

Abstract
Infection by severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), the etiological agent of coronavirus disease 2019 (COVID-19), may elicit uncontrolled and damaging inflammatory reactions, due to an excessive immune response and dysregulated production of cytokines and chemokines. Thus, it is critical to identify compounds able to inhibit virus replication and thwart the excessive inflammatory reaction and tissue lesions secondary to SARS-CoV-2 infection. Here, we show that the neuropeptides VIP and PACAP, molecules endowed with immunoregulatory properties, were able to inhibit SARS-CoV-2 RNA synthesis/replication in human monocytes and viral production in lung epithelial cells. VIP and PACAP protected these cells from virus-induced cytopathicity, reduced the production of proinflammmatory mediators, and prevented the SARS-CoV-2-induced NF-kB activation, which is critically involved in the production of inflammatory mediators. Both neuropeptides promoted CREB activation in infected monocytes, a transcription factor with antiapoptotic activity and also a negative regulator of NF-kB. As a possible host response to control patient inflammation, we identified that VIP levels were elevated in plasma from patients with severe forms of COVID-19, correlating with the inflammatory marker CRP and survival on those patients. Since a synthetic form of VIP is clinically approved in Europe and under two clinical trials for patients with COVID-19, our results provide the scientific evidence to further support clinical investigation of these neuropeptides against COVID-19.
Introduction
Individuals with coronavirus disease 2019 (COVID-19), caused by the severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2)1, may present asymptomatic or mild disease to severe lung inflammation and the life threatening acute respiratory distress syndrome (ARDS)2,3. Severe SARS-CoV-2 infection is characterized by elevated serum levels of proinflammatory mediators (hypercytokinemia, also known as cytokine storm) such as, for example, IL-2, IL-6, TNF-α, IL-8, IL-1b, IFN-γ2–5. The excessive immune response and the dysregulated production of cytokines and chemokines are hallmarks of SARS-CoV-2 infection and have been pointed as the main cause of the severe lung damage and unfavorable clinical progression of patients with COVID-193–6.
During the inflammatory response to human pathogenic coronaviruses (hCoVs), circulating monocytes migrate and infiltrate the lungs7,8 and there become the main producers of proinflammatory cytokines, exacerbating the disease9. In addition, MERS-CoV-and SARS-CoV-infected macrophages produce high levels of pro-inflammatory cytokines and chemokines10,11. More recently, Giamarellos-Bourboulis et al6 showed that lung monocytes from severe pneumonia caused by SARS-CoV-2 are potent producers of TNF-α and IL-6, whose levels were increased in the serum of the same patients.
Given that SARS-CoV-2 infection may trigger an uncontrolled and damaging inflammatory reaction, which might favor the virus propagation, it is critical to identify agents able to prevent the infection and concurrently thwart the prototypical excessive inflammatory reaction and tissue lesions secondary to SARS-CoV-2 infection. In this work, we evaluated whether the neuropeptides Vasoactive Intestinal Peptide (VIP) and Pituitary Adenylate Cyclase-Activating Polypeptide (PACAP), which present immunoregulatory functions12 and are also able to reduce viral production in HIV-1-infected macrophages13,14, can present the antiviral and anti-inflammatory activities needed to attenuate the damages caused by SARS-CoV-2 infection.
The neuropeptides VIP and PACAP, which are members of the Secretin/Glucagon family of peptides, share many biological properties through their interaction with the G protein-coupled receptors VPAC1, VPAC2 and PAC115. PACAP binds to all three receptors, with higher affinity to PAC1, while VIP interacts preferentially with VPAC1 and VPAC215,16. VIP and PACAP are produced by Th2 CD4+ and CD8+ T cells17–19, and their receptors are expressed by a variety of cell types, including T cells, macrophages and dendritic cells15.
VIP and PACAP have well-characterized regulatory effects on the immune system and anti-inflammatory properties, including control of cell activation and differentiation, down-regulation of inflammatory cytokines and reactive oxygen species and induction of the anti-inflammatory cytokine IL-1020–25. Based on their consistent anti-inflammatory activities, both neuropeptides have been considered as promising therapeutic agents for autoimmune disorders and chronic inflammatory illnesses12,26,27. In this paper, we report that both neuropeptides are able to inhibit SARS-CoV-2 gene replication in human monocytes and viral production in Calu-3 cells, and to reduce the cellular production of proinflammmatory mediators that have been described to contribute for the severe tissue damage occurring in the course of COVID-19.
Materials and Methods
Cells, virus and reagents
African green monkey kidney cells (Vero, subtype E6) and human lung epithelial cell lines (Calu-3) were expanded in high glucose DMEM (Vero) or MEM (Calu-3) with 10% fetal bovine serum (FBS; HyClone), with 100 U/mL penicillin and 100 μg/mL streptomycin (Pen/Strep; Gibco) at 37°C in a humidified atmosphere with 5% CO2.
Peripheral blood mononuclear cells (PBMCs) were isolated by density gradient centrifugation (Ficoll-Paque, GE Healthcare) from buffy-coat preparations of blood from healthy donors. PBMCs (2 × 106 cells) were plated onto 48-well plates (NalgeNunc) in RPMI-1640 with 5% inactivated male human AB serum (Merck) for 3 hours. Non-adherent cells were removed and the remaining monocytes were maintained in DMEM (low-glucose) with 5% human serum (Merck) and 100 U/mL penicillin and 100 μg/mL streptomycin (Pen/Strep; Gibco). The purity of human monocytes was above 90%, as determined by flow cytometric analysis (FACScan; Becton Dickinson) using anti-CD3 (BD Biosciences) and anti-CD14 (BD Biosciences) monoclonal antibodies.
SARS-CoV-2 was expanded in Vero E6 cells from an isolate contained on a nasopharyngeal swab obtained from a confirmed case in Rio de Janeiro, Brazil (GenBank accession no. MT710714). Viral isolation were performed after a single passage in a cell culture in a 150 cm2 flasks with high glucose DMEM plus 2% FBS. Observations for cytopathic effects were performed daily and peaked 4 to 5 days after infection. All procedures related to virus culture were handled in a biosafety level 3 (BSL3) multiuser facility according to WHO guidelines. Virus titers were determined as plaque forming units (PFU/mL). Virus stocks were kept in −80°C ultralow freezers. VIP (Vasoactive intestinal peptide), PACAP (Pituitary adenylate cyclase-activating polypeptide), and the VPAC1 agonist (Ala11,22,28)-VIP and VPAC2 agonist Bay 55-9837 were purchased from Tocris. The PAC1 agonist, Maxadilan was kindly donated to us by Dr. Ethan A. Lerner (Department of Dermatology, Massachusetts General Hospital, MA, USA).
Infections and virus titration
Infections were performed with SARS-CoV-2 at MOI of 0.01 (monocytes) or 0.1 (Calu-3) in low (monocytes) or high (Calu-3) glucose DMEM without serum. After 1 hour, cells were washed and incubated with complete medium with treatments or not. For viral titration, monolayers of Vero cells (2 × 104 cell/well) in 96-well plates were infected with serial dilutions of supernatants containing SARS-CoV-2 for 1 hour at 37°C. Cells were washed and 1.2% carboxymethyl cellulose (CMC)/DMEM (high glucose) semisolid medium with 2% FBS was added. At 3 days post-infection, plaque number was scored in at least 3 replicates per dilution by independent readers. The reader was blind with respect to source of the supernatant.
Molecular detection of virus RNA levels
The total RNA was extracted from cultures using QIAamp Viral RNA (Qiagen®), according to manufacturer’s instructions. Quantitative RT-PCR was performed using QuantiTect Probe RT-PCR Kit (Quiagen®) in an ABI PRISM 7500 Sequence Detection System (Applied Biosystems). Amplifications were carried out in 25 µL reaction mixtures containing 2× reaction mix buffer, 50 µM of each primer, 10 µM of probe, and 5 µL of RNA template. Primers, probes, and cycling conditions recommended by the Centers for Disease Control and Prevention (CDC) protocol were used to detect the SARS-CoV-2 (CDC 2020). The standard curve method was employed for virus quantification. For reference to the cell amounts used, the housekeeping gene RNAse P was amplified. The Ct values for this target were compared to those obtained to different cell amounts, 107 to 102, for calibration.
Measurements of Inflammatory Mediators, Cell Death, NF-kBp65, CREB and neuropeptides
The levels of IL-6, IL-8, TNF-α and MIF were quantified in the monocyte supernatants from infected and uninfected cells by ELISA (R&D Systems), following manufacturer’s instructions, and results are expressed as percentages compared to uninfected cells controls. Cell death was determined according to the activity of lactate dehydrogenase (LDH) in the culture supernatants using a CytoTox® Kit (Promega, USA) according to the manufacturer’s instructions. Results are expressed as percentages of released LDH compared to control cells lysed with 0.8% Triton X-100. Supernatants were centrifuged at 5,000 rpm for 1 minute, to remove cellular debris. Evaluation of NF-kBp65 and CREB activation was performed in monocytes, infected or not, according to manufacturer’s instructions: NFkB p65 (Total/Phospho) InstantOne™, CREB (Total/Phospho) Multispecies InstantOne™ ELISA Kits (Thermo Fisher). VIP and PACAP were quantified in plasma from patients or control volunteers, using standard commercially available ELISA and EIA Kits according to the manufacturer’s instructions (Abelisa).
Human subjects
We prospectively enrolled severe or mild/asymptomatic COVID-19 RT-PCR-confirmed cases, and SARS-CoV-2-negative controls. Blood and respiratory samples were obtained from the 24 patients with severe COVID-19 within 72 hours from intensive care unit (ICU) admission in two reference centers (Instituto Estadual do Cérebro Paulo Niemeyer and Hospital Copa Star, Rio de Janeiro, Brazil). Severe COVID-19 was defined as those critically ill patients, presenting viral pneumonia on computed tomography scan and requiring oxygen supplementation through either a nonrebreather mask or mechanical ventilation. Four outpatients presenting mild self-limiting COVID-19 syndrome, and two SARS-CoV-2-positive asymptomatic subjects were also included. All patients had SARS-CoV-2 confirmed diagnostic through RT-PCR of nasal swab or tracheal aspirates. Peripheral vein blood was also collected from 11 SARS-CoV-2-negative control participants as tested by RT-PCR on the day of blood sampling. The characteristics of severe (n = 24), mild/asymptomatic (n = 6) and control (n = 10) participants are presented in Table 1. Mild and severe COVID-19 patients presented differences regarding the age and the presence of comorbidities as obesity, cardiovascular diseases and diabetes (Table 1), which is consistent with previously reported patient cohorts2,28–30. The SARS-CoV-2-negative control group, however, included subjects of older age and chronic non-communicable diseases, so it is matched with mild and critical COVID-19 patients, except for hypertension (Table 1). All ICU-admitted patients received usual supportive care for severe COVID-19 and received respiratory support with either noninvasive oxygen supplementation (n=5) or mechanical ventilation (n= 19) (Supplemental Table 1). Patients with acute respiratory distress syndrome (ARDS) were managed with neuromuscular blockade and a protective ventilation strategy that included low tidal volume (6 mL/kg of predicted body weight) and limited driving pressure (less than 16 cmH2O) as well as optimal PEEP calculated based on the best lung compliance and PaO2/FiO2 ratio. In those with severe ARDS and PaO2/FiO2 ratio below 150 despite optimal ventilatory settings, prone position was initiated. Our management protocol included antithrombotic prophylaxis with enoxaparin 40 to 60 mg per day. Patients did not receive routine steroids, antivirals or other anti-inflammatory or anti-platelet drugs. The SARS-CoV-2-negative control participants were not under anti-inflammatory or anti-platelet drugs for at least two weeks. All clinical information was prospectively collected using a standardized form – ISARIC/WHO Clinical Characterization Protocol for Severe Emerging Infections (CCPBR). Clinical and laboratory data were recorded on admission in all severe patients included in the study and the primary outcome analyzed was 28-day mortality (n = 11 survivors and 13 non-survivors, Supplemental Table 2). Age and the frequency of comorbidities were not different between severe patients requiring mechanical ventilation or noninvasive oxygen supplementation neither between survivors and non-survivors (Supplemental Table 1 and 2).
Ethics statement
Experimental procedures involving human cells from healthy donors were performed with samples obtained after written informed consent and were approved by the Institutional Review Board (IRB) of the Oswaldo Cruz Foundation/Fiocruz (Rio de Janeiro, RJ, Brazil) under the number 397-07. The National Review Board approved the study protocol (CONEP 30650420.4.1001.0008), and informed consent was obtained from all participants or patients’ representatives.
Statistical analysis
Statistics were performed using GraphPad Prism software version 8. All the numerical variables were tested regarding their distribution using the Shapiro-Wilk test. One-way analysis of variance (ANOVA) was used to compare differences among 3 groups following a normal (parametric) distribution, and Tukey’s post-hoc test was used to locate the differences between the groups.
Comparisons between 2 groups were performed using the Student t test for parametric distributions or the Mann-Whitney U test for nonparametric distributions. Correlation coefficients were calculated using Pearson’s correlation test for parametric distributions and the Spearman’s correlation test for nonparametric distributions.
Results
VIP and PACAP inhibit SARS-CoV-2 RNA synthesis in human primary monocytes, protecting them from virus-mediated cytophatic effects
Based on our previous findings showing that VIP and PACAP present antiviral effects on HIV-1 infection13,14, and on their ability to control the inflammatory response, we initially investigated whether both neuropeptides could inhibit viral life cycle in SARS-CoV-2-infected monocytes. Thus, human primary monocytes were treated with either VIP or PACAP, then infected with SARS-CoV-2 and the replication of viral genome was measured 24 hours later. We found that VIP significantly reduced the SARS-CoV-2 RNA synthesis, achieving 33% and 45% inhibition at 5 nM and 10 nM, respectively (Fig. 1A). PACAP similarly inhibited levels of viral RNA synthesis with 5 nM (40%) and 10 nM (39%) (Fig. 1A). In parallel, we observed that VIP and PACAP, at 10 nM each, completely protected the SARS-CoV-2-mediated cytophatic effect, as measured by the LDH levels in the cell culture supernatant (Fig. 1B). These results show that both neuropeptides are similarly effective in their ability to reduce SARS-CoV-2 genome replication in monocytes and to prevent the virus-induced cell death.

(A) Monocytes were exposed to the indicated concentrations of VIP or PACAP during 30 min, and then infected with SARS-CoV-2. Virus RNA synthesis was evaluated by qPCR in the culture supernatants 20-24 hours after infection. (B) Cellular viability was analyzed by measuring LDH release in the supernatants of uninfected or SARS-CoV-2-infected monocytes, treated or not with VIP or PACAP. (C) Similar to (A), except that cells were treated with agonists for VIP and PACAP receptors, as indicated, at different concentrations. Data in A and C are shown normalized to infected cells kept only with culture medium, and in C represent means ± SD of absolute values. *p ≤ .05; **p ≤ .01.
Receptor contribution for the VIP and PACAP mediated inhibition of SARS-CoV-2 replication
Because VIP binds the receptors VPAC1 and VPAC2, and PACAP has high affinity for PAC1, but also binds the VPAC1 and VPAC215,16, we analyzed the role of the individual receptors in the neuropeptide-mediated inhibition of SARS-CoV-2 RNA replication. To this end, monocytes were treated with specific agonists to VPAC1, VPAC2 and PAC1 (Ala-VIP, Bay 55 9837 and Maxadilan, respectively), and then exposed to SARS-CoV-2. We found that the sole activation of VPAC2 at 1 nM, 5 nM and 10 nM, and of VPAC1 at 10 nM, significantly reduced the SARS-CoV-2 gene replication (Fig. 1C). The ligation of only VPAC1 at 1 and 5 nM, and of PAC1 at 5 and 10 nM also diminished the viral replication, without reaching, however, statistical significance. Of note, the optimal concentrations of the receptor agonists that significantly decreased the SARS-CoV-2 RNA synthesis were similar to those of the natural receptor ligands VIP and PACAP. Overall, these findings suggest that the three receptors contribute for the VIP- and PACAP-mediated SARS-CoV-2 inhibition, and that the sole activation of each of these receptors can lead to a diminished viral inhibition similar as that induced by the own neuropeptides.
VIP and PACAP inhibit SARS-CoV-2 replication in Calu-3 cells and protect them from virus-mediated cytophatic effects
We next evaluated whether VIP and PACAP could also be able to restrict the virus production in pulmonary cells, one of the major targets of SARS-CoV-2. Thus, Calu-3 cells, a lineage of epithelial lung cells highly susceptible to this virus, were infected with SARS-CoV-2 and treated with a range of concentrations of either peptide. We found that VIP decreased the productive infection, reaching 41% and 28% of inhibition with 1 nM and 5 nM, respectively (Fig.2A). PACAP also diminished virus production by 33%, 35% and 42% at concentrations equivalent to 5 nM, 10 nM and 50 nM (Fig. 2B). The different potency between VIP and PACAP in their ability to control the viral productive infection in Calu-3 cells might be explained by the relative abundance of the neuropeptide receptors, since it has been shown that these cells express only VPAC131. However, all three receptors are reported to be expressed in lungs, with some studies showing that VPAC1 levels are much higher than VPAC2 (the other VIP receptor) or PAC1 (to which PACAP binds with higher affinity than to VPAC1 and VPAC215,32–34). Likewise we found for monocytes, VIP and PACAP VIP also protected Calu-3 cells from the SARS-CoV-2-induced cytopathogenic effects, at the same optimal viral inhibitory concentrations (1 nM and 50 nM, respectively) of each neuropeptide (Fig. 2C).

Calu-3 cells were infected by SARS-CoV-2 and then left untreated (0) or treated with the indicated concentrations of VIP (A) or PACAP (B). After 60 hours, supernatants were collected and viral replication was evaluated by quantifying PFUs in plaque assay. (C) Cellular viability was analyzed by measuring LDH release in the supernatants of uninfected or SARS-CoV-2-infected Calu-3 cells, treated or not with VIP (1 nM) or PACAP (50 nM). (D, three bars on the left) SARS-CoV-2-infected Calu-3 cells were kept untreated (Nil) or received CM (50% final dilution) from VIP- or PACAP-treated uninfected monocytes. (D, three bars on the right) SARS-CoV-2-infected Calu-3 cells were kept untreated (Nil) or received CM (50% final dilution) from VIP- or PACAP-treated SARS-CoV-2-infected monocytes. After 60 hours, supernatants were collected and viral replication was evaluated by quantifying PFUs in plaque assay. Data are shown as means ± SD. *p ≤ .05; **p ≤ .01.
Given that monocytes and epithelial cells can be in close proximity in the lungs, and, thus, can interact and exchange molecules, we investigated whether conditioned medium (CM) obtained by SARS-CoV-2-infected monocytes exposed to VIP or PACAP could repress the viral production in human lung cells. While the transference of CM from uninfected monocytes, treated or nor with either neuropeptide, did not modify the virus growth, the addition of CM (at 50% vol/vol – the combination of fresh medium at this ratio did not altered the basal replication of SARS-CoV-2 in Calu-3 cells, Supp. Fig. 1) from infected monocytes, treated with VIP or PACAP, to Calu-3 cells, resulted in a 50% reduction of virus replication in these cells (Fig. 2D). This finding raises the hypothesis that both neuropeptides, besides inhibiting the SARS-CoV-2 gene replication, induce monocyte production and release of factors endowed with antiviral activity, which may increase the resistance of neighboring cells to SARS-CoV-2 growth. This bystander effect can reduce viral propagation, thus eventually protecting the tissues from lesions secondary to SARS-CoV-2-replication and dissemination. Future studies are needed to identify the nature of these putative soluble factors.
VIP and PACAP reduce production of proinflammatory cytokines by SARS-CoV-2-infected monocytes and Calu-3 cells
Given the hypothesis that controlling the production of proinflammatory cytokines may be critical for reducing SARS-CoV-2 replication and limiting tissue damages, and based on evidence that VIP and PACAP can regulate the inflammatory response24,35, we next evaluated whether both neuropeptides could attenuate the production of proinflammatory mediators by SARS-CoV-2-infected monocytes or lung epithelial cells. As shown in Fig. 3A, SARS-CoV-2-infected monocytes produced large amounts of the proinflammatory mediators IL-6, IL-8, TNF-α and MIF relative to uninfected cells (15, 4, 12 and 18 times more, respectively). In contrast, the treatment of monocytes with either neuropeptide after SARS-CoV-2 infection reduced to 66%, 50%, 66% and 50% the cellular production of IL-6, IL-8, TNF-α and MIF, respectively. Furthermore, VIP and PACAP reverted by approximately the same degree the release of IL-6 and IL-8 by Calu-3 cells (Fig. 3B), implying that VIP and PACAP may offer a critical protection to inflamed lungs affected by SARS-CoV-2 replication. Because proinflammatory cytokines may favor SARS-CoV-2 replication, which, in turn, can amplify the cellular synthesis of these mediators, these findings may support our assumption that VIP and PACAP offer tissue protection by inhibiting virus replication and regulating the boost of cytokine production.
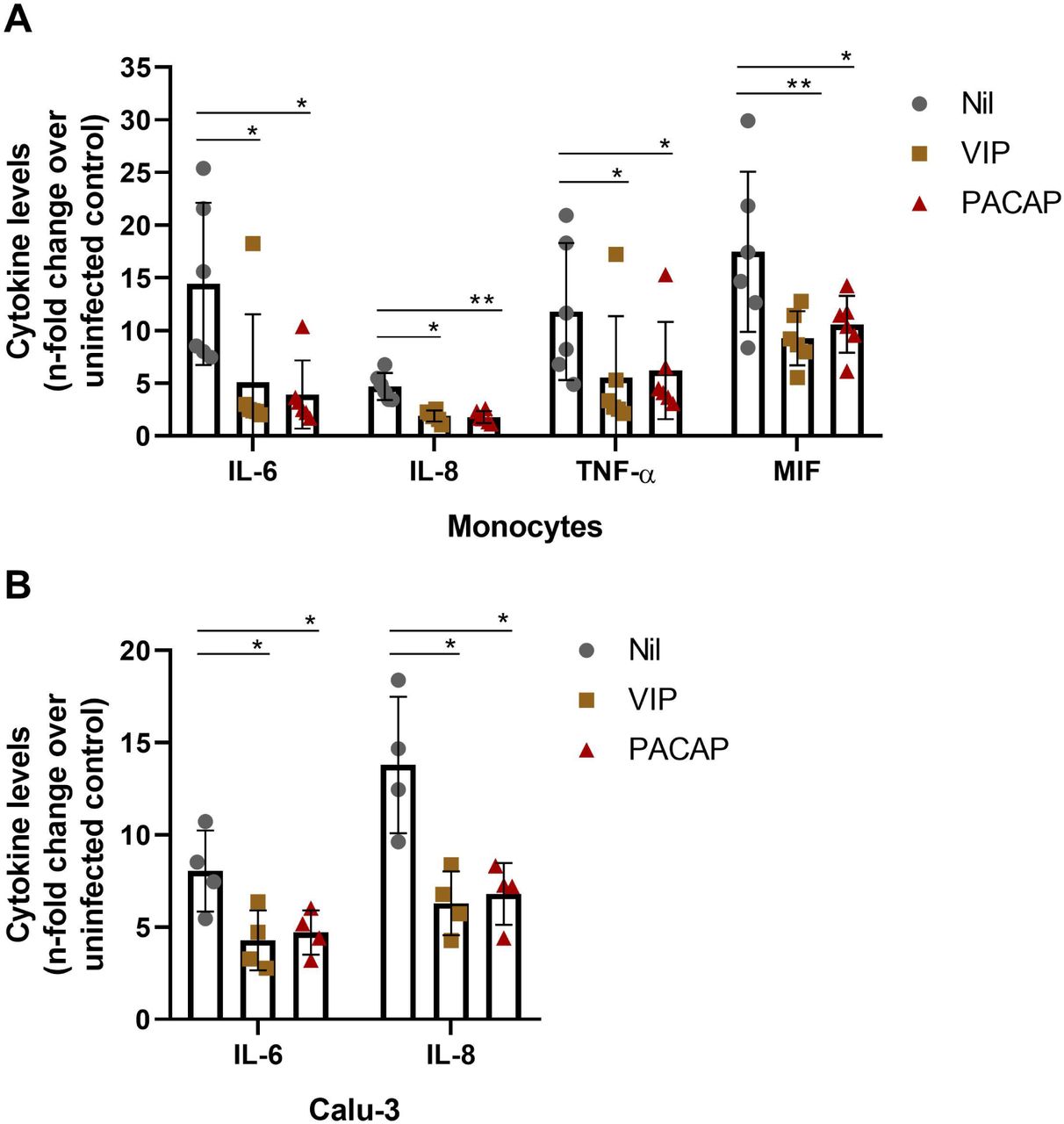
Monocytes and Calu-3 cells were infected with SARS-CoV-2 for 1 hour, and then exposed to VIP or PACAP (10 nM each for monocytes, 1 nM of VIP or 50 nM of PACAP for Calu-3 cells). The levels of IL-6, IL-8, TNF-α and MIF were measured in culture supernatants of monocytes after 24 hours (A), and of IL-6 and IL-8 after 60 hours for Calu-3 cells (B), by ELISA. Data represent means ± SD. *p ≤ .05; **p ≤ .01.
VIP and PACAP regulate the activation of transcription factors in SARS-CoV-2-infected monocytes
Based on the fact that the transcription factor NF-kB is critically involved in the cellular production of inflammatory mediators36, and on our own findings that VIP and PACAP can inhibit its activation in HIV-1-infected macrophages14, we investigated whether both neuropeptides would exert this same activity in SARS-CoV-2-infected monocytes. As can be seen in Fig. 4A, we firstly observed that NF-kB is up-modulated in infected cells (as measured by the increased amount of phosphorylated NF-kBp65 subunit) and, second, that VIP and PACAP reduced the NF-KBp65 phosphorylation, evidencing the significant anti-NF-kBp65 activity of VIP and PACAP. We believe that the SARS-CoV-2-induced NF-kB activation could have contributed for the elevated production of proinflammatory cytokines by SARS-CoV-2-infected monocytes, and that the inhibition of this transcription factor by VIP and PACAP was critical for the decreased synthesis of these mediators by the same cells.

Monocytes were infected by SARS-CoV-2 and, 1 hour after infection, the infected cells were exposed (or not – Nil) to VIP or PACAP (at 10 nM) and 24 hours later, cells were lysed and the ratios between phosphoNF-kBp65 / total NF-kBp65 (A) and phosphoCREB / total CREB (B) were quantified by ELISA in the cell lysates. “Control” indicates uninfected monocytes kept only with culture medium. Data in A and B represent means ± SD. *p ≤ .05; **p ≤ .01; ***p ≤ .001.
We also analyzed the effects of both neuropeptides on the activation of CREB, a transcription factor induced by several GPCRs ligands, including VIP and PACAP37. CREB and NF-kB share the CREB-binding protein/p300 (CBP/p300 protein) as a cofactor, and CREB activation results in the inhibition of NF-kB38. We initially observed that activation of CREB was diminished in SARS-CoV-2-infected monocytes, a finding coherent with NF-kB activation in the same cells (Fig. 4B). Consistent with this finding, VIP and PACAP promoted CREB activation (as measured by augmentation of CREB phosphorylation) in those infected monocytes, a result matching the inhibition of NF-kB and the reduction of cellular production of proinflammatory cytokines.
Plasma levels of VIP are elevated in patients with severe forms of COVID-19
Based on our results showing that VIP and PACAP, besides inhibiting viral replication and production, can also control the production of proinflammatory mediators by SARS-CoV-2-infected monocytes, we measured the levels of both neuropeptides in plasma samples of SARS-CoV-2-infected individuals. We found that the amount of VIP is significantly elevated in patients affected by the most severe forms of infection, relative to levels of uninfected healthy controls and asymptomatic/mild patients (Fig. 5A), and that some critically ill patients had increased levels of PACAP, but without statistical significance when considering the entire group (Fig. 5B). In addition, levels of VIP positively correlated with levels of C-reactive protein (Fig. 5C), but neither VIP nor PACAP levels correlated with other inflammatory markers, such as D-dimer, fibrinogen or IL-6 (data not shown), or with each other (Fig. 5D). Severe COVID-19 patients admitted to the ICU were further stratified between those requiring invasive mechanical ventilation or noninvasive O2 supplementation, or according to the 28-day mortality outcome as survivors or non-survivors, and, while for PACAP we did not observed statistical significance in the stratified groups (Fig. 5E and 5F), we detected that VIP plasma levels were significantly higher in survivors than in non-survivors (Fig. 5H).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
The levels of VIP (A) and PACAP (E) in the plasma of SARS-CoV-2-negative control participants, SARS-CoV-2-positive asymptomatic subjects, or symptomatic patients presenting mild to severe COVID-19 syndrome were quantified by ELISA. Correlation between VIP and CRP (B) and between levels of both neuropeptides (F) in the plasma from patients with COVID-19. (C and G) Severe COVID-19 patients admitted to the ICU were stratified between those requiring invasive mechanical ventilation or noninvasive O2 supplementation. (D and H) Severe COVID-19 patients were stratified according to the 28-day mortality outcome as survivors or nonsurvivors. One outlier (CRP = 621.64 mg/dL) was excluded from the analysis in panels B-D according to the ROUT test. Linear regression (with the 95 % confidence interval) and Spearman’s correlation were calculated according to the distribution of the dots (controls: grey; asymptomatics: brown; mild: green; severe: red). The horizontal lines in the box plots represent the median, the box edges represent the interquartile ranges and the whiskers indicate the minimal and maximal value in each group. *p ≤ .05; **p ≤ .01.
Taking into account that VIP and PACAP regulate inflammatory reactions, it is possible that their increased circulating amounts reflect a counter-regulatory effect elicited by the dysregulated immune response typical of the more severe clinical status of COVID-19 patients. The levels of VIP and PACAP in SARS-CoV-2-infected tissues should be a matter of future studies, to substantiate our hypothesis.
Discussion
In this work, we report that the neuropeptides VIP and PACAP, endogenous molecules presenting anti-inflammatory properties, are able to inhibit the SARS-CoV-2 RNA synthesis and viral production (in monocytes and human lung cells), and reduce the exacerbated synthesis of proinflammatory mediators due to infection. Also we identified that VIP was elevated in patients with severe form of COVID-19, correlating with levels of CRP and that the elevated levels also correlated with survival. Our findings attain great relevance, given that VIP (as a synthetic form, Aviptadil), in an intravenous39 and inhaled40 formulation, is under two clinical trials for COVID-19 patients with respiratory failure, whose first results are expected to be disclosed in September39/November40 this year. Clinical data, along with mechanism for antiviral inhibition, may allow larger trials.
We provide here experimental evidence showing that VIP and PACAP encompass critical activities for preventing the tissue damage caused by SARS-CoV-2 infection, i.e, inhibiting viral replication, maintaining the cellular viability, and controlling the overproduction of proinflammatory cytokines. These findings may support the assumption that the association of anti-inflammatory agents with antivirals should be suitable for COVID-19 treatment. In this sense, the recent reported administration of dexamethasone to patients with COVID-19 resulted in a significant clinical benefit for those affected with the more severe forms of this disease41.
VIP and PACAP are expressed in the central and peripheral nervous system and in many other sites, including the lymphoid tissue17–19 and the lungs15,42, where, together with brain and gut, the higher levels of VIP are found15,42. Both neuropeptides regulate the inflammatory response due to their ability to decrease the production of proinflammatory mediators, such as chemokines, reactive oxygen and nitrogen species, IL-1β, IL-6, IL-12, TNF-α and MIF, and to elicit the production of anti-inflammatory molecules, such as IL-4 and IL-1021–24. The receptors for VIP and PACAP, the G protein-coupled receptors VPAC1, VPAC2 and PAC1, are expressed in cells of the immune system and largely distributed in other tissues15. The systemic distribution of both neuropeptides and their receptors explain the pleiotropic effects of VIP and PACAP. While both neuropeptides have been extensively studied in chronic inflammatory illnesses, reports of their effects on infectious processes are scarce, and, specifically for viral infections, we reported that VIP and PACAP inhibit HIV-1 replication in human primary macrophages13,14.
We initially observed that VIP and PACAP decreased the genome replication of SARS-CoV-2 in monocytes, in parallel with protecting the cells from virus-induced cytopathicity. While at this moment it is unclear the mechanism by which both neuropeptides reduce the SARS-CoV-2 gene replication, it is possible to speculate that, by diminishing the intracellular levels of viral RNA and other viral molecules, VIP and PACAP could prevent the cell death by pyroptosis, which has been described as one of the main causes of cell damage during SARS-CoV-2 infection43. While maintaining the monocyte viability, VIP and PACAP also diminished the production of the proinflammatory cytokines IL-6, IL-8, TNF-α and MIF by these cells, a result which is in agreement with the reported ability of these neuropeptides to regulate the inflammatory response21–24,44. The ability of both neuropeptides to inhibit SARS-CoV-2 replication, protect cells from virus-mediated cytophaticity and reduce cellular production of inflammatory mediators was also extended to lung epithelial cells, a finding that further support the assumption that VIP and PACAP may offer a critical protection to inflamed lungs affected by SARS-CoV-2 replication. Because these mediators have been found to be elevated in patients with COVID-19, and that VIP and PACAP regulate inflammatory reactions, it is possible that the higher amounts of VIP in patients affected by the most severe forms of infection may reflect a counter-regulatory feedback elicited by the dysregulated immune response typical of the more severe clinical status of COVID-19 patients.
Interestingly, the cellular protection conferred by VIP or PACAP to SARS-CoV-2-infected monocytes can be transferred to other neighboring virus target cells, as we observed that conditioned medium from neuropeptide-treated infected-monocytes diminished the productive infection in Calu-3 lung epithelial cells. Thus, we understand that this bystander effect can ultimately protect other SARS-CoV-2-infected cells not directly interacting with VIP or PACAP. Our findings warrant more investigations to identify the nature of the protecting factors presumably released by monocytes that survived the infection by SARS-CoV-2.
Our findings showing that VIP and PACAP inhibited the SARS-CoV-2-induced activation of the subunit p65 of the transcription factor NF-kB, which is a classical inducer of inflammatory mediators36, pertain to a complex molecular mechanism probably participating in the neuropeptide-mediated reduction of proinflammatory cytokines by SARS-CoV-2-infected monocytes. As we also previously described for HIV-1 infection in macrophages14, the ability of VIP and PACAP to prevent the SARS-CoV-2-mediated NF-kBp65 activation might be associated to a concomitant neuropeptide-induced CREB activation. In fact, we detected here that the transcription factor CREB, which can act as a negative regulator of NF-kB45,46, is down-regulated in SARS-CoV-2-infected monocytes, in opposition to NF-kB activation in the same cells. In some models47–52, CREB activation is related to induction of anti-inflammatory cytokines concomitant with reduction of pro-inflammatory molecules, mainly through direct transcription of its targets, like IL-10, or through competing with NF-kB by their shared co-activador protein CBP/p30045,46,52,53. Besides its role in the balance between pro- and anti-inflammatory cytokines, CREB activation is also involved with the anti-apoptotic response in monocytes and macrophages, during differentiation and inflammatory stimuli54,55. Thus, the imbalance between CREB and NF-kB, either as a direct effect of infection by SARS-CoV-2 or a consequence of exposure of bystander cells to viral products and inflammatory molecules, could be an important target for inhibition of SARS-CoV-2 deleterious effects, at least in monocytes and probably also in lung cells, as a similar imbalance between CREB and NF-kB was observed in an acute inflammatory pulmonary condition38.
Quite important for the understanding of the VIP and PACAP anti-SARS-CoV-2 effects and for COVID-19 pathogenesis is the definition that the engagement of the individual VIP and PACAP receptors by specific agonists recapitulate the actions of the native neuropeptides in monocytes. Because VIP and PACAP signaling pathway outcome is dependent of the combined action of the receptors activated by them (VIP and PACAP receptors can elicit cell signaling in homo and hetero dimers56, it is possible that all receptors are involved in the final outcome analyzed. Our assays suggest that the three receptors may have contributed for VIP- and PACAP-mediated reduction of SARS-CoV-2 RNA synthesis in monocytes. This finding is particularly interesting for VPAC1, whose solely activation in HIV-1-infected cells enhanced viral production in human primary macrophages13 and in CD4+ tumor cell lines and PBMCs57. The inhibition profile of SARS-CoV-2 by VIP and PACAP in Calu-3 cells may be biased regarding the expected action in the lungs, since Calu-3 cells appear to express only VPAC131. However, lung tissues, while reported to express high levels of VPAC1, also express VPAC2 and PAC132,34, and, more specifically, VPAC2 mRNA was detected in airway epithelial, glandular, and immune cells of the lung33. Therefore, while the inhibition curve of SARS-CoV-2 by VIP and PACAP in Calu-3 cells point to different optimal doses than those obtained for monocytes, it is possible that in normal lung cells and tissue, VIP and PACAP could present a broader range of action in the inhibition of SARS-CoV-2. In fact, VIP and specific agonists for VPAC1 or VPAC2 have been proposed and tested for respiratory conditions, like asthma58–60, pulmonary arterial hypertension (PAH)58,61,62 and chronic obstructive pulmonary disease (COPD)58,59,63, demonstrating that the anti-inflammatory actions of VIP/PACAP can be achieved in lung tissues. Future studies should define which of these receptors would preferentially be activated by specific agonists to restrain SARS-CoV-2 replication in lungs or other sites. Also, the loss of VIP capacity to inhibit virus production at the more elevated concentrations in Calu-3 cells might involve receptor desensitization, a common feature for GPCRs, which is also described for VIP/PACAP receptors64,65. Together with the possible differences of receptor expression and self-regulatory characteristics of GPCRs, a third regulation level of VIP and PACAP action on pulmonary cells can be achieved by the activity of proteases and peptidases, as lungs are described to express high levels of several of them in both normal and pathological conditions66–68. Some of these peptidases could target VIP and PACAP, thus altering the ligand/receptor ratio and modulating the signaling pathways. Studies are underway in our laboratory to verify whether the combination of these agonists could potentiate their individual effects.
It has been described that SARS-CoV-2 propagates throughout the organism, thus causing inflammatory damages in several tissues4. Since VIP and PACAP, and their receptors are systemically distributed, we suppose that the antiviral and anti-inflammatory effects of both neuropeptides would not be restricted to the respiratory tract. Just as an example, VIP and PACAP have been shown to protect nervous tissues form injuries caused by a number of neurotoxic agents69. This neuroprotective effects might be particularly important in the light of new indication that COVID-19 patients can be affected by a range of neurological lesions and disorders70.
Because up to now there are no antivirals specific to SARS-CoV-2, and that the hyper-inflammation may persist even after the lowering of the viral load, the searching for compounds that target the aberrant production of proinflammatory cytokines and, simultaneously, the own viral replication, should be stimulated. Our present results showing that VIP and PACAP hold these two critical activities point these neuropeptides or their analogue molecules as potential therapeutic agents for COVID-19.
Author Contribution
Conceived the study: JRT, TMLS, DCBH; Designed the experiments: JRT, PTB, TMLS, DCBH; Performed the experiments: JRT, CQS, NFR, CRRP, CSF, SSGD, ACF, MM, VCS, LT, IGAQ, PK; Analyzed the data: JRT, PTB, PK, FAB, TMLS, DCBH; Wrote the paper: JRT, PTB, TMLS, DCBH. All authors reviewed and approved the manuscript.
Acknowledgments and funding
We thank the Hemotherapy Service from Hospital Clementino Fraga Filho (Federal University of Rio de Janeiro, Brazil) for providing buffy-coats. Dr. Andre Sampaio from Farmanguinhos, platform RPT11M, and Dr. Lucio Mendes Cabral from Department of Drugs and Pharmaceutics, Faculty of Pharmacy, Federal University of Rio de Janeiro (UFRJ) are acknowledged for kindly donating the Calu-3 cell. The recombinant protein Maxadilan was kindly donated to us by Dr. Ethan A. Lerner (Department of Dermatology, Massachusetts General Hospital, MA, USA). The authors are thankful to Prof. Elvira M. Saraiva (Federal University of Rio de Janeiro, Brazil) for stimulating comments and invaluable suggestions. This work was supported by Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), Fundação de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ), and by Mercosur Fund for Structural Convergence (FOCEM, Mercosur, grant number 03/11). This study was financed in part by the Coordenação de Aperfeicoamento de Pessoal de Nível Superior – Brasil (CAPES), Finance Code 001. Funding was also provided by CNPq, CAPES and FAPERJ through the National Institutes of Science and Technology Program (INCT) to Carlos Morel (INCT-IDPN) Wilson Savino (INCT-NIM). Thanks are due to Oswaldo Cruz Foundation/Fiocruz under the auspicious of Inova program. The funding sponsors had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, and in the decision to publish the results.
References




