsl4 respond to low temperatures in comparison with control conditions (Supplementary Figure 2A). The rsl2 mutant was highly responsive to low temperatures in a similar manner to WT while rsl4 was impaired in the response to cold. The double mutant rsl2/rsl4 and the triple mutant rhd6/rsl1/rsl4 did not develop RHs in either of the two conditions. In addition, constitutive expression of RSL4 (35S:RSL4) as well as its expression under the control of the RH specific EXPANSIN7 promoter (EXP7p:RSL4) boosted basal RH growth without further enhancement in response to cold (Supplementary Figure 2B). These results demonstrate that RSL4 is a key factor mediating RHD6 activation of RH growth at low temperature, and RSL2 might participate to a lower extent.
Nutrient unavailability is known to activate RH expansion through a transcriptional reprogramming governed by RHD6 and downstream TFs. The quantification of RH growth of WT plants in response to increasing concentrations of nutrients (0.5X to 2.0X MS (Murashige and Skoog) medium) indicates that high concentrations impair RH growth triggered by low temperatures (Supplementary Figure 3A). In a similar way, an increase in agar concentration in the MS medium (from 0.8% to 2.5%), which likely re-strains nutrient mobility (Singha, S., Townsend, E.C. and Oberly, 1985; Nonami and Boyer, 1989; Ghashghaie et al., 1991; Buah et al., 1999) blocks cold-induced RH expansion (Supplementary Figure 3B). Altogether, these observations suggest that low temperatures restrict nutrient mobility and availability in the culture medium, leading to the promotion of polar RH growth.
APOLO directly modulates the three-dimensional chromatin conformation of the root hair specific locus RHD6
Among APOLO targets involved in RH development, we found the master regulator of RH initiation RHD6 (Menand et al., 2007; Pires et al., 2013). The epigenetic profile of the RHD6 locus corresponds to typical APOLO targets (Figure 2A, (Ariel et al., 2020)), including H3K27me3 deposition (track 1), LHP1 recognition (track 2, chromatin immunoprecipitation (ChIP)-Seq, (Veluchamy et al., 2016)), and APOLO binding regions (tracks 3 to 5, chromatin isolation by RNA purification (ChIRP)-Seq, (Ariel et al., 2020)). A GAAGAA box, shown to be important for APOLO target recognition (Ariel et al., 2020) is located in the RHD6 locus and coincides with APOLO binding site. In addition, a peak of DNA-RNA hybrid immunoprecipitation (DRIP)-Seq from root samples indicates the presence of an R-loop coinciding with APOLO recognition sites over RHD6 (tracks 6 to 8, (Xu et al., 2020)).

A. Epigenomic landscape of the RHD6 locus. Lane 1: H3K27me3 deposition by ChIP-Seq (Veluchamy et al., 2016). Lane 2: LHP1 deposition by ChIP-Seq (Veluchamy et al., 2016). Lane 3 to 5: APOLO recognition by ChIRP-Seq (Lane 3 and 4, using ODD and EVEN sets of probes against APOLO, respectively; Lane 5, negative control using LacZ probes)(Ariel et al., 2020). Lane 6 to 8: R-loop formation by DRIP-Seq (R-loop Atlas, root samples,(Xu et al., 2020), on Watson strand (Lane 6) and Crick strand (Lane 7). DRIP negative control after RNAseH treatment is shown in Lane 8. Gene annotation is shown at the bottom.
B. Chromatin loops identified in the RHD6 region by DpnII HiC (Liu et al., 2016). Colors of the loops are related to the corresponding q-values indicated below. Black boxes in A and B indicate the same genomic locations, where the bases of the chromatin loop correlate with R-loop formation and APOLO recognition (compared to gene annotations). In red the arrows indicate the position of the probes used for 3C-qPCR shown in D.
C. APOLO association to DNA of the RHD6 locus by ChIRP-qPCR in Col-0 and RNAi-APOLO plants. The background level was determined using a set of probes against LacZ RNA.
D. Relative chromatin loop formation by 3C-qPCR deduced from B, in Col-0 plants vs. 35S:APOLO and RNAi lines for the region upstream RHD6. The probes used for 3C-qPCR are indicated in red in panel B.
In C and D, the error bars represent the SD of 3 biological replicates. The asterisks indicate that the differences are significant (t test “*”<0.05, “***”<0.001).
Remarkably, APOLO recognition and R-loop formation are also detectable over RHD6 neighbor gene, located 3.2 kb upstream RHD6 transcription start site (Figure 2A). According to DpnII Hi-C datasets from Arabidopsis seedlings (Liu et al., 2016), a chromatin loop encompassing the intergenic region up-stream RHD6 was detected (Figure 2B), and coincides with APOLO binding-sites (Figure 2A, ChIRP-Seq). By performing a ChIRP-qPCR with two independent sets of biotinylated probes to purify APOLO (ODD and EVEN; (Ariel et al., 2020)) and one additional set used as a negative control (LacZ), we confirmed that APOLO RNA–RHD6 DNA interaction occurs in WT and is lost in APOLO knockdown (RNAi) seedlings ((Ariel et al., 2014); Figure 2C). In addition, the quantification of relative RHD6 loop formation in RNAi-APOLO and 35S:APOLO (Ariel et al., 2020) seedlings, revealed impaired loop formation in both lines (Figure 2D), hinting at a stoichiometric requirement of APOLO for RHD6 chromatin loop formation. Chromatin loop formation (Figure 2D) is in agreement with RHD6 basal levels in 35S:APOLO and RNAi-APOLO lines (Figure 1D), suggesting that the chromatin loop including RHD6 promoter region precludes transcription. Altogether, our results indicate that APOLO lncRNA directly regulates RHD6 transcriptional activity by fine-tuning local chromatin 3D conformation.
It was previously reported that PRC2 actively participates in the regulation of RH growth (Ikeuchi et al., 2015) and that the RHD6 locus exhibits H3K27me3 deposition and LHP1 recognition (Figure 2A; (Veluchamy et al., 2016)). Considering that the lncRNA APOLO interacts with the PRC1 component LHP1 in vivo (Ariel et al., 2014; Ariel et al., 2020), we decided to explore the role of PRC1 and PRC2 in APOLO-mediated RHD6 activation at low temperatures. At 22°C, RHD6 suffers a reduction of H3K27me3 in the PRC2 mutant curly leaf (clf), in contrast to the PRC1 mutant lhp1 (Supplementary Figure 4A; (Veluchamy et al., 2016)). Interestingly, we observed that H3K27me3 deposition and LHP1 binding diminish in WT roots treated for 24h at 10°C compared to 22°C (Supplementary Figure 4B), consistent with the induction of RHD6 in response to cold (Figure 2E). Moreover, lhp1 and clf mutants exhibit a basal decrease of RH length together with a slight decrease of cold-induced RH elongation in lhp1, and a strong decrease of cold-induced RH elongation in clf (Supplementary Figure 4D). Consistently, although the decrease in H3K27me3 deposition results in higher basal transcript levels of RHD6 in the clf background, RHD6 transcriptional activation by cold is abolished in the clf and lhp1 mutants (Supplementary Figure 4C), hinting at an important role of chromatin rearrangement for RHD6 activation in response to cold.
APOLO interacts with the transcription factor WRKY42 to coordinate the activation of RHD6
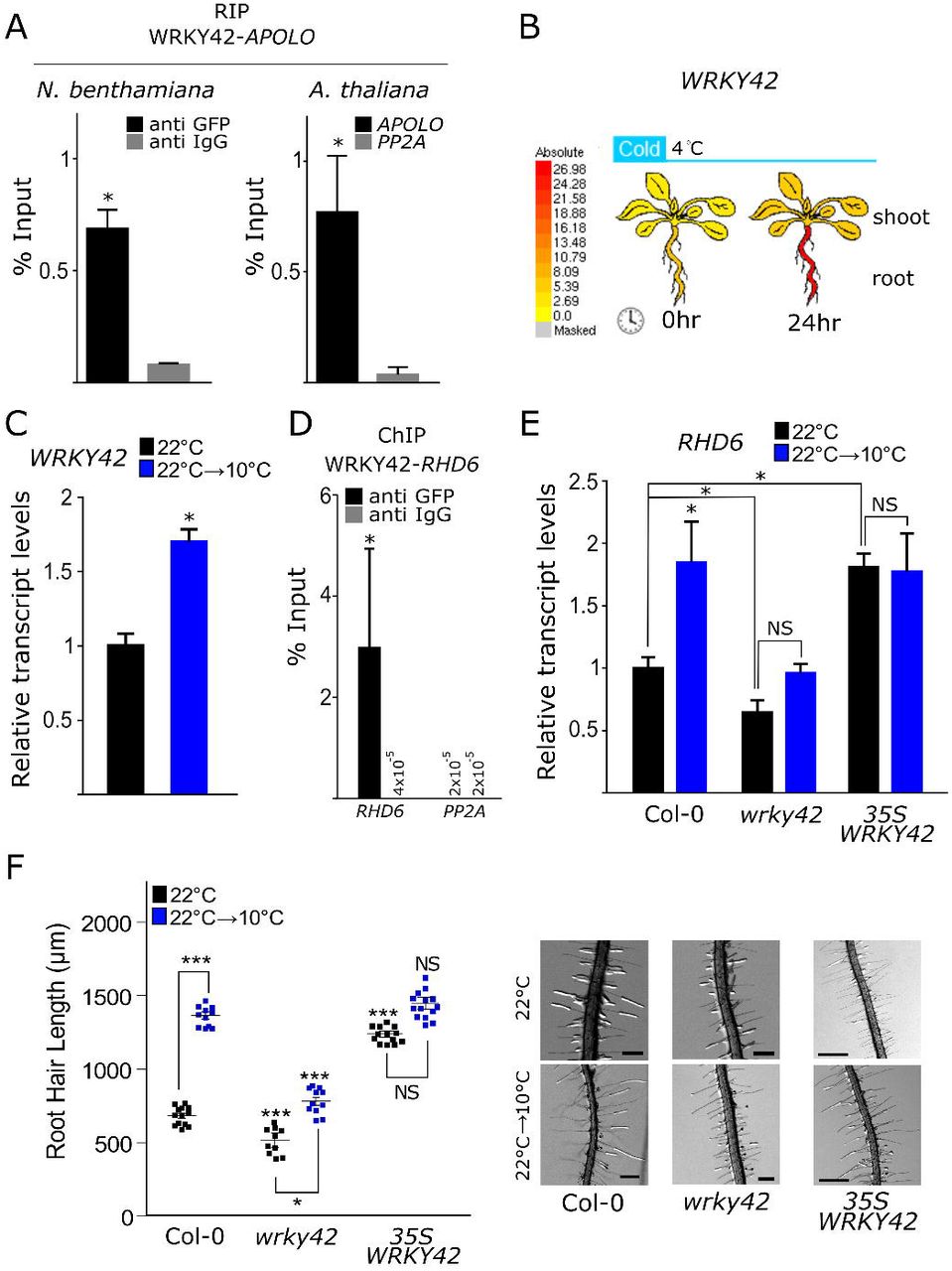
In order to uncover novel actors involved in cold-induced transcriptional regulation of RH growth, we aimed at identifying APOLO protein partners. To this end, we performed an exploratory ChIRP on WT seedlings using two independent set of biotinylated probes to purify APOLO (ODD and EVEN; (Ariel et al., 2014; Ariel et al., 2020)), and one additional set used as a negative control (against LacZ), as recently described (Rigo et al., 2020). Co-purified proteins were precipitated and analyzed by mass spectrometry. Among the potential APOLO partners (i.e. identified with at least two hits in ODD and EVEN samples, but absent in LacZ-ChIRP), we found the WRKY42 protein, a TF involved in the response to Pi starvation (Su et al., 2015), an environmental condition that promotes RH cell expansion (Bhosale, et al., 2018) in a similar manner to low temperatures. Thus, APOLO-WRKY42 interaction was validated by RNA immuno-precipitation (RIP-qPCR) in tobacco leaves and in Arabidopsis plants transitory or stably transformed with 35S:WRKY42:GFP, respectively (Figure 3A). Interestingly, according to the Arabidopsis eFP Browser (Waese et al., 2017), WRKY42 is induced in roots when seedlings are subjected to 4°C for 24h (Figure 3B). At 10°C, we observed that WRKY42 transcriptional accumulation augments significantly in roots (Figure 3C). Notably, 13 out of the 17 APOLO targets contained between 1 and 4 canonical WRKY TF binding sites (W-box) in their promoters, including RHD6 (Supplementary Table 1). By using a 35S:WRKY42:GFP line, we determined that WRKY42 can directly bind to the promoter region of RHD6 (Figure 3D). Accordingly, the over-expression of WRKY42 (35S:WRKY42:GFP line) led to a basal increase of RHD6 levels (Figure 3E) and RH elongation (Figure 3F) at ambient temperature, mimicking the effect of cold. On the contrary, cold-mediated induction of RHD6 is abolished in the wrky42 mutant ((Su et al., 2015); Figure 3E), which consistently exhibits shorter RHs at 22°C and almost no RH elongation at low temperatures (Figure 3F). Taken together, these results suggest that the APOLO-interacting TF WRKY42 is an important regulator of RHD6-mediated RH growth in response to cold.

A. APOLO-WRKY42 in vivo interaction demonstrated by RNA Immunoprecipitation (RIP)-qPCR in transiently transformed tobacco leaves and stably transformed Arabidopsis plants. In tobacco, WRKY42:GFP and APOLO were co-transformed under the control of the 35S constitutive promoter. In Arabidopsis, 35S:WRKY42:GFP transformed plants were used to detect the interaction with endogenous APOLO. AntiIgG antibodies were used as a negative control. The RNA PP2A was considered as a RIP negative control in Arabidopsis plants.
B. Arabidopsis eFP Browser (Waese et al., 2017) plot representing the increase of WRKY42 transcript levels in roots of seedlings treated for 24h at 4°C.
C. WRKY42 transcript levels measured by RT-qPCR in roots of plants grown at 22°C treated or not for 24h at 10°C.
D. Chromatin Immunoprecipitation (ChIP)-qPCR assay revealing regulation by WRKY42 of RHD6 by direct recognition of its promoter region. Probes amplifying PP2A were used as a negative control of the experiment. Anti-IgG antibodies were used as a negative control for each pair of probes.
E. RHD6 transcript levels measured by RT-qPCR in roots of Col-0 plants vs. wrky42 mutants and 35S:WRKY42:GFP lines grown at 22°C treated or not for 24h at 10°C.
F. Quantification of RH length of Col-0, wrky42 and 35S:WRKY42:GFP plants at 22°C and 10°C. Each point is the mean of the length of the 10 longest RHs identified in a single root. Representative images of each genotype are shown on the right. The scale bas represent 500μm. Asterisks indicate significant differences between Col-0 and the corresponding genotype at the same temperature (one way ANOVA followed by a Tukey-Kramer test; “*”<0.05, “***”<0.001).
In A, C, D and E, the error bars represent the SD of 3 biological replicates. The asterisks indicate that the differences are significant (t test p<0.05). NS stands for no statistically significant difference. In C and E, values are normalized using the constant housekeeping transcript PP2A.
We thus wondered to what extent WRKY42 regulates the epigenetic landscape of the RHD6 locus. We first observed that H3K27me3 deposition over RHD6 is significantly augmented in the wrky42 mutant background, in contrast to AZG2, an APOLO target non-related to WRKY42 (Figure 4A), consistent with reduced RHD6 basal levels reported in wrky42 (Figure 3E). Therefore, we evaluated the mutual contribution of APOLO and WRKY42 to their respective recognition of the RHD6 locus. APOLO ChIRP-qPCR in the WT and wrky42 mutant (Figure 4B) revealed similar binding to RHD6, indicating that WRKY42 does not participate in APOLO-target recognition. Reciprocally, we assessed the control of APOLO over WRKY42 recognition of the RHD6 locus. To this end, we transformed transiently A. thaliana leaves of WT, 35S:APOLO and RNAi-APOLO plants with the construct 35S:WRKY42:GFP. Observation using confocal microscopy indicated that WRKY42:GFP is localized in the nucleus in the three genetic backgrounds (Supplementary Figure 5). Remarkably, WRKY42:GFP ChIP-qPCR revealed that WRKY42 binding to RHD6 promoter is impaired both in 35S:APOLO and RNAi-APOLO plants compared to WT (Figure 4C), hinting at a stoichiometric role of APOLO in the modulation of TF-chromatin interaction. To further confirm our observations, chromatin was extracted from 35S:WRKY42:GFP seedlings and increasing amounts of in vitro transcribed APOLO were added before cross-link and regular WRKY42 ChIP over RHD6. Strikingly, increasing concentrations of APOLO gradually decoy WRKY42 away from the RHD6 locus (Figure 4D), further supporting the hypothesis of a stoichiometric regulation of APOLO over the activity of its partner TF.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
A. H3K27me3 deposition in Col-0 vs. wrky42 mutant 10-day-old seedlings grown at 23°C over RHD6 and a WRKY42-independent APOLO target (AZG2; (Ariel et al., 2020)). An immunoprecipitation (ChIP-qPCR) was performed using an anti-IgG antibody as a negative control.
B. APOLO association to DNA of the RHD6 locus by ChIRP-qPCR in Col-0 and wrky42 mutant plants. The background level was determined using a set of probes against LacZ RNA.
C. WRKY42 binding to RHD6 promoter in transiently transformed leaves of Col-0, RNAi-APOLO and 35S:APOLO plants. WRKY42:GFP was immunoprecipitated using the anti-GFP antibody (ChIP-qPCR). An immunoprecipitation was performed using an anti-IgG antibody as a negative control.
D. WRKY42 immunoprecipitation over RHD6 with the addition of increasing concentrations of in vitro transcribed APOLO prior to cross-linking.
E. Proposed model for the action of APOLO in RH growth under low temperatures. At 22 °C, RHD6 moderate transcription is regulated by APOLO, H3K27me3 and LHP1, maintaining a repressive chromatin loop encompassing the RHD6 promoter region. In response to cold, APOLO levels increase, still recognizing the RHD6 locus by sequence complementarity (R-loops). Then, LHP1 (brown balls) is decoyed away from chromatin and PRC2-dependent H3K27me3 (green balls) deposition decreases in a process involving WRKY42 and leading to the opening of the RHD6 promoter region. Additionally, APOLO and WRKY42 jointly activate RHD6 transcription. RHD6 activates the transcription of downstream TF genes RSL2 and 4 promoting RH cell expansion. Yet unknown factors activate RSL2 and 4 independently from APOLO and RHD6 in response to cold. Furthermore, APOLO directly regulates a subset of downstream genes, likely including a subset of them controlled also by WRKY42.
In A to D the error bar represents the SD of 3 biological replicates. The asterisk indicates that the difference is significant (t test “*”<0.05, “**”<0.01).
DISCUSSION
Our results indicate that the regulation of RHD6 expression in response to cold depends on Polycomb-dependent H3K27me3 dynamic deposition. The WRKY42-APOLO complex modulates the epigenetic environment of RHD6, activating its transcription and promoting RH growth at low temperatures. RHD6 activation further triggers the expression of RSL2 and RSL4 that control the transcriptional RH program inducing cell expansion in response to cold (Figure 4E).
Cell fate determination in the epidermis has been extensively studied (Ryu et al., 2005; Song et al., 2011; Schiefelbein et al., 2014; Balcerowicz et al., 2015). Once trichoblast cells differentiate in the root epidermis, RHs develop as fast polar growing protuberances in response to endogenous and environmental signals (Menand et al., 2007; Yi et al., 2010; Pires et al., 2013; Marzol et al., 2017). RHs are one of the main entry points in the roots for water-soluble macronutrients, such as Pi and nitrates. Pi is an essential element for plant growth and development, and the availability of this macronutrient is a factor limiting plant productivity. In Arabidopsis roots, low Pi in the soil triggers auxin synthesis and transport, enhancing RH elongation to promote Pi uptake (Bhosale et al., 2018). Thus, auxin mediates low Pi-induced promotion of RH cell expansion. Under low soil Pi, auxin synthesis is enhanced specifically in the root cap (Stepanova et al., 2008) and transported (mostly by AUX1, PIN2, and PGP4) from the apex to the differentiation zone, specifically leading to an increase of auxin levels in trichoblasts (Jones et al., 2009; Bhosale et al., 2018; Wang et al., 2020). In response to the high-auxin microenvironment, RHs protrude from the root epidermis controlled by RHD6 and RSL1(Menand et al., 2007; Pires et al., 2013). High levels of auxin in trichoblasts trigger a signaling cascade mediated by TIR1-ARF19 (and possibly also ARF7) which directly induces the expression of RSL4 (and likely of RSL2) and promote RH elongation (Yi et al., 2010; Mangano et al., 2017; Mangano et al., 2018b; Bhosale et al., 2018). ARF7 and ARF19 also activate other RH genes independently of RSL4 (Schoenaers et al., 2018). Interestingly, our results indicate that the lncRNA APOLO participates in the response to low temperatures. APOLO is directly activated by ARF7 and regulates the transcriptional activity of its neighboring gene PINOID (PID) by shaping local 3D chromatin conformation (Ariel et al., 2014; Ariel et al., 2020). PID encodes a kinase responsible for accurate auxin polar transport by localizing PIN2 in the root cell membrane (Friml et al., 2004). More recently, it was shown that APOLO can recognize a subset of distant genes across the Arabidopsis genome, most of them being related to auxin synthesis and signalling (Ariel et al., 2020). In this work, we demonstrate that a group of RH related genes is directly regulated by APOLO in response to cold, including the RH master regulator RHD6. Interestingly, RSL2 and RSL4 are still activated by cold in RNAi-APOLO roots in contrast to RHD6 (Figure 1C and Supplementary Figure 2A), suggesting that additional yet uncovered factors, which might include ARF TFs, may trigger RH expansion in an APOLO/RHD6-independent manner, and hinting at RSL4 as a key regulator. Furthermore, we found that in addition to RHD6, APOLO directly regulates 16 RH-related genes downstream RHD6 and RSLs (Supplementary Table 1), hinting at an intricate regulatory network controlling RH expansion. Collectively, our results uncover a lncRNA-mediated epigenetic link between environmental signals and auxin homeostasis modulating RH growth. Moreover, our observations suggest that low temperatures restrict nutrient mobility and availability in the culture medium, leading to the promotion of polar RH growth. Further research will be needed to determine what is the limiting nutrient mediating the effect of cold on RH growth.
Although substantial progress has been achieved in the identification of the molecular actors controlling RH development, the impact of chromatin conformation in the transcriptional regulation of central TFs remains poorly understood. In this study, we have revealed a new mechanism of gene regulation in RHs by which the lncRNA APOLO integrates chromatin-associated ribonucleoprotein complexes together with the TF WYRK42, participating in the transcriptional activation of RHD6 and the down-stream RH gene network (Figure 4E). APOLO directly regulates the chromatin 3D conformation of the genomic region encompassing the RHD6 locus and stoichiometrically recruits WYRK42, previously linked to Pi-starvation (Chen et al., 2009; Su et al., 2015). Low levels of APOLO fail to recruit WRKY42 to RHD6 promoter region, whereas high levels of APOLO likely decoy WRKY42 from target chromatin, as it was shown for APOLO regulation over LHP1 binding activity (Ariel et al., 2020). Notably, low and high levels of APOLO result in higher RHD6 basal transcriptional accumulation (through LHP1 activity and change in chromatin status), whereas both extremes impair RHD6 activation by cold (through WRKY42 binding modulation; a model is shown in Supplementary Figure 6). Our results suggest that an WRKY42-APOLO hub regulates RH cell elongation through the master regulator RHD6, although the APOLO-WYRKY42 hub potentially targets several additional cell wall related genes (Supplementary Table 1) at the end of the pathway controlled by RHD6 and the RHD6-downstream TFs RSL2/RSL4 (Mangano et al., 2017; Mangano et al., 2018a).
Participation of epigenetic factors in root cell identity determination strongly suggests the default pattern for epidermal cell fate that can be overridden by environmental stimuli (Guimil and Dunand, 2006). Interestingly, it was reported that the expression of GLABRA2 (GL2), a gene encoding a TF repressing RHD6 in atrichoblasts, is tightly regulated at the epigenetic level. By using 3D fluorescence in situ hybridization, it was shown that alternative states of chromatin organization around the GL2 locus are required to control position-dependent cell-type specification in the root epidermis (Costa and Shaw, 2006). Furthermore, GL2 epigenetic regulation was proposed to be responsive to salt stress (Beyrne et al., 2019). In addition, a comprehensive characterization of alternative mutant lines uncovered the role of PRC2 in the regulation of RH development (Ikeuchi et al., 2015). Loss-of-function mutants in different PRC2 subunits develop unicellular RHs but fail to retain the differentiated state, generating a disorganized cell mass from each single RH. It was shown that the resulting RHs are able to undergo a normal endoreduplication program, increasing their nuclear ploidy, although they subsequently reinitiate mitotic division and successive DNA replication. It was proposed that aberrant RH development in PRC2 related mutants is due to the epigenetic deregulation of key regulatory genes such as WOUND INDUCED DEDIFFERENTIATION 3 (WIND3) and LEAFY COTYLEDON 2 (LEC2) (Ikeuchi et al., 2015). Here, we showed that the single mutants clf (PRC2) and lhp1 (PRC1) are affected in RH growth. In addition, we found that H3K27me3 deposition and LHP1 binding to the RHD6 locus is modulated by cold. Moreover, we showed that in the clf background, H3K27me3 deposition throughout the RHD6 locus is partially impaired, and that the clf mutant is affected in RH elongation promoted by cold. Notably, basal transcriptional levels of RHD6 are higher in the clf mutant (Supplementary Figure 4C). A phenotypic characterization revealed that RH density is not altered in clf or lhp1 mutants, nor in APOLO and WRK42 deregulated lines (Supplementary Figure 7), suggesting that RHD6 is not ectopically expressed in epidermic cells. Thus, RHD6 over-accumulation may occur in inner cell layers of the root or in RHs, although their elongation in the clf mutant may be blocked by additional perturbations of Polycomb-associated regulation of downstream genes. Altogether, our results suggest that Polycomb proteins participate in the control of RH-related genes transcriptional reprogramming at low temperatures.
Notably, CLF and LHP1 were shown to interact with a subset of lncRNAs in Arabidopsis, modulating the activity of PRC target genes (Lucero et al., 2020). Interestingly, several lncRNAs have been linked to the control of transcription in response to cold. FLOWERING LOCUS C is regulated by at least three lncRNAs. First, the alternative splicing of a set of antisense transcripts, collectively named as COOLAIR, depends on the prolonged exposure to cold, epigenetically repressing FLC (Marquardt et al., 2014). The use of the COOLAIR proximal poly(A) site results in down-regulation of FLC expression in a process involving FLOWERING LOCUS D (FLD), an H3K4me2 demethylase (Marquardt et al., 2014). A second lncRNA called COLD ASSISTED INTRONIC NONCODING RNA (COLDAIR) is fully encoded in the sense strand of the first intron of FLC. Similar to COOLAIR, its accumulation oscillates in response to low temperatures. It was proposed that COLDAIR recruits the PRC2 component CLF to target FLC for H3K27me3 deposition (Heo and Sung, 2011). More recently, a third lncRNA modulating FLC transcription was identified (Kim and Sung, 2017). The cold-responsive lncRNA COLDWRAP is derived from the FLC proximal promoter and it also interacts with PRC2. It was suggested that COLDWRAP functions in cooperation with the lncRNA COLDAIR to retain Polycomb at the FLC promoter through the formation of a repressive intragenic chromatin loop (Kim and Sung, 2017). Another lncRNA named SVALKA was shown to mediate the response to low temperatures (Kindgren et al., 2018). Interestingly, the activation of SVALKA by cold triggers the transcription of a cryptic downstream lncRNA, which overlaps the antisense locus of the C-repeat/dehydration-responsive element Binding Factor 1 (CBF1), involved in the early response to cold in Arabidopsis. Antisense transcription causes Pol II head-to-head collision modulating transcriptional termination of CBF1 (Kindgren et al., 2018). Here, we show that the auxin-responsive lncRNA APOLO is also transcriptionally modulated by cold. The differential accumulation of 24nt siRNAs across the APOLO locus at low temperatures indicates that this activation is related to a decrease in RdDM. Moreover, we showed here that the intergenic region between PID and APOLO acting as a divergent promoter is also activated at low temperatures in RHs, as revealed by the reporter gene GFP. Thus, the lncRNA APOLO integrates external signals into auxin-dependent developmental outputs in Arabidopsis.
In the last decade, lncRNAs have emerged as regulators of gene expression at different levels, ranging from epigenetics to protein modifications and stability (Ariel et al., 2015). Notably, it has been shown in animals that noncoding transcripts can be recognized by TFs. In humans, it was proposed that the interaction with the lncRNA SMALL NUCLEOLAR RNA HOST GENE 15 (SNHG15) stabilizes the TF Slug in colon cancer cells. It was shown that SNHG15 is recognized by the zinc finger domain of Slug preventing its ubiquitination and degradation in living cells (Jiang et al., 2018). Also, the transcriptional activity of the human gene DIHYDROFOLATE REDUCTASE (DHFR) is regulated by a lncRNA encoded in its proximal promoter. It was proposed that the nascent noncoding transcript forms a hybrid with its parent DNA and decoys the regulatory TF IIB away from the DHFR promoter, dissociating the transcriptional pre-initiation complex in quiescent cells (Martianov et al., 2007). The lncRNA P21 ASSOCIATED ncRNA DNA DAMAGE ACTIVATED (PANDA) was identified in human cancer and it was activated in response to DNA damage (Hung et al., 2011). PANDA is transcribed from the promoter region of the CDKN1A gene and interacts with the TF NF-YA to limit the expression of pro-apoptotic genes. The activity of PANDA has been linked to the progression of different tumors (Kotake et al., 2016; Shi et al., 2019). Interestingly, it was shown that in addition to NF-YA, PANDA interacts with the scaffold-attachment-factor A (SAFA) as well as PRC1 and PRC2 to modulate cell senescence. In proliferating cells, SAFA and PANDA recruit Polycomb components to repress the transcription of senescence-promoting genes. Conversely, the loss of SAFA–PANDA– PRC interactions allows expression of the senescence program (Puvvula et al., 2014). In this work, we showed that the PRC1-interacting lncRNA APOLO can also be recognized by the TF WRKY42, hinting at general lncRNA-mediated mechanisms linking Polycomb complexes with the transcriptional machinery across kingdoms. Furthermore, our observations indicate that the deregulation of WRKY42 affects the epigenetic environment of RHD6. It was previously shown that the addition of in vitro transcribed APOLO to RNAi-APOLO chromatin extracts was able to partially restore R-loop formation over APOLO target genes, and that high levels of APOLO may titer LHP1 away from chromatin (Ariel et al., 2020). Here we show that the relative accumulation of the lncRNA APOLO can modulate the binding activity of its partner TF to common target genes. Collectively, our results strongly support that environmentally controlled cell fate in Arabidopsis relies on a transcriptional reprogramming governed by a network of epigenetic regulatory complexes, lncRNAs, TFs and effector proteins.
MATERIALS AND METHODS
Plant Material and Growth Conditions
All the Arabidopsis thaliana lines used were in the Columbia-0 (Col-0) background. WRKY42 over expression transgenic plants were generated through Agrobacterium tumefaciens (strain EHA105)-mediated transformation (Clough and Bent, 1998). 35S:WRKY42:GFP transformant lines were selected on MS/2 medium supplemented with kanamycin (40μg/mL) and WRKY42 expression levels were measured by RT-qPCR (primers used are listed in Supplementary Table 3). The wrky42 mutant line belongs to the SALK collection (SALK_121674C), as the one previously characterized (Su et al., 2015).The rhd6-3/rsl1-1/rsl4-1; rsl2; rsl4; rsl2rsl4, 35S:RSL4 and EXP7p:RSL4 transgenic lines were previously described and characterized (Yi et al., 2010; Hwang et al., 2017) Homozygous plants were obtained in our laboratory and genotyped using the oligonucleotides indicated in Supplementary Table 3. Seeds were surface sterilized and stratified at 4°C for 2d before being grown under long day conditions (16h light, 140μE.m−2.sec−1/8h dark), on ½-strength Murashige and Skoog media (1/2 MS) (Duchefa, Netherlands) with 0.8% plant agar (Duchefa, Netherlands).
Cloning procedure
The coding region of WRKY42 (AT4G04450) excluding the STOP codon was amplified by PCR, cloned into the Gateway entry vector pENTR/D-TOPO (Invitrogen) and recombined by Gateway technology (LR reaction) into the pK7FWG2,0 vector containing a p35S-GFP cassette (http://www.psb.ugent.be/gateway/index.php).
Root hair phenotype characterization
For quantitative analyses of RH phenotypes, 10 fully elongated RH from the root elongation zone of 15-20 roots, were measured on the same conditions for each particular case and grown on vertical plates with ½-strength Murashige and Skoog media (1/2 MS) (Duchefa, Netherlands) and 0.8% plant agar (Duchefa, Netherlands) for 5 days at 22° and 3 days at 10°C. Measurements were made after 8 days. Images were captured with an Olympus SZX7 Zoom Stereo Microscope (Olympus, Japan) equipped with a Q-Colors digital camera and software Q Capture Pro 7(Olympus, Japan). RH density was determined as the number of hairs in a representative area of the root elongation zone using the same setup stated above and ImageJ software. Results were expressed as the mean ± standard error (SE) and in the case of RH density, values were expressed per mm2. All measurements indicate the average of three independent experiments, each involving 15-20 seedlings.
Confocal microscopy analysis of root hairs
Confocal laser scanning microscopy was performed using Zeiss LSM5 Pascal (Zeiss, Germany) and a 40x water-immersion objective, N/A=1.2. Fluorescence was analyzed by using 488 nm laser for GFP excitation (Laser Intensity: 70%, Detector Gain:550, Amplifier Offset:0.1, Amplifier Gain:1), and emitted fluorescence was recorded between 490 and 525nm for GFP tag. Z stacks were done with an optical slice of 1μm, and fluorescence intensity was measured in 15μm ROI (Region Of Interest) at the RH tip and summed for quantification of fluorescence using ImageJ. Five replicates for each of ten roots and 15 hairs per root were observed. Col-0 wild type root hairs were used as a negative control, to check autofluorescence signal occurrence and no signal were detected in the wavelengths range stated above.
RNA extraction and RT-qPCR
Total RNA was extracted using TRIZol (Invitrogen) and 2 μg were subjected to DNase treatment according to the manufacturer’s protocol (Thermo Scientific). One μg of DNase-free RNA was reverse-transcribed using Maxima H Minus Reverse Transcriptase (Thermo Scientific). RT-qPCR were performed using the LightCycler 480 SYBR Green I Master Kit on a LightCycler480 apparatus (Roche) using standard protocols (40 cycles, 60°C annealing). PP2A (AT1G13320; primers are listed in Supplementary Table 3) was used as reference.
RNA Immunoprecipitation
RNA immunoprecipitation (RIP) assays were performed on transiently transformed N. benthamiana leaves as described in (Sorenson and Bailey-Serres, 2015), or in 10-day-old A. thaliana 35S:WRKY42:GFP seedlings as described in (Bardou et al., 2014), using anti GFP (Abcam ab290) and anti-IgG (Abcam ab6702). RIP was performed using Invitrogen Protein A Dynabeads. Precipitated RNAs were prepared using TRI Reagent (Sigma-Aldrich), treated with DNase (Fermentas) and subjected to RT-qPCR (High Capacity cDNA Reverse Transcription Kit (Thermo); primers used are listed in Supplementary Table 3). Total RNAs were processed in parallel and considered the input sample.
Chromatin Immunoprecipitation
Chromatin immunoprecipitation (ChIP) assays were performed on 10-day-old WT seedlings treated or not during 24h at 10 °C, using anti H3K27me3 (Diagenode pAb-195-050), anti LHP1 (Covalab pab0923-P) and anti-IgG (Abcam ab6702), as described in (Ariel et al., 2020). Cross-linked chromatin was sonicated using a water bath Bioruptor Pico (Diagenode; 30sec ON/30sec OFF pulses; 10 cycles; high intensity). ChIP was performed using Invitrogen Protein A Dynabeads. Precipitated DNA was recovered using Phenol:Chloroform:Isoamilic Acid (25:24:1; Sigma) and subjected to RT-qPCR (primers used are listed in Supplementary Table 3). Untreated sonicated chromatin was processed in parallel and considered the input sample. For in vitro competition assays, APOLO was transcribed using the T7 transcription kit (Promega; (Ariel et al., 2020)). After regular chromatin isolation from 10-day-old 35S:WRKY42:GFP seedlings, the sample was split in 4 independent tubes and diluted to 1ml in Nuclei Lysis Buffer without SDS. 0 μg, 0.1 μg, 1 μg and 10 μg of APOLO were added to each sample respectively, and incubated in rotation at 4 °C for 3h. Then, cross-linking was performed with 1% formaldehyde for 5 min at 4 °C, followed by 5 min with a final concentration of 50 mM glycine. SDS was added to a final concentration of 0.1% prior to sonication and the subsequent steps of a regular ChIP protocol.
For ChIP in transiently transformed leaves, 3-week-old A. thaliana were transformed as previously described (Zhang et al., 2020) In brief, Agrobacterium tumefaciens strain GV3101 carrying 35S:WRKY42:GFP construct were grown for 2 days in YEB-induced medium plates at 28 °C. Agrobacterium cells were scraped and resuspended in washing solution (10 mM MgCl2, 100 μM acetosyringone). Infiltration solution (¼MS [pH = 6.0], 1% sucrose, 100 μM acetosyringone, 0.005% [v/v, 50 μl/l] Silwet L-77) was prepared with the previous solution, adjusting the OD600=0.5. The infiltration was carried out in all leaves > 0.5cm in length of between 10 and 15 plants per genotype (WT, 35S:APOLO and RNAi-APOLO lines). After infiltration, plants were kept in light for 1h and then in darkness for 24h. Finally they were transferred back to light. Images and samples were obtained 3 days after infiltration. For image acquisition, infiltrated leaves were imaged with a Leica TCS SP8 confocal laser scanning microscope with excitation at 488 nm (Intensity=8%) and detection at 495-530 nm for GFP and 610-670 (gain 650) nm for chlorophyll fluorescence. Images were captured using 10X and 20X lenses, and processed using Fiji software (Schindelin et al., 2012).
Chromatin Isolation by RNA Purification followed by qPCR or mass spectrometry
A method adapted from the ChIRP protocol (Chu et al., 2012) was developed to allow the identification of plant DNA associated to specific lncRNAs, as described in (Ariel et al., 2014; Ariel et al., 2020). Briefly, plants were in vivo crosslinked and cell nuclei were purified and extracted through sonication. The resulting supernatant was hybridized against biotinylated complementary oligonucleotides that tile the lncRNA of interest and lncRNA-associated chromatin was isolated using magnetic streptavidin beads. One hundred pmol of probes against APOLO (ODD and EVEN set of probes (Ariel et al., 2014; Ariel et al., 2020)) and the corresponding negative set against LacZ were used for independent purification. Co-purified ribonucleoprotein complexes were eluted and used to purify RNA or DNA, which were later subjected to downstream assays for quantification as previously described (Ariel et al., 2020).
The exploratory identification of plant nuclear proteins bound to APOLO was performed as described in (Rigo et al., 2020). Plant samples were prepared as for ChIRP-qPCR. For protein extraction, approximately 250 g of 7-day-old Col-0 plants grown on solid half-strength MS medium was irradiated three times with UV using a CROSSLINKERCL-508 (Uvitec) at 0.400 J/ cm2. Protein purification samples for protein extraction were DNase-treated according to the manufacturer (Thermo Scientific). After addition of 1.8ml of TCA-acetone (5ml 6.1N TCA + 45ml acetone + 35μl β-mercaptoethanol), samples were incubated overnight at −80°C. After centrifugation at 20000rpm for 20min at 4°C, the supernatant was discarded and 1.8ml of acetone wash buffer (120ml acetone, 84μl β-mercaptoethanol) was added to the samples. Then, samples were incubated 1h at −20°C and centrifuged again at 20000 rpm for 20min at 4°C. The supernatant was discarded, and the dry pellet was used for mass spectrometry analyses.
Chromatin Conformation Capture
Chromosome conformation capture (3C) was performed basically as previously described in (Ariel et al., 2020) starting with 2g of seedlings. Digestions were performed overnight at 37°C with 400U DpnII (NEB). DNA was ligated by incubation at 16°C for 5h in 4 ml volume using 100U of T4 DNA ligase (NEB). After reverse crosslinking and Proteinase K treatment (Invitrogen), DNA was recovered by Phenol:Chloroform:Isoamilic Acid (25:24:1; Sigma) extraction and ethanol precipitation. Relative interaction frequency was calculated by qPCR (primers used are listed in Supplementary Table 3). A region free of DpnII was used to normalize the amount of DNA.
Author contributions
MM, JMP, LL, CFF, JRM, NM, AC, MB and FA performed the experiments. JB, MB, FI, MC, JE and FA analyzed the data. JE and FA conceived the project. FA, JE and CFF wrote the manuscript with the contribution of all authors.
Acknowledgments
We thank Chang Liu for providing the Hi-C plot of Figure 2B. This work was supported by grants from ANPCyT (PICT2016-0132 and PICT2017-0066) and Instituto Milenio iBio – Iniciativa Científica Milenio, MINECON to JME; ANPCyT (PICT2016-0007 and PICT2016-0289) and Fima Leloir Award to FA; UNL (CAI+D 2016) to LL; Laboratoire d’Excellence (LABEX) Saclay Plant Sciences (SPS; ANR-10-LABX-40) to MC; and CNRS (Laboratoire International Associé NOCOSYM) to MC and FA. LL, FI, JME and FA are researchers of CONICET; MM, JMP, CFF and JRM are fellows of the same institution. NM is a fellow of ANPCyT.
Footnotes
Several improvements has been added in the figures and text
References




