

Abstract
In the chloroplasts of photosynthetic organisms, the conversion of light energy into chemical energy through photosynthesis is essential for photoautotrophic growth. However, the conversion of excess light energy into thermal energy by non-photochemical quenching (NPQ) is important to avoid reactive oxygen species generation and maintain efficient photosynthesis. In the unicellular green alga Chlamydomonas reinhardtii, NPQ as a photoprotective mechanism is activated through wavelength-specific light signaling pathways mediated by the phototropin (blue light) and UVR8 (ultra-violet light, UV) photoreceptors. NPQ-dependent photoprotection improves cell survival under high-light conditions; however, the biological significance of photoprotection activated by different light qualities remains poorly understood. Here, we demonstrate that NPQ-dependent photoprotection is activated more rapidly by UV than visible light. We found that the induction of gene and protein expression related to photoprotection is significantly faster and greater in magnitude under UV treatment compared to that under blue- or red-light treatment. Furthermore, the action spectrum of UV-dependent induction of photoprotective factors implies that Chlamydomonas senses UV-A, whereas the model dicot plant Arabidopsis thaliana senses UV-B for induction of photoprotective responses. Therefore, we hypothesize that Chlamydomonas uses UV-A for the induction of preemptive photoprotection prior to the photoprotection response activated by visible light in the hydrosphere.
Introduction
Light absorption is the fundamental process of photosynthesis. Green photosynthetic organisms such as land plants and green algae possess light-harvesting complexes (LHC) in their photosynthetic apparatus that efficiently capture light energy1,2. This efficient light-harvesting system is advantageous under relatively weak light; however, under high-light (HL) conditions, excess light energy absorbed by LHCII potentially results in the formation of reactive oxygen species3, leading to photoinhibition4. Excess light energy absorption by photosynthetic organisms is compensated by a response that protects photosynthesis called non-photochemical quenching (NPQ)5,6.
NPQ is the mechanism through which excess light energy is dissipated, and NPQ is controlled by the photoprotective proteins LHC stress-related (LHCSR) and/or PSBS7. In vascular plants, a mutant lacking PSBS is deficient in NPQ activation under HL conditions8. LHCSR and PSBS effectively produce NPQ in the moss Physcomitrella patens; the corresponding mutants lacking either protein exhibit reduced rates of energy dissipation9. Similarly, the unicellular green alga Chlamydomonas reinhardtii possesses PSBS and two LHCSRs, namely LHCSR1 and LHCSR37. Chlamydomonas mutants lacking either LHCSR1 or LHCSR3 cannot survive under HL due to insufficient activation of NPQ10,11. Moreover, the molecular functions of LHCSR proteins in P. patens and Chlamydomonas have been clarified as energy quencher12–15 and/or energy distributor among photosystems16. In summary, each of these photoprotective factors has been confirmed to contribute to NPQ in green photosynthetic organisms.
In Chlamydomonas, LHCSRs are reported to be light inducible11,17. The blue-light receptor phototropin is essential for effective LHCSR3 gene and protein expression under HL18. Moreover, the ultra-violet (UV) light receptor UVR8 can initiate UV-dependent expression of LHCSR1 and PSBS genes and proteins10,19. Considering that both LHCSR1 and LHCSR3 has been associated with NPQ10,11, UV-induced activation of LHCSR1-dependent NPQ might have distinct significance compared with blue light-induced activation of LHCSR3-dependent NPQ.
Although UV and blue light are both clearly involved in the expression of photoprotective factors, the biological significance of different light wavelengths inducing different photoprotective factors in Chlamydomonas remains poorly understood. Previous studies showed that preacclimation to UV enabled Chlamydomonas survival under subsequent HL treatment, whereas cells not previously exposed to UV were severely bleached following HL treatment10,20. These results imply that UV-dependent activation of photoprotection in Chlamydomonas functions as “preemptive photoacclimation” before the “subsequent photoacclimation” enabled by HL-dependent photoprotection in nature.
To further evaluate this hypothesis, we characterized the molecular and physiological responses associated with UV-and visible light-dependent activation of photoprotection. Previous research showed that 16 hours of UV pre-acclimation was necessary for survival under subsequent HL treatment10; therefore, it is difficult to predict the advantages of UV-dependent activation of photoprotection in nature. Here, we present wavelength-specific differences in Chlamydomonas photoprotection activation kinetics. Under different monochromatic light conditions, experimental Chlamydomonas strains showed clear differences in both gene expression and NPQ activation kinetics. UV-dependent photoprotection was activated significantly faster than that activated by other light qualities. Further analysis revealed that UV-dependent photoprotection is indispensable for HL tolerance in strains lacking LHCSR3, the photoprotective factor predominantly activated via bluelight perception.
Results and Discussion
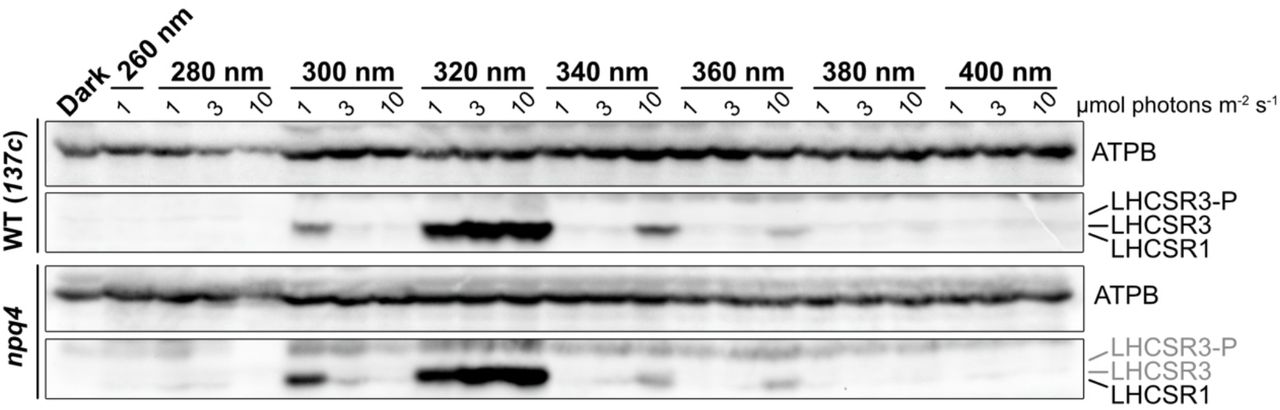
Although photoprotection in Chlamydomonas is reported to be activated via blue-light phototropin and UV-UVR8 signaling10,18, the precise wavelengths that trigger gene expression associated with photoprotection remain unclear. To investigate this, we first analyzed LHCSR1 and LHCSR3 protein accumulation in wild-type (WT) and npq4-mutant Chlamydomonas strains, the latter lacking LHCSR311, grown under 300–725 nm strong monochromatic light applied using a Okazaki Large Spectrograph (OLS)21. After 4 hours illumination with monochromatic light, subsequent immunoblot analysis of WT cells showed distinct profiles of LHCSR1 and LHCSR3 proteins (Figure 1). As expected, high-level LHCSR1 accumulation was observed in cells grown under light in the UV region (325–350 nm), whereas LHCSR3 accumulation was observed in response to growth under UV-A/blue (375–500 nm) and red (625–675 nm) light. UV-A-specific LHCSR1 accumulation was also observed in the npq4 mutant, confirming that the LHCSR protein signals observed reflected LHCSR1 accumulation rather than LHCSR3.

Protein was extracted from samples of wild-type (WT; 137c) and npq4-mutant strains. The cells were maintained in darkness (Dark) or treated with 100 μmol photons/m2/s of white light (WL) or different wavelengths of monochromatic light, as indicated, for 4 hours. Antibodies against ATPB or LHCSRs (recognizing both LHCSR1 and LHCSR3) were used for immunoblotting analysis. Shown are representative immunoblots from one of three replicated experiments, each performed using different biological samples.
LHCSR protein accumulation was not observed in WT or npq4 strains under monochromatic 300-nm UV illumination. The lower level of protein accumulation under 300-nm UV also extended to the loading control protein ATPB. Since strong UV light is known to induce photodamage of PSII via disruption of the Mn cluster in oxygen-evolving complexes22, the observed reduced protein accumulation under 300-nm UV may be attributed to cell death caused by strong photoinhibition.
To evaluate the action spectrum of LHCSR1 accumulation, we next irradiated the WT strain with monochromatic 300–340-nm UV light of relatively weak intensity (1 μmol photons/m2/s). Following 4-hour illumination, the action spectra of LHCSR1 protein accumulation (Figures 2A and S1) and NPQ induction (Figure 2B) exhibited peaks at 315–320 nm, suggesting that Chlamydomonas photoprotection activation is responsive to UV at the boundary of UV-B (280–315 nm) and in the UV-A region (315–400 nm). In line with the dynamics of protein levels shown in Figure 1, LHCSR1 protein was less abundant under 300-nm UV illumination. Cells exposed to 300-nm UV showed significantly lower photosynthetic activity, as indicated by the chlorophyll fluorescence parameter Fv/Fm (Figure 2C). Taken together with the results shown in Figure 1, Chlamydomonas cells seem to be severely photodamaged by 300-nm monochromatic UV light, which potentially results in cell death.

Protein was extracted from cells of wild-type (WT; 137c) and npq4-mutant strains. The cells were treated with different wavelengths of monochromatic UV light, as indicated, applied at 1, 3, and 10 μmol photons/m2/s for 4 hours, and compared to cells maintained in darkness as a control. Antibodies against ATPB or LHCSRs (recognizing both LHCSR1 and LHCSR3) were used for immunoblotting analysis. Shown are representative immunoblots from one of three replicated experiments, each performed using different biological samples.

Wild-type cells were treated with 1 μmol photons/m2/s of monochromatic UV light of different wavelengths, as indicated, for 4 hours. (A) Antibodies against ATPB or LHCSRs (recognizing both LHCSR1 and LHCSR3) were used for immunoblotting analysis. Shown are representative immunoblots from one of three replicated experiments, each performed using different biological samples. (B and C) Photosynthetic (B) and photoprotective (NPQ; C) activities of cells from A as measured by a FluorCAM system. Data are means ± SE, n = 3 biological replicates.
The action spectrum of LHCSR1 protein accumulation reported here is distinct from the UVR8 UV-absorption action spectrum in plants, which displays a peak in UV-B and UV-C (260–280 nm)23,24. This indicates that UV-response features differ between plants and green algae, which we presume to be the result of differences between their habitats. Land plants are frequently exposed to harmful UV-B/C, whereas green algae are mainly exposed to UV-A because UV-B/C is rapidly quenched (absorbed) in the hydrosphere25,26. Such habitat differences suggest that green algae developed a UV response activated by UV-A instead of UV-B/C.
To determine the biological significances of UV-A/B perception in Chlamydomonas, we next investigated the activation kinetics of photoprotective responses under UV (310–330 nm), blue (470 nm), and red (660 nm) light. We first analyzed protein accumulation associated with photoprotective factors and NPQ activity. To evaluate wavelength-dependent photoprotection kinetics, the WT strain was irradiated with the above-mentioned light qualities for 240 min, and cell samples were harvested at distinct time points as indicated in Figure 3. Subsequent immunoblot analysis again showed distinct patterns of LHCSR1 and LHCSR3 accumulation under different light qualities (Figure 3A). In agreement with the OLS action spectrum shown in Figure 1, LHCSR1 protein mainly accumulated under UV light, whereas LHCSR3 accumulated under blue and red light (Figure 3A).

(A) Wild-type cell samples were collected after 15-, 30-, 60-, 120-, and 240-min irradiation with three different qualities of light, as indicated, and compared to control samples maintained in low light (LL) for the duration of the experiment. LHCSR1 and LHCSR3 protein levels were detected using an antibody against LHCSRs (recognizing both LHCSR1 and LHCSR3). The ATPB protein detected using a specific antibody was used as a loading control. Shown are representative immunoblots from one of three replicated experiments, each performed using different biological samples. (B) Nonphotochemical quenching activities of cells from A as measured by a FluorCAM system. Data are means ± SE, n = 3 biological replicates.
Interestingly, NPQ activation kinetics were faster and NPQ was induced to a greater extent in UV-irradiated cells compared to that in blue or red light-irradiated cells (Figure 3B). Although LHCSR protein accumulation was not detectable within 15–30 min of UV irradiation, NPQ activity at these time points was much higher in UV-illuminated cells compared to that in blue or red light-treated cells, the latter of which seemed to correlate with LHCSR3 protein accumulation. These results imply that, in addition to activating LHCSR accumulation, UV treatment activates other photoprotection-related molecules such as PSBS. Considering that PSBS rapidly and temporarily accumulated before LHCSRs during activation of photoprotection27–29, it is plausible that rapid UV-dependent activation of NPQ involves both PSBS and LHCSRs in Chlamydomonas.
Since UV-A/B-dependent signal transduction is independent of photosynthesis10,30, it is possible that the expression kinetics of UV-A/B-induced photoprotective genes is much faster than that controlled by retrograde signaling via photosynthesis under HL exposure in the visible part of the spectrum18. To further investigate whether the prompt photoprotective response of Chlamydomonas under UV light increases with increasing photoprotection-associated gene expression, we analyzed the expression of genes encoding LHCSRs and PSBS over the same time course as used for the NPQ analysis shown in Figure 3. LHCSR1 and PSBS1 genes encoding UV-inducible photoprotective factors were immediately induced and reached maximum expression levels after 15–30 min UV treatment (Figure 4, UV), whereas blue or red light-treated cells showed slower induction kinetics of LHCSR3.2 gene expression (Figure 4, 470 nm and 660 nm). Moreover, LHCSR1 and PSBS1 expression was almost undetectable in red light-treated cells (Figure 4, 660 nm). These data imply there are differences in expression kinetics between genes encoding NPQ-associated photoprotective factors that are induced by either UV (LHCSR1 and PSBS1) or visible light (LHCSR3.2)18,27,31. Together with the activation kinetics of photoprotective responses shown in Figure 3, these gene expression data indicate that UV is the most effective light quality for prompt NPQ activation in Chlamydomonas.

RNA was extracted from wild-type cell samples collected after 15-, 30-, 60-, 120-, and 240-min irradiation with three different qualities of light, as indicated. The expression levels of PSBS1, LHCSR1, and LHCSR3.2 genes related to photoprotection were semi-quantitatively analyzed by RT-PCR. The CBLP gene was used as a loading control. Shown are representative gels from one of three replicated experiments, each performed using different biological samples.
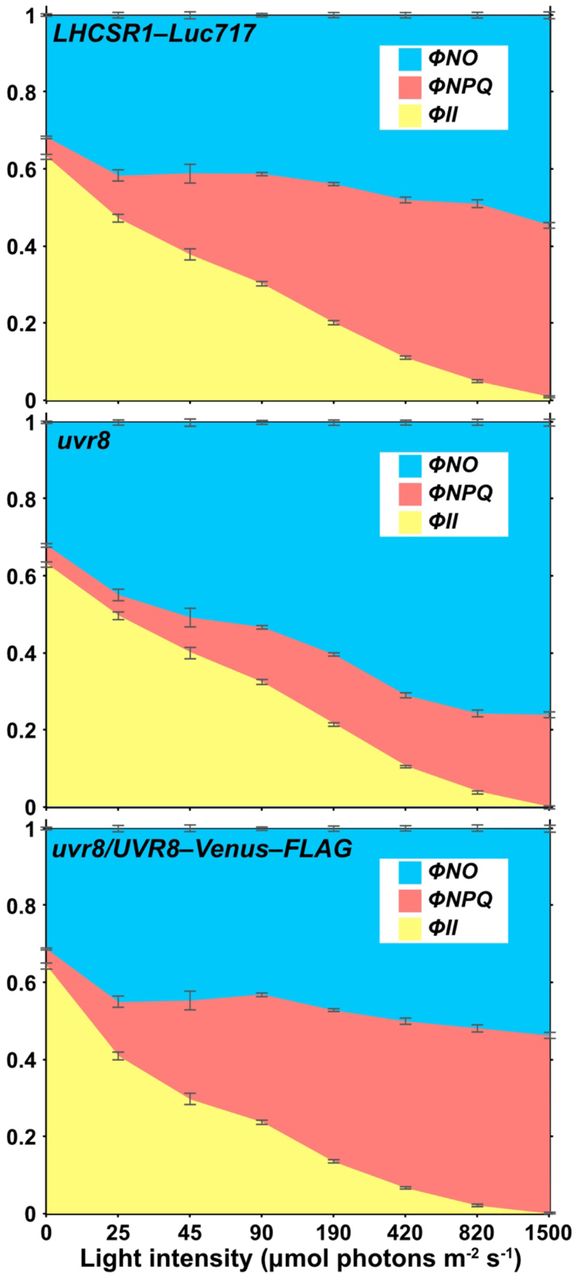
Concerning how photosynthetic organisms recognize UV light, the UVR8 photoreceptor is capable of perceiving UV-B in both land plants32 and Chlamydomonas10. Using a previously obtained uvr8 mutant30, we performed semi-quantitative RT-PCR analysis to evaluate whether UVR8 was responsible for UV-inducible, photoprotection-associated gene expression in Chlamydomonas. In line with a previous study10, the uvr8 strain showed reduced expression of UV-induced photoprotective components, which was reflected in both mRNA and protein levels (Figure 5). Moreover, the UV-dependent rapid activation of photoprotection was severely reduced in uvr8 cells, leading to a significantly lower NPQ light-response curve compared to the WT strain, even under low light (LL, ~50 μmol photons/m2/s; Figure 6). We also confirmed that the observed uvr8 phenotypes were complemented by UVR8, which was fused with the yellow fluorescent protein variant Venus and a FLAG epitope tag (Venus–FLAG) and over-expressed in the uvr8 strain (Figures 6 and S2). These results further confirm that the UV-B photoreceptor UVR8 is responsible for the UV-dependent rapid activation of NPQ observed here.

The quantum efficiency of internal energy relaxation of Chl (ϕNO), non-photochemical quenching (ϕNPQ), and PSII photochemistry (ϕII) calculated from the dataset used in Figure 6 are presented in a stacked bar chart. The top, middle, and bottom chart represents the parameters of the control (LHCSR1–Luc717), uvr8-mutant, and complemented (uvr8/UVR8–Venus–FLAG) strains, respectively. Data are means ± SE, n = 3 biological replicates.

RNA and protein were extracted from samples of control (CS; LHCSR1–Luc717) and uvr8-mutant strains illuminated with three different qualities of light, as indicated, for 1 hour and compared to control samples maintained in low light (LL). (A) The expression levels of PSBS1, LHCSR1, and LHCSR3.2 genes related to photoprotection were semi-quantitatively analyzed by RT-PCR. The CBLP gene was used as a loading control. (B) LHCSR1 and LHCSR3 protein levels were detected using an antibody against LHCSRs (recognizing both LHCSR1 and LHCSR3). The ATPB protein was used as a loading control. Shown are representative gels and immunoblots from one of three replicated experiments, each performed using different biological samples.

Cells of control (LHCSR1–Luc717), uvr8-mutant, and complemented (uvr8/UVR8–FLAG) strains were treated with UV light for 1 hour to induce photoprotective mechanisms, and compared to control samples maintained under low light (LL). (A) The UVR8–Venus–FLAG and UVR8 proteins were detected using antibodies against FLAG and CrUVR8, respectively. LHCSR1 and LHCSR3 protein levels were detected using an antibody against LHCSRs (recognizing both LHCSR1 and LHCSR3). The ATPB protein was used as a loading control. (B) Non-photochemical quenching was recorded by a moni-PAM system (Walz). The light curve was set at 0, 45, 90, 190, 420, 820, and 1,500 μmol photons/m2/s. Glycolaldehyde at 10 mM final concentration was added 3 min before the measurements. Data are means ± SE, n = 3 biological replicates.
Our data establishes that UVR8-dependent activation of photoprotection is promptly established under low-level UV illumination (Figure 4). To clarify whether this UV-dependent rapid activation of photoprotection is of biological significance under subsequent HL treatment, we next evaluated the chlorophyll bleaching phenotypes of WT and mutant Chlamydomonas strains. All strains were UV treated for 1 hour to induce rapid activation of photoprotection before subsequent low-level UV-supplemented HL treatment. Although the uvr8 strain showed less NPQ compared to WT (Figures 6 and S2), it did not show chlorophyll bleaching under the HL conditions applied here (Figure 7). The photoprotection-compromised, LHCSR3-lacking npq4 strain was also tolerant to our HL conditions and showed negligible chlorophyll bleaching (Figure 7). However, the npq4 uvr8 double-mutant strain exhibited significant HL-induced chlorophyll bleaching, implying that UV-dependent rapid activation of NPQ is indispensable under the HL conditions applied here in the absence of LHCSR3.

(A) Wild-type (WT; 137c), control (CS; LHCSR1–Luc717), and uvr8-, npq4-, and npq4 uvr8-mutant strains in a 24-well plate after treatment with low light (LL; 30 μmol photons/m2/s) or high light (HL; 500 μmol photons/m2/s) containing UV for 10 hours. Cell concentration was normalized at 5 × 106 cells/mL. Shown are representative sample pictures from one of three replicated experiments, each performed using different biological samples. (B) Chlorophyll contents of samples from A were normalized to cell number. Data are means ± SE, n = 3 biological replicates.
In the present study, we reveal that the green alga Chlamydomonas has a distinct UV-response compared to that of land plants. The Chlamydomonas UV response is initiated by relatively long-wavelength UV including that in the UV-A region (Figures 1 and S1), whereas the land-plant UV response is initiated by UV-B32–34. It should be noted that although there is a difference in wavelength responsiveness, both Chlamydomonas and land-plant UV responses activate acclimation mechanisms that suit environmental light conditions20,23. The difference in wavelength responsiveness may be attributed to differing natural habitats. Chlamydomonas primarily inhabits the hydrosphere, wherein the majority of UV radiation is in the UV-A region because UV-B is easily absorbed by dissolved or suspended organic/inorganic compounds in the water column26. Plants on land, by contrast, are exposed to more harmful UV-B35. This habitat difference may explain why green photosynthetic organisms have developed a UV response to UV-A in the hydrosphere and to UV-B in terrestrial environments. We anticipate that further insight into the UV-response strategies of these photosynthetic organisms will improve our understanding of plant evolution.
Materials and Methods
Algal strains and growth conditions: Chlamydomonas reinhardtii 137c (wild type; WT) strain was obtained from the Chlamydomonas Center (https://www.chlamycollection.org/). The npq4 strain was isolated in previous report11 and was then backcrossed with the WT strain for several times16. DSR1 (uvr8) and DSR1-comp (uvr8/UVR8–Venus–FLAG) strains were generated in previous studies19,30. LHCSR1–Luc717 strain harboring a reporter construct expressing a LHCSR1-Luciferase fusion30, was used as a control strain (CS) for DSR1. For generating double-mutant strains harboring npq4 and uvr8 mutations, the npq4 and DSR1 strains were crossed. All strains and mutants were grown in tris-acetate-phosphate (TAP) medium. Strains were grown under 50 μmol/m2/s light (Osram FL40SS D/37 white-light) at 25°C for all experiments. Once grown, cells were harvested and resuspended at 2 × 106 cells/mL in high-salt (HS) medium36, modified to include 20 mM MOPS and K2HPO4/KH2PO4 at an altered concentration of 1 mM. Cell resuspensions were then subject to experimental light treatments as described in the text and individual figure legends.
Standard mRNA quantification: Total RNA from light-treated cells was extracted with the Maxwell RSC instrument (Promega) equipped with the Maxwell RSC simplyRNA Tissue Kit (Promega). The isolated RNA was quantified with the QuantiFluor RNA System (Promega) prior to reverse transcription. Reverse transcription and semi-quantitative PCR were carried out with the ReverTra Ace qPCR RT kit with gRemover (TOYOBO) and the KOD FX Neo DNA polymerase (TOYOBO) on the SimpliAmp Thermal Cycler (ThermoFisher Scientific). For semi-quantitative RT-PCR, the gene encoding the G protein ß-subunit-like polypeptide (CBLP) was chosen as the housekeeping gene during light treatment. The primers used for the semi-quantitative RT-PCR were as described in previous studies19,30.
Immunoblotting: Protein samples of whole-cell extracts (corresponding to ~2.0 × 106 cells, unless stated otherwise) were loaded on 11% SDS-PAGE gels containing 7 M urea and blotted onto nitrocellulose membranes. Antiserum against ATPB was obtained from Agrisera (AS05 085, rabbit polyclonal); antiserum against LHCSRs (recognizing both LHCSR1 and LHCSR3) was raised and affinity-purified against the peptide LGLKPTDPEELK as reported previously30; antiserum against UVR8 was raised and affinity purified against the peptide MGPDDMGTAGDSRD (Eurofins Genomics); antiserum against FLAG fusion proteins was obtained from Sigma-Aldrich (F1804, mouse monoclonal). An anti-rabbit horseradish peroxidase-conjugated antiserum (#7074, Cell Signaling Technology) or an anti-mouse horseradish peroxidase-conjugated antiserum (#330, MBL Lifescience) was used as secondary antibody. The blots were developed with ECL detection reagent EzWestLumi plus (ATTO) and images of the blots were obtained using a CCD imager ChemiDocTouch System (Bio-Rad Laboratories). The upper LHCSR3 band represents the phosphorylated form of LHCSR318.
Chlorophyll fluorescence-based photosynthesis analysis: For measurement of NPQ activation kinetics (Figure 4B), minimum (Fo) and maximum (Fm) fluorescence yield in darkness were measured with a FluorCAM (Photon System Instruments) after weak far-red (<5 μmol photons/m2/s) treatment for 30 min. Maximum fluorescence yield in light (Fm’) was measured following subsequent actinic irradiation at 750 μmol photons/m2/s for 30 sec. For measurement of the light response curve (Figure 6B), Fo and Fm were measured with a Moni-PAM system (Walz) after weak far-red (<5 μmol photons/m2/s) treatment for 30 min. Fm’ at different light intensities were measured following subsequent actinic irradiation at 0, 45, 90, 190, 420, 820, and 1,500 μmol photons/m2/s for 2 min. Glycolaldehyde at a final concentration 10 mM was added 3 min prior to the measurements to mimic carbon limitation. Photosynthetic parameters were calculated as follows:

Wavelength-dependent photoprotection assay: WT cells in HS media were irradiated with low-level UV-supplemented fluorescence light (using a ReptiSun10.0 UV fluorescent bulb30 at 10 μmol photons/m2/s, Figure S3), 470-nm LED light, or 660-nm LED light at 110 μmol photons/m2/s. The total light intensities were measured by a sun spectroradiometer (S-2442 HIDAMARI mini, SOMA OPTICS, LTD.) with a range of 300 to 800 nm. Total cellular protein extracts were obtained from culture samples taken following 15, 30, 60, 120, and 240 min of light treatment.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
The light emission spectra of the 300–800 nm UV light source used in the present study as obtained by a sun spectroradiometer (S-2442 HIDAMARI mini, SOMA OPTICS, LTD.).
High-light tolerance assay: All algal strains in HS media were pretreated with low-level UV-A/B for 1 hour. Cells were then irradiated with either low light (LL; white fluorescence light at 30 μmol photons/m2/s) or UV-supplemented high light (HL; 500 μmol photons/m2/s) for 10 h. Total cellular protein extracts were obtained from culture samples taken following 1, 2, 4, 6, and 8 h of light treatment. Chlorophyll amounts and cell numbers were determined by Porra calculation37 and a TC20 automated cell counter (Bio-Rad Laboratories), respectively. The calculated chlorophyll amounts were normalized to total cell number (pg of chlorophyll per cell). For culture photos, LL- or HL-treated strains were adjusted to 5 × 106 cells/mL and transferred into a multi-well plate to be photographed.
Author contributions
RT designed the research. RT performed transcriptional, biochemical, and pigment bleaching analyses. RT, TY, and KF-K generated the DSR1 mutant. RT and KO analyzed the chlorophyll fluorescence quenching of the alga. RT wrote the manuscript. JM supervised the research and provided the resources. All authors contributed to revise the manuscript and approved the final version of the manuscript.
Acknowledgements
We thank Mr. Tamaki Uchikawa and Ms. Maki Kondo for technical support in OLS experiments. Dr. Yasuhiro Kamei is thanked for fruitful discussion about UV photoreceptors. We also thank Mrs. Tamaka Kadowaki for providing technical assistance with genetic crossing of the alga. This work was supported by JSPS KAKENHI (Grant Numbers JP15H05599 and JP20H03282 to RT, and JP16H06553 to JM), the Nakajima Foundation to RT, and NINS program for cross-disciplinary study (Grant Number 01311701 to RT). This study was carried out under the NIBB Cooperative Research for the Okazaki Large Spectrograph (g-16-705).
References




