

Abstract
Only two de novo biosynthetic routes to nicotinamide adenine dinucleotide (NAD+) have been described, both of which start from a proteinogenic amino acid and are tightly controlled. Here we establish a C3N pathway starting from chorismate in Escherichia coli as a third NAD+ de novo biosynthesis pathway. Significantly, the C3N pathway yielded extremely high cellular concentrations of NAD(H) in E. coli. Its utility in cofactor engineering was demonstrated by introducing the four-gene C3N module to cell factories to achieve higher production of 2,5-dimethylpyrazine and develop an efficient C3N-based whole-cell bioconversion system for preparing chiral amines. The wide distribution and abundance of chorismate in most kingdoms of life implies a general utility of the C3N pathway for modulating cellular levels of NAD(H) in versatile organisms.
Introduction
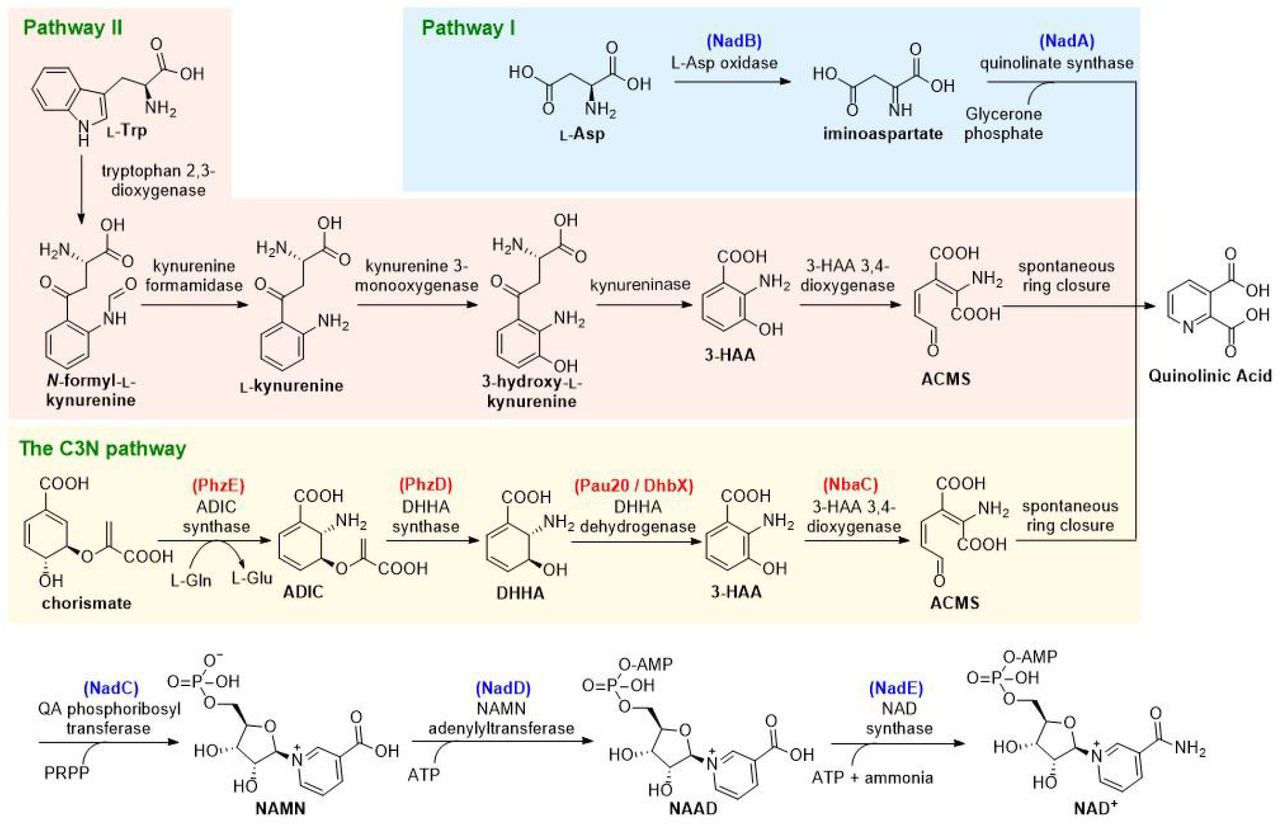
Nicotinamide adenine dinucleotide (NAD+) is a universally essential central metabolite consisting of adenosine monophosphate (AMP) linked to nicotinamide mononucleotide (NMN). Since its discovery as the first ‘cozymase’ over 100 years ago, two de novo biosynthesis pathways for NAD+ have been reported (Fig. 1)1. Both pathways use a proteinogenic amino acid as the precursor for nicotinamide and converge at a common intermediate quinolinic acid (QA). In plants and most bacteria, the de novo biosynthesis of NAD+ starts from L-aspartate (L-Asp), which is converted to QA by L-Asp oxidase (NadB) and quinolinate synthase (NadA) (Pathway I); in mammals, fungi, and some bacteria, nicotinamide is derived from conversion of L-tryptophan (L-Trp) to 3-hydroxyanthranilic acid (3-HAA) by four reactions of the kynurenine pathway (Pathway II). Oxidation of 3-HAA by 3-HAA 3,4-dioxygenase yields 2-amino-3-carboxymuconate semialdehyde (ACMS), which undergoes spontaneous ring closure to form QA1–3. The QA generated in both pathways is converted to NAD+ via a common three-enzyme pathway (catalyzed by enzymes NadC-E): phosphoribosylation of QA to generate nicotinic acid mononucleotide, AMP addition to form nicotinic acid adenine dinucleotide, and a final amidation to afford NAD+. In addition to the two de novo biosynthesis pathways, NAD+ is also regenerated by different salvage pathways using varied pyridines (nicotinic acid, nicotinamide, and nicotinamide riboside) as precursors1, 4.

The enzymes used in this work are in parenthesis and color-coded as either red (from secondary metabolic processes like natural product biosynthesis and aromatic compound degradation) or blue (from NAD+ biosynthesis). ADIC: 2-amino-2-deoxyisochorismate; DHHA: 2,3-dihydro-3-hydroxyanthranilic acid; ACMS: 2-amino-3-carboxymuconate semialdehyde; 3-HAA: 3-hydroxyanthranilic acid; PRPP: 5-phosphoribosyl diphosphate; NAMN: nicotinic acid mononucleotide; NAAD: nicotinic acid adenine dinucleotide.
NAD+ and its reduced form NADH serve as hydride acceptor and donor, respectively, in thousands of redox reactions in vital metabolic pathways such as glycolysis, citric acid cycle, fatty acid degradation, and oxidative phosphorylation5. Although cellular levels of NAD(H) are usually sufficient to support NAD(H)-dependent oxidoreductases under physiological conditions, NAD(H) concentrations may become rate-limiting when these enzymes are overproduced in engineered cell factories6. Indeed, many such examples of limiting NAD(H) have been documented since the first report of Escherichia coli cells overexpressing Vibrio harveyi luciferase in 19827, 8. NAD(H) deficiency is also medically significant; for example carcinoid syndrome patients, in whom L-Trp is depleted by excessive production of serotonin, exhibit pellagra-like symptoms caused by NAD(H) deficiency9.
NAD+ has also been recently identified as the substrate for NAD+-consuming enzymes involved in signal transduction pathways that regulate crucial biological processes including DNA repair, transcription, and cell cycle progression10–12. In some cases, cellular levels of NAD(H) are reduced dramatically by NAD+-consuming enzymes; for example, the hyper-activation of poly(ADP-ribose) polymerase-1 reduced the NAD+ pool by about 80%11. Consequently, increasing cellular NAD(H) concentrations can stimulate the NAD+-consuming enzymes and modulate downstream biological processes. For example, increasing NAD(H) levels can activate sirtuins and improve mitochondrial homeostasis and extend the lifespan of different species13, 14.
Several strategies have been developed to enhance the catalytic efficiencies of NAD(H)-dependent or NAD+-consuming enzymes by increasing cellular NAD(H) levels. These include supplementation with NAD+ or its pyridine precursors, limiting NAD+ consumption, and reinforcing NAD+ salvage or de novo biosynthesis pathways15–17. However, each strategy has its limitations: supplementation is restricted by poor cellular uptake of NAD+; reducing NAD+ consumption and accelerating its recycling via the salvage pathways can only replenish but cannot expand cellular NAD(H) pools. Considering that the highest cellular NAD(H) levels are limited in large part by NAD+ de novo biosynthetic capacity, manipulating these de novo pathways should efficiently expand cellular NAD(H) pools. The most frequently used strategy for increasing cellular NAD(H) levels is by reinforcing the NAD+ salvage pathways instead of manipulating de novo biosynthesis. However, the few reported attempts to manipulate NAD+ de novo pathways yielded only very limited18 or nonexistent increases in cellular NAD(H) levels19, probably due to the stringent regulation of NAD+ de novo biosynthesis at transcriptional20, translational21, and post-translational levels22. In addition, because both NAD+ de novo biosynthesis pathways start from a proteinogenic amino acid, their activation may deplete amino acid pools required to efficiently overproduce proteins in engineered cell factories.
In this study, we decoupled NAD+ de novo biosynthesis and protein synthesis by designing a C3N pathway as the third NAD+ de novo biosynthesis pathway, which uses chorismate as the precursor for nicotinamide. The C3N pathway was constructed in E. coli by combining genes from secondary metabolism with the latter steps of pathway I. It effectively circumvents the tight regulatory controls on NAD+ de novo biosynthesis and enables extremely high cellular concentrations of NAD(H) in the recombinant E. coli strains. We genetically packaged the C3N pathway as a four-gene C3N module for plug-and-play installation to significantly boost cellular NAD(H) concentrations in E. coli. Its utility in cofactor engineering was demonstrated by improving the bioconversion efficiency of 2,5-dimethylpyrazine (DMP) and by developing a C3N-based whole-cell system for efficient production of chiral amines.
Results and Discussion
Conceptual design of a third NAD+de novo biosynthesis pathway
The conceptual basis for an alternative NAD+ de novo biosynthesis pathway arose from the observation that several secondary metabolites contain structures derived from 3-HAA23–28 (Supplementary Fig. 1a), which also occurs in primary metabolism as a key intermediate in NAD+ de novo biosynthesis pathway II (Fig. 1). The biosynthetic gene clusters encoding these natural products indicate that chorismate is converted to 3-HAA by three sequential reactions catalyzed by 2-amino-2-deoxyisochorismate (ADIC) synthase, 2,3-dihydro-3-hydroxyanthranilic acid (DHHA) synthase, and DHHA dehydrogenase (Fig. 1 and Supplementary Fig. 1). Chorismate is an ideal precursor for a new pathway because as a natural branch point for many primary and secondary metabolic processes it is already metabolically promiscuous and abundant in most cells29, 30. We therefore designed a third de novo biosynthesis pathway to NAD+ by combining the chorismate-to-3-HAA pathway with 3-HAA 3,4-dioxygenase (the enzyme converting 3-HAA to QA) and the common three-step process of pathways I and II converting QA to NAD+. This synthetic route was designated as C3N pathway based on its precursor chorismate, the key intermediate 3-HAA and the final product NAD+, and the four enzymes converting chorismate to QA comprised the C3N module.
Stringent regulatory control of NAD+ de novo biosynthesis is directed mainly at the first biosynthetic genes like nadB and occurs at the transcriptional, translational, and post-translational levels20–22. By constructing the C3N pathway using defined promoters, ribosomal binding sites, and enzymes from secondary metabolism, these regulatory controls may theoretically be circumvented. Moreover, the recruitment of chorismate as the precursor for nicotinamide should decouple the C3N pathway from protein synthesis and make it suitable for use in engineered cells that overexpress proteins.
Characterization of Pau20 as a DHHA dehydrogenase
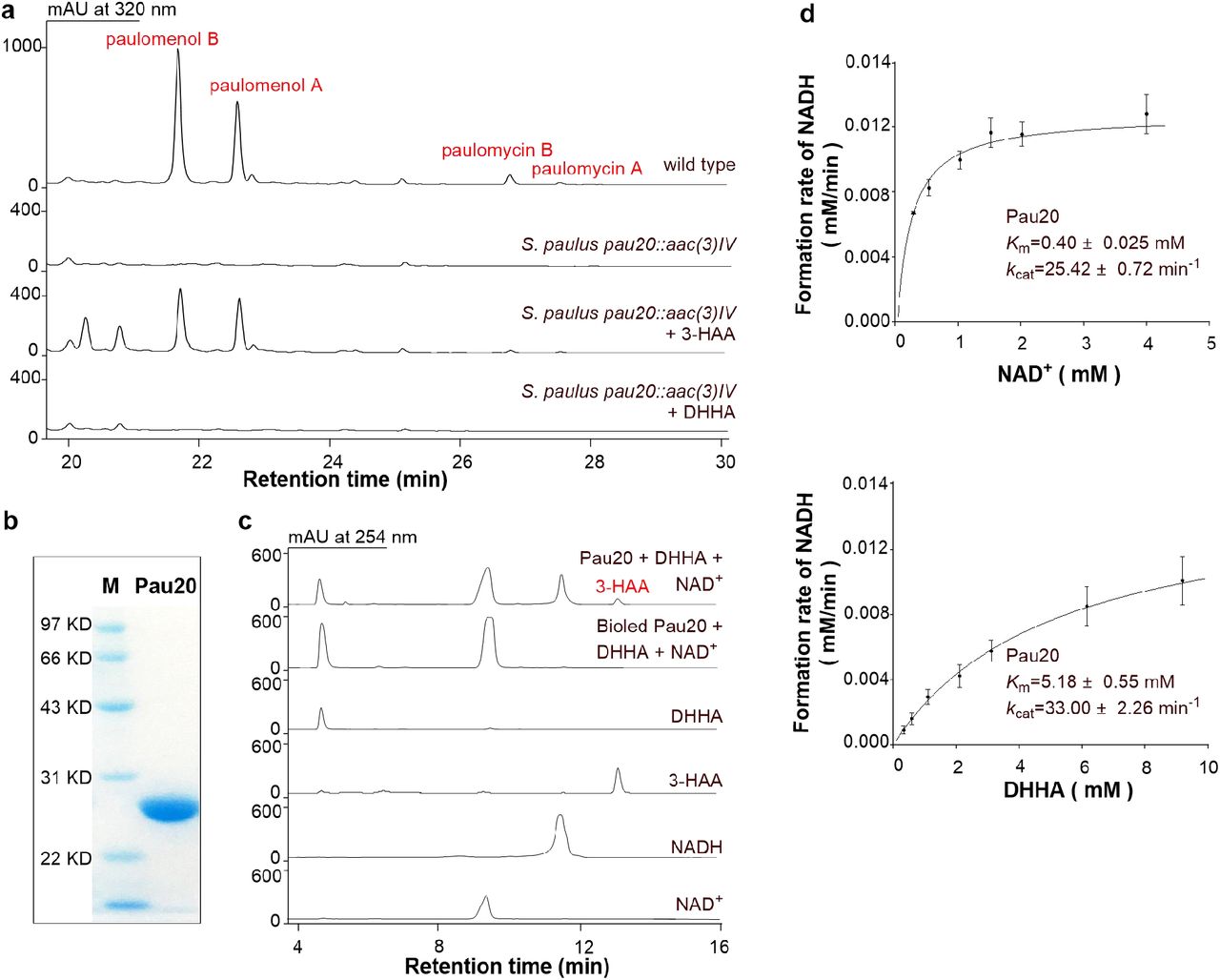
Although the ADIC synthase and DHHA synthase enzymes catalyzing the first two steps of the C3N pathway have been well studied31, the third enzyme DHHA dehydrogenase has yet to be biochemically characterized (Supplementary Fig. 1). We chose the putative DHHA dehydrogenase Pau20 from the paulomycin biosynthetic gene cluster for characterization27. The pau20 gene was inactivated by gene replacement in Streptomyces paulus NRRL 8115 to construct the S. paulus pau20∷aac(3)IV mutant (Supplementary Fig. 2a), in which the production of paulomycins was totally abolished. In feeding experiments of S. paulus pau20∷aac(3)IV, the production of paulomycins could be restored by 3-HAA but not by DHHA, indicating that Pau20 is responsible for the conversion of DHHA to 3-HAA (Fig. 2a). N-His6-tagged Pau20 was then overexpressed in E. coli BL21, purified, and incubated with DHHA and NAD+. Efficient oxidation of DHHA to 3-HAA was observed, verifying Pau20 as a DHHA dehydrogenase (Fig. 2b and 2c).

a, HPLC metabolite profiles of S. paulus wild type, the pau20 inactivated mutant S. paulus pau20∷aac(3)IV, and S. paulus pau20∷aac(3)IV complemented with 3-HAA or DHHA. b, SDS-PAGE analysis of Pau20. M, protein marker; Pau20, N-His6-tagged Pau20. c, Representative enzymatic assays of Pau20. d, Steady-state kinetic analysis of Pau20.
Previous studies showed that 3-HAA is prone to oxidation under alkaline conditions and exposure to air32. Indeed, when the Pau20 assay was performed at buffers over pH 7.2, spontaneous oxidation of 3-HAA could be observed (Supplementary Fig. 3a). Therefore, Pau20 characterization was first performed at pH 7.0 and its optimal temperature of 37 °C (Supplementary Fig. 3b). Steady-state kinetic analysis of Pau20 under these conditions revealed Michaelis-Menten behavior for all substrates and kinetic constants consistent with Pau20 acting as an efficient DHHA dehydrogenase (Fig. 2d and Supplementary Fig. 3f). Characterization Pau20 in vitro not only supported a useful DHHA dehydrogenase for testing the designed C3N pathway, but for the first time unambiguously verified the chorismate to 3-HAA pathway originally proposed for a number of natural products.
Constructing the C3N pathway in E. coli
The feasibility of the C3N pathway was initially tested by enabling 3-HAA production in E. coli, which lacks the kynurenine pathway and therefore cannot synthesize 3-HAA. A pau20-phzDE cassette consisting of pau20 and two genes encoding the well-characterized ADIC synthase (PhzE) and DHHA synthase (PhzD) from the phenazine biosynthesis pathway of Pseudomonas aeruginosa PAO133, 34 was synthesized and inserted into the medium-copy-number plasmid pXB1s downstream of the arabinose-inducible promoter PBAD to construct pXB1s-HAA. This plasmid was introduced into E. coli BW25113 to generate E. coli BW-pXB1s-HAA, which produced 3-HAA at a titer of 9.91 ± 0.14 mg/L (Fig. 3a).

a, HPLC analysis of the metabolite profiles of E. coli BW25113 and BW-pXB1s-HAA with or without Ara (arabinose, 10mM). b, Growth of E. coli BW25113 (wild type) and ΔnadAB on M9 plates with or without QA (10 mM) addition. c, Growth of E. coli ΔnadAB-pXB1s-QA and the control strain E. coli ΔnadAB-pXB1s on M9 plates with or without arabinose (10 mM) addition. d, The cellular NAD(H) (NAD+ and NADH total) and NADH concentrations of different E. coli strains.
To establish the complete C3N pathway in E. coli, the nbaC gene encoding an efficient 3-HAA 3,4-dioxygenase from the aromatic compound degradation pathway of Pseudomonas fluorescens KU-735 was inserted into plasmid pXB1s-HAA to generate pXB1s-QA, in which the four-gene cassette nbaC-pau20-phzDE dictates a C3N module converting chorismate to QA. In addition, to test whether cells could survive with the synthetic C3N pathway as the sole de novo source of NAD+, we constructed E. coli ΔnadAB (Supplementary Fig. 2b), a null mutant of NAD+ de novo biosynthesis, which was unable to grow on M9 plate unless QA was supplemented (Fig. 3b). Indeed, growth could be restored by introducing pXB1s-QA into E. coli ΔnadAB to afford E. coli ΔnadAB-pXB1s-QA, a recombinant strain with the complete C3N pathway. It grew well on M9 plates when the inducer arabinose was added. As a control, E. coli ΔnadAB with empty pXB1s could not grow on M9 plates even when supplemented with arabinose (Fig. 3c). Cellular NAD(H) levels measured at early stationary phase in liquid M9 medium with 10 mM arabinose were slightly higher in the C3N strain E. coli ΔnadAB-pXB1s-QA (1.18 ± 0.18 mM) than in E. coli BW25113 in M9 medium with (0.85 ± 0.05 mM) or without arabinose (0.89 ± 0.04 mM) (Fig. 3d). These results indicate that the C3N pathway was active in E. coli ΔnadAB-pXB1s-QA and could supply NAD+ efficiently for its growth.
Optimizing the C3N pathway in E. coli
To assess the potential of the C3N pathway for improving cellular NAD(H) levels, we cloned the nbaC-pau20-phzDE cassette into the high-copy-number vector pAB1s. Transformation of the resultant plasmid into E. coli ΔnadAB afforded the strain E. coli ΔnadAB-pAB1s-QA with cellular NAD(H) concentrations as high as 4.43 ± 0.14 mM. HPLC analysis of the E. coli ΔnadAB-pAB1s-QA metabolic profile revealed that DHHA was accumulated in the fermentation broth (Supplementary Fig. 3c), implying that NAD(H) levels could be further increased by using more efficient DHHA dehydrogenases.
Six putative DHHA dehydrogenases (NatDB, CbxG, BomO, DhbX, CalB3, and StnN) were overproduced and purified as N-His6-tagged proteins and assayed along with Pau20 at pH 7.0 and 37 °C. All six enzymes displayed DHHA dehydrogenase activities, with DhbX, CalB3, and StnN outperforming the other three (Supplementary Fig. 3d-f). Steady-state kinetic analysis revealed that DhbX had the highest specificity constants among all tested DHHA dehydrogenases (Supplementary Fig. 3f). To exploit the efficient DhbX enzyme, we constructed plasmid pAB1s-QA* by synthesizing the nbaC-dhbX-phzDE cassette and cloning it into pAB1s. Introduction of pAB1s-QA* into E. coli ΔnadAB generated E. coli ΔnadAB-pAB1s-QA*, in which the accumulation of DHHA was clearly reduced (~35%) and its cellular NAD(H) concentration increased to 7.21 ± 0.53 mM (Fig. 3d and Supplementary Fig. 3c). When E. coli ΔnadAB-pAB1s-QA* was cultured in 250 mL flasks that contained 50 mL medium, the cellular NAD(H) concentration reached 9.28 ± 0.76 mM (Fig. 3d). Under the same cultivation conditions, the cellular NAD(H) levels of E. coli BW25113 and the control strain E. coli BW-pAB1s (E. coli BW25113 with empty pAB1s) were 0.96 ± 0.13 mM and 0.65 ± 0.09 mM, respectively.
The 9.7-fold increase in cellular NAD(H) concentration generated solely by the C3N pathway compares favorably with previous cofactor engineering efforts. Specifically, cellular NAD(H) levels could be enhanced 7-fold (to 7.03 mM) in E. coli BL21 by reinforcing NAD+ salvage via overexpression of the nicotinic acid phosphoribosyltransferase PncB and NAD+ synthetase NadE15. Other attempts to either limit NAD+ consumption or reinforce NAD+ de novo pathways achieved no more than 4-fold increases to cellular NAD(H) levels17, 18. Interestingly, an upper limit to cellular NAD(H) concentrations was proposed for the E. coli BW25113-derived strain YJE003. By inactivating nadE to block NAD+ synthesis and expressing the NAD(H) transporter ntt4 gene to enable NAD+ intake, the maximum NAD(H) level of YJE003 was 9.6-fold (8.5 mM) higher than BW2511336. Similarly, our C3N-generated cellular NAD(H) concentration for E. coli ΔnadAB-pAB1s-QA* was 9.7-fold (9.3 mM) higher than BW25113, suggesting that the C3N pathway is optimized for NAD(H) yield and therefore harbors great potential for more general applications that require expanded cellular NAD(H) pools.
Increasing DMP production via the C3N module
We envisioned that the C3N pathway converting chorismate to QA may represent a powerful tool to increase cellular NAD(H) levels. Specifically, we can consider the advantages of a packaged ‘C3N module’: (i) it can be easily introduced into targeted strains as a four-gene cassette; (ii) the cellular NAD(H) pools can be expanded significantly by the established C3N pathway in E. coli; and (iii) it is compatible with other NAD(H) level increasing strategies (Fig. 4a). We first tested this by introducing pAB1s-QA* into E. coli DMP, a cell factory for producing the food flavor additive 2,5-dimethylpyrazine (DMP), which is also the synthetic precursor of 5-methyl-2-pyrazinecarboxylic acid, a key component of widely used pharmaceuticals like Glipizide and Acipimox37. E. coli DMP overexpresses two genes encoding Tdh and SpNox. Tdh is an NAD+-dependent L-threonine-3-dehydrogenase that oxidizes L-threonine to (2S)-2-amino-3-oxobutanoate, which then spontaneously undergoes decarboxylation and dimerization to yield DMP38; SpNox is an NADH oxidase (H2O forming) for NAD+ recycling39. Compared with E. coli DMP-Con (E. coli DMP with empty pAB1s), the NAD(H) levels of the new strain E. coli C3N-DMP increased 2.7-fold, and DMP production was 3.8-fold higher (Fig. 4b). These results supported the application of the C3N module as a simple and effective tool to increase cellular NAD(H) levels and boost yields in microbial cell factories.

a, General overview of improving the bioconversion efficiencies of cell factories by C3N module introduction. b, Improvement of DMP production by C3N module introduction. c, Evaluation of different C3N-based E. coli whole-cell bioamination systems. d, Evaluation of the bioamination efficiencies of E. coli C3N-ChA3 using different rac-alcohols as substrates. The bioconversions were performed in 100 mM KPi buffer (pH 8.5) with 10% DMSO, 4 M NH4COOH, OD600 of 50, at 37 °C, 230 rpm for 10 h.
Construction of a C3N-based whole-cell system for preparing chiral amines
Encouraged by the convenience and efficacy of the C3N module towards boosting DMP production, we next sought to develop a C3N-based whole-cell system for preparing chiral amines. Chiral amines are important for synthetic intermediates of pharmaceuticals and other bioactive molecules40. Preparative scale amination of alcohols to enantiomerically pure chiral amines has been achieved using a biocatalytic hydrogen-borrowing cascade employing an NAD+-dependent alcohol dehydrogenase (ADH) coupled with an NADH-dependent amine dehydrogenase (AmDH) to enable efficient internal recycling of the nicotinamide coenzyme40 (Fig. 4c). A recent report described the first whole-cell conversion of alcohols to chiral amines by combining an enantioselective AA-ADH with a chimeric amine dehydrogenase Ch1-AmDH. The bioamination efficiencies in those whole-cell systems were enhanced by increasing cellular NAD(H) levels via the supplementation of NAD+ to lyophilized E. coli resting cells (external NAD+ cannot diffuse into cells without lyophilization) or by adding glucose to promote NADH recycling. This system could transform the starting alcohol (S)-1a to the chiral amine product (R)-1c (amphetamine) with 46 ± 14% yield (Fig. 4c). Adding the R-selective LBv-ADH enabled conversion of the racemic alcohol substrate rac-1a to product (R)-1c with 21 ± 6% yield41.
The critical role of the expensive reagent NAD(H) in this dual-enzyme system suggested it could benefit from C3N-boosted NAD(H) levels, and inspired us to develop an efficient whole-cell system for preparing chiral amines. We chose TeSADH, an ADH capable of accepting racemic secondary alcohols42, and CalAmDH, an AmDH with high activities toward phenylacetones43, to construct the dual-enzyme cascade. The medium copy number plasmid pCDF-TesADH-CalAmDH was transformed into E. coli BW25113(DE3) recombinant strains BW-C3N (harboring pAB1s-QA*) and BW-Con (harboring empty pAB1s) to afford E. coli C3N-ChA1 and the control strain ChA1-Con, respectively. The cellular NAD(H) concentration of E. coli C3N-ChA1 (4.92 ± 0.50 mM) was much higher than that of E. coli ChA1-Con (1.52 ± 0.03 mM) (Fig. 4c). With respect to increased production, the rac-1a substrate was turned over to product (R)-1c (ee > 99%) with 5.6-fold higher conversion for E. coli C3N-ChA1 (9.5 ± 0.4%) relative to E. coli ChA1-Con (1.7 ± 0.4%) (Fig. 4c).
Although incorporating the C3N module led to 5.6-fold higher conversion to the final product, further optimization was required to increase the bioamination efficiency of the whole-cell system. We therefore optimized the whole-cell bioconversion procedure for E. coli C3N-ChA1 with rac-1a (Supplementary Fig. 4a-d) and the selected optimal condition (100 mM KPi buffer, pH 8.5; 4 M NH4COOH; 5 mM rac-1a; OD600 = 50; 37 °C, 230 rpm for 10 h) doubled conversion to 20.6 ± 2.0 % of product (R)-1c. Further improvements were realized by additional gene copies to increase protein levels of the ADH-AmDH catalysis module. First, introduction of pACYC-CalAmDH, an expression plasmid of CalAmDH, into E. coli C3N-ChA1 afforded E. coli C3N-ChA2 with a similar conversion to (R)-1c of 21.5 ± 0.7 %. Next, the high copy number plasmid pRSF-TesADH-CalAmDH was transformed into E. coli BW-C3N to generate E. coli C3N-ChA3, which doubled conversion to 41.4 ± 4.2 % of product (R)-1c. Finally, E. coli C3N-ChA4 was obtained by transforming pACYC-CalAmDH into E. coli C3N-ChA3. It exhibited comparable bioamination capacity to E. coli C3N-ChA3 (Fig. 4c and Supplementary Fig. 4e).
Having constructed a set of strains with variable bioamination efficiencies, we sought to assess whether these efficiencies could be linked to C3N-generated NAD(H). Indeed, NAD(H) measurements revealed a clear positive correlation between bioamination efficiency and cellular NAD(H) levels of the resting cells (Fig. 4c). To further verify the effect of the C3N pathway on bioamination, a second control strain E. coli ChA3-Con was constructed by transforming pRSF-TesADH-CalAmDH into BW-Con. Under the optimized conditions the cellular NAD(H) levels of E. coli ChA3-Con (3.60 ± 0.47 mM) were much lower than that of E. coli C3N-ChA3 (7.03 ± 1.57 mM), and the conversion to final amine product was halved (18.7 ± 0.5%, (R)-1c) (Fig. 4c). When the concentration of rac-1a was increased to 20 mM, the conversion to product (R)-1c was much lower for E. coli ChA3-Con (6.7 ± 1.3%) than that for E. coli C3N-ChA3 (48.3 ± 4.1 %, Fig. 4d), further highlighting the important contribution of the C3N pathway.
We next explored the substrate promiscuity of the system using a series of phenyl-containing secondary alcohols (rac-2a to rac-5a). In all cases amination by E. coli C3N-ChA3 achieved remarkable amine conversions (40-50%) and optical purity (ee > 99% (R)), (Fig. 4d). To demonstrate the applicability of this whole-cell bioamination system at larger scale, preparative biotransformation of rac-2a (138 mg, 0.9 mmol) was carried out in a 30 mL mixture of E. coli C3N-ChA3 resting cells and resulted in 38.1 ± 1.9 % conversion to the product (R)-2c. When scaled up to 100 mL (using 462 mg rac-2a, 3 mmol), about 30% conversion was observed (producing 138 mg (R)-2c).
The C3N-based whole-cell system for chiral amine preparation revealed that the C3N module can be used as a powerful and expedient tool for cofactor engineering in E. coli. Moreover, we believe it has the potential to be used in a versatile range of host organisms because: (i) chorismate, the precursor of the C3N pathway, is a product of the shikimate pathway, which exists widely in bacteria, archaea, fungi, and plants44; (ii) the shared final three steps of NAD+ de novo biosynthesis pathways I and II exist in almost all organisms. In most cases, the only requirement for constructing the C3N pathway is to express the C3N module genes in the targeted organisms.
Author Contributions
Conceptualization, Y. D., X. L., and Y. C.; Methodology, Y. D., X. L., B. W., and Y. C.; Investigation, Y. D., X. L., P. L., M. W., J. L., Z. Z., and W. L.; Writing – Original Draft, Y. D., X. L., G. P. H., and Y. C.; Writing –Review & Editing, Y. D., G. P. H., B. W., Y. T., and Y. C.; Funding Acquisition, Y. C.; Resources, B. W., Y. T., and Y. C.; Supervision, Y. C.
Competing interests
The authors declare no competing financial interest.
Methods
Strains and plasmids
E. coli JM109 was used for DNA cloning. E. coli BL21(DE3) was used for expressing recombinant proteins. E. coli BW25113 and BW25113(DE3) were used for constructing C3N pathway and the C3N pathway-based whole-cell systems for preparing chiral amines, respectively. LB, M9 and M9Y media (M9 with 1 g/L yeast extract) were used for growth of E. coli strains at 37 °C except where noted specifically45. S. paulus NRRL 8115 and its mutants were grown on mannitol/soya (MS) agar for spore collections46. GS-7 medium was used as the seed culture and R5α medium was used for paulomycin production47, 48. All Streptomyces strains were cultured at 28 °C, 220 rpm. Strains and plasmids used in this study are summarized in the Supplementary Table 1.
General DNA manipulations and sequence analyses
DNA synthesis and sequencing were performed in Generay Biotech (Shanghai, China) and Biosune (Shanghai, China) respectively. PCRs were performed with KOD-Plus DNA polymerase (Toyobo, Osaka, Japan) or Q5 DNA polymerase (NEB, Beijing). The one-step cloning method was employed in plasmid construction according to the manufacturer’s instructions (Vazyme, Nanjing, China). Restriction enzyme digestions, ligations and transformations were carried out following general methods45. E. coli-Streptomyces conjugations were performed as described46. A BLASTP search was used to find protein homologs (https://blast.ncbi.nlm.nih.gov/Blast.cgi). Protein sequence alignments were performed using Clustal Omega (https://www.ebi.ac.uk/Tools/msa/clustalo/).
Construction of S. paulus pau20∷aac(3)IV
A blue-white screening-based gene inactivation system was used to inactivate gene pau2049. The 1.9-kb upstream fragment and the 2.2-kb downstream fragment of pau20 were amplified using primer pairs pau20-s2/pau18R and pau22-R/pau20-R and inserted into the MunI and BlnI sites of pCIMt002 respectively to afford the pau20 disruption plasmid, which was then introduced into S. paulus NRRL 8115 via E. coli-Streptomyces conjugation. Both blue and white exconjugants were obtained on the plate. One of the white exconjugants with apramycin resistance was selected as the desired pau20 gene inactivation mutants S. paulus pau20∷aac(3)IV, which was verified by PCR with primer pair pau20-s2/ pau20-R (Supplementary Fig. 2a).
Production of paulomycins
For paulomycin production, 50 μL spores of S. paulus NRRL 8115 or pau20∷aac(3)IV were inoculated into GS-7 liquid medium and cultured at 28 °C, 220 rpm for 2 days. The resulting seed culture was inoculated into 50 mL R5α liquid medium at a 2% ratio (v/v). After 4 days fermentation, the broth was harvested by centrifugation, extracted with 50 mL ethyl acetate three times, and concentrated in vacuo. The samples were re-dissolved in 1 mL acetonitrile and subjected to HPLC analysis.
Complementation of S. paulus pau20∷aac(3)IV by feeding DHHA or 3-HAA
For the feeding experiments, S. paulus pau20∷aac(3)IV was cultured at the paulomycin production condition. DHHA or 3-HAA was added to the cultures to a final concentration of 2.5 mM two days after seed inoculation. After culturing at 28 °C, 220 rpm for 2 days, the broth was harvested, extracted, and concentrated as described above. The samples were re-dissolved in 1 mL acetonitrile and analyzed by HPLC.
Expression and purification of the DHHA dehydrogenases
The 0.7-kb pau20 gene was cloned using the S. paulus NRRL 8115 genome as a template with primer pair pauN10ES/pauN10ER, verified by sequencing, and inserted into the NdeI/BamHI sites of pET-28a to generate pET-28a-pau20. A single transformant of E. coli BL21(DE3)/pET-28a-pau20 was inoculated into LB medium with 50 μg/mL kanamycin and cultured overnight at 37 °C, 220 rpm. The overnight seed culture was inoculated to the same medium at 1% ratio (v/v) and incubated at 37 °C, 220 rpm until OD600 reached 0.6. Expression of Pau20 was then induced by the addition of isopropyl-β-thiogalactoside (IPTG, 0.05 mM) and further cultured at 16 °C, 180 rpm for 12-16 h.
Purification of Pau20 was carried out with the Ni-NTA affinity column at 4 °C following the manufacturer’s instructions. The E. coli cells harvested by centrifugation (4 °C, 3000 × g, 10 min) were re-suspended in lysis buffer (20 mM Tris-HCl, 500 mM NaCl, 5 mM imidazole, pH 7.9) and burst by ultrasonication. After removing the cell debris by centrifugation (4 °C, 16,000 × g, 30 min), the remaining supernatant was subjected to the Ni-NTA affinity column and washed with washing buffer (lysis buffer with 60 mM imidazole) and then elution buffer (lysis buffer with 500 mM imidazole). The purified protein was desalted and concentrated by ultrafiltration using a Millipore centrifugal filter (15 mL, 10 kDa MWCO) and stored at −80 °C in phosphate-buffered saline (PBS) buffer (pH 7.4) with 20% glycerol.
The other six DHHA dehydrogenase genes, natDB (NatDB Accession: CEK42820), cbxG (CbxG Accession: KDQ70111), dhbX (DhbX Accession: CDG76955), bomO (BomO Accession: ALE27507), calB3 (ClaB3 Accession: AEH42481), and stnN (StnN Accession: AFW04566), were synthesized and inserted into the NdeI/BamHI sites of pET-28a to afford plasmids pET-28a-natDB, pET-28a-cbxG, pET-28a-dhbX, pET-28a-bomO, pET-28a-calB3, and pET-28a-stnN respectively. The plasmids were transformed into E. coli BL21(DE3) individually to afford the protein expression strains. Protein expression and purification of the six DHHA dehydrogenases were carried out using the same procedures as those of Pau20. The purified enzymes were stored at −80 °C in PBS buffer with 20% glycerol. Protein concentrations were measured with a BCA protein assay kit (Takara, Dalian, China).
Enzymatic assays of the DHHA dehydrogenases
The DHHA dehydrogenase activity of Pau20 was tested in a 100 μL mixture containing 50 mM PBS buffer (pH 7.4), 2 mM NAD+, 2 mM DHHA, and 5 μM Pau20 at 30 °C for 3 h. The reaction was quenched by equal volume of CHCl3. After centrifugation, the aqueous phase of the reaction (20 μL) was analyzed by HPLC. The pH value optimization of Pau20 was carried out at 37 °C in buffers of pH ranging from 6.6 to 8.5 (200 mM phosphate buffer for pH 6.0-7.6, and 50 mM Tris-HCl buffer for pH 8.5) for 30 min. Temperature optimization was performed in a 100 μL reaction mixture in 200 mM phosphate buffer (pH 7.0) for 30 min. The steady-state kinetic analysis of Pau20 was carried out in 200 mM phosphate buffer (pH 7.0) at 37 °C. The enzymatic assays of the other six DHHA dehydrogenases and the steady-state kinetic analyses of DhbX, CalB3, and StnN were all carried out using the same reaction conditions as Pau20 in 200 mM phosphate buffer at 37 °C.
Construction and evaluation of E. coli BW-pXB1s-HAA
A 2.6-kb DNA fragment containing the 0.6-kb phzD (PhzD Accession: AAC64487) and the 1.9-kb phzE genes (PhzE Accession: AAC64488) was synthesized and the homolog sequences for ligation independent cloning at its both ends were added by a PCR amplification using primer pair pHAAphzDE-F/pHAAphzDE-R. The 0.7-kb DNA fragment containing the pau20 gene was amplified from the S. paulus NRRL 8115 genome using primer pair pHAApau20-F/pHAApau20-R. The two DNA fragments were inserted into the NcoI/EcoRI sites of pXB1s (p15A origin, medium-copy-number) using one-step cloning kit (Vazyme, Nanjing, China) to generate pXB1s-HAA. E. coli BW-pXB1s-HAA was constructed by introducing pXB1s-HAA into E. coli BW25113. E. coli BW-pXB1s-HAA was cultured in 3 mL LB medium with 50 μg/mL streptomycin at 37 °C overnight. The seed culture was then inoculated into 50 mL M9 medium with 50 μg/mL streptomycin at 1% ratio (v/v). After cultured at 37 °C, 220 rpm for 3 h, 10 mM arabinose was added and the culture was further fermented at 37°C, 220 rpm for 20 h. The broth was centrifuged to remove the cell pellet and the supernatant was subjected to HPLC analysis directly.
Construction of E. coli ΔnadAB-pXB1s-QA, ΔnadAB-pAB1s-QA and ΔnadAB-pAB1s-QA*
E. coli BW25113 nadA∷aph is a commercially available nadA disrupted mutant, in which gene nadA is replaced by the kanamycin resistance gene cassette aph flanked by two FRT (FLP recognition targets) sites. After introducing plasmid pCP20 with the FLP-recombinase gene into E. coli BW25113 nadA∷aph, one of the kanamycin sensitive strains was selected as the nadA deleted mutant E. coli ΔnadA and was verified by PCR using primer pair nadA-F/nadA-R. To knock-out nadAB, P1 bacteriophage was used for dissociation of the nadB gene inactivated mutant E. coli BW25113 nadB∷aph and then introduced into E. coli ΔnadA by transduction (Thomason et al., 2007). One of the kanamycin resistant strains was selected and verified as E. coli ΔnadAB by PCR analysis with primer pairs nadA-F/nadA-R and nadB-F/nadB-R (Supplementary Fig.2b). The 0.6-kb nbaC (NbaC Accession: WP_006094629) was synthesized and the homolog sequences for ligation independent cloning at its both ends were added by a PCR amplification using primer pair NbaC-F/NbaC-R. The nbaC gene was then inserted into the NcoI site of pXB1s-HAA to generate pXB1s-QA, which was transformed into E. coli ΔnadAB to obtain E. coli ΔnadAB-pXB1s-QA.
Plasmids pAB1s-HAA (ColE1 origin, high-copy-number) was constructed similarly as that of pXB1s-HAA by insertion of the 2.6-kb fragment containing genes phzDE and the 0.7-kb fragment containing gene pau20 into the NcoI/EcoRI sites of pAB1s. The 0.6-kb nbaC gene was then inserted into the NcoI sites of pAB1s-HAA to generate pAB1s-QA, which was transformed into E. coli ΔnadAB to obtain E. coli ΔnadAB-pAB1s-QA.
The 0.7-kb dhbX gene was synthesized and the homolog sequences for ligation independent cloning at its both ends were added by a PCR amplification using primer pair DhbX-F/DhbX-R. It was then inserted into the XbaI site of pAB1s-QA to replace the pau20 gene (excised by XbaI digestion) to generate pAB1s-QA*, which was transformed into E. coli ΔnadAB to obtain E. coli ΔnadAB-pAB1s-QA*.
Growth of E. coli strains on M9 plates
The E. coli strains were cultured in 3 mL LB medium with appropriate antibiotics at 37 °C, 220 rpm overnight and harvested by centrifugation (4 °C, 3000 × g, 5 min). To remove extracellular NAD(H) and its salvage pathway precursors, the cells were washed twice with ddH2O and diluted to an OD600 of 0.1. For the growth experiments on agar plates, 2 μL dilutions (OD600 of 0.1) of different E. coli strains were spotted onto M9 agar with varied supplements. E. coli ΔnadAB was spotted onto M9 agar supplemented with or without 10 mM QA; E. coli ΔnadAB-pXB1s-QA and ΔnadAB-pAB1s-QA and their controls with empty vectors were inoculated onto M9 agar (50 μg/mL streptomycin) with or without 10 mM arabinose. The E. coli strains were grown at 37 °C.
Construction and evaluation of the DMP cell factory
A 2.4-kb DNA fragment containing the 1.0-kb EcTdh (EcTDH, Accession No.: WP_000646007) and the 1.4-kb SpaNox (SpaNox, Accession No.: WP_002989788) genes was synthesized and inserted into the NcoI/XhoI sites of pRSFDuet-1 to afford plasmid pRSF-EcTdh-SpaNox. E. coli C3N-DMP and the control strain E. coli DMP-Con were constructed by introducing pRSF-EcTdh-SpaNox into E. coli BL21-C3N (containing plasmid pAB1s-QA*) and BL21-Con (containing plasmid pAB1s), respectively. Briefly, E. coli strains C3N-DMP and DMP-Con were cultured in 3 mL LB medium with 50 μg/mL streptomycin and 50 μg/mL kanamycin at 37 °C overnight. The seed cultures were inoculated into 100 mL M9Y medium (M9 Medium containing 0.1% yeast extract) with 50 μg/mL streptomycin and 50 μg/mL kanamycin at 1% ratio (v/v). After culturing at 37 °C, 220 rpm for 3 h, 0.1 mM IPTG and 10 mM arabinose were added and the cultures were further cultured at 37 °C, 220 rpm for 24 h. The cells were then collected by centrifugation (4 °C, 3,000 × g, 20 min), washed with ddH2O and resuspend in 100 mM Tris-HCl buffer (pH 8.0). The whole-cell conversions were started in 2 mL Eppendorf tubes containing 1 mL 100 mM Tris-HCl buffer (pH 8.0) under 28 °C and 230 rpm for 12 h. The initial concentrations of L-threonine and the resting cells were set as 200 mM and OD600 of 35, respectively. To detect the production of DMP, the reaction samples were centrifuged (4 °C, 16,000 × g, 10 min) to remove the pellets, diluted 4 times with acetonitrile, and subjected to HPLC analysis.
Construction of E. coli strains BW-C3N-ChA1 to BW-C3N-ChA4
The codon-optimized alcohol dehydrogenase TesADH gene (TesADH, Accession No.: WP_041589967, point mutations W110A and G198D were modified according to Thompson et al., 2017) and amine dehydrogenase CalAmDH gene (CalAmDH, Accession No.: WP_007505854, point mutations K68S and N266L were modified according to previous reported43) were synthesized and inserted into the NcoI/BamHI sites of pET-28a to afford plasmids pET28a-TesADH and pET28a-CalAmDH, respectively. The 1.1-kb DNA fragment containing the TesADH gene was amplified from the pET28a-TesADH using primers T7/Cal-P23R and inserted into the NcoI/SalI site of pCDFduet-1a (derivate from pCDFduet-1, AmpR) to generate pCDF-TesADH. The 1.1-kb DNA fragment containing the CalAmDH gene was amplified from the pET28a-CalAmDH using primers CALRS-F/CALRS-R and inserted into the NdeI/XhoI site of pCDF-TesADH to generate pCDF-TesADH-CalAmDH. E. coli BW-C3N-ChA1 was obtained by introducing pCDF-TesADH-CalAmDH into E. coli BW-C3N (E. coli BW25113(DE3) containing plasmid pAB1s-QA*), and the control strain E. coli BW-ChA1-Con was constructed by introducing the same plasmid into E. coli BW-Con (E. coli BW25113(DE3) containing plasmid pAB1s). To construct E. coli BW-C3N-ChA2, the CalAmDH gene was cloned from pET28a-CalAmDH with primers T7/Cal-his-R and inserted into the NcoI/SalI site of pACYduet-1 to generate pACYC-CalAmDH, and the resultant plasmid was introduced into E. coli BW-C3N-ChA1. The 2.3-kb DNA fragment containing genes TesADH and CalAmDH was amplified from pCDF-TesADH-CalAmDH using primers T7/Cal-P23R and cloned into the NcoI/XhoI site of pRSFduet-1 (RSF origin, high-copy-number) to generate pRSF-TesADH-CalAmDH, which was transformed into E. coli BW-C3N and BW-Con respectively to afford E. coli BW-C3N-ChA3 and the control strain BW-ChA3-Con. E. coli BW-C3N-ChA4 was obtained by introducing pACYC-CalAmDH into E. coli BW-C3N-ChA3.
General procedures for bioamination of rac-alcohols
The engineered E. coli strains was inoculated in 3 mL LB broth with appropriate antibiotics when necessary (50 μg/mL kanamycin, 50 μg/mL streptomycin, 100 μg/mL ampicillin, 20 μg/mL chloramphenicol) and cultured at 37 °C, 220 rpm overnight. The seed cultures were then inoculated into 100 mL M9Y medium at 1% ratio (v/v). After culturing at 37 °C, 220 rpm for 3 h, 0.1 mM IPTG and 10 mM arabinose were added and the cultures were further fermented for 24 h. The cells were then harvested by centrifugation (4 °C, 3,000 × g, 20 min), washed by ddH2O and re-suspended in appropriate buffers for the whole-cell bioconversions. For comparison of the bioamination efficiencies of E. coli BW-ChA1-Con and BW-C3N-ChA1, the bioconversions were carried out in 100 mM KPi buffer (pH 8.5) with 10% DMSO in 1 mL reaction volume with 5 mM 1a, 2 M NH4COOH under 30 °C, 230 rpm for 10 h. To find an optimal bioamination procedure, varied buffers (100 mM KPi, 100 mM Tris-HCl, and 2 M NH3/NH4Cl (all buffers were adjusted to pH 8.5 and with 10% DMSO), concentrations of NH4+ (range from 0.5 M to 4 M), amounts of resting cells (OD600 of 30, 50 and 70), and reaction temperatures (25 °C, 30 °C, 37 °C, and 42 °C) were sequentially optimized in 1 mL reactions with 5 mM substrate 1a and keeping the agitation at 230 rpm.
The analytical scale bioamination reactions of E. coli BW-C3N-ChA1 to BW-C3N-ChA4 with different alcohol substrates were then carried out under the optimal condition (100 mM KPi buffer (pH 8.5) with 10% DMSO, 5 or 20 mM rac-alcohols, 4 M NH4COOH, OD600 of 50, 37 °C, 230 rpm) in 2 mL Eppendorf tubes containing 1 mL reaction volume for 10 h. To scale up the biotransformation system, E. coli BW-NAD-ChA3 was cultured, collected and resuspended in a 250 mL flask containing 30 mL or 100 mL KPi buffer (100 mM, pH 8.5) with 10% DMSO, 30 mM rac-2a, 4 M NH4COOH, OD600 of 50 resting cells, and the whole-cell bioconversions were performed at 37 °C, 230 rpm for 24 h. All biotransformation reactions were quenched using 100 μL 10 M KOH per mL reaction mixture. After centrifugation at 16,000 × g for 10 min, 200 μL supernatant was extracted by the same volume of ethyl acetate three times. The organic phase (20 μL) was then subjected to HPLC analysis.
Derivatization of the chiral amine samples
The enantiomeric excess of the amine products was analyzed using the FDAA (Marfey’s Reagent) pre-column derivatization method. Briefly, the reaction mixture (200 μL) was extracted by ethyl acetate, dried in vacuum, and re-suspended in 20 μL acetone. To perform the derivatization, the sample was incubated with 40 μL FDAA (1% pre-dissolved in acetone, w/v) and 40 μL 1 M NaHCO3 at 40 °C for 1 h. The reaction was quenched by adding 80 μL 1 M HCl and then diluted with 800 μL ethyl acetate. The supernatant was subjected to HPLC analysis after centrifugation.
Measurement of intracellular NAD(H) concentrations
The E. coli strain was cultivated in 3 mL LB medium with appropriate antibiotics at 37 °C, 220 rpm overnight, and the overnight culture was then inoculated into 3 mL or 50 mL M9 medium at 1% ratio (v/v) and grown at 37°C, 220 rpm for 18-24 h. To measure the intracellular NAD(H) concentration, the cells were harvested by centrifugation (4 °C, 3,000 × g, 5 min) at the time point of four hours after entering stationary phase. The cell pellets were washed with ice-cold PBS buffer, re-suspended in the same buffer and adjusted to OD600 of 1.0. The concentrations of NAD(H) (total NAD+/NADH) and NADH only were measured by the NAD+/NADH assay kit (Abcam, Cambridge, UK) according to the manufacturer’s instructions. E. coli cell concentration was calculated as 109 cells/mL at OD600 of 1.0 and the volume of one E. coli cell was 10−12 mL36, 50 (Zhou et al., 2011; Brumaghim et al., 2003).
Spectroscopic analysis
HPLC analysis was carried out on a Shimadzu HPLC system (Shimadzu, Kyoto, Japan). The analysis of paulomycin production was performed using an Apollo C18 column (5 μm, 4.6 × 250 mm, Alltech, Deerfield, IL, USA), which was developed with a linear gradient using acetonitrile and water with 0.1% trifluoroacetic acid at a flow rate of 0.8 mL/min. The ratio of acetonitrile was maintained at 5% for 5 min and changed linearly from 5% to 90% over 5-25 min and from 90% to 100% over 25-30 min. The detection wavelength was 320 nm.
The formation of NADH in the DHHA dehydrogenase reactions was monitored by BioTek Synergy H4 Hybrid Reader (BioTek, Vermont, USA) at 340 nm. HPLC analysis of the DHHA dehydrogenase reactions and production of 3-HAA in E. coli was carried out with a ZORBAX SB-Aq StableBond Analytical column (5 μm, 4.6 × 250 mm, Agilent Teologies, Santa Clara, CA, USA). A linear gradient of acetonitrile and water with 0.1% trifluoroacetic acid was used for development of the column at a flow rate of 0.8 mL/min. The ratio of acetonitrile was changed linearly from 1% to 50% over 0-30 min, from 50% to 100% over 30-32 min, and maintained at 100% for 5 min. The detection wavelength was 254 nm or 294 nm.
The production of DMP was analyzed using an Apollo C18 column (5 μm, 4.6 × 250 mm, Alltech, Deerfield, IL, USA) using an isocratic elution of acetonitrile/water (0.1% trifluoroacetate, v/v) (20:80, v/v) at 0.8 mL/min. The detection wavelength was 275 nm.
The bioamination samples were analyzed using an Apollo C18 column (5 μm, 4.6 mm × 250 mm, Alltech, Deerfield, IL, USA) using an isocratic elution of acetonitrile/water (0.1% trifluoroacetate, v/v) (30:70, v/v) at 0.8 mL/min. The detection wavelength was 254 nm. HPLC analysis of the enantiomeric excess of chiral amine products was carried out with the same Apollo C18 column with a linear gradient using acetonitrile with 0.1% formic acid and water with 0.1% formic acid at a flow rate of 1 mL/min. The percentage of acetonitrile started being held constant at 20% for 10 min, changed from 20 to 60% over 30 min and from 60 to 100% over 5 min. The detection wavelength was 340 nm.
LC-MS analysis was performed with the ZORBAX SB-Aq StableBond Analytical column (5 μm, 4.6 × 250 mm, Agilent Teologies, Santa Clara, CA, USA) on an Agilent 1260/6460 Triple-Quadrupole LC/MS system (Santa Clara, CA, USA).
Chemicals
The racemic alcohols 3a and 5a were chemically synthesized by reduction of the commercially available ketones 3b and 5b; the ketone 1b was chemically synthesized by oxidation of 1a. Enantiomeric standard (S) and (R) amines of 1c-5c were synthesized by stereoselective amination using previously described enzymatic methods51.
Acknowledgments
This work was supported in part by the National Key R&D Program of China (2018YFA0901600 and 2018YFA0901900), the National Natural Science Foundation of China (31670032 and 31522001). Y.C. is an awardee for the ‘Hundred Talents Program’ of CAS.
Footnotes
↵5 Lead Contact





{kind=link}
{kind=link}
{kind=link}
{kind=link}