

Abstract
Plant autotoxicity has proved to play an essential role in the behaviour of local vegetation. We analyse a reaction-diffusion-ODE model describing the interactions between vegetation, water, and autotoxicity. The presence of autotoxicity is seen to induce movement and deformation of spot patterns in some parameter regimes, a phenomenon which does not occur in classical biomass-water models. We aim to analytically quantify this novel feature, by studying travelling wave solutions in one spatial dimension. We use geometric singular perturbation theory to prove the existence of symmetric, stationary and non-symmetric, travelling pulse solutions, by constructing appropriate homoclinic orbits in the associated 5-dimensional dynamical system. In the singularly perturbed context, we perform an extensive scaling analysis of the dynamical system, identifying multiple asymptotic scaling regimes where (travelling) pulses may or may not be constructed. We discuss the agreement and discrepancy between the analytical results and numerical simulations. Our findings indicate how the inclusion of an additional ODE may significantly influence the properties of classical biomass-water models of Klausmeier/Gray–Scott type.
1 Introduction
Self-organized vegetation patterning has been proved in recent years to play an important role in our understanding of climate change and catastrophic shifts [44], in particular in arid and semi-arid environments (e.g., [9, 10]). Several reasons have been hypothesised to explain the occurrence of such patterns, focusing on spatial interactions between vegetation and the abiotic environment (e.g., [11, 20, 23, 27, 45, 49, 50]). The main mechanisms that have been identified in vegetation self-organization rely on the transport of water and nutrients toward vegetated patches through a scale-dependent (short-range positive, long-range negative) feedback, allowing plants to act as ecosystems engineers [16, 32]. Water availability alone, however, fails to explain the emergence of vegetation patterns in environments where water availability is not limited (see [46] and references therein).
A mechanism that is by now well established in the description of vegetation dynamics is plant-soil negative feedback (see, e.g., [15, 34, 42]). This feedback, which was already well-known in agriculture since ancient time, underlies the practice of “crop rotation” after repeated monoculture to avoid “soil sickness” - a situation where the same plant species cannot be grown within the same region after a certain amount of time. The origin of this feedback has been linked to several biological phenomena, such as the presence of soilborne pathogens, the changing composition of soil microbial communities [31], and the accumulation of autotoxic compounds from decomposing plant litter [4, 39]. In particular, it has been proved that plant-soil negative feedback occurs because of inhibitory effects of extracellular DNA [40]. Extensive studies have revealed that this negative feedback plays an important role in several aspects of vegetation dynamics, such as species coexistence and biodiversity [38], as well as and spatial organisation of plants by means of clonal rings [5, 6] and patterns [35].
In [35], in particular, the authors have found that the presence of autotoxicity induces – for some parameter ranges, and in agreement with experimental observations – the occurrence of dynamic patterns, with a non-symmetric distribution of biomass. The governing process is represented by the attempt of the biomass to “escape” areas with high levels of toxicity. The analytical foundation for such ecologically relevant results, however, is still lacking.
In this paper, we aim to take a first step in analytically justifying the numerical findings in [35] by using recently developed analytical techniques [14, 48]. In particular, we prove existence of both stationary and travelling pulses in one spatial dimension. To this aim, we use a convenient scaling, which allows us to transform the model introduced in [35] into an extension of the classical Klausmeier/Gray-Scott (KGS) model. For this classical KGS model, a large number of extensive analytical and numerical studies have been performed, investigating the existence, stability and dynamics of several types of patterns [12, 13, 28, 29, 41, 49, 50]; note that these references are but an incomplete selection of the vast literature on pattern formation in the KGS model.
We show that the extension of the KGS model with a third, spatially homogeneous, ordinary differential equation that models autotoxicity, can significantly influence the development and behaviour of pattern solutions, inducing the formation of travelling pulses which have been proven not to exist in the corresponding toxicity-free case [13].
From an analytical point of view, the inclusion of an additional model component significantly increases the complexity of the model, and hence introduces a potential obstacle for the successful analysis of pattern formation in this extended KGS model. One of the main results of this paper is that several asymptotic scaling regimes can be identified where the techniques developed in a two-component setting [14] nevertheless can be extended to analyse the existence of pulse patterns in this new three-component ‘reaction-diffusion-ODE’ model.
In the original KGS model, the existence and stability of pulse solutions sensitively depends on the asymptotic scaling of model components and parameters, see e.g. [2]. In order to analytically understand certain numerically observed behaviour, the ‘proper’ asymptotic scaling regime has to be identified. In the classical KGS model, this has already proven to be a subtle problem (see e.g. [48]); the inclusion of an additional model component exacerbates this subtility by increasing the number of scaling regimes that can be considered. Since choosing a particular asymptotic scaling is, to a certain extent, a prerequisite for the application of the analytical techniques presented in [14], the in-depth investigation of asymptotic scaling regimes relevant for investigating the existence of stationary and travelling pulse solutions, is a major part of this paper.
The paper is structured as follows: in Section 2, we present the original (dimensional) biomass-water-autotoxicity model [35], and the nondimensionalised, scaled model version we use in our subsequent analysis. Section 3 revolves around a collection of numerical results obtained by simulating both the original model in two spatial dimensions, and the rescaled, extended KGS model in a one-dimensional domain, showing the influence of autotoxicity. Section 4 is devoted to the existence for stationary and travelling pulses. Finally, a discussion of the presented results and an outline for future work in Section 5 conclude the paper.
2 The model
The original model introduced in [35] describes the dynamics of biomass (B), water (W), and autotoxicity (T) on a two-dimensional spatial domain, all measured in kg m−2, as follows:


 with time t ≥ 0 measured in days (d) and space
with time t ≥ 0 measured in days (d) and space  in meters (m);
in meters (m);  is the usual planar Laplace operator. System (2.1) is an extension of the classical Klausmeier model [27] on flat terrain, hence the absence of an advection term in (2.1a) (see e.g. [26]). An overview of the model parameters is given in Table 1.
is the usual planar Laplace operator. System (2.1) is an extension of the classical Klausmeier model [27] on flat terrain, hence the absence of an advection term in (2.1a) (see e.g. [26]). An overview of the model parameters is given in Table 1.
We rescale and nondimensionalise (2.1), obtaining the following system:


 which is defined on a two-dimensional spatial domain
which is defined on a two-dimensional spatial domain  for
for  . For the definition of the new model components and parameters in terms of the original components and parameters, see Appendix A.
. For the definition of the new model components and parameters in terms of the original components and parameters, see Appendix A.
The original KGS model can be recovered by setting  in (2.2), thereby decoupling the toxicity variable S from the water-biomass variable pair (U, V). For the decoupled system, there is extensive literature on the existence and dynamics of several types of patterns, both in one and two spatial dimensions (e.g. [2, 7, 12, 13, 28, 29, 41, 48, 49, 50, 54]). In particular, the existence and dynamics of pulse solutions in the original KGS model has been studied in great detail, using a variety of techniques [12, 13, 28, 29, 41, 49, 50]. In this paper, we adopt the analytical approach as outlined in [14]. Moreover, model (2.2) has been rescaled to enable parameter identification with the original KGS model as analysed in [13]. We want to emphasise that this does not imply our model formulation is in any way preferable over alternative formulations, such as those used in [28, 29] or [49, 50]; our choice merely reflects the similarity to the method employed in [13, 48], to enable easy comparison with previous results.
in (2.2), thereby decoupling the toxicity variable S from the water-biomass variable pair (U, V). For the decoupled system, there is extensive literature on the existence and dynamics of several types of patterns, both in one and two spatial dimensions (e.g. [2, 7, 12, 13, 28, 29, 41, 48, 49, 50, 54]). In particular, the existence and dynamics of pulse solutions in the original KGS model has been studied in great detail, using a variety of techniques [12, 13, 28, 29, 41, 49, 50]. In this paper, we adopt the analytical approach as outlined in [14]. Moreover, model (2.2) has been rescaled to enable parameter identification with the original KGS model as analysed in [13]. We want to emphasise that this does not imply our model formulation is in any way preferable over alternative formulations, such as those used in [28, 29] or [49, 50]; our choice merely reflects the similarity to the method employed in [13, 48], to enable easy comparison with previous results.
3 Numerics
In this section we illustrate the results obtained by performing numerical simulations of equations (2.1) and (2.2) on a two-dimensional and one-dimensional spatial domain, respectively. The main aim of this section is to highlight the differences caused by the presence of toxicity when compared to ‘classical’ biomass-water models without negative plant-soil feedback. When such negative plant-soil feedback is particularly strong (i.e. for low precipitation rate, high sensitivity to toxicity, and low toxicity decay rate), two effects arise, which are not detected in biomass-water models without toxicity: the emergence of spatio-temporal (dynamic) patterns and an asymmetric biomass distribution within the patterns.
3.1 Simulations on a two-dimensional domain
We consider} the dimensional biomass-water-toxicity model (2.1) on a two-dimensional domain Ω = {0 ≤ x ≤ Lx, 0 ≤ y ≤ Ly} with boundary and initial conditions
 and
and
 respectively. Here, ∂Ω is the boundary of Ω, ∂n is the normal derivative on ∂Ω, B0 and W0 are initial spatial distributions of biomass and water, respectively. Our simulations are based on a numerical scheme which uses finite differences in space, and forward Euler in time. The two-dimensional domain Ω is discretised via a square lattice of 100 100 elements, with Δx = Δy = 1 and Lx = Ly = 99 meters. The initial datum for the biomass B0 satisfies B0 = 0.2 in N0 = 5000 randomly selected elements (total initial biomass
respectively. Here, ∂Ω is the boundary of Ω, ∂n is the normal derivative on ∂Ω, B0 and W0 are initial spatial distributions of biomass and water, respectively. Our simulations are based on a numerical scheme which uses finite differences in space, and forward Euler in time. The two-dimensional domain Ω is discretised via a square lattice of 100 100 elements, with Δx = Δy = 1 and Lx = Ly = 99 meters. The initial datum for the biomass B0 satisfies B0 = 0.2 in N0 = 5000 randomly selected elements (total initial biomass  and B0 = 0 in the remaining nodes, while W0(x) = 40 uniformly. The simulation time tmax = 1000 days consists of 105 time-steps Δt, with Δt = 0.01 days.
and B0 = 0 in the remaining nodes, while W0(x) = 40 uniformly. The simulation time tmax = 1000 days consists of 105 time-steps Δt, with Δt = 0.01 days.
Since our goal is to investigate the difference between the case without toxicity (s = 0) and the one with strong influence of toxicity (s ≠ 0, small k), we fix all other parameter values in (2.1) as follows
 and consider two particularly representative cases for s and k (see Figure 1). In the case without toxicity (s = 0, Figure 1(a)), numerical simulations show the emergence of a stable stationary spatial pattern in the form of spots, consistent with existing literature results (see e.g. [26]). On the other hand, for s = 0.3 and k = 0.01, strong negative feedback due to plant toxicity induces the formation of dynamic patterns that continuously evolve in time with a propagation speed of about 3 meter/year. Moreover, the biomass distribution within the pattern is not symmetric, yielding crescent moon shapes (see Figure 1 (b)). These crescent moon shapes are observed to travel with constant velocity in the direction of highest local biomass concentration. Moreover, the velocity distribution is seen to be isotropic. Such non-symmetric patterns are usually linked to the presence of an advection term in the water equation, which models water transport due to slope (see e.g. [52, Figure 11]). Negative feedback due to autotoxicity is hence able to reproduce such non-symmetric shapes on flat terrain, in agreement with experimental observations (see [35]).
and consider two particularly representative cases for s and k (see Figure 1). In the case without toxicity (s = 0, Figure 1(a)), numerical simulations show the emergence of a stable stationary spatial pattern in the form of spots, consistent with existing literature results (see e.g. [26]). On the other hand, for s = 0.3 and k = 0.01, strong negative feedback due to plant toxicity induces the formation of dynamic patterns that continuously evolve in time with a propagation speed of about 3 meter/year. Moreover, the biomass distribution within the pattern is not symmetric, yielding crescent moon shapes (see Figure 1 (b)). These crescent moon shapes are observed to travel with constant velocity in the direction of highest local biomass concentration. Moreover, the velocity distribution is seen to be isotropic. Such non-symmetric patterns are usually linked to the presence of an advection term in the water equation, which models water transport due to slope (see e.g. [52, Figure 11]). Negative feedback due to autotoxicity is hence able to reproduce such non-symmetric shapes on flat terrain, in agreement with experimental observations (see [35]).

Biomass patterns obtained simulating equations (2.1) with parameter values (3.2). Darker shades of gray correspond to higher biomass density B(x, tmax). (a) s = 0, any k. In this case, the influence of toxicity is absent (as equations (2.1b) and (2.1c) are decoupled), and we recover the symmetric stationary spots present in ‘classical’ biomass-water models (see [26, 27]). (b) s = 0.3, k = 0.01. Toxicity induces both a non-symmetric distribution of biomass within the spots, and movement of the spots with constant velocity – that is, the formation of a dynamic spatio-temporal pattern.
3.2 Simulations on a one-dimensional domain
We consider equations the nondimensionalised, rescaled model (2.2) on a one-dimensional domain  with boundary and initial conditions
with boundary and initial conditions
 and
and

Our numerical simulations are again based on a numerical scheme which uses finite differences in space, and forward Euler in time. We consider  and discretize the domain by fixing m = 801 nodes at equal distance
and discretize the domain by fixing m = 801 nodes at equal distance  . The simulation time
. The simulation time  consists of 107 time steps
consists of 107 time steps  with
with  .
.
For  system (2.2) reduces to the classical KGS model. In one spatial dimension, the KGS model has been extensively studied, revealing the formation of self-replicating patterns (see e.g. [13, 28, 43]). In particular, in [13], the non-existence of travelling solitary pulses and the occurrence of pulse splitting leading to a stationary periodic pulse pattern have been shown, both analytically and numerically. In this section, our aim is to compare the numerical results presented in [13] for the toxicity-free scenario, with the scenario where (4.1b) and (4.1c) are coupled (i.e.,
system (2.2) reduces to the classical KGS model. In one spatial dimension, the KGS model has been extensively studied, revealing the formation of self-replicating patterns (see e.g. [13, 28, 43]). In particular, in [13], the non-existence of travelling solitary pulses and the occurrence of pulse splitting leading to a stationary periodic pulse pattern have been shown, both analytically and numerically. In this section, our aim is to compare the numerical results presented in [13] for the toxicity-free scenario, with the scenario where (4.1b) and (4.1c) are coupled (i.e.,  versus
versus  ). To this end, we fix all parameters except for
). To this end, we fix all parameters except for  as follows:
as follows:

These values are commensurate with the parameter choices in the two-dimensional simulations (3.2), with in addition k = 0.01, and s = 0 (no toxicity sensitivity) or s = 0.3 (high toxicity sensitivity), see also Appendix A. As initial data, we take  uniformly, which is commensurate with the choice W0(x) = 40 made in subsection 3.1. For V, we consider the following two initial profiles:
uniformly, which is commensurate with the choice W0(x) = 40 made in subsection 3.1. For V, we consider the following two initial profiles:
a Gaussian peak in the centre of the domain

a “half-bare–half-vegetated” domain


When  (see Figure 2), we observe the formation of a stationary, periodic, spotted pattern as a consequence of pulse splitting, independent of the choice of the initial V-profile. This observation is in agreement with the results presented in [13, 26] and with the two-dimensional numerical simulations presented in subsection 3.1. When we now ‘turn on’ the influence of toxicity by choosing
(see Figure 2), we observe the formation of a stationary, periodic, spotted pattern as a consequence of pulse splitting, independent of the choice of the initial V-profile. This observation is in agreement with the results presented in [13, 26] and with the two-dimensional numerical simulations presented in subsection 3.1. When we now ‘turn on’ the influence of toxicity by choosing  , the numerical simulations show the emergence of travelling pulses. In particular, when the initial profile V0(
, the numerical simulations show the emergence of travelling pulses. In particular, when the initial profile V0( ) is chosen as in (3.5), the initial pulse symmetrically splits into two pulses which periodically move away from each other and toward each other within the domain (see Figure 3(a)). On the other hand, if we take the “half-vegetated–half-bare” initial profile (3.6), we observe that a single pulse forms, which periodically travels back and forth within the domain (see Figure 3(b)). In both cases, numerical results reveal that the speed of such pulses is approximately 0.0384, which could be interpreted
) is chosen as in (3.5), the initial pulse symmetrically splits into two pulses which periodically move away from each other and toward each other within the domain (see Figure 3(a)). On the other hand, if we take the “half-vegetated–half-bare” initial profile (3.6), we observe that a single pulse forms, which periodically travels back and forth within the domain (see Figure 3(b)). In both cases, numerical results reveal that the speed of such pulses is approximately 0.0384, which could be interpreted  for this choice of ε. This value is of the same order of magnitude with respect to the numerical propagation speed computed for the 2D simulations.
for this choice of ε. This value is of the same order of magnitude with respect to the numerical propagation speed computed for the 2D simulations.

Evolution of V in space  and time
and time  obtained by numerically simulating equations (2.2) with parameters as in (3.4) and
obtained by numerically simulating equations (2.2) with parameters as in (3.4) and  . (a) Simulations obtained for an initial datum
. (a) Simulations obtained for an initial datum  as in (3.5). (b) Here,
as in (3.5). (b) Here,  is fixed as in (3.6). In both cases, decoupling the dynamics of V from those of S leads to a symmetric, stable, regular pulse pattern through a process of pulse splitting.
is fixed as in (3.6). In both cases, decoupling the dynamics of V from those of S leads to a symmetric, stable, regular pulse pattern through a process of pulse splitting.

Evolution of V in space  and time
and time  obtained by numerically simulating equations (2.2) with parameters as in (3.4) and
obtained by numerically simulating equations (2.2) with parameters as in (3.4) and  . (a) Simulations obtained for an initial datum
. (a) Simulations obtained for an initial datum  as in (3.5). The strong influence of S on the dynamics of V induces the formation of a symmetric pair of pulses, which travel within the domain, moving away from and towards each other periodically. (b) Here,
as in (3.5). The strong influence of S on the dynamics of V induces the formation of a symmetric pair of pulses, which travel within the domain, moving away from and towards each other periodically. (b) Here,  is chosen as in (3.6). In this case, a single pulse forms, which travels back and forth along the spatial domain. Note that, in both cases, V is not symmetrically distributed within the pulse.
is chosen as in (3.6). In this case, a single pulse forms, which travels back and forth along the spatial domain. Note that, in both cases, V is not symmetrically distributed within the pulse.
The results presented in this section appear to be independent of boundary conditions, as it persists when considering different (larger) domains which mimick an infinite spatial interval. A snapshot of the travelling pulse is shown in Figure 4.
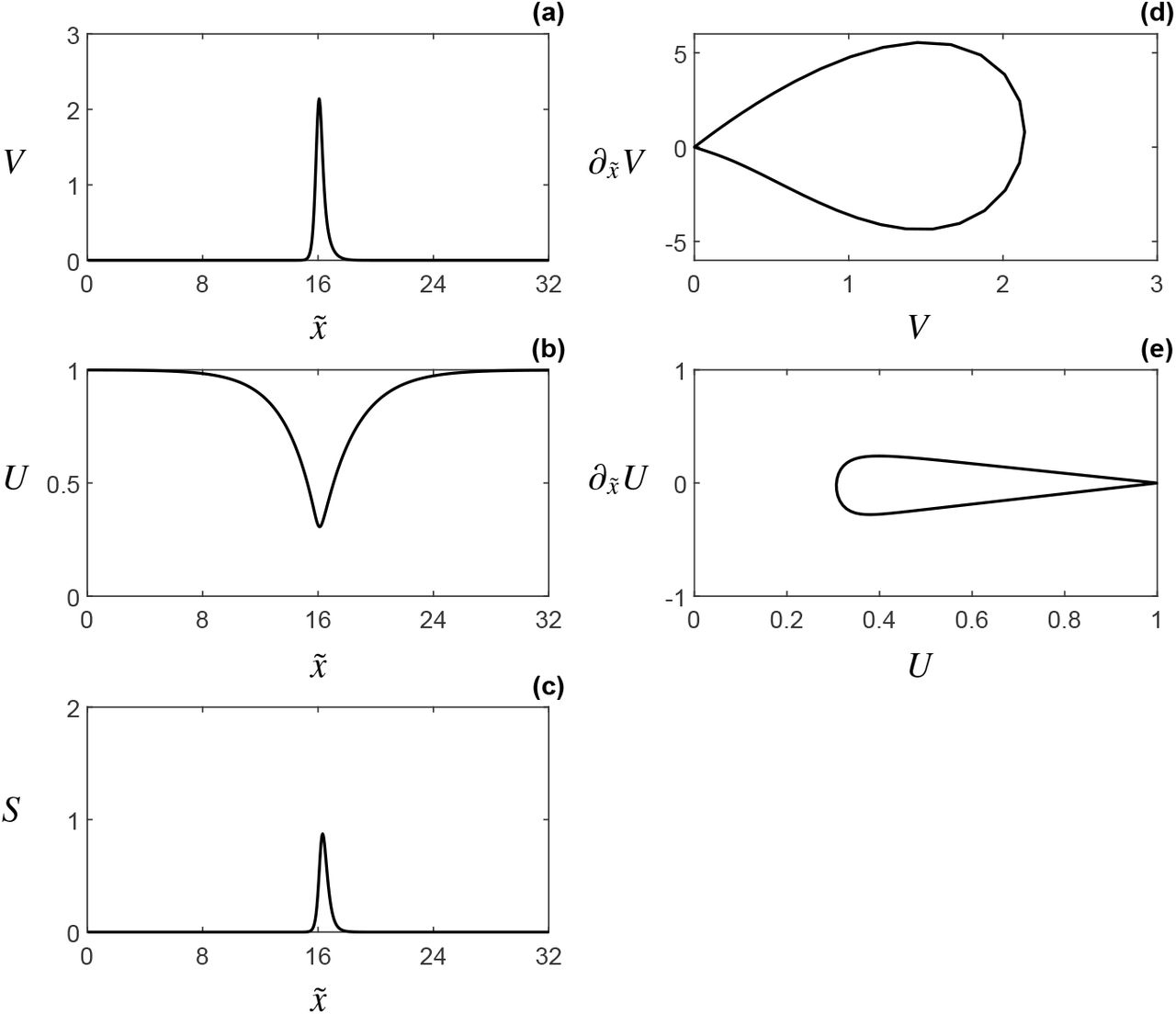




A preliminary, order-of-magnitude investigation into the role of the parameters  , and
, and  is presented in Table 2. An important observation is that the existence of pulse solutions not only clearly depends on the magnitude of
is presented in Table 2. An important observation is that the existence of pulse solutions not only clearly depends on the magnitude of  and
and  , but also on the fact that
, but also on the fact that  and
and  have to be chosen (approximately) equal. Secondly, by varying
have to be chosen (approximately) equal. Secondly, by varying  , we are able to find stationary pulse solutions. In addition, for relatively low values of
, we are able to find stationary pulse solutions. In addition, for relatively low values of  and
and  , simulations show strongly asymmetric, travelling pulse solutions –or travelling fronts with a decaying back– as a transient phase towards bare soil, see Figure 5. To our knowledge, transients of this shape have not been observed in the context of the classical KGS model.
, simulations show strongly asymmetric, travelling pulse solutions –or travelling fronts with a decaying back– as a transient phase towards bare soil, see Figure 5. To our knowledge, transients of this shape have not been observed in the context of the classical KGS model.
An overview of numerical simulation results of system (2.2) for several choices of parameters  and
and  . Throughout,
. Throughout,  and ε = 0.1. (*) In the case
and ε = 0.1. (*) In the case  , the pulses seem to be slowing down over the simulated time interval, suggesting a possible transience to stationary pulses.
, the pulses seem to be slowing down over the simulated time interval, suggesting a possible transience to stationary pulses.

Evolution of V in space  and time
and time  obtained by numerically simulating equations (2.2) with and
obtained by numerically simulating equations (2.2) with and  ε = 0.1. (a) Initial datum
ε = 0.1. (a) Initial datum  as in (3.5). Here, two travelling fronts form, meet at the centre of the domain, and then merge and decay to a bare soil state. (b)
as in (3.5). Here, two travelling fronts form, meet at the centre of the domain, and then merge and decay to a bare soil state. (b)  as in (3.6). In this case, a single travelling front emerges, and eventually decays to the bare soil state as well.
as in (3.6). In this case, a single travelling front emerges, and eventually decays to the bare soil state as well.
4 Existence of pulse solutions
In this section, we investigate the existence of stationary and travelling pulse solutions to system (2.2) on an unbounded, one-dimensional spatial domain  . System (2.2) in one spatial dimension is restated here for completeness:
. System (2.2) in one spatial dimension is restated here for completeness:


 We introduce a co-moving frame coordinate
We introduce a co-moving frame coordinate
 in which system (4.1) takes the form
in which system (4.1) takes the form



We look for pulse solutions, i.e. solutions to (4.3) that are bi-asymptotic to the trivial background state (U, V, S) = (1, 0, 0), that is, solutions (U(z), V(z), S (z)) for which limz→±∞(U(z), V(z), S (z)) = (1, 0, 0). To that end, we make the crucial assumption that ε, which measures the biomass diffusion rate with respect to the water diffusion rate, is an asymptotically small parameter, i.e. 0 < ε ≪ 1. This is consistent with the experimental data collected in arid environments (see, e.g., [45] and references therein). Next, in accordance with previous work [12, 13, 48, 54], we allow all model variables and parameters to scale with (a power of) this small parameter ε. While the introduction of these scalings can seem unnecessary complicated and confusion due to the large number of new parameters that are introduced as scaling exponents, previous analysis [12, 13, 48, 54] has shown that several solution types –and pulse patterns in particular– exist only in regions of parameter space that scale with ε in a particular way; moreover, the resulting pattern amplitude may also scale with ε. A preparatory asymptotic scaling analysis of system (4.3), along the lines of [13, 48], can be found in Appendix B. The resulting rescaling can be summarised as follows:
 with the additional assumptions that
with the additional assumptions that

Note that the positivity of the original model parameters  implies that their rescaled counterparts a, b, d, h are positive and
implies that their rescaled counterparts a, b, d, h are positive and  in ε; however, the associated scaling exponents may be negative. This also ap-plies to the model variables U, V, S and their rescaled counterparts u, v, s. The speed of the moving frame,
in ε; however, the associated scaling exponents may be negative. This also ap-plies to the model variables U, V, S and their rescaled counterparts u, v, s. The speed of the moving frame,  can take any sign.
can take any sign.
Application of rescaling (4.4) to (4.3) yields the following 5-dimensional dynamical system:





Pulse solutions to (4.3) can now be identified with orbits in system (4.6) that are homoclinic to the (rescaled) trivial background state (ε−κ, 0, 0, 0, 0). In the following, we will use techniques from geometric singular perturbation theory [12, 13, 14, 48] to constructively establish the existence of such homoclinic orbits in system (4.6).
4.1 Stationary pulses
For stationary pulses, we have  in (4.2), hence c = 0 in (4.6). We obtain the differential-algebraic system
in (4.2), hence c = 0 in (4.6). We obtain the differential-algebraic system




 where we have defined the two small parameters
where we have defined the two small parameters

Note that ε1,2 are indeed asymptotically small by the scaling conditions (4.5).
First, we observe that the algebraic equation (4.7e) can be solved for s, yielding
 which (as s is positive) implies that
which (as s is positive) implies that

In other words, the dynamics of (4.7) take place on the 4-dimensional invariant manifold  embedded in 5-dimensional (u, p, v, q, s)-phase space, that is given by
embedded in 5-dimensional (u, p, v, q, s)-phase space, that is given by
 see also Figure 8 (a). At this point, the introduction of
see also Figure 8 (a). At this point, the introduction of  can seem somewhat superfluous; however, this manifold will also play a role in the upcoming analysis of travelling pulses (see Section 4.2), and its introduction can illuminate similarities between the analysis of the current section and that of section 4.2.
can seem somewhat superfluous; however, this manifold will also play a role in the upcoming analysis of travelling pulses (see Section 4.2), and its introduction can illuminate similarities between the analysis of the current section and that of section 4.2.
On  the dynamics are given by the 4-dimensional dynamical system
the dynamics are given by the 4-dimensional dynamical system




Note that, for h = 0, this is exactly the system studied in [12, 13]. For future reference, we introduce the 2-dimensional hyperplane

We note that  is invariant under the flow of (4.7); moreover,
is invariant under the flow of (4.7); moreover,  (4.11).
(4.11).
4.1.1 Fast dynamics
Following the approach of geometric singular perturbation theory, we study the fast reduced limit of (4.12) by letting ε1 → 0, and obtain the planar, Hamiltonian, fast reduced system



The dynamics of p can be obtained by direct integration of pz = u0v2. Note that  (4.13) consists of hyperbolic (trivial) equilibria of the fast reduced system (4.14). We introduce
(4.13) consists of hyperbolic (trivial) equilibria of the fast reduced system (4.14). We introduce
 to obtain
to obtain
 with Hamiltonian
with Hamiltonian

Analogously to [12, 13, 14, 48], we look for a homoclinic orbit to the origin in system (4.16); note that this orbit lies on the level set Hf = 0. Straightforward phase plane analysis reveals that such a homoclinic orbit exists as long as 0 < δ < δmax, and the maximally attained w-value of the associated ‘spike’ solution is given by the unique positive solution to

The value for δmax can now be determined by considering the situation when (4.18) is degenerate, which is when δ = w(1 − w). Hence, we find that
 where wmax is the unique positive solution to
where wmax is the unique positive solution to
 the approximate numerical values are
the approximate numerical values are

See Figure 6 for an illustration of the dynamics of (4.16).
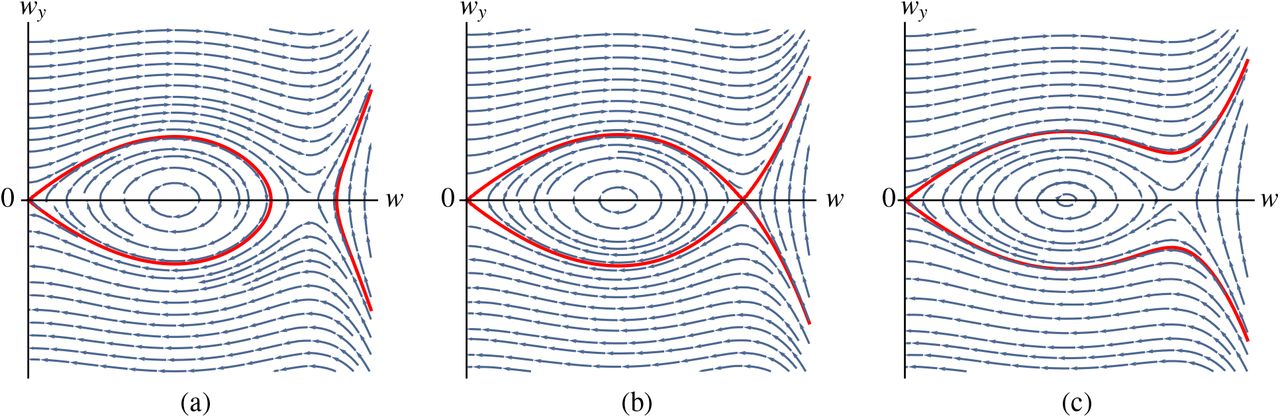
The phase plane of (4.16) for varying values of δ. The level set Hf = 0 is indicated in red. (a) 0 < δ < δmax, a planar homoclinic orbit exists. (b) δ = δmax, the planar homoclinic orbit deteriorates in a pair of heteroclinic orbits. (c) δ > δmax, no planar homoclinic orbit to the origin exists.
Note that, although the unique homoclinic solution wh(y; δ) to (4.16) does not have a closed-form expression, it can be approximated reasonably accurately by substitution of an asymptotic expansion in δ, as 0 < δ < δmax ≈ 0.23 (4.21). Writing
 we find for the first terms
we find for the first terms

 see Figure 7 for a visualisation of the approximation accuracy of series (4.22).
see Figure 7 for a visualisation of the approximation accuracy of series (4.22).




The geometric construction of the stationary pulse solution underlying Theorem 4.1. (a) The 4-dimensional invariant manifold  (4.11) embedded in 5-dimensional (u, p, v, q, s)-phase space. (b) On
(4.11) embedded in 5-dimensional (u, p, v, q, s)-phase space. (b) On  , the slow (u, p)-dynamics take place on the invariant manifold
, the slow (u, p)-dynamics take place on the invariant manifold  (4.13). The fast dynamics normal to
(4.13). The fast dynamics normal to  , can be used to connect the unstable and stable subspaces Eu and Es (4.25) through the fast homoclinic orbit wh (see subsection 4.1.1); see also [13, Figures 2 and 3].
, can be used to connect the unstable and stable subspaces Eu and Es (4.25) through the fast homoclinic orbit wh (see subsection 4.1.1); see also [13, Figures 2 and 3].
4.1.2 Slow dynamics
As noted at its definition,  (4.13) is a normally hyperbolic manifold that is invariant under the flow of (4.7). The flow on
(4.13) is a normally hyperbolic manifold that is invariant under the flow of (4.7). The flow on  is given by
is given by


This flow is linear; it has a unique saddle equilibrium at  , with stable resp. unstable manifolds given by the stable resp. unstable linear subspaces
, with stable resp. unstable manifolds given by the stable resp. unstable linear subspaces


Note that the dynamics of (4.24) are slow in z; the eigenvalues of the saddle equilibrium are of order 
4.1.3 Constructing a stationary pulse solution
We establish the existence of a stationary pulse solution to (4.1) by constructing a homoclinic orbit in system (4.7). This construction, which uses techniques and concepts from geometric singular perturbation theory, is based directly on the equivalent construction of pulses in the classical KGS model, as carried out in e.g. [13]. Due to the abundance of high quality sources, we choose to highlight central concepts in the pulse construction below. For detailed arguments and proofs, we refer to [12, 13, 14, 48].
The pulse solution that we want to construct, is an orbit that is homoclinic to the equilibrium ( , 0, 0, 0, 0) in (4.7). This equilibrium lies in particular on the invariant manifold
, 0, 0, 0, 0) in (4.7). This equilibrium lies in particular on the invariant manifold  (4.13). In order to construct a homoclinic orbit, we need to consider the flow both on
(4.13). In order to construct a homoclinic orbit, we need to consider the flow both on  and normal to
and normal to  . The latter can be studied from a geometric viewpoint by considering the unstable and stable submanifolds of
. The latter can be studied from a geometric viewpoint by considering the unstable and stable submanifolds of  under the flow (4.7), denoted by
under the flow (4.7), denoted by  respectively. To leading order in ε (and hence to leading order in ε1,2), the flow normal to
respectively. To leading order in ε (and hence to leading order in ε1,2), the flow normal to  is given by the fast reduced system (4.14), as studied in subsection 4.1.1. The existence of a homoclinic orbit in this planar system implies that
is given by the fast reduced system (4.14), as studied in subsection 4.1.1. The existence of a homoclinic orbit in this planar system implies that  and
and  intersect. Moreover, using the reversibility symmetry of (4.7) (i.e. the invariance of the flow (4.7) of under the reflection (z; p, q)→(−z, −p, −q)), one can show that this intersection is transversal; for details, see [13, 48]. Using the homoclinic solution wh(y; δ) to (4.16), we can track the fast flow normal to
intersect. Moreover, using the reversibility symmetry of (4.7) (i.e. the invariance of the flow (4.7) of under the reflection (z; p, q)→(−z, −p, −q)), one can show that this intersection is transversal; for details, see [13, 48]. Using the homoclinic solution wh(y; δ) to (4.16), we can track the fast flow normal to  through this intersection. We define the interval
through this intersection. We define the interval
 chosen such that I f is asymptotically large in z but asymptotically small in ε1z. We calculate the change of the slow variables u and p through the fast flow, over this interval, as follows. From (4.7), we see that uz is
chosen such that I f is asymptotically large in z but asymptotically small in ε1z. We calculate the change of the slow variables u and p through the fast flow, over this interval, as follows. From (4.7), we see that uz is  hence u is constant to leading order in z; we write
hence u is constant to leading order in z; we write

Moreover, pz is to leading order slaved to u and v; we write

We observe that, to leading order, p − p0 is odd in z. The change of u and p over I f can now be calculated to leading order as

 cf. (4.15). Geometric singular perturbation theory [13, 14, 22, 30] enables us to construct an orbit by concatenating orbits on
cf. (4.15). Geometric singular perturbation theory [13, 14, 22, 30] enables us to construct an orbit by concatenating orbits on  and normal to
and normal to  ; in particular, any orbit homoclinic to
; in particular, any orbit homoclinic to  first flows away from this equilibrium along (and exponentially close to) Eu (4.25). Then, it takes an excursion away from
first flows away from this equilibrium along (and exponentially close to) Eu (4.25). Then, it takes an excursion away from  , during which its evolution is to leading order determined by the fast reduced flow (4.14). After touching down again exponentially close to
, during which its evolution is to leading order determined by the fast reduced flow (4.14). After touching down again exponentially close to  , the u-component has not changed its value to leading order, but the p-component has, cf. (4.29). For the orbit to be biasymptotic to the equilibrium (
, the u-component has not changed its value to leading order, but the p-component has, cf. (4.29). For the orbit to be biasymptotic to the equilibrium ( , 0, 0, 0, 0), this change needs to be such that the touchdown point lies exponentially close to Es (4.25), such that the last, slow, orbit component takes us back to the (
, 0, 0, 0, 0), this change needs to be such that the touchdown point lies exponentially close to Es (4.25), such that the last, slow, orbit component takes us back to the ( , 0, 0, 0, 0) along Es; see Figure 8 for a sketch of the geometric situation and construction.
, 0, 0, 0, 0) along Es; see Figure 8 for a sketch of the geometric situation and construction.
For this concatenation construction to work, we find the condition that the ‘ vertical’ p-distance between Eu and Es must be equal to the leading order change in p during the fast excursion over the interval I f, as calculated in (4.29), that is,

Using techniques from geometric singular perturbation theory, one can show that, for sufficiently small ε1, condition (4.30) is not only necessary, but also sufficient to prove the existences of a homoclinic orbit in (4.7) that is asymptotically close to its singular concatenation. This existence follows from the persistence of 0 and its stable and unstable manifolds  , together with the observation that
, together with the observation that  and
and  intersect transversally; for more details and proofs, see [13, 14, 48]. For the purposes of this paper, it is sufficient to consider the outcome of previous equivalent analyses, namely that stationary pulse solutions are completely characterised by the existence condition (4.30), in the following way.
intersect transversally; for more details and proofs, see [13, 14, 48]. For the purposes of this paper, it is sufficient to consider the outcome of previous equivalent analyses, namely that stationary pulse solutions are completely characterised by the existence condition (4.30), in the following way.
Let ε1 be sufficiently small, and let u∗ be a nondegenerate solution to
 where wh(y; bh/u∗) is the unique, positive, nontrivial solution to (4.16) for which limy±∞ wh(y; bh/u∗) = 0. Then, system (4.7) admits an orbit that is homoclinic to the equilibrium (
where wh(y; bh/u∗) is the unique, positive, nontrivial solution to (4.16) for which limy±∞ wh(y; bh/u∗) = 0. Then, system (4.7) admits an orbit that is homoclinic to the equilibrium ( , 0, 0, 0, 0). To leading order in ε1, this orbit is given by
, 0, 0, 0, 0). To leading order in ε1, this orbit is given by



Conversely, if (4.31) has no solution, then no orbit homoclinic to ( , 0, 0, 0, 0) exists.
, 0, 0, 0, 0) exists.
Proof. The proof is completely analogous to that of [13, Theorem 4.1], with the fast planar homoclinic wh(y; δ) taking the role of v0 [13, equation (3.3)] therein. Since the arguments of the proof of [13, Theorem 4.1] depend only on the existence of such a planar homoclinic orbit, and not on its specific functional form, the reasoning in [13] holds ad verbatim for the construction described in section 4.1 which leads to the statement of Theorem 4.1. □
For the construction of pulse solutions in general systems of singularly perturbed reaction-diffusion equations, see [14].
Note that condition (4.30) can be formulated in terms of δ as
 taking the limit h → 0, i.e. δ → 0 (4.16), yields
taking the limit h → 0, i.e. δ → 0 (4.16), yields
 cf. (4.22) and (4.23). This is in accordance with the observation that the limit h → 0 of system (4.7) yields the classical KGS system, for which (4.34) has been derived as the existence condition of stationary pulse solutions [13, equation (4.2)]. See Figure 9 for the behaviour of u∗ for increasing values of h, and for a plot of the pulse solution given in Theorem 4.1.
cf. (4.22) and (4.23). This is in accordance with the observation that the limit h → 0 of system (4.7) yields the classical KGS system, for which (4.34) has been derived as the existence condition of stationary pulse solutions [13, equation (4.2)]. See Figure 9 for the behaviour of u∗ for increasing values of h, and for a plot of the pulse solution given in Theorem 4.1.

(a) The value of u∗, solving the existence condition (4.31) for increasing values of h (with a and b fixed), in blue. The set of admissible values, for which  is bounded below by the dashed line
is bounded below by the dashed line  . (b) A plot of the pulse solution established in Theorem 4.1, with uh in blue, vh in orange and sh in green.
. (b) A plot of the pulse solution established in Theorem 4.1, with uh in blue, vh in orange and sh in green.
4.2 Travelling pulses
Pulses travelling with nonzero speed  i.e c ≠ 0, are homoclinic orbits to system (4.6). In contrast to the stationary case studied in section 4.1, the dynamics of this system are fully 5-dimensional. For clarity of presentation, we introduce the small parameter
i.e c ≠ 0, are homoclinic orbits to system (4.6). In contrast to the stationary case studied in section 4.1, the dynamics of this system are fully 5-dimensional. For clarity of presentation, we introduce the small parameter
 note that ε3 is indeed asymptotically small by conditions (4.5). Using the previously defined small parameters ε1,2 (4.8), we can rewrite system (4.6) as
note that ε3 is indeed asymptotically small by conditions (4.5). Using the previously defined small parameters ε1,2 (4.8), we can rewrite system (4.6) as





This system can be treated in a similar fashion as in section 4.1. However, there are important differences. First, we observe that, unlike in the stationary situation (see section 4.1.1), the time scale of the dynamics of (v, q, s) in system (4.36) is not immediately clear, because the asymptotic magnitude of ε δ+β is not yet determined. In particular, depending on the sign and magnitude of δ + β, the dynamics of s may be faster, equivalent, or slower than those of (v, q). We examine all three cases, and the subsequent construction of a travelling pulse, in subsections 4.2.2, 4.2.3 and 4.2.4. Secondly, it is important to note that that the invariant 2-dimensional hyperplane  (4.13) that was defined in the context of stationary pulses, is also invariant under the flow of the ‘nonzero speed system’ (4.36). The dynamics of (4.36) on
(4.13) that was defined in the context of stationary pulses, is also invariant under the flow of the ‘nonzero speed system’ (4.36). The dynamics of (4.36) on  will be analysed in subsection 4.2.1.
will be analysed in subsection 4.2.1.
4.2.1 Slow dynamics
On the invariant hyperplane  (4.13), the dynamics of (4.36) are determined by the planar system
(4.13), the dynamics of (4.36) are determined by the planar system


As in the stationary case (see subsection 4.1.2), the dynamics on  are linear. Again, the only equilibrium is the saddle at
are linear. Again, the only equilibrium is the saddle at  . However, due to the advection term, the flow is not symmetric anymore with respect to reflection in the u-axis (compare (4.24) and (4.25)); instead, the stable resp. unstable manifolds of the saddle equilibrium are given by the stable resp. unstable linear subspaces
. However, due to the advection term, the flow is not symmetric anymore with respect to reflection in the u-axis (compare (4.24) and (4.25)); instead, the stable resp. unstable manifolds of the saddle equilibrium are given by the stable resp. unstable linear subspaces


The dynamics of (4.37) are slow in z, as the eigenvalues of the saddle equilibrium are of order  – that is, asymptotically small.
– that is, asymptotically small.
4.2.2 Constructing a travelling pulse solution, case I: s faster than(v, q)
We assume that the s-dynamics are faster than the (v, q)-dynamics, that is, ε3ε δ+β → 0 as ε → 0. Taking the limit ε → 0 in (4.36) then yields





This is precisely the same system as studied in subsection 4.1.1. In particular, the same algebraic equation as (4.7e) defines the same 4-dimensional manifold  (4.11). Introducing the fast coordinate
(4.11). Introducing the fast coordinate
 system (4.36) takes the form
system (4.36) takes the form




 with singular limit
with singular limit


It is easy to check using (4.42) that  which consists entirely of equilibria of the reduced fast system (4.42), is normally hyperbolic. Hence, by geometric singular perturbation theory (see e.g. [22, 30]), for sufficiently small ε, there exist a normally hyperbolic manifold
which consists entirely of equilibria of the reduced fast system (4.42), is normally hyperbolic. Hence, by geometric singular perturbation theory (see e.g. [22, 30]), for sufficiently small ε, there exist a normally hyperbolic manifold  that is invariant under the flow of the full 5-dimensional system (4.36); moreover,
that is invariant under the flow of the full 5-dimensional system (4.36); moreover,  is
is  close to
close to 
In contrast to the stationary case (4.7), the dynamical system we are investigating here (4.41) is fully 5-dimensional; hence, we need to look at the dynamics normal to  . Since these normal dynamics are one-dimensional,
. Since these normal dynamics are one-dimensional,  is either uniformly attracting or uniformly repelling, depending on the sign of c. In either case, it follows that any bounded (in particular, any homoclinic) orbit of (4.36) must lie entirely on
is either uniformly attracting or uniformly repelling, depending on the sign of c. In either case, it follows that any bounded (in particular, any homoclinic) orbit of (4.36) must lie entirely on  . By the uniform asymptotic proximity of
. By the uniform asymptotic proximity of  to
to  we can determine the dynamics on
we can determine the dynamics on  by a regular perturbation expansion of system (4.36) in powers of ε3ε δ+β.
by a regular perturbation expansion of system (4.36) in powers of ε3ε δ+β.
We also observe that, as  is invariant under the full flow of (4.36), it necessarily holds that
is invariant under the full flow of (4.36), it necessarily holds that  . To construct a travelling pulse solution on
. To construct a travelling pulse solution on  , we study the unstable and stable manifolds of
, we study the unstable and stable manifolds of  in their intersection with
in their intersection with  ; depending on the sign of c, this effectively means we disregard the fast expansion (c > 0) or contraction (c < 0) in the (normal) s-direction. Defining the 3-dimensional manifolds
; depending on the sign of c, this effectively means we disregard the fast expansion (c > 0) or contraction (c < 0) in the (normal) s-direction. Defining the 3-dimensional manifolds
 we want to determine whether
we want to determine whether  and
and  intersect transversally, in order to repeat the construction procedure outlined in subsection 4.1, see also [12, 13, 14, 48]. From the existence of a planar homoclinic orbit in the fast singular limit (4.39) (see subsection 4.1.1), it follows that the singular limits of
intersect transversally, in order to repeat the construction procedure outlined in subsection 4.1, see also [12, 13, 14, 48]. From the existence of a planar homoclinic orbit in the fast singular limit (4.39) (see subsection 4.1.1), it follows that the singular limits of  and
and  coincide, as in the stationary case. However, due to the fact that the full system (4.36) does not exhibit reversibility symmetry due to the presence of the c-dependent terms, the question whether
coincide, as in the stationary case. However, due to the fact that the full system (4.36) does not exhibit reversibility symmetry due to the presence of the c-dependent terms, the question whether  and
and  intersect transversally for nonzero ε cannot be answered based on symmetry arguments alone, as was the case in section 4.1.3, see also [13, 48]. Here, we need to perform a Melnikov calculation, along the lines of the analysis in [13, 48]. The Melnikov calculation is based on the observation that the singular limit system (4.39), in particular its planar reduction (4.14), is Hamiltonian. The associated conserved quantity Hf (4.17) is zero along the singular planar homoclinic wh(y); moreover, we see that Hf identically vanishes on
intersect transversally for nonzero ε cannot be answered based on symmetry arguments alone, as was the case in section 4.1.3, see also [13, 48]. Here, we need to perform a Melnikov calculation, along the lines of the analysis in [13, 48]. The Melnikov calculation is based on the observation that the singular limit system (4.39), in particular its planar reduction (4.14), is Hamiltonian. The associated conserved quantity Hf (4.17) is zero along the singular planar homoclinic wh(y); moreover, we see that Hf identically vanishes on  . Hence, along any orbit in
. Hence, along any orbit in  (which is necessarily biasymptotic to
(which is necessarily biasymptotic to  ), the total change of Hf must vanish.
), the total change of Hf must vanish.
In terms of the original system variables (u, p, v, q, s), Hf takes the form

Hence, the change of Hf in z is given by
 by (4.36). Using (4.36e), we can write
by (4.36). Using (4.36e), we can write
 so that we obtain
so that we obtain

Following the analysis in [14, 48], we use the previously defined interval I f (4.26) to express the total change of Hf along an orbit  in as
in as

Now, we can use previous analysis on the reduced system (4.41) to obtain leading order expressions for (v, q, s), which can be used to obtain a leading order expression for  . Defining the positive integrals
. Defining the positive integrals


 we can use (4.9), (4.15), (4.27) and (4.28) to obtain the condition
we can use (4.9), (4.15), (4.27) and (4.28) to obtain the condition
 for
for  to vanish to leading order. Note that the h 0 (i.e. δ 0) limit of (4.50) can be calculated using the asymptotic expansion (4.22)-(4.23), yielding
to vanish to leading order. Note that the h 0 (i.e. δ 0) limit of (4.50) can be calculated using the asymptotic expansion (4.22)-(4.23), yielding
 which is equivalent to [13, equation (3.14)] and [48, equation (2.24)].
which is equivalent to [13, equation (3.14)] and [48, equation (2.24)].
As in the stationary case (cf. subsection 4.1.3), the second step in the construction of a pulse solution is to match the fast evolution through  with the evolution on
with the evolution on  , such that the fast excursion normal to
, such that the fast excursion normal to  can be concatenated with the slow flow on
can be concatenated with the slow flow on  along Eu,s (4.38), in order to construct an orbit that is homoclinic to the equilibrium point
along Eu,s (4.38), in order to construct an orbit that is homoclinic to the equilibrium point  . To that end, we need to calculate the change of the slow componentents (u, p) during the fast excursion. Since the dynamical equations for u and p in (4.36) differ from those in the stationary system (4.7) only by an asymptotically small term ε3ε2cp, and the leading order fast flow of (4.41) is the same as that of the stationary system (4.7), the leading order calculations (4.29) apply in the current setting as well. However, as the unstable and stable manifolds of the slow flow (4.38) differ from those in the stationary case (4.25), the resulting matching condition is different, and yields
. To that end, we need to calculate the change of the slow componentents (u, p) during the fast excursion. Since the dynamical equations for u and p in (4.36) differ from those in the stationary system (4.7) only by an asymptotically small term ε3ε2cp, and the leading order fast flow of (4.41) is the same as that of the stationary system (4.7), the leading order calculations (4.29) apply in the current setting as well. However, as the unstable and stable manifolds of the slow flow (4.38) differ from those in the stationary case (4.25), the resulting matching condition is different, and yields

Note that at this point, the asymptotic magnitude of the fraction  is still undetermined. However, as the right hand side of the matching condition (4.52) is
is still undetermined. However, as the right hand side of the matching condition (4.52) is  in ε and ε1,2,3, the left hand side must be as well, hence it follows that either
in ε and ε1,2,3, the left hand side must be as well, hence it follows that either  or
or 
Moreover, we can calculate the value of p0 in the existence condition (4.50) using (4.28), by writing
 by (4.38); note that this is the same calculation as found in [48, equations (2.22-25)] and [13, equation (3.18)]. However, once we substitute this result into (4.50), we obtain the leading order existence condition
by (4.38); note that this is the same calculation as found in [48, equations (2.22-25)] and [13, equation (3.18)]. However, once we substitute this result into (4.50), we obtain the leading order existence condition
 where we observe that c occurs as a (nonzero) prefactor; hence, the leading order value of c cannot be determined from (4.54). It follows that c must be determined from (4.52); to be able to do this, we see that
where we observe that c occurs as a (nonzero) prefactor; hence, the leading order value of c cannot be determined from (4.54). It follows that c must be determined from (4.52); to be able to do this, we see that  , and thus c is the solution of
, and thus c is the solution of

Moreover, to satisfy (4.54), we see that ε δ+β or  must be equal to one, since all integrals
must be equal to one, since all integrals  are positive and at least one between the first
are positive and at least one between the first  and the third
and the third  term must balance
term must balance  ; from the same argument, it follows that neither term can be asymptotically large, hence ε δ+β 1 and
; from the same argument, it follows that neither term can be asymptotically large, hence ε δ+β 1 and  . Thus, three distinct situations arise, where only the first
. Thus, three distinct situations arise, where only the first  term, only the third
term, only the third  term, or both the first and the third term balance the second
term, or both the first and the third term balance the second  term in the existence condition (4.54) In Figure 10, several parameter-dependent solution curves for these three different scaling choices of ε δ+β and
term in the existence condition (4.54) In Figure 10, several parameter-dependent solution curves for these three different scaling choices of ε δ+β and  are shown.
are shown.

Solution curves of the existence condition (4.54), for different scaling choices of ε δ+β and  where a and b are fixed throughout. The set of admissible values, for which
where a and b are fixed throughout. The set of admissible values, for which  is bounded below by the dashed line
is bounded below by the dashed line  . (a) Dominant balance between the second
. (a) Dominant balance between the second  and third
and third  term, i.e.
term, i.e.  and ε δ+β « 1. Here, u0 is independent of d. (b) Dominant balance between the first
and ε δ+β « 1. Here, u0 is independent of d. (b) Dominant balance between the first  and the second
and the second  term, i.e.
term, i.e.  and ε δ+β = 1. Here, u0 is plotted as a function of h for increasing values of d. (c) Equal balance between all three terms, i.e. both
and ε δ+β = 1. Here, u0 is plotted as a function of h for increasing values of d. (c) Equal balance between all three terms, i.e. both  and ε δ+β = 1. Again, u0 is plotted as a function of h for increasing values of d.
and ε δ+β = 1. Again, u0 is plotted as a function of h for increasing values of d.
We summarise the findings of this subsection in the following theorem:
Let ε be sufficiently small. Moreover, let  , and assume at least one of the following conditions holds:
, and assume at least one of the following conditions holds:


Then, system (4.36) admits a homoclinic orbit to the equilibrium ( , 0, 0, 0, 0), provided equation (4.55) admits a real solution for c, where δ is chosen such that condition (4.54) is satisfied to leading order in ε
, 0, 0, 0, 0), provided equation (4.55) admits a real solution for c, where δ is chosen such that condition (4.54) is satisfied to leading order in ε
The homoclinic orbit is, to leading order in ε, given by



Conversely, if either (4.55) has no solution for real c, or (4.54) cannot be satisfied, then no orbit homoclinic to ( , 0, 0, 0, 0) exists.
, 0, 0, 0, 0) exists.
Remark 4.3. The existence result of Theorem 4.2 does not contradict the non-existence result [13, Theorem 5.1] for travelling pulses, as the scaling regimes (4.56a), (4.56b) were not considered in [13]; see also [12, Remark 4.1].
Proof. For sufficiently small ε3ε δ+β, Fenichel’s invariant manifold theorem [22, 30] provides the existence of  , which is invariant under the flow of (4.36) and
, which is invariant under the flow of (4.36) and  close to
close to  (4.11). Specifically, there exists ε0,A > 0 such that for all 0 < ε < ε0,A, the persistence of
(4.11). Specifically, there exists ε0,A > 0 such that for all 0 < ε < ε0,A, the persistence of  as an invariant manifold is guaranteed. On this 4-dimensional invariant manifold
as an invariant manifold is guaranteed. On this 4-dimensional invariant manifold  , the transversal intersection of the restricted unstable and stable manifolds of
, the transversal intersection of the restricted unstable and stable manifolds of  (4.43),
(4.43),  follows from leading order calculations presented in subsection 4.2.2 combined with geometric arguments from the proof of [13, Theorem 4.1] and/or [48, Theorem 2.1]. The latter only needs to be augmented in the construction of the take-off and touchdown curves, where the intersection with the hyperplane q = 0 now needs to be considered in its intersection with
follows from leading order calculations presented in subsection 4.2.2 combined with geometric arguments from the proof of [13, Theorem 4.1] and/or [48, Theorem 2.1]. The latter only needs to be augmented in the construction of the take-off and touchdown curves, where the intersection with the hyperplane q = 0 now needs to be considered in its intersection with  . Geometric perturbation theory now provides the existence of ε0,B > 0 such that for all 0 < ε < ε0,B, a homoclinic orbit to (
. Geometric perturbation theory now provides the existence of ε0,B > 0 such that for all 0 < ε < ε0,B, a homoclinic orbit to ( , 0, 0, 0, 0) exists within
, 0, 0, 0, 0) exists within  ; defining the global upper ε-bound
; defining the global upper ε-bound  completes the proof.
completes the proof.
It is useful to explore the ramifications of the scaling choices made in this subsection for the original system (4.1). As mentioned in Theorem 4.2, the condition  fixes
fixes  which, in combination with condition (4.5), implies β − κ > 0.
which, in combination with condition (4.5), implies β − κ > 0.
The scaling choice (4.56a), which balances  with
with  implies β > 2. Taken together with κ = 2, we find for the scaling exponents of the original model parameters (4.4):
implies β > 2. Taken together with κ = 2, we find for the scaling exponents of the original model parameters (4.4):
 while the sign of δ, the scaling exponent of
while the sign of δ, the scaling exponent of  is not fixed. In particular, we see that
is not fixed. In particular, we see that  scales with a relatively high power of ε; hence, a travelling pulse whose existence is established by Theorem 4.2 with scaling (4.56a), can only be observed for very small values of
scales with a relatively high power of ε; hence, a travelling pulse whose existence is established by Theorem 4.2 with scaling (4.56a), can only be observed for very small values of  . In addition, the h → 0 limit of (4.54) yields u0 = 1 and
. In addition, the h → 0 limit of (4.54) yields u0 = 1 and  (see also Figure 10 (a)). That is, in the classical KGS limit, the amplitude of the pulse does, to leading order, not depend on the system parameters; this is in clear contrast to the stationary pulses studied in this paper (see Theorem 4.1) and to the travelling pulses constructed in [48].
(see also Figure 10 (a)). That is, in the classical KGS limit, the amplitude of the pulse does, to leading order, not depend on the system parameters; this is in clear contrast to the stationary pulses studied in this paper (see Theorem 4.1) and to the travelling pulses constructed in [48].
On the other hand, the scaling choice (4.56a), which balances  with
with  , only implies β > 0. Taken together with 0 < κ ≤ 2, we find for the scaling exponents of the original model parameters (4.4):
, only implies β > 0. Taken together with 0 < κ ≤ 2, we find for the scaling exponents of the original model parameters (4.4):

Compared with (4.58), the conditions on the scaling components are rather mild. Still, the condition β − κ > 0 implies that  – that is,
– that is,  and
and  differ at least by two orders of magnitude in ε. Note that the h → 0 limit of (4.54) yields u0 = 0 (see also Figure 10 (b)), accentuating the fact that this solution branch does not exist in the classical KGS system.
differ at least by two orders of magnitude in ε. Note that the h → 0 limit of (4.54) yields u0 = 0 (see also Figure 10 (b)), accentuating the fact that this solution branch does not exist in the classical KGS system.
4.2.3 Constructing a travelling pulse solution, case II:(v, q, s)-dynamics on the same scale
We assume that the s-dynamics occur on the same scale as the (v, q)-dynamics, that is, ε3ε δ+β = 1. Taking the limit ε → 0 in (4.36) now yields





The construction method of the previous sections can be applied if and only if the reduced system (4.60) admits a homoclinic orbit. In contrast to the previous cases (cf. (4.14) and (4.39)), where the effective phase space was planar, the dynamics of (4.60) are effectively three-dimensional. This poses a considerate analytical challenge, as the dynamics of nonlinear three-dimensional dynamical systems can be highly complex. That being said, it seems sensible to start from a situation where the existence of a homoclinic orbit is known – that is, in the singular limit cd = 0. In this limit, the analysis of section 4.1.1 applies, and provides us with an homoclinic orbit (vh(z), qh(z), sh(z)). Now, we can use Melnikov theory [24, section 2.1] to determine whether the intersection of the stable and unstable manifolds of the origin (v, q, s) = (0, 0, 0) persists for cd > 0. To that end, we use the conserved quantity Hf in terms of the original variables (v, q) (4.45). However, we find that
 is always positive, which suffices to conclude that the intersection for cd = 0 does not persist for small positive values of cd.
is always positive, which suffices to conclude that the intersection for cd = 0 does not persist for small positive values of cd.
While, in principle, the existence of homoclinic orbits in (4.60) for non-small values of cd is not ruled out by the above argument, their analytical inaccessibility makes the situation created by the scaling choice considered in this section, fall outside the scope of this paper. Hence, we disregard the (limiting) case ε3ε δ+β = 1.
4.2.4 Constructing a travelling pulse solution, case III:s slower than(v, q)
We assume that the s-dynamics are slower than the (v, q)-dynamics, that is  as ε → 0. Multiplying (4.36e) with
as ε → 0. Multiplying (4.36e) with  and subsequently taking the limit ε → 0 in (4.36) then yields
and subsequently taking the limit ε → 0 in (4.36) then yields





The existence of a homoclinic orbit in (4.62) now follows directly from the analysis in [13, 48], yielding

In principle, one could mimic the analytical steps taken in subsection 4.2.2, to obtain existence conditions along the lines of (4.54), which would depend on s0. However, for reasons that will become clear below, we will consider the dynamics of the slower variables (u, p, s) first.
The set of equilibria of the reduced system (4.62) is given by the 3-dimensional hyperplane

Like  (4.13),
(4.13),  is invariant under the flow of the full 5-dimensional system (4.36). The (u, p)-dynamics on
is invariant under the flow of the full 5-dimensional system (4.36). The (u, p)-dynamics on  are the same as those on
are the same as those on  , and are therefore given by system (4.24). However, the s-dynamics on
, and are therefore given by system (4.24). However, the s-dynamics on  are given by
are given by
 which are unbounded for z → ∞. Hence, we can conclude that in the scaling chosen in this subsection, any orbit homoclinic to (
which are unbounded for z → ∞. Hence, we can conclude that in the scaling chosen in this subsection, any orbit homoclinic to ( , 0, 0, 0, 0) in (4.36), being necessarily bounded, must have a trivial s-component – that is, must be a solution to the classical KGS system.
, 0, 0, 0, 0) in (4.36), being necessarily bounded, must have a trivial s-component – that is, must be a solution to the classical KGS system.
5 Conclusion and outlook
The aim of this paper is to analytically investigate the influence of autotoxicity on vegetation patterns, through the analysis of travelling pulse solutions to the biomass-water-toxicity model (2.1). In particular, we want to analytically explain the occurence of travelling, asymmetric pulses, as observed in numerical simulations of the biomass-water-toxicity system, see e.g. Figure 3. The numerical evidence presented in section 3 suggests that the presence of toxicity is a prerequisite for the occurence of these travelling pulses in particular (compare Figures 2 and 3), and for previously unobserved dynamic spatio-temporal patterns in general (cf. Figure 1).
However, while the inclusion of an additional model component to the classical KGS model enriches the class of spatial patterns produced by this model with a number of provocative spatiotemporal patterning phenomena, it also significantly increases the mathematical complexity of the model, and subsequently of its analysis. Where the occurence of several types of patterns in the classical KGS model sensitively depends on the asymptotic scaling of the model components and parameters (see e.g. [12, Figure 2]), this is even more so the case in the extended model (2.2) considered in this paper. Therefore, the main emphasis of the analytical part of this paper (section 4) is on the balance of asymptotic scalings. The result of this analysis is that, within the scaling preparatory scaling choices made in appendix B, stationary and travelling pulses can only be constructed in specific asymptotic scalings. This ‘positive’ outcome, as summarised in Theorems 4.1 and 4.2, is balanced by the ‘negative’ outcome of subsections 4.2.3 and 4.2.4.
Comparing the statement of Theorem 4.2 and its ramifications for the parameter scalings in the nondimensionalised model (2.2), it is clear that the travelling pulses constructed in subsection 4.2.2 are not the travelling pulses observed in numerical simulations, shown in Figure 3. In particular, the numerically observed pulses vanish when the parameters  and
and  are not approximately equal (see Table 2), whereas the travelling pulses from Theorem 4.2 only exist when
are not approximately equal (see Table 2), whereas the travelling pulses from Theorem 4.2 only exist when  and
and  differ by two orders of magnitude in ε (cf. (4.58) and (4.59)). This discrepancy between analytical results and numerical observations could be explained in several ways.
differ by two orders of magnitude in ε (cf. (4.58) and (4.59)). This discrepancy between analytical results and numerical observations could be explained in several ways.
Boundary effects. The numerical simulations have been carried out on a bounded domain, whereas the analytical results assume an unbounded spatial domain. Hence, the existence of travelling pulses could be accredited to boundary effects – in particular, to self-interaction through the boundary. This would mean that a single travelling pulse on an unbounded domain is not the proper mathematical abstraction of the numerical observations as in Figure 3. Rather, one should consider a pair of interacting pulses, along the lines of [12]. However, our numerical investigations reveal that the observed travelling pulse solutions persist (both shape and speed) when doubling of the domain length, which is not in line with the hypothesis of self-interaction.
Transient behaviour. The numerical simulations have been carried out for a finite time, whereas the existence analysis is time-independent. Hence, the observed travelling pulses could be the manifestation of a slow transient process from initial state to bare soil, where these travelling pulses only exist for a finite time. This would mean that a time-independent travelling pulse, that is, a stationary pulse in a co-moving frame with fixed speed, is not the proper mathematical abstraction of the numerical observations as in Figure 3. However, our numerical investigations reveal that the observed travelling pulse solutions persist when doubling the simulation time. This does not rule out the hypothesis of metastability, but it does severely limit the evolution speed of the wave profile. Transitional pattern formation phenomena have be studied analytically, see e.g. [3]; one could adopt the approach outlined in [3] to study (2.2). However, the absence of integrable structure might prohibit this approach.
Scaling assumptions. As outlined in the introduction of section 4, the applicability of the geometric singular perturbation theory approach to pulse patterns, as used in [12, 13, 14, 48], is closely tied to the asymptotic scaling of the underlying model (2.2). The arguments leading to the preparatory scaling choices (4.4) are specified in appendix B. However, it is important to note that not every scaling choice made in appendix B is necessary for the application of the geometric construction techniques. The two scaling choices pertaining to the dynamics of the toxicity component s ((B.17) and (B.20)) are convenient, rather than necessary. In particular, the assumption underlying the last scaling choice (B.20) – namely, that the stationary pulse constructed in section 4.1 should be the ‘c = 0’ member of a family of travelling (‘c ≠ 0’) pulses – could prove to be too restrictive. After all, families of travelling waves that do not include a neighbourhood of c = 0 regularly occur [8, 51, 53]. In addition, the ‘proper’ scaling needed to understand the travelling pulse shown in Figure 4 need not be uniform: abandoning scalings (B.17) and (B.20) might lead to the situation where the nonlinear interaction terms govern the singular behaviour of the pulse solution. In such a case, one would need to use geometric blowup techniques to construct a singular concatenated homoclinic orbit, along the lines of [25].
The analysis of travelling pulses incorporating this generalised scaling is ongoing work.
It is important to note that, while the analytical approach advocated in this paper applies to the existence of stationary and travelling pulse solutions, the question of pulse stability is still very much open. We plan to apply the techniques developed for the stability analysis of pulses in two-component reaction-diffusion systems, as presented in [14], to the three-component reaction-diffusion-ODE system (2.2). The method presented in [14], which is based on an Evans function approach, does not in principle depend on the number of model components, and can be applied to pattern solutions in n-dimensional reaction-diffusion systems [47]. Moreover, this approach is amenable to be extended to systems of mixed (reaction-diffusion-ODE) type. The stability of pulse solutions to the biomass-water-toxicity model (2.1) will be investigated in a separate work.
Moreover, numerical simulations in [35] on two-dimensional domains reveal the presence of both crescent (travelling) moon spots and double-scale patterns for some parameter regimes. In these double-scale pattens, the pulses/fronts travel at a micro-scale level within a pattern, which appears stable on a macroscopic scale. Such multiple-scale behaviour has been connected with pattern robustness in a different, but related, ecological context [33]. Future challenges hence involve the analytical investigation of such structures – in the first case (travelling crescent moons), an approach along the lines of [21] would be a prime candidate.
The biomass-water-toxicity model by Marasco et al. (2.1) has proven to be a rich and inspiring source of previously unobserved patterning phenomena. It is important to emphasise that autotoxicity can be used to explain and recreate experimentally observed dynamical patterns, without having to assume a specific domain topography, in contrast to previous work [1, 2, 52]. In this respect, the model is interesting both from an ecological perspective, and from the more general viewpoint of mathematical modelling. Systems of reaction-diffusion-ODE type have been the subject of recent investigations; in particular, the shape and stability properties of patterns have proven to be significantly different from their ‘classical reaction-diffusion’ counterparts, sometimes leading to counterintuitive results [36, 37]. We hope that the work presented in this paper, though exploratory, will lead to a deeper understanding and a broader appreciation of systems of this type, of which the biomass-water-toxicity model (2.1) is an intriguing example.
Acknowledgements
AI acknowledges partial support from the New Frontier’s grant NST-0001 of the Austrian Academy of Sciences ÖAW. FV was supported by a Humboldt Research Fellowship.
A Nondimensionalisation and rescaling
We take the original Marasco et al. model (2.1), and define the nondimensional space and time variables

Next, we define the nondimensional model components

The model in these new model components (U, V, S) takes the form (2.2), with (nondimensional) parameters defined as
 while the (square root of the) diffusivity ratio gives rise to the natural small parameter
while the (square root of the) diffusivity ratio gives rise to the natural small parameter

Note that decoupling the autotoxicity equation by setting  is equivalent to taking the toxicity sensitivity s = 0.
is equivalent to taking the toxicity sensitivity s = 0.
B Preparatory asymptotic scaling of system (4.3)
Our goal is to scale system (4.3) in such a way, that we can apply the approach taken in e.g. [13, 14, 48] to construct a travelling pulse solution. For similar asymptotic scaling analyses (with the same goal), see [13, Appendix] and [48, section 2.1].
A priori, every parameter, component, and variable in system (4.3) admits an asymptotic scaling in the small parameter ε. Hence, we scale
 and introduce a rescaled coordinate
and introduce a rescaled coordinate  to obtain
to obtain



We now choose z to be the variable on which the (fast) v-dynamics take place. Moreover, we assume that the equilibrium (u, v, s) = (ε−κ, 0, 0) is hyperbolic in v. This fixes the asymptotic scaling of the coordinate z as
 which, dividing the second equation in (B.2) by εβ+λ, yields
which, dividing the second equation in (B.2) by εβ+λ, yields



In the geometric construction as carried out in [13, 14, 48], a pivotal element is the existence of a homoclinic orbit (spike) in v for a fixed value of u. In particular, this means that the initial exponential growth close to v = 0 due to the term −bv in (B.4) must be balanced by a positive, nonlinear term. We fix the asymptotic scaling of v such, that such an homoclinic spike solution has  amplitude in v, which implies
amplitude in v, which implies
 With this scaling, dividing the third equation in (B.4) by εσ, we obtain
With this scaling, dividing the third equation in (B.4) by εσ, we obtain



Furthermore, based on the numerical observations from section 3, we conjecture that a homoclinic spike solution in v is, to leading order, symmetric in z. This implies that the advection term cvz in the v-equation is perturbative, hence

Next, we consider the behaviour of u. For the type of pulse solutions we consider in this paper, there is a clear spatial scale separation between the u-component and the v-component, see Figure 4. In particular, u is slow in z in comparison to v. Rewriting the first equation in (B.6) as
 this implies that
this implies that
 where the third inequality already follows from (B.7). Moreover, both from the simulation results presented section 3 and the interpretation of the model (2.2) in light of its dimensional version (2.1), it is clear that the U-variable takes values in between the homogeneous dry (U = 0) and the homogeneous wet (U = 1) states. As U = εκu, this implies
where the third inequality already follows from (B.7). Moreover, both from the simulation results presented section 3 and the interpretation of the model (2.2) in light of its dimensional version (2.1), it is clear that the U-variable takes values in between the homogeneous dry (U = 0) and the homogeneous wet (U = 1) states. As U = εκu, this implies

The coupling of v into the u-equation plays a central role in the geometric construction outlined in section 4 of this paper, and in the main references [12, 13, 14, 48]. In particular, the question is whether the stable and unstable manifolds of the invariant manifold  (4.13) intersect transversally, thereby guaranteeing the existence of a (travelling) pulse solution to (2.2). A necessary condition for this intersection to exist, is that the uv2-term in (B.8) is leading order – that is,
(4.13) intersect transversally, thereby guaranteeing the existence of a (travelling) pulse solution to (2.2). A necessary condition for this intersection to exist, is that the uv2-term in (B.8) is leading order – that is,

Moreover, this term should match, in a specific way, the dynamics of u on  – that is, the dynamics of u when v = 0. From (B.8), we see that these dynamics are linear, with u = ε−κ being the only equilibrium. This equilibrium is a saddle, with stable resp. unstable eigenvalues
– that is, the dynamics of u when v = 0. From (B.8), we see that these dynamics are linear, with u = ε−κ being the only equilibrium. This equilibrium is a saddle, with stable resp. unstable eigenvalues

We now assume that there exists an asymptotic scaling such that both eigenvalues (B.12) are nonzero and of the same order, which implies

For the same eigenvalues, we consider the associated linear stable and unstable subspaces Es and Eu in (u, uz)-phase space. The geometric matching condition now stipulates that the vertical distance between these two subspaces, which is given by
 should be of the same asymptotic order as the leading order term in (B.8), that is, ε2+β−2κuv2. We use this condition to fix the scaling of
should be of the same asymptotic order as the leading order term in (B.8), that is, ε2+β−2κuv2. We use this condition to fix the scaling of  , yielding
, yielding

This scaling choice further specifies (B.6) as



Next, we consider the aim of this paper – that is, to investigate the influence of toxicity on the dynamics of vegetation patterns. The coupling between the toxicity component s and the biomass component v is mediated by the term hvs. We fix the scaling of the parameter  such, that the toxicity-coupling term has
such, that the toxicity-coupling term has  influence on the biomass dynamics, which implies
influence on the biomass dynamics, which implies

Applying this scaling choice to system (B.16), we obtain



As a last scaling choice, we consider the question of existence of stationary pulse solutions – that is, when  in (4.2), and as a consequence, c = 0 in (B.18). From the resulting (algebraic) s-equation
in (4.2), and as a consequence, c = 0 in (B.18). From the resulting (algebraic) s-equation
 it follows that for a stationary pulse to have a nontrivial s-component, the terms in (B.19) all need to have the same asymptotic scaling. When one considers a stationary pulse solution to be a particular travelling pulse solution –namely, one with zero speed–, it can be argued that both solution types have to exist within the same asymptotic scaling. Hence, we decide to fix the asymptotic amplitude of s as
it follows that for a stationary pulse to have a nontrivial s-component, the terms in (B.19) all need to have the same asymptotic scaling. When one considers a stationary pulse solution to be a particular travelling pulse solution –namely, one with zero speed–, it can be argued that both solution types have to exist within the same asymptotic scaling. Hence, we decide to fix the asymptotic amplitude of s as

With this scaling, dividing the first equation of (B.18) by εβ−2+κ, we obtain


 which is equivalent to the 5-dimensional system (4.6).
which is equivalent to the 5-dimensional system (4.6).
References
- [1].↵
- [2].↵
- [3].↵
- [4].↵
- [5].↵
- [6].↵
- [7].↵
- [8].↵
- [9].↵
- [10].↵
- [11].↵
- [12].↵
- [13].↵
- [14].↵
- [15].↵
- [16].↵
- [17].
- [18].
- [19].
- [20].↵
- [21].↵
- [22].↵
- [23].↵
- [24].↵
- [25].↵
- [26].↵
- [27].↵
- [28].↵
- [29].↵
- [30].↵
- [31].↵
- [32].↵
- [33].↵
- [34].↵
- [35].↵
- [36].↵
- [37].↵
- [38].↵
- [39].↵
- [40].↵
- [41].↵
- [42].↵
- [43].↵
- [44].↵
- [45].↵
- [46].↵
- [47].↵
- [48].↵
- [49].↵
- [50].↵
- [51].↵
- [52].↵
- [53].↵
- [54].↵





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}