

ABSTRACT
Cytokinins play an important role in plant stress responses via a multistep signaling pathway, involving the histidine phosphotransfer proteins (HPs). Here, the AHP4 possesses a negative regulatory role in drought response in Arabidopsis thaliana as evidenced by higher survival rates of ahp4 than wild-type (WT) plants under drought, and the down-regulated AHP4 expression in WT during dehydration. Transcriptome analysis of ahp4 and WT plants revealed AHP4-mediated expression of dehydration- and/or abscisic acid (ABA)-responsive genes important for plant drought acclimation. ahp4 plants showed increased wax crystal accumulation in stems, thicker cuticles in leaves, greater sensitivity to exogenous ABA, narrower stomatal apertures, higher leaf temperatures during dehydration, and longer root length under osmotic stress. Additionally, ahp4 plants showed greater photosynthetic efficiency, lower levels of reactive oxygen species, reduced electrolyte leakage and lipid peroxidation, and increased anthocyanin contents under drought. Findings could make editing of AHP4 a promising approach for generating drought-tolerant crops.
INTRODUCTION
Abiotic stresses, including drought, have detrimental effects on the growth and productivity of many important crops, resulting in significant yield losses that may lead to food shortages and threaten agricultural sustainability (Daryanto et al., 2016; Abdelrahman et al., 2018; Lamaoui et al., 2018). To cope with environmental stresses, plants have developed a range of cooperative physiological, biochemical and molecular mechanisms regulated by complex signaling networks (Sah et al., 2016; Choudhury et al., 2017). Phytohormones, the key regulators of plant growth and development, control different physiological and biochemical processes in plant responses to environmental stresses (Peleg and Blumwald, 2011; Verma et al., 2016; Mostofa et al., 2018; Wybouw and De Rybel, 2019). Among the various phytohormones, cytokinin (CK) is one of the central regulators in plant abiotic stress responses, and coordinates an array of functions, enabling plants to adapt to different types of stress (Ha et al., 2012; Zwack and Rashotte, 2015; Li et al., 2016; Cortleven et al., 2018; Li et al., 2019). CK has been reported to play a protective role in plant drought tolerance through enhancement of the endogenous CK level at the onset of drought in transgenic plants (Prerostova et al., 2018). More recently, by studying various CK-deficient and CK-signaling mutants, CK was found to play a negative regulatory role in plant drought responses (Li et al., 2016; Cortleven et al., 2018; Ramireddy et al., 2018). Thus, CK has been suggested to have multifaceted actions, playing both stimulatory and quiescent roles in plant drought tolerance (Prerostova et al., 2018).
The CK signaling pathway starts with histidine-kinase receptors (HKs), moves through histidine phosphotransfer proteins (HPs), and terminates in the activation of response regulators (RRs) which mediate the expression of downstream genes (Kieber and Schaller, 2018; Romanov et al., 2018). In Arabidopsis thaliana, there are three membrane-bound CK receptors (AHK2, AHK3 and CRE1/AHK4), which consist of a conserved CK-binding domain, a histidine kinase domain and a receiver domain (Keshishian and Rashotte, 2015; Pekarova et al., 2016; Romanov et al., 2018). Because the receptors are fixed to the membrane of endoplasmic reticulum, and RRs are primarily located in the nucleus, the intermediate HPs are necessary to help to relay the CK signal through the pathway (Keshishian and Rashotte, 2015; Romanov et al., 2018). There are five authentic AHPs (AHP1, 2, 3, 4 and 5) in Arabidopsis, which act as mediators in the multistep phosphorelay by transferring a phosphoryl group from the receiver domain of an activated AHK receptor to the receiver domain of an ARR (Keshishian and Rashotte, 2015; Kieber and Schaller, 2018). There are 24 RRs in Arabidopsis divided into two typical ARR groups, type-A (11 members) and type-B (10 members), and one atypical type-C (3 members) (Wybouw and De Rybel, 2019). Both type-A and type-B ARRs contain receiver domains; however, only type-B ARRs have a long C-terminal region that includes a MYB-like DNA-binding domain. Type-B ARRs, which play positive regulatory roles in CK signaling, function as transcription factors (TFs) in the final step of CK signaling by regulating downstream target gene expression, whereas type-A ARRs act as negative feedback regulators of CK signaling (Keshishian and Rashotte, 2015; Zwack and Rashotte, 2015; Kieber and Schaller, 2018; Romanov et al., 2018; Wybouw and De Rybel, 2019). Several type-B ARRs, namely ARR1, ARR2 and ARR12, were shown to be degraded by the kiss me deadly (KMD) F-box proteins (Kim et al., 2013).
Previous studies in Arabidopsis reported that several CK signaling members showed negative and redundant roles in drought responses of Arabidopsis plants, which included AHK2 and 3, AHP2, 3 and 5, type-B ARR1, 10 and 12 (Tran et al., 2007; Kang et al., 2012; Nishiyama et al., 2013; Kumar and Verslues, 2015; Nguyen et al., 2016). Among the 5 authentic AHPs, AHP4 is intriguingly distinct from the other AHPs not only evolutionarily, but also functionally. For instance, whereas AHP1, AHP2, AHP3, and AHP5 have been shown to act as positive regulators and have partially redundant functions in CK signaling, AHP4 was reported to play a negative regulatory role in CK signaling in some cases, such as later root formation (Hutchison et al., 2006). This raises a critical question regarding how the AHP4 acts within the CK signaling pathway in plant response to water deficit.
In the present study, using the loss-of-function approach, we initially characterize the detailed functions of AHP4 in regulating the response of Arabidopsis to drought. In contrast with its function in later root formation, our results indicate that AHP4 plays a negative role in plant response to drought similar to that played by AHP2, AHP3 and AHP5. Furthermore, this study showed increased responsiveness to ABA in ahp4 mutant plants, which may contribute to higher drought tolerance of ahp4 relative to wild-type (WT), and provide evidence for crosstalk between ABA and CK signaling. Additionally, higher leaf relative water content (RWC), lower leaf temperature, maintenance of higher cell membrane stability, increased reactive oxygen species (ROS)-scavenging enzyme activities and enhanced anthocyanin biosynthesis were all identified to potentially contribute to the enhanced drought tolerance of the ahp4 mutant plants. Comparative transcriptome analysis of the ahp4 mutant and WT plants revealed several potential pathways, including ROS detoxification- and anthocyanin biosynthesis-related pathways, which were activated in ahp4 plants under both normal and water-deficit conditions, further strengthening our findings.
RESULTS
AHP4 is Down-regulated in Arabidopsis Plants during Dehydration
To determine the involvement of AHP4 in dehydration responses, we first used real-time quantitative PCR (RT-qPCR) to analyze the expression of the AHP4 gene in Arabidopsis WT plants exposed to a dehydration treatment. Twenty-one-day-old WT plants were grown on plates of germination medium and exposed to dehydration treatment under ambient conditions. Rates of plant water loss were recorded during the treatment (Fig. 1A). AHP4 expression in both whole plants and shoots rapidly declined in response to dehydration (Fig. 1B), with the lowest levels of expression of AHP4 observed after 4 h of dehydration, followed by an incomplete recovery of expression after 6 h of dehydration (Fig. 1B). AHP4 expression in the roots of WT plants did not show the consistent responses observed in both whole plants and shoots under dehydration, but instead it showed a slight initial increase followed by a decline to a minimal value after 4 h of dehydration, finally followed by an increase in expression (Fig. 1B). The well-known dehydration-responsive responsive to desiccation 26 (RD26) gene (Fujita et al., 2004) was used as a marker to check the efficacy of the dehydration treatment (Fig.1C). The data presented in Fig. 1C showed that the RD26 expression levels in the shoots, roots and whole plants were elevated throughout the dehydration treatment, with the highest levels of expression found after 4 h of dehydration (∼76% water loss). Results of expression analysis suggest a dehydration-related function for AHP4 in Arabidopsis plants.

A, Water loss rate of 21-day-old WT plants grown on germination medium and subjected to a dehydration treatment. Data represent the means and standard errors (SEs) (n = 5). B, Expression of the AHP4 gene in 21-day-old WT plants subjected to dehydration treatment. C, Expression of the stress-inducible responsive to desiccation 26 (RD26) gene, which was used as a marker gene for checking the efficacy of dehydration treatment. Relative expression levels were normalized to a value of 1 in the respective control plants. Data represent the means and SEs (n = 3). Asterisks indicate significant differences as determined by a Student’s t-test (*P < 0.05, **P < 0.01, ***P < 0.001).
Arabidopsis ahp4 Mutant Plants Have Improved Drought Tolerance
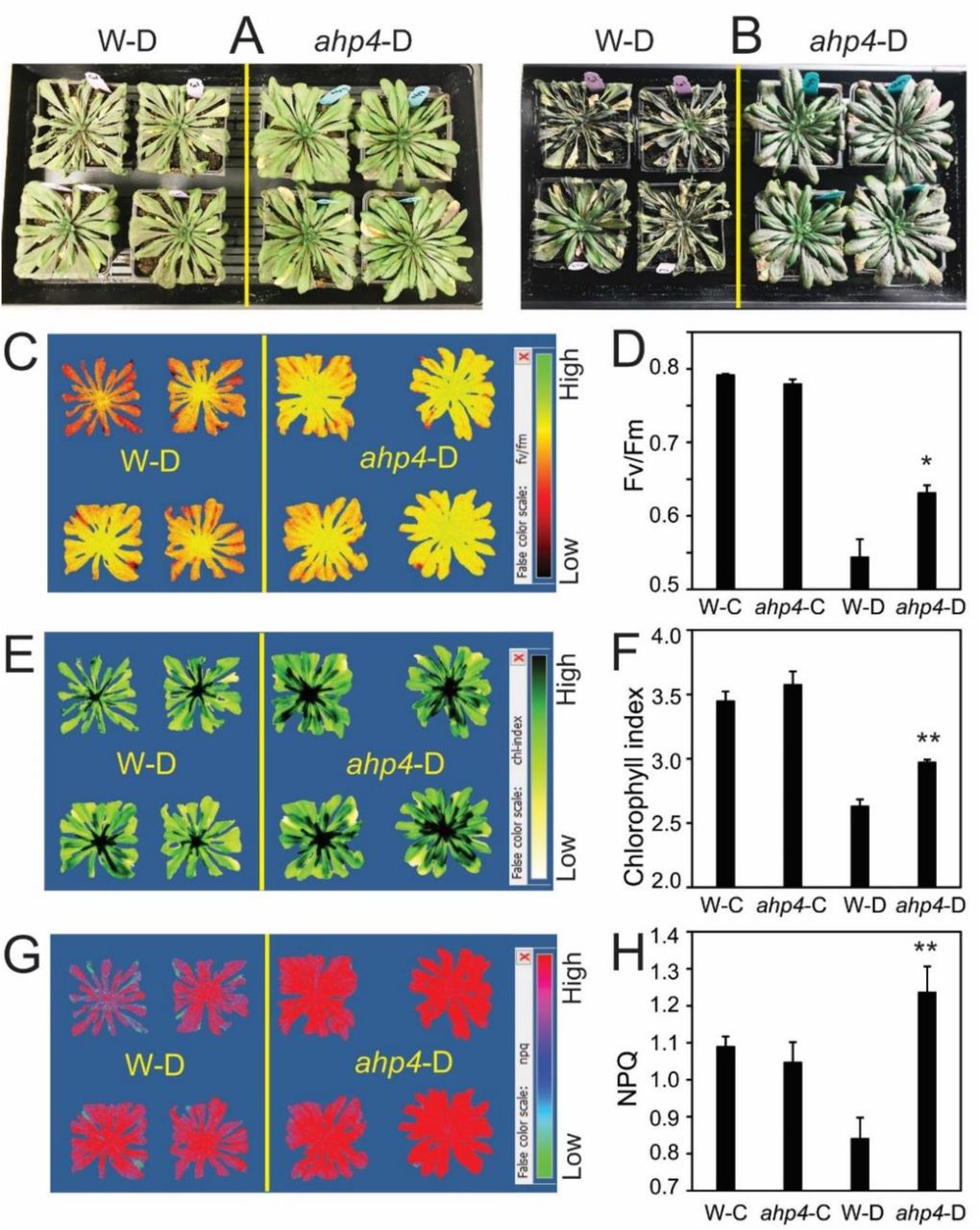
To further elucidate the role of the AHP4 gene in plant response to drought, a loss-of-function approach was used. The ahp4 mutant and WT plants were tested for drought tolerance using biomass reduction (Fig. 2), recovery (Fig. 3) and survival assays (Supplementary Fig. S1). Under well-watered conditions, ahp4 mutant plants showed insignificant difference in shoot growth compared with WT plants (Fig. 2A-B; Supplementary Fig. S1A, C). However, under water-deficit conditions ahp4 mutant plants had a greater plant biomass than WT plants (Fig. 2B). Compared with the well-watered plant groups, the stressed WT and ahp4 mutant plants showed 37.1 and 17.7% biomass reduction, respectively (Fig. 2C). Furthermore, ahp4 plants had better recovery (Fig. 3A-B) and higher survival rates than WT plants under water-deficit conditions (Supplementary Fig. S1B, D). Moreover, to test whether drought stress affects the photosynthetic system, we measured the maximum potential quantum efficiency of photosystem (PS) II (Fv/Fm), chlorophyll index and nonphotochemical chlorophyll fluorescence quenching (NPQ). Data showed that ahp4 mutant plants exhibited greater Fv/Fm (Fig. 3C-D), chlorophyll index (Fig. 3E-F) and NPQ (Fig. 3G-H) than WT plants under water-deficit conditions, suggesting that ahp4 mutant plants had lower photoinhibition and higher photosynthetic efficiency under drought than WT plants. In addition, we observed that the primary roots of ahp4 mutant plants grew better than that of WT plants, when they were treated with 200 or 300 mM of mannitol (Supplementary Fig. S2), suggesting that AHP4 controlled primary root growth in response to mannitol-induced water stress. Taken together, these results clearly indicated that loss-of-function of the AHP4 gene resulted in a drought-tolerant phenotype, and that AHP4 acts as a negative regulator of responses involved in drought tolerance of Arabidopsis plants.

A, Representative rosettes of ahp4 and wild-type (WT) plants in the well-watered control and soil-drying treatments. B, Biomass of aph4 and WT plants under well-watered and soil-drying conditions. Data represent the means and standard errors (SEs) (n = 12/genotype). C, Biomass reduction of soil-dried aph4 and WT plants relative to respective well-watered control plants. Data represent the means and SEs (n = 12/genotype). Asterisks indicate significant differences between treatments as determined by a Student’s t-test (***P < 0.0001). D, Averaged losses of ahp4 and WT pot weights relative to initial pot weight during soil-drying (n = 12/genotype). Black arrow reveals when water was added to 70% of the initial pot weight. Blue arrow reveals when biomass was measured.

A, Representative of ahp4 and WT plants after 14 days of drought. B, Representative of ahp4 and WT plants were exposed to drought for 15 days, and then rewatered for 3 days. C, Representative image of Fv/Fm of ahp4 and WT plants after 14 days of drought. D, The Fv/Fm of 49-day-old ahp4 and WT plants under well-watered control conditions and after 14 days of drought treatment. E, Representative image of Chl index of ahp4 and WT plants after 14 days of drought. F, Chl index of 49-day-old ahp4 and WT plants under well-watered control conditions and after 14 days of drought treatment. G, Representative image of NPQ of ahp4 and WT plants at 14 days of drought. H, NPQ of 49-day-old ahp4 and WT plants under well-watered control conditions and after 14 days of drought treatment. Data represent the means and standard errors (SE) (n = 4). Asterisks indicate significant differences as determined by a Student’s t-test (*P < 0.05, **P < 0.01). W-C, WT under well-watered control; W-D, WT under drought; ahp4-C, ahp4 under well-watered control; ahp4-D, ahp4 under drought.
Comparative Transcriptome Analyses of ahp4 and WT Plants under Non-stressed and Dehydration Conditions
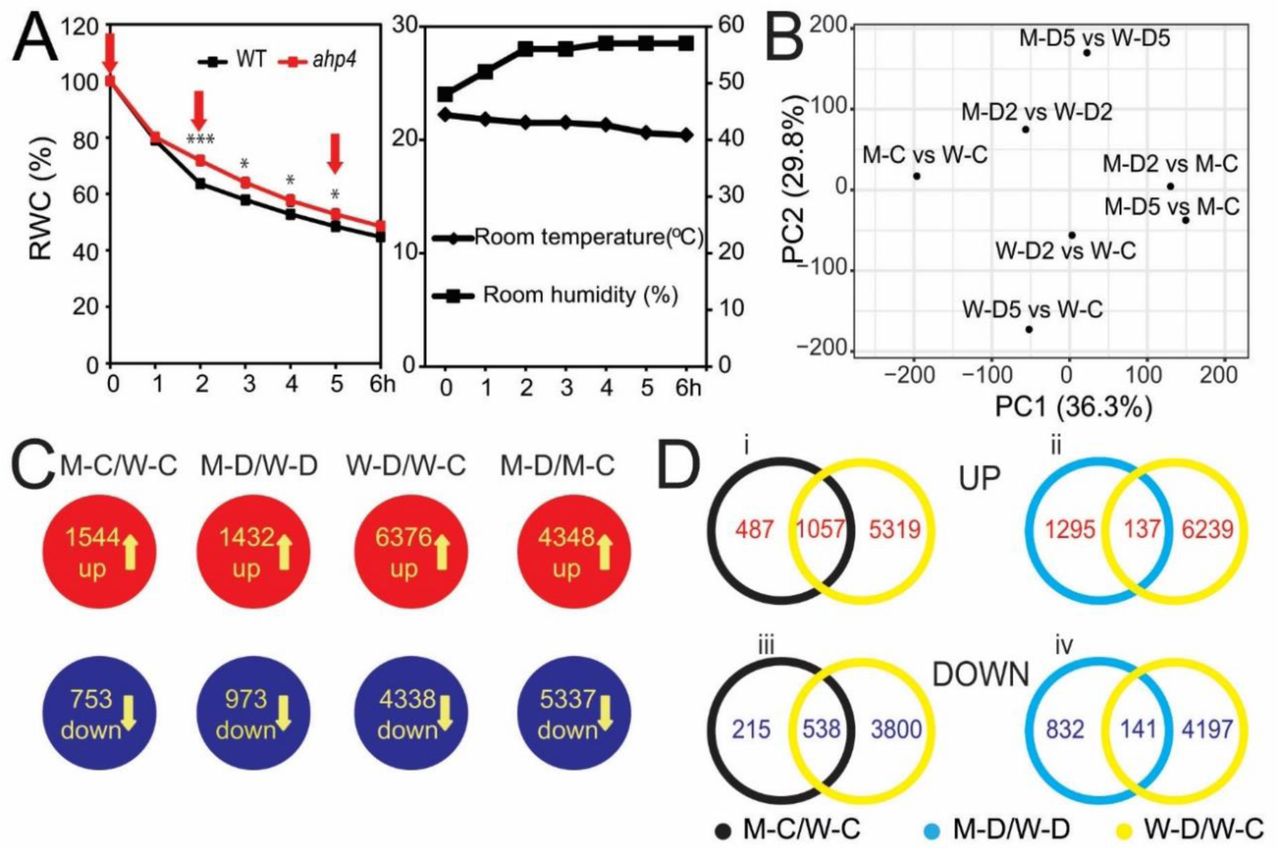
Based on AHP4 expression and drought-tolerant test results, we performed a microarray analysis to investigate AHP4-dependent CK signaling-mediated downstream genes involved in plant adaptation to water deficiency. The experimental design for comparing transcriptome data obtained from the leaves of ahp4 and WT plants under non-stressed and dehydration conditions are illustrated in Fig. 4A, and the results of the microarray analysis are summarized in Fig. 4B-D and Supplementary Table S1. Confirmation of the microarray analysis, which was carried out using RT-qPCR of five selected genes, supported the results of microarray data (Supplementary Fig. S3). Compared with WT plants under non-stressed conditions (M-C/W-C), the analysis of differentially expressed genes (DEGs) found 1544 up-regulated and 753 down-regulated genes (Fig. 4C; Supplementary Table S2A-B). Comparison of DEGs in ahp4 versus WT plants under dehydration (M-D/W-D) revealed a total of 1432 up-regulated genes (Fig. 4C; Supplementary Table S3A), with more genes being up-regulated after 2-h (M-D2/W-D2) dehydration than after 5-h (M-D5/W-D5) dehydration (Supplementary Table S3B-C). A smaller number of genes (973) were down-regulated in (M-D/W-D) (Fig. 4C; Supplementary Table S3D), with more genes being repressed with increased duration of dehydration (Supplementary Table S3E-F).

A, Fourteen-day-old ahp4 and WT plants grown on GM plates were transferred to soil and grown for 10 additional days under well-watered conditions. The aerial portions of 24-day-old ahp4 and WT plants were subjected to a dehydration stress and recorded the relative water content (RWC). Data represent the means and standard errors (n = 5/genotype). Asterisks indicate significant differences between the two genotypes as determined by a Student’s t-test (*P < 0.05, ***P < 0.001). Rosette leaves, which were collected at 0, 2, and 5 hours (h) (arrows) of dehydration treatment, were used for microarray analysis. Room temperature and relative room humidity were recorded during the dehydration treatment. B, Principal component (PC) analysis plots of ahp4 and WT in various comparisons. C, Diagrams showing the compilation of genes with changed expression in each comparison. Data were extracted from the results of three independent microarray experiments. D, Venn diagram analysis showing the overlapping and non-overlapping up- and down-regulated gene sets. M-C/W-C, ahp4 well-watered control 0 h versus WT well-watered control 0 h; M-D2/W-D2, ahp4 dehydrated 2 h versus WT dehydrated 2 h; M-D5/W-D5, ahp4 dehydrated 5 h versus WT dehydrated 5 h; W-D2/W-C, WT dehydrated 2 h versus WT well-watered control 0 h; W-D5/W-C, WT dehydrated 5 h versus WT well-watered control 0 h; M-D2/M-C, ahp4 dehydrated 2 h versus ahp4 well-watered control 0 h; M-D5/M-C, ahp4 dehydrated 5 h versus ahp4 well-watered control 0 h; M-D/W-D, ahp4 dehydrated 2 and/or 5 h versus WT dehydrated 2 and/or 5 h; M-D/M-C, ahp4 dehydrated 2 and/or 5 h versus ahp4 well-watered control 0 h; W-D/W-C, WT dehydrated 2 and/or 5 h versus WT well-watered control 0 h.
Using Venn diagram analysis, many dehydration-inducible genes belonging to the groups of genes were identified as being up-regulated in the M-C/W-C and/or M-D/W-D comparisons (Fig. 4D, i-ii; Supplementary Tables S2A, S3A and S4A-B). Dehydration-repressible genes were also found to belong to the groups of genes that were down-regulated genes in the M-C/W-C and/or M-D/W-D comparisons (Fig. 4D, iii-iv; Supplementary Tables S2B, S3D and S4C-D). These changes in gene expression likely resulted in drought-tolerant enhancement in ahp4 mutant plants. MapMan analysis was then used to classify the DEGs, identified from the M-C/W-C and/or M-D/W-D comparisons, into functional groups (Supplementary Fig. S4), and to provide an overview of changes in general metabolism in ahp4 versus WT plants (Supplementary Fig. S5) under non-stressed and dehydration conditions.
A detailed analysis of the DEGs obtained from the comparison of ahp4 versus WT transcriptomes under non-stressed and dehydration conditions was then conducted to identify dehydration- and/or ABA-inducible genes, whose up-regulation might contribute to the drought-tolerance of the ahp4 mutant plants. A number of dehydration- and/or ABA-inducible DEGs were annotated to encode AP2- and MYB-type TFs, C2H2-like zinc finger proteins, leucine-rich repeat kinases, glycine-aspartate-serine-leucine (GDSL) esterase/lipase, late embryogenesis abundant (LEA) proteins, α/β-hydrolases, β-glucosidases and glycoside hydrolases (Supplementary Tables S2 and S3). Many of the dehydration-inducible genes found to be up-regulated in dehydrated ahp4 versus dehydrated WT plants (M-D/W-D) were associated with secondary metabolism, which included genes that are known to be involved in flavonoid biosynthesis like those encoding flavonol synthase 4, UDP-glucosyl transferase 73C6 and UDP-glucosyl transferase 78D1 (Supplementary Figs. S5 and S6, Table S3A). Furthermore, several genes associated with cuticular wax biosynthesis, including AT1G34490, AT1G34500, AT5G51420 and AT5G55360, were found to be up-regulated in ahp4 mutant plants under dehydration (M-D/W-D) (Supplementary Figs. S5 and S6, Table S3A). In addition, several ROS-related genes were found to have altered expression patterns in dehydrated ahp4 versus dehydrated WT plants, suggesting that alteration in ROS metabolism could also participate in drought tolerance of ahp4 mutant plants (Supplementary Fig. S4, Tables S2 and S3). In summary, the above results suggest that the enhanced drought tolerance of ahp4 mutant plants (Figs. 2-3; Supplementary Fig. S1) was the result of differential regulations of a set of DEGs associated with a range of metabolic processes.
Reduced Stomatal Apertures, Induced Photosynthetic Efficiency and Enhanced ABA Responsiveness in ahp4 Plants
Plants respond to drought/dehydration stress by inducing stomatal closure and/or reducing stomatal density which help them retain water under water-deficit conditions. To determine the role of AHP4 in controlling stomatal activity, we conducted several assays under both normal and dehydration conditions. As reported earlier under drought (Supplementary Fig. S1F) and long-term dehydration (Fig. 4A) conditions, ahp4 plants also showed greater RWC than WT plants during the short-term dehydration (Fig. 5A). Furthermore, ahp4 plants exhibited higher leaf temperatures than WT plants under both non-stressed and the same dehydration conditions (Fig. 5B), suggesting a relationship between water transpiration and stomatal density and/or stomatal movement. Because no significant differences were observed in stomatal density between ahp4 mutant and WT plants (Fig. 5C), the higher leaf temperatures observed in ahp4 plants (Fig. 5B) resulted from their narrower stomatal apertures in comparison with WT plants under both non-dehydrated and dehydrated conditions as shown in Fig. 5D-E. Additionally, using different light intensity treatments, we found that the stomatal conductance and transpiration rates of ahp4 mutant showed lower trends than WT plants under non-stressed conditions, but they were comparable under dehydration conditions (Supplementary Fig. S7).

A, RWC of 32-day-old soil-grown ahp4 and WT plants subjected to a dehydration treatment. Data represent the means and standard errors (SEs) (n = 5). B, Relative leaf temperatures of 32-day-old soil-grown ahp4 and WT plants were recorded during the dehydration treatment. The rainbow color scale indicates the relative temperatures. C, Average stomatal densities of rosette leaves from 32-day-old soil-grown ahp4 and WT plants. Stomata were counted from eight different areas on each leaf. Data represent the means and SEs (n = 5). D, Guard cells of 32-day-old soil-grown ahp4 and WT plants were subjected to a dehydration treatment for 0 (Control), 30 and 60 minutes. E, Average size of the stomatal aperture of rosette leaves from 32-day-old soil-grown ahp4 and WT plants subjected to a dehydration treatment for 0 (Control), 30 and 60 minutes. Data represent the mean and SEs (n = 5 plants/genotype; for each plant the average of 22 stomatal measurements from a single leaf was calculated). Asterisks indicate significant differences between the two genotypes as determined by a Student’s t-test (*P < 0.05, **P < 0.01, ***P < 0.001).
In addition, ABA-mediated regulation of stomatal opening/closure, and/or ABA-associated mechanisms that promote cellular dehydration tolerance are all important for drought tolerance in plants (Osakabe et al., 2014; Kuromori et al., 2018). To determine if any such ABA-related relationships could be responsible for the differences in the RWC of ahp4 and WT plants and contributed to the differences in drought tolerance, we next compared the stomatal apertures of the two genotypes with or without exogenous ABA treatment. In agreement with stomatal closure data shown in Fig. 5D-E under non-stressed conditions, ahp4 plants showed narrower stomatal apertures than WT did in the absence of ABA; however, both ahp4 and WT showed comparable stomatal apertures in the presence of ABA (Supplementary Fig. S8). This collective data demonstrated that the lower rate of water loss in ahp4 mutant plants, relative to that of WT plants, was caused by a decrease in water transpiration rate due to the impairment of stomatal closure, contributing to the increased drought tolerance of ahp4 mutant plants.
To determine if increased drought tolerance of the ahp4 mutant plants was also associated with increased ABA responsiveness, which may induce downstream ABA-responsive genes that are not related to stomatal activity processes (Fujita et al., 2005), we conducted a seed germination and root growth assay on medium supplemented with different concentrations of ABA. Results showed that the ahp4 mutant plants were more sensitive to ABA than WT plants at both germination and seedling stages (Supplementary Fig. S9A-D). Furthermore, we observed that the expression of AHP4 gene in WT plants was significantly down-regulated in whole plants and roots treated with ABA at all the time points of dehydration period (Supplementary Fig. S9E). Interestingly, AHP4 expression was mostly un-changed in shoots of ABA-treated WT plants at earlier time points, and was then significantly down-regulated after 6 h of dehydration (Supplementary Fig. S9E). These findings indicate that enhanced response of ahp4 mutant plants to ABA might contribute to their improved drought tolerance, and repression of AHP4 transcription by stress-induced ABA might help plants adapt to adverse environmental conditions.
Enhancement of Cell Membrane Integrity and Cuticle Thickness of ahp4 Plants
The greater RWC observed in ahp4 mutant plants compared with WT plants under water deficit suggests the possibility of changes in cell membrane integrity (Figs. 4A and 5A; Supplementary Fig. S1F) (Verslues et al., 2006). To determine if there were any differences in cell membrane integrity induced by water deficit, tissue electrolyte leakage in ahp4 and WT plants under drought was measured (Supplementary Fig. S1G). Our data demonstrated that the loss-of-function mutation in AHP4 led to a considerably lower electrolyte leakage in ahp4 mutant than in WT plants under water deficit (Supplementary Fig. S1G).
The cuticle, which composes of cutin and waxes, covers the epidermis and controls the water permeability, limiting non-stomatal water loss (Sieber et al., 2000; Yeats and Rose, 2013; Jetter and Riederer, 2016). We next evaluated whether there is difference in cuticle structure between ahp4 and WT plants, which would differentiate the transpiration rates of ahp4 mutant and WT plants. First, we conducted a chlorophyll leaching assay of rosette leaves of ahp4 mutant and WT plants grown under normal conditions to compare cuticle permeability. The chlorophylls were found to leach more slowly from ahp4 leaves than WT leaves (Fig. 6A), suggesting lower cuticle permeability in ahp4 compared with WT plants. Second, toluidine blue (TB) staining images showed lower TB uptake by the leaves of ahp4 mutant as compared with that of WT plants (Fig. 6B). These data infer that loss-of-function of AHP4 might result in enhanced cuticle structure of ahp4 mutant plants, which could prevent non-stomatal water loss under adverse conditions. Third, scanning electron microscopy (SEM) imaging of the surface wax ornamentation of the stems of ahp4 and WT plants indicated a higher density of surface wax in ahp4 mutant versus WT plants (Fig. 6C). Fourth, transmission electron microscopy (TEM) imaging of the cuticle layers of the fifth leaves showed thicker cuticle layer in ahp4 mutant versus WT plants (Fig. 6D-E). Taken together, these results clearly indicate that increased cuticle thickness in ahp4 mutant plants may help to prevent non-stomatal water loss from leaf and stem surfaces. To investigate the underlying genetics associated with increased cuticle thickness, transcript levels of several genes involved in wax biosynthesis, namely decrease wax biosynthesis (DEWAX), fatty acyl-coenzyme A reductase 6 (FAR6) and shine 1 (SHIN1), were compared in ahp4 mutant and WT plants (Cui et al., 2016). Results revealed that the expression of DEWAX was down-regulated in ahp4 mutant under dehydration conditions, while that of FAR6 and SHIN1 was up-regulated in ahp4 mutant plants under both non-dehydrated and dehydrated conditions (Fig. 6F). This difference in expression levels might contribute to increased cuticle thickness (Fig. 6D-E). These collective results suggest that AHP4 may play an important role in controlling wax biosynthesis in Arabidopsis plants acclimatizing to water stress.

A, Fourteen-day-old ahp4 and WT plants grown on germination medium plates were transferred to soil and grown under well-watered conditions for 21 additional days. Chlorophyll leaching of 35-day-old ahp4 and WT plants were measured at indicated time points. Data represent the means and standard errors (n = 5). B, TB staining patterns of leaves of 35-day-old ahp4 and WT plants grown on soil as described in (A). C, Wax surface ornamentation of stems of 35-day-old ahp4 and WT plants grown on soil as described in (A) were detected by scanning electron microscope. D, Cuticle of the fifth leaves (adaxial side) of 35-day-old ahp4 and WT plants grown on soil as described in (A) were also observed by transmission electron microscope. The green arrows indicate cuticular layer, and yellow arrows indicate cuticle proper. E, Cuticle thickness of the fifth leaves (adaxial side) of ahp4 and WT plants was measured by ImageJ software. Data represent the means and SEs (n = 5, where each repeat was counted from eight different areas). F, Expression of several wax-related genes in ahp4 and WT plants with or without 5 h of dehydration treatment as described in Fig. 4 Data represent the means and SEs (n = 3). Asterisks indicate significant differences between the two genotypes as determined by a Student’s t-test (*P < 0.05, **P < 0.01, ***P < 0.001). CW, cell wall; MC/WC, ahp4 well-watered control 0 h versus WT well-watered control 0 h; MD/WD, ahp4 dehydrated 5 h versus WT dehydrated 5 h. DEWAX, decrease wax biosynthesis; FAR6, fatty acyl-coenzyme A reductase 6; SHIN1, shine 1.
Decreased ROS Accumulation and Oxidative Damage, and Increased ROS-scavenging Antioxidant Capacity in ahp4 Plants
Plants respond to various environmental stresses, including drought, via many adaptive mechanisms, including maintaining a balance between ROS production and detoxification, which is important for the maintenance of many cellular functions (Wang et al., 2015; Choudhury et al., 2017; Farooq et al., 2019; Huang et al., 2019; Xie et al., 2019). As the microarray analyses showed that AHP4 is involved in regulating the expression of genes involved in ROS metabolism, we decided to further investigate the role of AHP4 in ROS homeostasis. We did so by examining ROS production (O2•− and H2O2) in the leaves of ahp4 mutant and WT plants under well-watered and water-deficit conditions using the staining approach (Fig. 7A-B). We observed a comparable accumulation of O2•− and H2O2 in the leaves of well-watered ahp4 and WT plants. In contrast, under water deficit conditions, the levels of O2•− and H2O2 in ahp4 mutant remained much lower than in WT plants (Fig. 7A-B). To determine if oxidative damage was in fact reduced in ahp4 mutant, the contents of malondialdehyde (MDA), a byproduct of lipid peroxidation and common marker of oxidative stress (Mostofa et al., 2015), in ahp4 and WT plants were compared. While no significant difference in MDA content was observed in ahp4 mutant and WT plants under well-watered conditions, after 13 days of drought stress ahp4 mutant displayed much lower MDA level than did the WT (Fig. 7C). These data suggest that loss-of-function of AHP4 mitigates oxidative damage in Arabidopsis plants by reducing drought-induced ROS accumulation and lipid peroxidation.

A, Nitro blue tetrazolium staining of superoxide; and B, diamino-benzidine staining of hydrogen peroxide of ahp4 and WT plants exposed to drought for 12 days. C, MDA content of ahp4 and WT plants under drought. Data represent the means and standard errors (SEs) (n = 3). D, Superoxide dismutase (SOD); E, ascorbate peroxidase (APX); F, glutathione peroxidase (GPX); and G, glutathione S-transferase (GST) enzyme activities in ahp4 and WT plants under soil-drying. Data represent the means and SEs (n = 3). Different superscripted letters (a, b, c, d, e and f) within the column reveal statistically significant differences between the two genotypes, and among the treatments, which were determined by Duncan’s multiple range test (P < 0.05). DAS, days after stress; WT-C, wild-type control; WT-D, wild-type drought; ahp4-C, ahp4 control; ahp4-D, ahp4 drought.
The results on drought-induced oxidative stress combined with the transcriptome data indicate that under the water-deficit conditions ahp4 mutant plants have a greater antioxidant capacity to more efficiently detoxify ROS, when compared with WT plants. To investigate this further, first the activities of several key ROS-scavenging enzymes like superoxide dismutase (SOD), ascorbate peroxidase (APX), glutathione peroxidase (GPX) and glutathione S-transferase (GST) were determined in WT and ahp4 plants under well-watered and water-deficit conditions (Fig. 7D-G). Under well-watered conditions, no differences were observed in the activities of APX, GPX and GST in the leaves of WT and ahp4 mutant plants, and only SOD activity showed higher activity in ahp4 mutants than in WT plants. In contrast, under water-deficit conditions the activities of the examined ROS-scavenging enzymes were generally greater in the leaves of ahp4 than in WT plants (Fig. 7D-G). Second, although there were no significant differences in anthocyanin contents between ahp4 mutant and WT plants under well-watered conditions (Fig. 8A-B), under water-deficit conditions ahp4 mutant plants possessed higher anthocyanin contents than WT plants (Fig. 8A-B). Concurrent with this observation, under water-deficit conditions, the expression of genes involved in anthocyanin/flavonoid biosynthesis showed a greater up-regulation in ahp4 mutant than in WT plants (Fig. 8C). These findings suggest that enhanced accumulation of anthocyanins, which may act as an antioxidant under different types of environmental stress, including drought (Nakabayashi et al., 2014; Lotkowska et al., 2015; Nguyen et al., 2016; Nguyen et al., 2016; Li et al., 2017), was associated with the drought-tolerant phenotype of ahp4 mutant plants. Collectively, these results suggested that loss-of function of AHP4 enhanced antioxidant defense as a preventive measure to protect ahp4 plants from drought-induced oxidative damage.

A, Representative of ahp4 and WT plants under control and 12 days after soil-drying. B, Anthocyanin contents in ahp4 and WT plants under control and drought conditions. C, Expression of anthocyanin/flavonoid-related genes in ahp4 and WT plants under control and drought conditions. Data represent the mean and standard errors (n = 5). Different superscripted letters (a, b, c, d, e, f and g) within the column indicate statistically significant differences between the two genotypes, and among the treatments determined by Duncan’s multiple range test (P < 0.05). DW, dry weight; DAS, days after stress; WT-C, wild-type control; WT-D, wild-type drought; ahp4-C, ahp4 control; ahp4-D, ahp4 drought; PAP1/PAP2, production of anthocyanin pigment 1/2; DFR, dihydroflavonol 4-reductase; F3H, flavanone 3-hydroxylase; F3’H, flavonoid 3’-monooxygenase; GL3, glabra 3.
DISCUSSION
Globally abiotic stresses, including drought, are the primary factors that impact plant survival and cause yield losses in crop plants (Lamaoui et al., 2018). To acclimatize to environmental stresses, plants need to be able to regulate a variety of developmental, physiological and cellular mechanisms to survive (Osakabe et al., 2014; Sah et al., 2016; Choudhury et al., 2017; Zandalinas et al., 2018). In this study, we found that the expression of AHP4 gene was down-regulated by dehydration or ABA treatment (Fig. 1B; Supplementary Fig. S9E), suggesting a possible involvement of APH4 in negative regulation of Arabidopsis adaptation to drought in ABA-dependent manner. This hypothesis is strongly supported by the fact that the aph4 mutant plants exhibited a drought-tolerant phenotype (Figs. 2 and 3; Supplementary Fig. S1A-D). Next, we used various physiological and biochemical assays and a genome-wide transcriptome analysis to investigate, identify and critically evaluate numerous AHP4-regulated genes, several developmental and physiological mechanisms, and selected pathways that might contribute to AHP4-mediated drought responses in Arabidopsis. Our results demonstrate that AHP4 does indeed play a negative regulatory role in a wide range of mechanisms, as summarized in Fig. 9.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Under drought stress, expression of AHP4 is repressed, weakening CK signaling. Downregulation of AHP4results in changes in various physiological and biochemical processes, including impairment of stomatal closure, enhanced photosynthetic efficiency, increased cell membrane integrity and cuticle thickness, and improvement of ROS detoxification through increasing both enzymatic and non-enzymatic antioxidants.
First, we investigated if loss-of-function of AHP4 would enhance primary root growth in plants exposed to concentrations of mannitol high enough to cause osmotic stress. Around 200 or 300 mM mannitol, ahp4 mutant plants showed increased primary root growth compared with WT plants (Supplementary Fig. S2). These results suggest that the enhancement of primary root growth observed in ahp4 mutant plants under osmotic stress treatment might contribute to ahp4 mutant’s drought-tolerant phenotype. Increasing downward root growth is an essential mechanism which can enable plants to absorb water from deep layers within the soil when water in shallower soil layers is limited (Feng et al., 2016). The negative regulatory role of AHP4, and thus CK signaling, in root growth and development observed in this study under osmotic stress, is supported by previous studies which reported significant increases in root length and biomass in both transgenic plants with reduced CK content (Werner et al., 2010) and CK-signaling ahk2,3 double mutant (Riefler et al., 2006; Li et al., 2019) even under normal growth conditions, and enhanced root elongation in ahk3 mutant plants exposed to low water potential (Kumar and Verslues, 2015). Additionally, the CK-deficient and ahk2,3 plants were better able to survive under drought (Tran et al., 2007; Werner et al., 2010; Nishiyama et al., 2011; Kang et al., 2012).
Second, the drought tolerance of ahp4 mutant plants was found be enhanced by their ability to retain higher RWC and Fv/Fm when compared with WT (Figs. 3C-D, 4A and 5A; Supplementary Fig. S1F) to better survive water deficit (Figs. 2 and 3A-B; Supplementary Fig. S1B, D). Maintaining higher RWC and better drought tolerance requires the down-regulation of APH4 in order to activate a range of biochemical and physiological mechanisms that help plants cope with water deficit as visualized in the model shown in Fig. 9. These mechanisms include increased maximum photochemical efficiency of PSII (Fig. 3), reduced stomatal apertures (Fig. 5D-E; Supplementary Fig. S8), decreased cell membrane damage and cuticle permeability (Fig. 6; Supplementary Fig. S1G), increased ABA responsiveness (Supplementary Fig. S9), and increased antioxidant defense (Figs. 7 and 8).
During drought, plants alter the photosynthetic machinery to maintain cell water status for better survival (Wang et al., 2018). For evaluation of drought tolerance of plants in terms of their ability of maintain photosynthesis, the Fv/Fm, stomatal conductance and transpiration rate parameters are widely used as photosynthetic indicators (Wang et al., 2018; Utsumi et al., 2019). Previous studies reported that the drought-tolerant plants have high Fv/Fm, NPQ, and low stomatal conductance and transpiration rate to maintain water status in plants during water scarcity (Wang et al., 2018; Utsumi et al., 2019). In this study, ahp4 plants showed greater Fv/Fm and NPQ under drought (Fig. 3C-D, G-H), higher leaf temperature during dehydration (Fig. 5B), lower stomatal conductance and transpiration rates under well-watered conditions (Supplementary Fig. S7A-B) when compared with those in WT, suggesting that the enhanced drought tolerance of ahp4 plants might be attributed to an alteration in photosynthetic responses. In addition, the observed greater photosynthetic efficiency of ahp4 plants (Fig. 3C-D) suggests that ahp4 plants may absorb light better than WT plants. In support of this premise, the transcript levels of the photosystem light harvesting complex (LHC)-related genes, such as LHCB2.1, 2.2, 2.3 and 4.1, were up-regulated in ahp4 plants when compared with WT plants under dehydration conditions (Supplementary Table S5). These findings indicate the important role of AHP4 in regulation of photosynthetic processes in plant response to drought.
ABA is known to be a key factor in plant abiotic stress responses as it is involved in regulating the expression of many ABA- and/or stress-inducible genes (Osakabe et al., 2014; Osakabe et al., 2014; Ding et al., 2015; Sah et al., 2016). Previous studies demonstrated that ABA and CK signaling pathways have antagonistic actions in various processes, including germination, senescence and drought tolerance (Nishiyama et al., 2011; Wang et al., 2011; Nguyen et al., 2016; Abdelrahman et al., 2017; Huang et al., 2018). In the present study, the enhancement of the ABA response in ahp4 mutant plants (Supplementary Fig. S9) could result in the up-regulation of downstream ABA- and/or stress-inducible genes under water deficit. These DEGs include the LEA, ABA insensitive 3 (ABI3), ABA-responsive element-binding factor 3 (ABF3) and β-glucosidase 1 (BG1) genes (Supplementary Fig. S3; Table S3A). These genes are known to have various important functions, including membrane protection, maintenance of osmotic homeostasis and repair of cell damage, in plants under water deficit, leading to improvement of the overall performance of plants exposed to drought (Fujita et al., 2005; Verslues et al., 2006). ABI3 encodes a AP2/ERF-type TF important for normal embryogenesis and seed development (Rohde et al., 2000; Roscoe et al., 2015), as well as for plants under dehydration stress (Bedi et al., 2016). ABI3 might have a positive regulatory function in Arabidopsis plants under drought as its transcription is up-regulated following dehydration, and its transcript levels are maintained during the subsequent stress recovery phases (Bedi et al., 2016; Bedi and Chaudhuri, 2018). On the other hand, the drought-induced ABF3 encodes a member of the ABF/AREB subfamily of bZIP-type TFs (Fujita et al., 2013; Yoshida et al., 2015; Zandkarimi et al., 2015; Wang et al., 2016). Overexpression/ectopic expression of AtABF3 in many plant species, including Arabidopsis, rice (Oryza sativa), grape (Vitis vinifera), lettuce (Lactuca sativa), peanut (Agrostis mongolica), alfalfa (Medicago sativa) and cotton (Gossypium hirsutum), results in increased tolerance of transgenic plants to various stresses, including dehydration, cold, high temperatures and oxidative stress (Kang et al., 2002; Kim et al., 2004; E. Vanjildorj, 2005; Oh et al., 2005; Abdeen et al., 2010; Pruthvi et al., 2014; Wang et al., 2016; Kerr et al., 2017). These reports together strengthen the idea that up-regulation of ABI3 and ABF3 genes by loss-of-function of AHP4 might contribute to enhanced drought tolerance of the ahp4 mutant plants. Additionally, in the present study, the AtBG1 gene, which encodes a β-glucosidase, was also expressed at higher levels in ahp4 mutant plants than WT plants under water deficit (Supplementary Table S3A). β-glucosidases release glucose from glucose-conjugates, such as the ABA-glucose ester by hydrolysis, releasing free ABA and increasing active ABA levels in plants (Lee et al., 2006; Dong et al., 2014). BG1 expression is highly induced in WT plants treated with exogenous ABA, NaCl and high concentrations of mannitol (Dong et al., 2014). Overexpression of AtBG1 in creeping bentgrass plants (Agrostis stolonifera) resulted in greater free ABA levels and increased drought tolerance in comparison with WT plants (Han et al., 2012). In contrast, atbg1 mutant plants were shown to be more sensitive to drought than WT (Lee et al., 2006). The above findings support a role for AHP4, as a negative regulator of AtBG1 expression, in the rapid ABA production pathway that is important for plants adapting to drought.
Under water deficit, plants commonly regulate water loss by controlling stomatal number and size of stomatal aperture (e.g. stomatal opening and closing) (Nilson and Assmann, 2007; Kerr et al., 2017; Saradadevi et al., 2017; Zandalinas et al., 2018). In the present study, a lower rate of water loss from ahp4 mutant plants, compared with WT plants under water deficit, was partly due to reduced stomatal aperture (Fig. 5D-E; Supplementary Fig. S8). Similar to ahp4 mutant, the ahp2 single, ahk2,3 double and arr1,10,12 triple mutant plants also showed narrower stomatal apertures than WT (Marchadier and Hetherington, 2014; Nguyen et al., 2016). Furthermore, ahk2,3 and arr1,10,12 mutants exhibited higher tolerance to drought than WT (Tran et al., 2007; Kang et al., 2012; Nguyen et al., 2016). Additionally, AHP4 was reported to interact with the type-B ARR1 in the CK signaling pathway (Dortay et al., 2006), and both AHP4 (this study) and ARR1 (Nguyen et al., 2016; Huang et al., 2018) act as negative regulators of drought tolerance. These findings collectively suggest that loss-of-function of AHP4 may result in weakened CK signaling, leading to reduced stomatal apertures and enhanced drought tolerance. Thus, CK signaling is an important negative regulator of stomatal activity with respect to plant adaptation to drought.
Cuticular wax is a protective barrier, containing predominantly long-chain hydrocarbons, which plays a crucial role in plant responses to various environmental stresses, including drought (Shepherd and Wynne Griffiths, 2006; Kosma et al., 2009; Seo et al., 2011; Zhu and Xiong, 2013; Lee et al., 2014). Changes in the cuticle formation, especially cuticle thickness, have been reported to be associated with drought tolerance in a range of plant species, such as Arabidopsis thaliana, rice and Camelina sativa (Aharoni et al., 2004; Shepherd and Wynne Griffiths, 2006; Seo et al., 2011; Zhu and Xiong, 2013; Lee et al., 2014; Zhu et al., 2014; Zhou et al., 2015). Adaptation of plants to drought often requires a reduction in both stomatal and non-stomatal water loss, and the latter can be achieved by increasing the thickness of the cuticles found on the surface of leaves and stems (Lee et al., 2014; Jetter and Riederer, 2016). Our results demonstrate that AHP4 negatively regulates the thickness of the cuticle layer as ahp4 mutant plants produced thicker cuticles (Fig. 6D-E); and thus, showed lower cuticle permeability than WT (Fig. 6A-B). In support of this finding, transcriptional analysis also revealed that AHP4 mediates plant drought responses by down-regulating wax biosynthesis-related genes, as indicated by increased expression of the SHN1 and FAR6 genes in ahp4 mutant plants under both well-watered and dehydration conditions (Fig. 6F) (Aharoni et al., 2004; Cui et al., 2016). On the other hand, expression of the DEWAX gene in ahp4 mutant plants was down-regulated under dehydration conditions (Fig. 6F). The DEWAX belonging to the AP2/ERF-type TF family was reported to act as a negative regulator of cuticular wax biosynthesis in Arabidopsis as indicated by a greater wax load in the dewax mutant and lower wax load in the DEWAX-overexpressing lines as compared with that in WT (Go et al., 2014; Suh and Go, 2014). DEWAX was also shown to negatively regulate the expression of several wax biosynthesis-related genes, including FAR6 (Go et al., 2014). As a result of enhanced wax biosynthesis, a thicker cuticle layer with reduced water permeability might help ahp4 mutant plants maintain higher RWC (Figs. 4A, 5A and 6A-E; Supplementary Fig. S1F) than WT plants under water deficit, which is likely to be one of the major factors contributing to the greater drought tolerance of ahp4 mutant plants.
In plants, the production of ROS can increase under drought conditions, leading to an imbalance in ROS-generation and -scavenging, which can result in oxidative damage (Mittler, 2017; Zandalinas et al., 2018; Huang et al., 2019). Therefore, a vibrant antioxidant defense is requisite to save plants from drought-induced oxidative stress (Miller et al., 2010; Zandalinas et al., 2018). A previous study reported that overproduction of endogenous CKs promoted ROS generation and decreased ROS-scavenging enzyme activities in Arabidopsis, resulting in enhanced salt sensitivity (Wang et al., 2015). In this study, lower ROS levels were found in ahp4 mutant plants than WT plants under water deficit (Fig. 7A-B), indicating that the mechanisms involved in ROS elimination might be activated in ahp4 mutant. In both well-watered and water-stressed ahp4 mutant plants, down-regulated expression of the ferric reduction oxidase 1 (FRO1) gene, which encodes a ferric-chelate reductase involved in production of O2•−, was observed (Supplementary Tables S2 and S3); possibly helping to maintain low O2•− concentrations (Fig. 7A) (Mittler et al., 2004). Results of enzymatic antioxidant assays confirmed that SOD, APX, GPX and GST exhibited enhanced activity levels under water deficit in ahp4 mutant plants when compared with WT plants (Fig. 7D-G). The enhanced SOD activity in the leaves of ahp4 may help in the elimination of excessive O2•− by enhancing the conversion of O2•− into H2O2 (Fig. 7A, D) (Liu and He, 2016). The reduced level of H2O2 in ahp4 mutant plants corresponded to the heightened activities of APX, GST and GPX, which are involved in the removal of H2O2 produced under unfavorable stress conditions (Fig. 7B, E-G). GPX and GST also play important roles in protecting the cellular membrane by removing organic peroxides and lipid hydroperoxides produced when plants undergo abiotic stresses (Miller et al., 2010; Mostofa et al., 2015). Therefore, enhanced activity of GPX and GST implies that these enzymes might significantly contribute to the protection of cellular membrane from drought-induced reactive peroxides. In addition, higher anthocyanin concentrations were found in ahp4 mutant than WT plants under drought (Fig. 8B). Since anthocyanins have been shown to function as non-enzymatic antioxidants (Nakabayashi et al., 2014; Li et al., 2017), this may also increase the capacity of ahp4 mutant plants to limit drought-induced ROS accumulation. The increase in anthocyanin levels in ahp4 mutant plants during drought may be explained by transcriptional regulation of the anthocyanin/flavonoid biosynthetic pathway as indicated by both the RT-qPCR and transcriptome data (Fig. 8C; Supplementary Figs. S4-S6). Several studies have found positive correlations between drought tolerance and anthocyanin levels in Arabidopsis owing to their ROS-scavenging functions (Nakabayashi et al., 2014; Nguyen et al., 2016; Li et al., 2017; Li et al., 2020a; Li et al., 2020b). The findings of the present study suggest that enhanced anthocyanin/flavonoid biosynthesis is an important drought acclimation mechanism in plants mediated by AHP4, and perhaps through ARR1, ARR10 and/or ARR12 as anthocyanin biosynthesis was reportedly enhanced in the triple arr1,10,12 mutant under drought (Nguyen et al., 2016).
CONCLUSIONS
Transcriptome analysis of ahp4 mutant and WT plants has provided valuable insight into the regulatory roles of AHP4, and thus CK signaling, in plant drought adaptation (Fig. 9). Loss-of-function of APH4 altered the expression of many genes associated with plant response to water deficit, including dehydration- and/or ABA-responsive genes important for drought tolerance. In comparison with WT plants, ahp4 mutant plants showed enhanced ABA responsiveness and photosynthetic efficiency, increased root elongation, reduced stomatal apertures, increased wax crystal accumulation and thicker cuticles. All of these factors contribute to increased water retention in ahp4 mutant under water deficit. Additionally, under drought, ahp4 mutant plants had lower ROS accumulation, less electrolyte leakage and lower lipid peroxidation levels. These changes indicate decreased levels of drought-induced cellular damage in ahp4 mutant plants, which were likely due to increased expression of genes encoding enzymatic (e.g. SOD, APX, GST and GPX) and non-enzymatic (e.g. anthocyanins) antioxidants participated in antioxidant defense. The aforementioned observations collectively indicate that AHP4 has the ability to negatively regulate multiple protective mechanisms associated with drought tolerance (Fig. 9). Therefore, AHP4 is a promising candidate gene to be identified in various crops for gene editing to generate drought-tolerant crop cultivars.
METHODS
Plant Materials, Growth and Treatments
The Arabidopsis ahp4 mutant used in this study is from the Columbia genetic background, and it was obtained from a previous work (Hutchison et al., 2006). For dehydration treatments, 21-day-old WT plants were grown on germination medium (GM) agar plates (22°C, 16-h light/8-h dark cycle, 60 µmol m−2 s−1 photon flux density) and exposed to dehydration for the indicated time periods. For ABA treatments, 21-day-old WT plants were grown on GM agar plates and treated in a solution containing 0 µM ABA (control) or 50 µM ABA for 0, 1, 2, 4 or 6 h. After the treatments, whole plant, shoot and root samples were collected in three biological replicates, and immediately frozen in liquid nitrogen and stored at - 80°C for further analyses.
Drought Tolerance Assay
For examining drought-tolerant phenotypes, we followed the published gravimetric method for biomass reduction assay (Harb and Pereira, 2011; Nguyen et al., 2018), and the survival test was done in the same-tray system (Nishiyama et al., 2011). Biomass reduction of aph4 and WT plants under drought stress relative to respective well-watered control plants was measured at 5 days after rewatering. Per genotype, 12 pots under either well-watered or drought conditions were used for measuring the plant biomass reduction. During soil-drying, the soil moisture content was recorded at five different positions per genotype using HydroSense (Campbell Scientific Australia Pty. Ltd, Australia). Photographs were taken 3 days after rewatering and removal of inflorescences from the surviving plants. For the survival test, the survival rates were calculated from three independent experiments, in which each replicate was calculated from 30 plants per genotype.
Osmotic Tolerance Assay
To examine root growth under osmotic stress, 7-day-old ahp4 mutant and WT plants were grown on GM plates, and then transferred onto 0.5 × Murashige and Skoog (MS) plates containing 1.2% agar and 0, 100, 200 or 300 mM mannitol. Primary root length of 14-day-old plants was measured after 7 days of incubation (22°C, 16-h light/ 8-h dark, 60 µmol m−2 s−1 photon flux density).
RWC and Electrolyte Leakage under Drought
The ahp4 mutant and WT plants were grown and subjected to drought stress as described in the survival test in the same-tray system (Nishiyama et al., 2011). RWC and electrolyte leakage of the detached aerial portions of the plants during soil-drying were measured at 11, 12, 13 and 14 days after drought, in accordance with the methods previously described (Nishiyama et al., 2011). Five biological replicates for each genotype were used for all experimental measurements.
ROS Accumulation and MDA Content
The ahp4 mutant and WT plants were grown and subjected to drought stress as described in the survival test in the same-tray system (Nishiyama et al., 2011). ROS accumulation of ahp4 mutant and WT plants, which were either exposed to 12 days of drought or well-watered control conditions, was determined by using nitro blue tetrazolium and diaminobenzidine staining, in accordance with the previous methods (Mostofa et al., 2015). MDA contents of ahp4 mutant and WT plants exposed to drought for 12 and 13 days or well-watered control conditions were measured using the previous procedure (Mostofa et al., 2015). Three biological replicates of each treatment were used for measuring MDA contents.
Anthocyanin Contents and Antioxidant Enzyme Activities
The ahp4 mutant and WT plants were grown and subjected to drought stress as described in the survival test using the same-tray system (Nishiyama et al., 2011). After 12 and 13 days of drought stress, rosette leaves of stressed and non-stressed plants were separately collected for determining anthocyanin contents and antioxidant enzyme activities. The anthocyanin contents of freeze-dried rosette leaves of stressed and non-stressed ahp4 mutant and WT plants were measured as previously described (Li et al., 2017). Antioxidant enzyme activities of rosette leaves of stressed and non-stressed plants were measured following previously described methods (Mostofa et al., 2015). Total soluble protein contents were measured using the Bradford method (Bradford, 1976).
RWC, Leaf Temperature, Stomatal Density and Aperture, Transpiration Rate and Stomata Conductance Measurements under Dehydration
To examine these characteristics, 32-day-old ahp4 mutant and WT plants grown on soil (22°C in light period/18°C in dark period, 12-h light/12-h dark cycle, 200 µmol m−2 s−1 photon flux density, 50% relative room humidity) were exposed to dehydration for the indicated time periods. The RWC of 32-day-old ahp4 mutant and WT plants were recorded during dehydration treatment according to the method previously described (Nishiyama et al., 2011). Five biological replicates for each genotype were used for RWC measurements. The relative leaf temperatures of 32-day-old ahp4 mutant and WT plants during dehydration treatment were detected using the FLIR ONE camera (FLIR Systems, Wilsonville, Oregon, USA) and Thermal Analysis software. The stomatal density and dehydration-induced stomatal closure of 32-day-old ahp4 mutant and WT plants were performed at 0 (control), 30 and 60 minutes of dehydration treatment. At the indicated time points, the stomata of the fifth leaf were embedded using Parkell Cinch hydrophilic vinyl polysiloxane (dental impression material) and Cartridge gun (Parkell Inc., Edgewood, New York, USA) for 1 h. The epidermal cells were peeled and then dried using Sally Hansen Double Duty (Sally Hansen, USA) for 1h. The epidermal cells were then used for determination of stomatal density and apertures using Leica DM750 microscope (Leica Microsystems Inc., Buffalo Grove, Illinois, USA) and ImageJ software (https://imagej.nih.gov/ij/). Five biological replicates for each genotype were used for stomatal measurements.
For transpiration rate and stomatal conductance measurements: for the dehydration samples, the whole rosette leaves of 32-day-old ahp4 mutant and WT plants were exposed to dehydration for 30 minutes, followed by 20 minutes of dark adaptation before measurement; for the well-watered samples, plants were kept for 20 minutes under dark before measurement. The constant systems of 2-cm chamber were set during the measurement (22°C of leaf temperature, 10,000 rpm fan speed, 500 μmol s−1 flow rate, 400 ppm CO2 level). The fifth rosette leaves of 32-day-old ahp4 mutant and WT plants were selected for determining the transpiration rate, stomata conductance at 30 minutes of dehydration or well-watered (control) conditions by using LICOR6800 system (LICOR Biosciences, Lincoln, Nebraska, USA). Three biological replicates for each genotype were used for all experimental measurements.
The Photosynthetic Efficiency, Chlorophyll Index and NPQ under Drought
The ahp4 mutant and WT plants were grown on soil under well-watered conditions (22°C in light period/18°C in dark period, 12-h light/12-h dark cycle, 200 µmol m−2 s−1 photon flux density, 50% relative room humidity) for 35 days, and then exposed to drought stress for 14 days. The plants were kept for 20 minutes under dark before measurement. The photosynthetic efficiency (Fv/Fm), chlorophyll index and NPQ of 49-day-old ahp4 mutant and WT plants under well-watered or drought conditions were measured using the CropReporter system (CID AgTech, Camas, Washington, USA). Four biological replicates for each genotype were used for all experimental measurements.
Germination and Root Inhibition Assays for Evaluation of ABA Responsiveness, and ABA-induced Stomatal Closure
Germination assay was conducted on GM medium containing 1% sucrose and various concentrations of exogenous ABA as previously described (Nishiyama et al., 2011). For root inhibition assay, 7-day-old ahp4 mutant and WT plants grown on GM plates were transferred onto 0.5 × Murashige and Skoog (MS) plates containing 1.2% agar and 0 or 20 μM ABA. Primary root growth was measured after 4 and 7 days of incubation (22°C, 16-h light/ 8-h dark, 60 µmol m−2 s−1 photon flux density). The ABA-induced stomatal closure was performed following the previous method (Osakabe et al., 2013). Fourteen-day-old ahp4 mutant and WT plants grown on GM plates were transferred to soil and grown for 7 additional days under well-watered conditions. Rosette leaves from 21-day-old plants were then used for determination of stomatal apertures under 0 (control), 30 and 50 μM ABA treatments (Osakabe et al., 2013).
Chlorophyll Leaching, TB Staining, and Determination of Epicuticular Wax Density and Cuticle Thickness
For chlorophyll leaching assay, the ahp4 mutant and WT plants were grown on soil under well-watered conditions (22°C in light period/18°C in dark period, 12-h light/12-h dark cycle, 200 µmol m−2 s−1 photon flux density, 50% relative room humidity) for 35 days. The chlorophyll leaching rates from rosette leaves of ahp4 and WT plants were determined as previously described (Li et al., 2017). TB staining was conducted following the published procedure (Tanaka et al., 2004). The aerial portions of 35-day-old plants grown on soil were submerged into a solution containing water or 0.05% (w/v) TB for 3 h. Treated aerial portions were subsequently transferred to water and gently shaken to remove excessive TB, and were then photographed using the Leica DM750 microscope. For the assay of epicuticular wax density in stems, 14-day-old ahp4 mutant and WT plants grown on GM plates were transferred to soil and grown for 21 additional days under well-watered conditions in the same-tray system (Nishiyama et al., 2011). The main stems of 35-day-old plants were then selected for measuring wax crystal surface by SEM (Ukitsu et al., 2007). The fifth rosette leaves of 35-day-old plants were also selected for determining cuticle layer thickness by TEM following previously reported procedures (Ukitsu et al., 2007). ImageJ software was used to measure the cuticle thickness, with eight different areas measured for each replicate.
Gene Expression Analyses
Total RNA was purified using the RNeasy Plant Mini Kit (Qiagen, Hilden, Germany). The cDNA synthesis and RT-qPCR were conducted according to previous methods (Le et al., 2011). UBQ10 was used as a reference gene for RT-qPCR data analysis. Gene-specific primers used for RT-qPCR are presented in Table S6.
Dehydration Sampling and Microarray Analysis
Fourteen-day-old ahp4 mutant and WT plants grown on GM plates were transferred to soil and grown for 10 additional days under well-watered conditions. The aerial portions of 24-day-old plants were then subjected to dehydration treatments as previously described (Ha et al., 2014). Rosette leaves of ahp4 mutant and WT plants treated by dehydration for 0, 2 and 5 h were collected in three biological repeats, and were then used for transcriptome analysis using the Arabidopsis Oligo 44K DNA microarray (Version 4.0, Agilent Technology) (Nishiyama et al., 2012). To search for DEGs, the criteria of |fold-change| ≥ 2 and a false discovery rate corrected P-value (q-value) of < 0.05 were used. The detailed protocol and raw microarray data have been deposited in the Gene Expression Omnibus database (GSE95614). ClustVis (https://biit.cs.ut.ee/clustvis/) and MapMan (http://mapman.gabipd.org) were used to analyze the data.
Statistical analysis
Analysis of variance (ANOVA) or Student’s t-test was used for data analysis. Different superscripted letters within the column reveal statistically significant differences between the two genotypes and among the treatments as determined by Duncan’s multiple range test (using IBM SPSS software package 21.0). Asterisks demonstrate significant differences as assessed by the Student’s t-test (*P < 0.05, **P < 0.01, ***P < 0.001).
AUTHOR CONTRIBUTIONS
L.-S.P.T. designed the research; C.V.H., K.H.N., M.G.M., C.D.T., Y.W., W.L., Y.O., M.S., K.T., M.T., C.A., R.Z. and M.S. performed the research; K.T., and M.S. contributed research materials, reagents and analytic tools; C.V.H., M.G.M., C.A. and R.Z. analyzed the data with the input of D.J.B. and L.-S.P.T.; and C.V.H., D.J.B. and L.-S.P.T. critically interpreted the study and wrote the manuscript.
ACKNOWLEDGMENTS
The authors would like to thank Dr. Sona Pandey, Donald Danforth Plant Science Center, Rie Nishiyama and Yu Li for their support in this research, Mayumi Wakazaki, RIKEN CSRS, for supporting the TEM observations, and Audrey Dodds for editing the English.
Footnotes
Data deposition: The data reported in this paper have been deposited in the Gene Expression Omnibus (GEO) database, www.ncbi.nlm.nih.gov/geo (accession no. GSE95614). ID:chienhavan. Password: Hana18289.
REFERENCES




