

Abstract
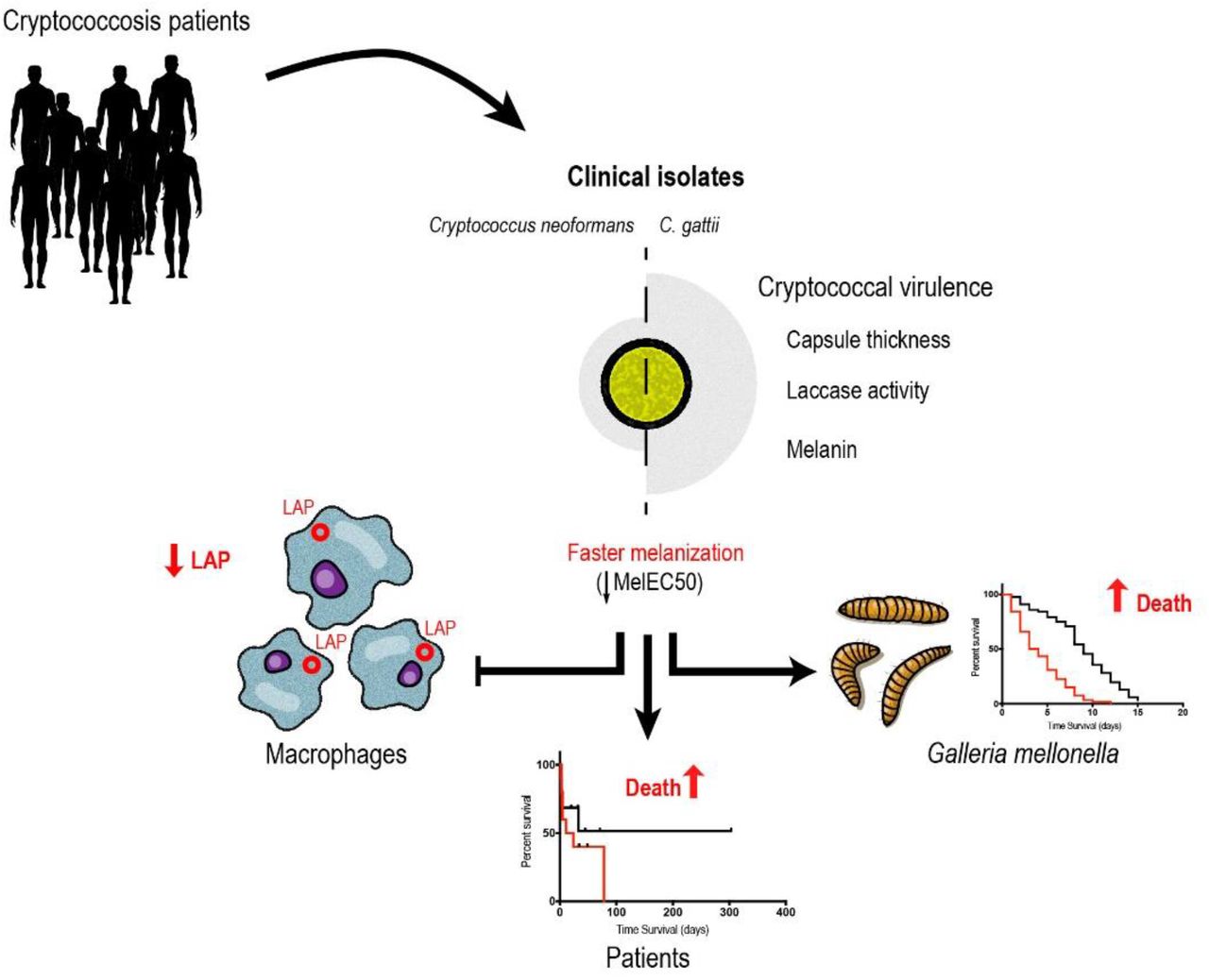
Cryptococcus spp. are important human pathogens responsible for about 180,000 deaths per year. Studying their virulence attributes can lead to better cryptococcosis prevention and treatment. In this work, we systematically investigated virulence attributes of Cryptococcus spp. clinical isolates and correlated them with patient data. We collected 66 C. neoformans and 19 C. gattii isolates from Brazilian patients and analyzed multiple phenotypes related with their capsule, production of laccase, melanin and extracellular vesicles. We also tested their virulence in Galleria mellonella and ability to evade macrophage LC3-associated phagocytosis (LAP). All phenotypes analyzed varied widely among the isolates, but C. neoformans isolates tended to melanize faster and more intensely and produce thinner capsules in comparison with C. gattii. We also observed correlations that match previous studies, such as that between secreted laccase – but not total melanin production – and disease outcome in patients. The most striking results, though, came from our measurements of Cryptococcus colony melanization kinetics, which followed a sigmoidal curve for most isolates. Faster melanization correlated positively with LAP evasion, virulence in G. mellonella and worse prognosis in humans. These results suggest that the speed of melanization, more than the total amount of melanin Cryptococcus spp. produces, is crucial for virulence.

Introduction
Cryptococcosis is estimated to cause 181.000 deaths per year, mostly in low and middle-income countries (1). Infection occurs through inhalation of spores or desiccated yeast cells of the species complexes Cryptococcus neoformans or C. gattii. In most cases, the disease happens in hosts with defective immunity secondary to AIDS, cancer or medication (2). The association of Amphotericin B and flucytosine is the gold standard treatment (3), but even in high income countries the case fatality rates are around 20%. In regions where these drugs are not available and fluconazole is the only choice, the case fatality rates can be higher than 60% (1). Combined with the fact that there are no vaccines, there is a clear need for more effective preventative and therapeutic options.
C. neoformans and C. gattii have several well-studied virulence factors such as the ability to produce melanin (4), the presence of a polysaccharide capsule (5), the ability to grow at 37 °C and the secretion of urease and other extracellular enzymes (6). Of these, the capsule and melanin contribute almost half of the C. neoformans virulence (7). The cryptococcal capsule is composed mainly of polysaccharides, such as glucuronoxylomannan (GXM). Its presence and thickness interfere with macrophage phagocytosis, and capsular polysaccharides interfere with the activation and differentiation of T cells (5, 8). Studies in mice and translational studies with cryptococcosis patient samples have shown the importance of GXM secretion by C. neoformans on central nervous system infections by C. neoformans (9, 10).
Melanin is a brown or black, hydrophobic, high molecular weight, negatively charged pigment. It is present in the cryptococcal cell wall and protects the yeast cell against host and environmental stresses (11). The pigment also increases resistance to Amphotericin B and affects susceptibility to fluconazole (12). In pathogenic species of the genus Cryptococcus, melanin production is dependent on laccase enzymatic action on biphenolic compounds (13). A translational study with C. neoformans clinical isolates demonstrated that laccase also has other roles that are crucial for fungal survival in the cerebrospinal fluid, and that isolates with more effective melanin-independent secreted laccase roles were associated with poorer patient outcomes (14).
Given that several tools presently used to prevent and treat infectious diseases target microbial virulence factors, we delved further into the roles played by capsule, melanin, laccase and extracellular vesicles in the interaction between Cryptococcus spp. and their host. The strategy consisted of collecting clinical isolates and patient data, characterizing the isolates in the laboratory and correlating the experimental results with patient outcomes. We found several correlations that confirm previous observations, but also important correlations between the melanization kinetics and outcomes of the interaction between Cryptococcus spp. and macrophages or G. mellonella. Most importantly, we also found that faster melanization, but not the final amount of melanin on cryptococcal colonies, correlated with the survival of HIV-positive patients with severe cryptococcosis. These findings help us better understand the mechanisms used by Cryptococcus spp. to survive and cause disease in their hosts.
Results
Clinical and epidemiological data
Cryptococcus spp. clinical isolates analyzed in this study are from two different sources. Twenty-eight (16 C. neoformans and 12 C. gattii) are from a cohort of patients treated in Rio de Janeiro, Brazil, about whom we have no clinical information. The second group of clinical isolates are from an ongoing epidemiological study in Brasília, Brazil. It contains 54 isolates (7 C. gattii and 47 C. neoformans) from 41 patients. From these, 37 were from the cerebrospinal fluid (CSF), 1 from a blood culture, 1 from a tissue biopsy and 2 from bronchoalveolar lavage fluid. Thirty-seven patients were infected by C. neoformans only, 2 by C. gattii only and the other 2 had C. neoformans/C. gattii mixed infections. Available information about them and the patients from which they were isolated can be found in Table S1.
All patients from the Brasília study (Table 1) were diagnosed and treated in public hospitals, according to the standards used by the services in which they were assisted. Most were male (68.3%), and their mean age was 42 years. HIV infection was reported in 68.3%; of the 12 HIV-negative patients, two had diabetes, three were using corticosteroids, one was using corticosteroids plus a second immunosuppressive drug, one had a primary immunodeficiency and eight had no known risk factor. Among the 39 patients that we were able to follow until death or hospital discharge, the 2-week and 10-week mortality rates were respectively 30.8% and 41%.
Melanization kinetics
The method we used to measure melanin production by each isolate quantitatively was based on a previously published protocol (15) (Figure S1A). To do so, we spotted a specific number of fungal cells in 24-well plates filled with solid medium containing the melanin precursor L-DOPA. These plates were photographed at regular intervals during incubation, and the resulting digital images processed to quantify how dark the colonies had become at each point in time. For most isolates, the resulting data fit very well in a sigmoidal curve. This method was highly reproducible, as shown by the similarity between the curves obtained from five independent experiments performed in different days with the control strain H99 (Figure S1B). As shown in Figures S1C-D, some isolates, such as H99, melanized faster and became black at the end of the experiment, whereas others, such as CNB017.1, melanized slowly and never became black. We also found differences in the pattern of colony melanization, such that some isolates (e.g. H99 and CNB017.1) showed homogeneously pigmented colonies whereas others had more intense melanization either in the periphery (CNB013.1) or in the center (CGF007) of the colony.
Using logistic regression, we quantitatively evaluated the kinetics of melanin production by each isolate. This regression resulted in five melanization parameters, three of which with a specific biological meaning:
Bottom – median gray level of the colony at the first time point.
Top – median gray level of the colony at the end of the experiment. Indicates how dark the colony becomes, and thus the final amount of melanin it produces.
Span: Top minus Bottom.
Hill Slope: steepness of the curve. Indicates how fast the colony produces melanin during the time in which melanization is occurring.
EC50: Time it takes for the colony to reach half of its final melanization intensity. Measures both how soon the colony starts melanizing and how fast it produces melanin once it has started.
A great variety was observed in the melanization parameters between isolates (Figure 1A-C), except for the melanization slope, which has a less dispersed frequency distribution (Figure 1D). To validate this new methodology, we compared its results with a semi-quantitative analysis we had previously done of 16 clinical isolates (Figure S2A). These isolates were grown in solid melanin-inducing medium and photographed every 12 h for 7 days. After cropping all photos of each colony together, we visually ranked them based on how fast they melanized and how dark they eventually became. The isolates were then given a score of 1 to 7: 1 for those with slowest melanization and less dark colonies and 7 for those with the highest rate of colony pigmentation. We found a significant correlation between this visual score and the logistic regression parameter Top (Figure S2B), indicating that our image analysis method matches the visual ranking. As expected, the visual melanization score also correlated directly with Span (r = 0.456) and Hill Slope (r = 0.226) and inversely with EC50 (r = −0.254), but these correlations were not statistically significant (p-values of 0.066, 0.379 and 0.321, respectively).

(A-D) Histograms showing the distribution of the melanization kinetics parameters from clinical isolates. Images of each colony taken throughout 168 h of incubation in melanizing medium were processed and fitted to sigmoidal curves to obtain: (A) melanization EC50 - time in hours for the colony to reach half maximum melanization; (B) melanization Top - median gray level of the colony at the end of the experiment, which indicates how dark the colony became; (C) melanization Span (difference in median gray levels of the colony at the beginning and end of the experiment); (D) melanization Slope (slope of the sigmoidal curve at the inflection point, an expression of how fast the colony melanizes. (E-F) Histograms showing the specific laccase activity from clinical isolates. Frequency distribution of secreted laccase activity, n = 82 and (F) Frequency distribution of whole-cell laccase activity, n = 84.
In addition to melanization, we also measured laccase activity both on washed whole cells (n= 84) and on the culture supernatants (n = 82). We observed greater dispersion in the frequency distribution of secreted laccase activity (Figure 1E) than whole-cell laccase activity (Figure 1F). Interestingly, the laccase activity in culture supernatants, but not on whole cells, correlated well with the visual melanization score (Figure S2C) and melanization EC50 (Figure S2D).
Clinical isolates demonstrate the variation of secreted GXM and capsule size in different culture media
To evaluate the capsule from each clinical isolate, we grew them in different media, photographed the cells with India ink and measured their capsule thickness. The media we used included Sabouraud, a rich medium in which the cryptococcal capsule is not induced, and three capsule inducing media: Sabouraud diluted 1:10 with MOPS pH 7.5 (Sab-MOPS), minimum medium (MM) and CO2-independent medium (CIM). The baseline capsule thickness in non-inducing medium varied from 0.5 to 5 μm for different isolates, although more than 90% of the clinical isolates had capsules 1 to 2 μm thick (Figure 2A). Representative pictures of clinical isolates at both extremes variation of capsule thickness is shown in Figure 2B. To determine the capacity for capsule induction, we measured the capsules for each isolate in each inducing medium and divided the value by that obtained in Sabouraud (Figure 2C-F). Sab-MOPS medium resulted in the greatest capsule induction, the most notable of which that of CNF016 (Figure 2 D). However, even in this medium some isolates such as CGB009.1 maintained a capsule thickness that was very similar to that in Sabouraud, indicating that different isolates may respond differently to the signals that induce capsule.

(A) Frequency distribution of the capsule thickness in Sabouraud medium (Sab), n = 48. (B) Representative photo of isolates at both ends of the capsule thickness distribution, CNF001.1 isolate less than 1 μm thick and CGF005 isolate greater than 5 μm. (C) Frequency distribution of capsule induction in CO2-independent medium (CIM) relative to Sabouraud (CIM / Sabouraud), n = 47. (D) Representative photo of isolates at both ends of the relative distribution, isolate CGB009.1, which maintained the same capsule thickness in both media, and isolate CNF016, whose capsule was about 3 times larger in Sab-MOPS media (n = 48) and (E) Frequency distribution of capsule induction in Sab-MOPS relative to Sabouraud (Sab-MOPS / Sabouraud). (F) Frequency distribution of capsule induction in Minimum Medium (MM) relative to Sabouraud (MM / Sabouraud), n = 48. (G) Frequency distribution of secreted GXM concentration, n = 46. Experiments were repeated at least twice and had similar results. Scale bars: 5 μm.
As capsular polysaccharides can be secreted in soluble form, we also used a capture ELISA to determine the concentrations of GXM on the culture supernatants of the clinical isolates. As observed with other virulence factors, the values we obtained varied widely across clinical isolates (Figure 2G).
Clinical isolates present different profiles of interaction with macrophages
Macrophages are crucial effector cells in the immune response to Cryptococcus spp. A specific type of autophagy, LC3 associated-phagocytosis (LAP), is important for the fungicidal activity of macrophages (16). As previous studies with Aspergillus fumigatus showed that melanin inhibited LAP (17, 18), which is important in immunity against C. neoformans (19), we quantified LAP in macrophage-like J774 cells infected with antibody-opsonized clinical isolates.
The infected J774 cells were processed for LC3 immunofluorescence microscopy and imaged. For each isolate, we evaluated the images to calculate two variables: the proportion of macrophages with LC3 recruitment to at least one phagosome containing C. neoformans and the proportion of macrophages where all internalized fungi were on LC3-positive phagosomes. Figure 3A shows representative immunofluorescence images with the two isolates that had the lowest (CNB020) and the highest (CNB042) proportion of LC3-positive phagosomes. In none of the isolates tested all internalized fungi were noticed in LC3-positive phagosomes (Figure 3B), possibly indicating that Cryptococcus spp. has mechanisms to avoid this type of macrophage response.

Representative photo of the autophagy assessment by immunofluorescence, measured by means of phagocytosis associated with LC3 (LAP). Clinical isolate CNB020 with low LAP induction and CNB042 with high LAP induction. (B) Frequency distribution of the proportion of J774 cells with LC3-positive phagosomes, n =24. Experiments were repeated at least twice in different days and had similar results. Scale bars: 10 μm.
C gattii isolates have larger capsules and more secreted laccase activity, but C. neoformans melanizes faster and more intensely
After systematically measuring virulence and host-pathogen interaction attributes, we began mining them for important insights into cryptococcal virulence. We observed important differences in the expression of virulence factors between C. neoformans and C. gattii. A larger proportion of C. neoformans isolates (23 out of 57) had a homogeneous colony melanization pattern, in comparison with just one out of 19 C. gattii isolates (p = 0.004, Fisher’s exact test). C. neoformans isolates also melanized faster (lower melanization EC50) and accumulated more melanin at the end of the experiment (higher melanization Top), with no differences on Hill Slope and Span (Figure 4 A-D). The two species also differed in laccase activity, with more secreted laccase activity in the supernatants – but not whole cells – of C. gattii isolates (Figure 4 E-F).

(A) Differences in melanization EC50 (time in hours for the colony to reach half maximum melanization); melanization HillSlope (slope of the sigmoidal curve at the inflection point, proportional to how fast the colony melanizes); melanization Span (difference in median gray levels of the colony at the beginning and end of the experiment - difference between top and bottom) and melanization Top (median gray level of the colony image at the end of the experiment - proportional to how dark the colony turned), C. gattii, n = 19 and C. neoformans n = 60. (B) Differences in secreted laccase activity (C. gattii, n = 18; C. neoformans, n = 64) and whole-cell laccase activity (C. gattii, n = 19; C. neoformans, n = 65). (C) Capsule thickness in Sabouraud medium (Sab) ratio with H99, induction in Sab-MOPS, Minimum Medium (MM) and independent CO2 relative to Sabouraud (C. gattii, n = 16; C. neoformans, n = 32). Experiments were repeated at least twice and had similar results. To compare the groups we used two-tailed t-test for independent samples.
C gattii isolates had thicker capsules than C. neoformans in non-inducing Sabouraud medium (Figure 4G). C gattii isolates also induced their capsules to a larger extent than C. neoformans in all capsule-inducing media, with the differences being statistically significant in all but minimal medium (Figure 4H-J). The slightly higher amount of GXM secreted into the supernatant of C. gattii cultures was not significant (p = 0.319, two-tailed t-test) (data not shown).
The amount of secreted extracellular vesicles correlates with capsule thickness, melanization and secreted laccase activity
Given that extracellular vesicles (EVs) are necessary for the export of capsular polysaccharides and laccase (20, 21), we also studied the EVs isolated from a subset of C. neoformans isolates. We used an indirect method to quantify them, measuring the concentration of ergosterol in cell-free supernatants (22) and dividing this by the number of cells (Figure 5A). Besides, we also measured the hydrodynamic diameters and polydispersity indices of the vesicle preparations by dynamic light scattering (DLS) (Figure 5B-C). We observed significant correlations between EV-ergosterol content and the visual melanization score and melanization EC50 (Figure 5D-E), but not with melanization Top and Span (Figure 5F-G). EV-ergosterol also showed significant correlation with whole-cell laccase activity and secreted laccase activity (Figure 5H-I). The indirect EV measurement in the supernatants of clinical isolates also correlated well with their basal capsule thickness in Sabouraud, but not with their ability to induce capsule in any of the three tested media (Figure 5J-N).

EVs preparations were measured by Dynamic Light Scattering (DLS), (A) Frequency distribution of the sterol quantification by sterol amplex red kit (Invitrogen). (B) Frequency distribution of the hydrodynamic diameter (intensity) and (C) Frequency distribution of the polydispersity index. (D) Correlation between EV-Ergosterol content (indicates amount of EV) and classical melanization score (r = 0.634, p = 0.004). (E) Correlation between EV-Ergosterol content and Melanization EC50 (represents the speed of melanization index from non-linear regression curve of median gray value) (r = −0.381, p = 0.030). (F) No correlation between EV-Ergosterol content with Melanization Top (maximum melanization index from non-linear regression curve of median gray value) (r = 0.271, p = 0.290) or (G) melanization Span (difference in median gray levels of the colony at the beginning and end of the experiment - difference between top and bottom) (r = 0.321, p = 0.004). (H) Correlation between EV-Ergosterol content with cell-wall laccase activity (r = 0.634, p <0.001) and (I) secreted laccase activity (r = 0.573, p = 0.013). (J) No correlation between EV-Ergosterol content and secreted GXM concentration (r = −0.022, p = 0.935). (K) Correlation between EV-Ergosterol content and capsule thickness in medium rich Sabouraud (Sab) (r = 0.490, p = 0.039). (L) No correlation between EV-Ergosterol content with capsule induction in medium Sab-MOPS (r = 0.228, p = 0.362), (M) capsule induction in medium minimal (r = 0.259, p = 0.298) and (N) capsule induction in CO2-independent medium (r = 0.067, p = 0.791). All samples (n = 18) were analyzed in duplicate and under the same conditions. All correlations were made with Spearman rank.
Melanization kinetics of clinical isolates affect the ability to escape from LC3-associated phagocytosis in macrophages
We correlated the LAP proportions described above with melanin and capsule phenotypes. The proportion of macrophages with LC3-positive phagosomes correlated strongly with melanization EC50 and inversely with melanization Top and Span (Figure 6A-C), but not with secreted laccase activity (Figure 6D). In contrast, no significant correlation between the proportion of macrophages with LC3-positive phagosomes and any of the capsule variables was found (Figure S3).

Using classical murine macrophages J774, without previous activation stimuli. (A) Correlation between LC3-associated phagocytosis with melanization EC50 (represents the speed of melanization index from non-linear regression curve of median gray value) (r = 0.686, p = 0.0002), (B) with melanization Top (maximum melanization index from non-linear regression curve of median gray value) (r = −0.483, p = 0.016), (C) with melanization Span (difference in median gray levels of the colony at the beginning and end of the experiment - difference between top and bottom) (r = −0.463, p = 0.022) and no correlation with secreted laccase activity (r = 0.194, p = 0.375). All correlations were made with Spearman rank.
Melanization kinetics, laccase activity and the capsule of clinical isolates affect survival in G. mellonella
We infected wax moth (G. mellonella) larvae with the clinical isolates to evaluate the role of melanization, laccase activity and capsule on virulence. This experiment was made with 15 C. gattii and 31 C. neoformans isolates divided into three lots. The survival curves for one of these lots is shown as an example in Figure 7A, whereas the distribution of median G. mellonella larvae survival times is shown in Figure 7B.

(A) Survival curve of G. mellonella infected with clinical isolates. In this experimental batch, all isolates were C. neoformans of the molecular type VNI. (B) Frequency distribution of the median survival G. mellonella infected with clinical isolates (n = 46). Twelve individuals were infected per isolate.
Using Cox proportional hazards regression, a multivariate survival tool, we generated a statistical model which evaluated the impact of virulence phenotypes upon survival time for all infected larvae (n = 588). In addition to virulence variables, we added as possible confounding variables the Cryptococcus species of the isolate and diet fed to G. mellonella larvae used for infections. The Cox regression model (X2 = 258.9, df = 10, p < 0.001) showed significant impacts on G. mellonella survival of capsule induction in Sab-MOPS (HR 1.7, 95% CI 1.2–2.4, P = 0.003), melanization EC50 (HR 0.8, 95% CI 0.7–0.9, P = 0.02), melanization Top (HR 2.9, 95% CI 1.6–5.2, P < 0.001), secreted laccase activity (HR 2.6, 95% CI 2.0–3.3, P < 0.001) and the confounding co-variable cereal diet (HR 7.5, 95% CI 5.0–11.2, P < 0.001). Variables that were not significant on this Cox regression model are capsule thickness in Sabouraud, capsule induction in MM and CIM, whole-cell laccase activity and species of Cryptococcus.
Secreted laccase activity and melanization EC50 increases the risk of death in patients with disseminated cryptococcosis
To study the impact of these phenotypes on the human disease, we used survival data from patients of the Brasília study for a Cox proportional hazards regression. Of the 41 patients, we included in the regression model only those with systemic disease caused by C. neoformans or mixed C. neoformans/C. gattii (n = 34). Eight patients were HIV negative and 23 HIV positive. The maximum follow-up time was 303 days. Patients who died during the follow-up time were accounted as events (n = 19), whereas the other ones were released after treatment (n = 14) or lost to follow up while still alive (n = 1). The Cox regression model (X2 = 13.7, df = 6, p = 0.032) showed significant impacts on patient survival of the melanization EC50 (HR 0.23, 95% CI 0.06–0.83, P = 0.025) and secreted laccase activity (HR 5.07, 95% CI 1.22–21.09, P = 0.025). Covariates that were not significant on the Cox regression model were age at the time of diagnosis, HIV status, whole-cell laccase activity and melanization Top.
Discussion
Capsule and melanin are two of the most important and studied virulence factors in the Cryptococcus genus, allowing the fungal cells to subvert host immunity and cause cryptococcosis. Most studies on these virulence factors are made either in vitro or in animal models, which are both highly informative but not completely applicable to human disease. We used a translational approach to study virulence phenotypes in clinical isolates, their interaction with experimental hosts and finally to associate them with the outcome of the disease in the patients from whom the isolates were obtained. We were thus able to obtain novel information on the role these two virulence attributes play in the pathogenesis of cryptococcal disease.
The most striking observations were related to melanin, a pigment that protects cryptococcal cells against oxidative stress, extreme temperatures and UV radiation (23–27). All isolates tested were able to produce the pigment, although some produced it faster than others and appeared darker at the end of the experiment. This melanization speed correlated negatively with the survival of cryptococcosis patients, suggesting a poor prognosis in infections caused by fast melanin forming strains. Sabiiti and colleagues (14) have previously shown that the amount of secreted laccase correlated well with cryptococcal survival in the cerebrospinal fluid and patient outcome, but when they analyzed the amount of melanin made, they were unable to establish a statistically significant correlation at the p = 0.05 level, which was possibly at Type II error. What they measured, however, was the total amount of melanin in the cells of a subset of ten clinical isolates, which might be similar to the Melanization Top variable we measured. Our observations suggest that the melanization speed, rather than the final amount of melanin, could be more important in determining the outcome of human cryptococcosis. This observation makes sense given that melanization protects cells against immune mechanisms and cells that melanize earlier would have a survival advantage.
Because laccase synthesizes melanin, melanization parameters are expected to depend to some extent on laccase production, which means that teasing their roles on outcomes of infection apart can be challenging. However, the present study suggests these roles are at least partly independent. Specifically, figure S2D shows that while laccase secretion generally correlates with melanization speed, outliers do exist that either secrete relatively large amounts of the enzyme, but melanize slowly, and melanize fast while secreting relatively low amounts of the enzyme. Differences in the efficiency of melanin anchoring in the cell wall, in the proportion of free and cell wall-associated laccase, or in the proportion of chitin in the cell wall (the polymer to which melanin is anchored (28)) may perturb the correlation between the two measurements. Statistically, residual plots of our correlation data are symmetric around zero for both variables, which also suggests the assumptions of our model are correct. Moreover, laccase has well-documented effects in infection outcomes that are melanin-independent, such as detoxification of iron (29), prostaglandin production (30) and neutralization of the fungicidal properties of cerebrospinal fluid (14).
Extracellular vesicles (EVs) are associated with several biological roles (31). In fungi, especially Cryptococcus, they are associated with the transport of various important virulence molecules like melanin, laccase, nucleic acids and others (20, 21). A C. neoformans mutant with impaired EV secretion, obtained through silencing of the SEC6 gene, presented a decrease in secreted laccase activity and was hypovirulent in mice (20). The formation of capsule and melanin are dependent on the secretion of EVs (32, 33). Hence, we quantified the association between EVs and the phenotypes of the capsule, laccase and melanin of a subset of the isolates, all of the same species of C. neoformans and molecular type VNI. The amount of EVs secreted in minimal medium had a strong correlation with laccase activity. Interestingly, we also found that the amount of EVs are associated with faster melanization and the larger capsule thickness in rich medium. On the other hand, we found no correlation with the total amount of melanin, the total amount of secreted GXM or the thickness of the capsule in nutrient-deficient inducing media. These findings highlight the role of EVs in the expression of C. neoformans virulence factors. Taken together, our data indicate that factors other than the amount of EVs are important for inducing capsule in nutrient-deficient media.
Melanization phenotypes and laccase activity are associated with greater virulence of clinical Cryptococcus spp. Isolates (8, 14, 34–38). Genetic and environmental factors contribute to the variation in the melanin production of C. neoformans (39, 40), which is under complex cellular regulation (40, 41). Furthermore, the experimental quantification of the pigment presents significant methodological challenges (42). C. neoformans showed better melanization capacity compared to C. gattii. But interestingly, C. gattii showed a greater capsule thickness in all media, except for minimal medium (which mimics the environment of cryptococcal meningitis infection, the main manifestation in most severe cases of cryptococcosis). We found that isolates with higher basal capsule thickness (Sabouraud) had a significant correlation with isolates with lower melanization speed (high melanization EC50), but this melanization index did not correlate with capsule thickness in MM. We found significant correlations between secreted laccase activity and relative capsule thickness in Sab-MOPS /Sabouraud medium after 24h of culture. Overall, these data suggest that melanization kinetics, secreted laccase activity and capsule thickness have different expression mechanisms between species. In addition, laccase activity differs not only between species but also at the cellular location where it is quantified. Perhaps higher melanin production and lower capsule thickness are important factors that favor the dissemination and survival of C. neoformans in the central nervous system, unlike C. gattii, which is strongly associated with pulmonary cryptococcosis (43).
The evolutionary divergence of proteins and signaling cascade configurations between C. neoformans and C. gattii may explain the differences in the expression of virulence factors. For example, TPS1 and TPS2 genes were found to be critical for thermotolerance, pathogenicity, capsule and melanin production in C. gattii (44), but the homologous genes in C. neoformans were required only for thermotolerance, not for capsule or melanin production (45). This may indicate that the expression of the capsule is influenced not only by the yeast environment, but also by different genetic traits between the two species (10). Here we hypothesize that C. gattii ability to infect immunocompetent individuals can be partly explained by the increased basal expression of important virulence factors such as the polysaccharide capsule.
Macrophages are crucial effector cells against fungi. However, some facultative intracellular pathogens, like Cryptococcus spp., can survive and replicate inside macrophages. These cells trigger autophagy as part of their response to intracellular pathogens (46, 47). The presence of LC3 (microtubule-associated protein 1 light chain 3 alpha) is associated with autophagosome maturation (48). The autophagy route called LC3 associated-phagocytosis (LAP) is important for the fungicidal activity of macrophages (16). Fungal virulence factors, such as melanin in Aspergillus fumigatus, inhibited LAP and increased virulence, in vitro and in a murine model (17, 18). In a subset of the clinical isolates, we observed a strong correlation between the melanization kinetics and the inhibition of LAP in murine macrophages.
Wax moth larvae (Lepidoptera) are an invertebrate animal model extensively used for in vivo studies of Cryptococcus virulence, host innate immunity after infection and the activity of antifungal compounds (49–51). Bouklas and collaborators showed that intracellular phagocytosis, killing by murine macrophages, capsule thickness and laccase activity did not correlate with C. neoformans virulence in G. mellonella. They found high-uptake strains to have significantly increased laccase activity, and virulence in mice, but not in G. mellonella (52). It should be added that methodological differences might explain the discrepancy in observations: Bouklas et al. measured laccase activity by melanin accumulation in a liquid culture over 16 hours at 37 °C, plus 24 hours at 25 °C, which corresponds roughly to the whole-cell laccase activity quantitation protocol we used, whereas we measured secreted laccase separately. In other words, there is no disagreement between our data and theirs, since in our work whole-cell laccase also had no detectable influence on the outcome of Galleria infection. Another study found that C. gattii strains exhibited similar virulence between murine inhalation models and G. mellonella infection (53). Following evidence of a deterministic system in G. mellonella cryptococcal infection (54), our results support the idea that virulence is an emerging property that cannot be easily predicted by a reductionistic approach, but can be partially resolved by the multivariate regression model. Furthermore, our result is in agreement with a previous report (51) that melanin synthesis was directly related with the level of virulence of four major molecular types of C. gattii in G. mellonella.
In summary, our study showed that melanization kinetics, secreted laccase activity and capsule growth in different inducing media are each associated with the virulence of clinical Cryptococcus strains in an invertebrate animal model. EVs, laccase secretion and melanin production represent a continuum that seems to exert the major influence on infection outcomes. A limitation of this study is that our patients were treated with different regimens in distinct health services. In our cohort of patients, faster colony pigmentation together with secreted laccase activity had a significant association with mortality attributed to disseminated cryptococcosis. The clinical isolates C. neoformans CNB004.1, CNB007.1 and CNB020 presented the lowest MelEC50 values of (faster melanization). Accordingly, they were found to be highly virulent in that the patients that harbored them all died within 25 days after diagnosis with cryptococcal meningoencephalitis. Altogether, we have seen strong correlations and trends, and although this does not necessarily imply causality, the strength and pattern of the associations indicate that melanization kinetics plays a key role in cryptococcal disease. These findings highlight the importance of the role of the laccase-dependent melanin pathway and its relevance to the human clinical outcomes, which suggests the EV-laccase-melanin nexus as an important source of targets for future therapeutic approaches to disseminated cryptococcosis.
Methods
Patients and Cryptococcus spp isolates
The isolates used in the study were obtained from two different sources in Brazil. One set came from the Culture Collection of Pathogenic Fungi at Fiocruz, in the city of Rio de Janeiro. The other one from an ongoing epidemiological study in the city Brasília. This study was approved by the Ethics Committee of the Foundation for Teaching and Research in Health Sciences (CEP-FEPECS). Patients or their legal guardians gave written informed consent for the collection of clinical data and specimens, including those from which fungal isolates were obtained.
A total of 82 clinical isolates were used. We added two control strains (H99 and B3501). The 28 isolates from Rio de Janeiro had been previously typed by MLST as C. neoformans molecular type VNI and C. gattii molecular type VGII, whereas the isolates from Brasília were typed by Sanger-sequencing an URA5 amplicon and comparing the sequence to that of standard strains. With this analysis, 65 isolates were determined to belong to the C. neoformans species complex, molecular type VNI, and 19 were typed as belonging to the C. gattii species complex, molecular type VGII. We were able to obtain clinical information from the medical records of 41 patients, which are shown in Table 1.
Cryptococcus culture
All yeasts were kept frozen in 35% glycerol on a −80°C freezer. From this stock, the isolates were streaked onto Sabouraud agar medium (2% dextrose, 1% peptone, 1.5% agar, pH 5.5) and grown for 72 hours at 30 °C. Isolated colonies were incubated for 72 hours in Sabouraud-dextrose (Sab) liquid medium (4% dextrose, 1% peptone, pH 5.5; Sigma-Aldrich) at 37°C with 250 rpm shaker rotation. Laboratory reference strains H99 (C. neoformans var. grubii, serotype A) and B3501 (C. neoformans var. neoformans, serotype D) were used as controls.
Melanin production evaluation
The minimal medium (MM) (15 mM d-Glucose, 10 mM MgSO4, 29.4 mM KH2PO4, 13 mM glycine, 3 μM thiamine-HCl) (26) was used in these experiments. The 2X concentrated MM and 3% agar were prepared separately. As thiamine and L-DOPA can be degraded at high temperatures, the agar was heated separately and, after reaching the temperature of 60°C, was mixed with the 2X MM, reaching a final concentration of 1.5% agar and 1X MM. After that, the medium was supplemented with 1 mM L-DOPA (Sigma-Aldrich). Then 1 mL of the MM agar was added to each well in a 12 well plate protected from light. In the center of each well, 5 μL of each isolate containing a total of 105 cells were inoculated. All inocula were made in duplicates, in two different wells. The plates were incubated at 37 °C protected from light and monitored for 7 days. After 24 hours of incubation, we photographed each plate every 12 hours. The photographs were taken inside a biosafety cabinet, using a mirror apparatus specially prepared to photograph culture plates. With these conditions, it was possible to standardize the illumination and photography parameters during the capture of all images. A Nikon D90 digital single-lens reflex (D-SLR) camera equipped with and 85 mm lens was used, with fixed focal length, exposure time, ISO setting, aperture, and white balance. The colony melanization phenotype was also evaluated and classified into heterogeneous (melanization mostly at the center or edges of the colony) or homogeneous colony melanization.
Semi-quantitative melanization score
Based on the images collected as described above, the isolates were qualitatively categorized into 7 groups ordered from 1 to 7. Group 1 contains the colony with less intense and slower melanization and group 7 the isolates with more intense and faster melanin production. Each isolate was scored independently by two researchers that were blinded to each other’s evaluation, and the outcomes were very similar.
Melanin quantification in grayscale
We also created a protocol based on previous publications (15, 55) to quantitatively analyze the images described above. All images were manipulated using Adobe Photoshop CS6 and ImageJ version 2.0.0-rc-65 / 1.52a. No nonlinear modifications were done on the original images. The photos were adjusted in Photoshop to show each plate horizontally and with a size of 2400 × 1800 pixels. Each image was then exported to ImageJ, converted to 8-bit gray scale and inverted. The “ROI” tool was used to measure the colonies to obtain the median, mean and area values. The median gray level values were the averaged for the two colonies from each isolate. These values were fit to a nonlinear regression equation [Agonist] vs. response - Variable slope (four parameters) on GraphPad Prism. Strain H99 was used as an internal control in each experiment. All values were normalized by the value obtained with H99 in the respective experiment.
Laccase activity
Each strain was inoculated into 5 mL YPD (2% glucose, 2% yeast extract) and incubated with 200 rpm agitation at 30°C for 24 hours. The cells were harvested by centrifugation at 1,000 × g for 10 minutes and the culture was resuspended and incubated at 30°C, for 5 days, in 5 mL of asparagine salts medium with glucose (0.3% glucose, 0.1% L-asparagine, 0.05% MgSO4, 1% of solution 1 M (pH6.5) Na2HPO4, 3 μM thiamine, 0.001% of solution 0.5 M CuSO4). The cells were harvested by centrifugation at 1,000 × g for 10 minutes and washed once with 5 mL 50 mM Na2HPO4 pH 7.0 and washed once with 3 mL asparagine salts medium without glucose. Yeast cells were counted in a hemocytometer and adjusted to achieve an inoculum 108 cells/mL. The same number of cells were resuspended in 5 mL of the medium without glucose and incubated at 30°C for 72 hours to induce laccase expression. After incubation, the number of yeast cells per sample were counted in a hemocytometer to normalize the secreted laccase activity in the supernatant by the number of cells present at the end of the culture. The supernatants were harvested at 4,000 g for 5 minutes, and secreted laccase activity was measured in a 96-well plate by adding 180 μL of supernatant from each isolate and 20 μL of 10 mM L-DOPA. To measure laccase activity in whole cells, we prepared suspensions of 108 cells/mL of each isolate. In 24-well plates, we placed 450 μL of each isolate suspension plus 50 μl 10 mM L-DOPA. In 96-well plates, we placed 180 μL of the yeast suspension and 20 μl 10 mM L-DOPA. The blank for laccase activity in living cells was 500 and 200 μL cell suspension, respectively. The amount of pigment produced was determined spectrophotometrically at a 480-nm wavelength read every hour for 6 to 48 hours. Absorbance values (OD) converted into laccase activity per yeast with the equation: laccase activity (μmol/number yeast) = mean of OD/(7.9 × number yeast at the end of culture). For L-DOPA we used a standard molar extinction coefficient of 7.9 μmol−1 (56, 57). Results were reported as the mean of two or three experiments. To adjust for interexperimental variation, laccase activity of the clinical isolates was expressed as a ratio to H99 (positive control).
ELISA for GXM quantitation secreted in supernatant
Inocula containing 105 cells/mL of each isolate were incubated in 3 mL of minimal medium for 72 hours at 30°C under 200 rpm shaking. The cells were precipitated by centrifugation at 4,000 × g for 10 minutes, 4°C and 1.5 mL of the supernatant was transferred to microcentrifuge tubes. Cells debris were precipitated by centrifugation at 15,000 × g for 30 minutes, 4°C and 1 mL of the supernatant was filtered through a 0.45 μm syringe filter (polycarbonate membrane) and kept at −20 ° C until the time of the ELISA assay. The supernatants were diluted 5,000 and 10,000-fold and analyzed for GXM by enzyme-linked immunosorbent assay (ELISA). A 96-well high binding polystyrene plate (Costar, #25801) was coated for 1 hour at 37°C with supernatants samples and standard curve of GXM from 0 to 10 μg/mL (H99 GXM standard) and then blocked with 1% bovine serum albumin for 1 hour at 37°C. The plates were then washed three times with a solution of Tris-buffered saline (0.05%Tween 20 in PBS), followed by the detection of GXM with 50 μL of the monoclonal IgG1 18B7 (1 μg/ml) for 1 hour at 37°C. The plates were washed three times again and binding of 18B7 was detected with 50 μL of alkaline phosphatase-conjugated GAM IgG1 (Fisher) (1 μg/mL) for 1 hour at 37°C. The plates were washed three more times and developed with 50 μL of p-nitrophenyl phosphate disodium hexahydrate (Pierce, Rockford) (1 mg/mL). The absorbance was measured at 405 nm after 15 minutes in an EON Microplate Spectrophotometer (Biotek Inc). After subtraction of the blank values, sample measurements were interpolated with a standard four-parameter sigmoidal curve. Values represent the averages of three independent culture and ELISA experiments, performed in different days. The result of each clinical isolate was normalized by the number of cells at the end of the culture time.
Capsule formation
Inocula were made from cultures grown overnight in Sabouraud medium. These cells were grown in 24-well culture plates with four distinct liquid media, with a starting density of 106 yeasts/mL. A non-capsule-inducing medium (Sabouraud dextrose - Sab) and three capsule-inducing media (58, 59) were used: minimal medium (MM), ten-fold diluted Sabouraud in 50 mM MOPS (SabMOPS) and CO2-independent medium (Thermo Fisher Scientific) (CIM) for 24 hours at 37°C. Afterwards, 10 μL of yeasts cells were stained using 1:1 India ink. The slides were photographed in a Zeiss Z1 Axio Observer inverted microscope using a 40X objective (EC Plan Neofluar 40X/0.75 Ph 2; Carl Zeiss GmbH) and an MRm cooled CCD camera (Carl Zeiss GmbH). Images were collected and the capsules measured with the ZEN 2012 software. Capsule thickness measurements were normalized by the value obtained with H99 in each experiment. The expressed results are a mean of three independent experiments performed on different days, with 20 cells measured per experiment.
Analysis of extracellular vesicles (EVs)
EVs were obtained from the culture supernatants of each clinical isolate as previously outlined (32), with modifications. Briefly, fungal cells were cultivated in 40 mL of minimal medium for 3 days at 30°C with shaking. The cultures were then sequentially centrifuged to remove smaller debris, filtered through a 0.8 μm filter and the supernatants ultra-centrifuged (Beckman Coulter optima I-90k, SW28 rotor). The precipitate was resuspended in 2 mL of the remaining culture medium. With 1 mL of EV preparations, the hydrodynamic diameter (intensity) and the polydispersity index were measured by Dynamic Light Scattering (DLS) (ZetaSizer Nano ZS90 (Malvern,UK). Vesicle quantification was performed based on the analysis of sterol in their membranes, using a quantitative fluorimetric Amplex Red sterol assay kit (Invitrogen, catalog number A12216), according to the manufacturer’s instructions. All samples were analyzed in duplicate and under the same conditions.
Interaction with macrophages from immunofluorescence microscopy and LC3-associated phagocytosis (LAP)
The J774.16 cell line (J774) was purchased from the American Type Culture Collection (ATCC) to study the interaction of clinical isolates with macrophages. Cells were maintained at 37°C in the presence of 5% CO2 in Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10% heat-inactivated fetal calf serum (FCS) and 1% penicillin–streptomycin (fresh medium) (all from Invitrogen). Cells were used between 10 and 35 passages. J774 cells (2 × 105 cells) were plated in a 13 mm round glass coverslip (previously treated with 5% HCL and heated to 90°C for 10 minutes) placed inside a flat-bottom 24-well tissue culture plate (Kasvi) and allowed to adhere for 24 hours. The cell monolayers were then infected with each IgG1-opsonized (mAb 18B7, 10μg/mL) clinical isolate in a proportion of two fungi per macrophage. The cells were co-incubated for 12 hours and then fixed and permeabilized with methanol at −20°C for 10 minutes. The cells were then incubated with rabbit polyclonal antibody to LC3 (Rabbit IgG anti-LC3-beta, Santa Cruz Biotechnology) followed by a fluorescein-conjugated secondary (Goat anti-Rabbit IgG conjugated Alexa Fluor 488, Invitrogen). After staining, the coverslips were sealed in ProLong Gold Antifade (Invitrogen). Images were collected on a Zeiss Z1 Axio Observer inverted microscope at 63X (Plan-Apochromatic 63X/1.4 NA, Carl Zeiss GmbH) and an MRm cooled CCD camera (Carl Zeiss GmbH) using the ZEN Blue 2 software. For each isolated, 100 macrophages with internalized C. neoformans cells were counted and scored as positive for LAP if at least one of the phagocytosed fungi was in an LC3-positive vacuole.
G. mellonella median survival time
G. mellonella larvae were reared in glass jars, at 30 °C in darkness. To maintain the colony, sufficient amounts of an artificial diet were added to the jars at least three times a week. Last instar larvae in the 200 mg weight range were injected in the terminal left proleg with 104 yeast cells in ten microliters of PBS containing ampicillin at 400 μg/mL. Twelve individuals were infected per group and the larvae were kept at 37°C after infection. Deaths were counted daily and the experiment was terminated when all individuals in the PBS group molted. Molted individuals in any group were censored from the analysis at the day of their molting. The median survival for each clinical isolate was normalized with that observed for H99 in each experiment.
Negative control (inoculated with PBS) and positive control (inoculated with H99) groups were repeated in each experimental set. Two of the three sets were made with G. mellonella larvae that had been fed a cereal-based diet, whereas the third set with 11 clinical isolates were fed beeswax and pollen.
Statistics
Values of p lower than 0.05 were considered significant. Spearman’s correlation was used to determine the correlation between in vitro phenotypes and in vivo patient outcomes. Differences between groups were determined using the two-tailed t-test for normally distributed data. Survival in G. mellonella infection studies was obtained by log-rank (Mantel-Cox) test using GraphPad Prism software. Results from G. mellonella survival studies and were evaluated with Cox proportional hazards regression using IBM SPSS software.
Study approval
Clinical cryptococcal isolates used in this study were obtained from patients enrolled in a clinical study in Brazil. All trials were approved by the Ethics Committee of Fiocruz, Brazil and by the Research Ethics Committee of - FEPECS/SES-DF, Brazil, number: 882,291 and 942,633. Patients or their legal guardians gave written informed consent, including authorization for the storage and use of their clinical isolates for future research. All patients were monitored and treated free of charge by the Brazilian national health system.
Author contributions
Conceived and designed the experiments: HRS, AMN, PA. Performed the experiments: HRS, SOF, GPOJ, CPR, KMG, EMG. Analyzed the data: HRS, AMN. Supplied funding, assisted in data analysis and oversaw writing of the paper: MSSF, ISP, PA, HCP and AC. Obtaining and tracking clinical isolates and data: WFF, AFC, JLJ, VLPJ, LT and MSL. Wrote the first manuscript draft: HRS and AMN. The manuscript draft was revised and approved by all authors.
Supplemental data

Reproducibility of the new method for quantifying melanin and colony melanization profile. (A) Colony image capture system (mirror plate reflector device developed and manufactured in our research group). (B) Five biological replicates of H99 performed in duplicate technique reveal high reproducibility of the methodology developed to quantify melanin. Three image processing programs were used (Adobe Photoshop CC version 19.0, ImageJ version 1.50i and Prism 7 version 7.0a, respectively) to calculate the median gray values of each colony over time. (C) Representative photo of the macroscopic melanization profile of three clinical isolates and the standard H99 internal control strain in all experiments. Two strains with homogeneous colony melanization (H99 and CNB017.1) and two heterogeneous (CNB013.1 and CGF007). (B and D) Non-linear regression curve of median gray value (represents amount of melanin measured at each observed time).

Validation and correlation of the new method for quantifying melanin. (A) Classic semi-quantitative melanization score. The isolates were qualitatively categorized into 7 groups ordered from 1 to 7. Group 1 contains the colony with smaller and later melanization and group 7 the most effective isolates for melanin production. (B) Correlation between Melanization Top (maximum melanization index from non-linear regression curve of median gray value) and classical melanization score (r = 0.501, p = 0.048). (C) Correlation between secreted laccase activity and classical melanization score (r = 0.74, p <0.001). (D) Correlation between secreted laccase activity and Melanization EC50 (represents the speed of melanization index from non-linear regression curve of median gray value) (r = 0.426, p <0.0004). Melanization score defined with double blind control of two experienced individuals. All correlations were made with Spearman rank.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Capsule thickness no affect the ability of clinical isolates to escape from LC3-associated phagocytosis. (A) No correlation between LC3-associated phagocytosis with capsule thickness in medium rich Sabouraud (Sab) (r = −0.013, p = 0.973), (B) with capsule induction in medium Sab-MOPS (r = −0.118, p = 0.716), (C) capsule induction in medium minimal (r = 0.314, p = 0.319) and (D) capsule induction in medium CO2 independent (r = −0.025, p = 0.869). All correlations were made with Spearman rank.
Acknowledgements
A.M.N was funded by FAP-DF awards 0193.001048/2015-0193.001561/2017 and the CNPq grant 437484/2018-1. M.S.S.F was supported by FAP-DF/PRONEX award 193.001.533/2016. AC was supported by the National Institutes of Health grants 5R01A1033774, 5R37AI033142, and 5T32A107506, and CTSA grants 1 ULI TR001073-01, 1 TLI 1 TR001072-01, and 1 KL2 TR001071 from the National Center for Advancing Translational Sciences. The authors would like to thank Jhones Dias and Alicia Ombredane for experimental assistance, as well as Emma Camacho and Radamés Cordero for the critical reading of the manuscript.
Footnotes
Conflict of interest statement: The authors have declared that no conflict of interest exists.
References




