

Abstract
Extinct large volant birds and pterosaurs are thought to have used wind-dependent soaring flight, similar to modern large birds. There are two types of soaring: thermal soaring, used by condors and frigatebirds, which involves the use of updrafts to ascend and then glide horizontally; and dynamic soaring, used by albatrosses, which involves the use of differences in wind speed with height above the sea surface. However, it is controversial which soaring styles were used by extinct species. In this study, we used aerodynamic models to comprehensively quantify and compare the soaring performances and wind conditions required for soaring in two of the largest extinct bird species, Pelagornis sandersi and Argentavis magnificens (6–7 m wingspans), two pterosaur species, Pteranodon (6 m wingspan) and Quetzalcoatlus (10 m wingspans), and extant soaring birds. For dynamic soaring, we quantified how fast the animal could fly and how fast it could fly upwind. For thermal soaring, we quantified the animal’s sinking speed circling in a thermal at a given radius and how far it could glide losing a given height. Consistently with previous studies, our results suggest that Pteranodon and Argentavis used thermal soaring. Conversely, the results suggest that Quetzalcoatlus, previously thought to have used thermal soaring, was less able to ascend in updrafts than extant birds, and Pelagornis sandersi, previously thought to have used dynamic soaring, actually used thermal soaring. Our results demonstrate the need for a comprehensive assessment of performance and required wind conditions when estimating soaring styles of extinct flying species.
Introduction
Flying animals have evolved a wide range of body sizes. Among them, there have been incredibly large species of birds and pterosaurs (Fig. 1). Pelagornis sandersi and Argentavis magnificens are the largest extinct volant birds. Their estimated wingspans reached 6–7 m (1–4), twice as large as that of the wandering albatross, the extant bird with the longest wingspan (Table 1). Several large species of pterosaurs appeared in the Cretaceous period. Pteranodon, presumably the most famous pterosaur, is estimated to have had a wingspan of 6 m (Table 1) (5). The azhdarchids are one of the most successful Cretaceous pterosaur groups and include many large species with wingspans of approximately 10 m (Table 1) (6–9). Quetzalcoatlus northorpi, one of the azhdarchid species, is regarded as one of the largest flying animals in history.

Discoveries of these giant winged animals have fascinated paleontologists and biologists for over a century. Their flight ability has been of great interest because their huge size must significantly affect their flight. With increasing size, flapping becomes more costly; the power required to fly increases faster than the power muscles can produce. Two main arguments are often debated about the flight of extinct giants, both stemming from this physical constraint: the first is about whether they were able to take off (4, 10–12); the second is about what kind of soaring flight style they employed (1, 4, 13–15). This study focuses on the latter debate. Due to the high cost of flapping, large extant birds prefer to fly utilizing wind energy, that is, they prefer to soar. It is presumed that extinct large animals also employed soaring flight as their primary mode of transportation (1, 4, 14). For example, in Quetzalcoatlus, flapping flight might have required anaerobic exercises that were difficult to sustain for long periods of time (14).
There are two main soaring flight styles among extant birds: dynamic soaring and thermal soaring (16). In dynamic soaring, birds extract flight energy from wind shear—the vertical gradient in horizontal wind speed over the ocean (Fig. 2A). In extant birds, seabirds (e.g., albatrosses, shearwaters, and petrels) employ this soaring style and can routinely travel hundreds of kilometers per day over the sea. In thermal soaring, birds first fly circling in warm rising-air columns (thermals). They climb to a substantial height and then glide off in the desired direction while losing their height (Fig. 2C-E). By repeating this up-down process, birds travel over vast distances. Various terrestrial bird species (e.g., vultures, eagles, and storks) and seabirds (e.g., frigatebirds and pelicans) employ thermal soaring.

This study investigated which soaring styles were employed by four large extinct species, that is, Pelagornis sandersi, Argentavis magnificens, Pteranodon, and Quetzalcoatlus. Their soaring capabilities were quantified using two factors. The first is performance, for example, the available speed and efficiency of soaring. The second is the minimum wind speed required for sustainable soaring flight.
For thermal soaring, performance was evaluated in the gliding and soaring up phases. In the gliding phase, a bird’s performance is its glide ratio, which is the ratio of distance a bird traverses to the height the bird loses to cover that distance, i.e., (glide ratio) = (horizontal speed)/(sinking speed) (Fig. 2E). A bird with a higher glide ratio can traverse a longer distance for the same amount of height lost as a bird with a lower glide ratio. Plotting a curve known as a ‘glide polar’ is the conventional method to quantify glide ratio and the associated speed in artificial gliders and birds (16). This curve is a plot of the sinking speed against horizontal speed when a bird glides in a straight line. We can determine the flyer’s highest glide ratio and the associated travel speed by finding the line that passes the origin and tangents of the glide polar.
The inverse of the line’s slope and the speed at the tangent point correspond to the highest glide ratio and the associated horizontal speed, respectively. The performance in the soaring up phase is represented by the sinking speed during circling in a given radius. To achieve thermal soaring, a bird needs a higher upward wind speed than the circling bird’s sinking speed (Fig. 2D). Because thermals have a stronger updraft in the center (Fig. 2C), the animal needs to achieve not only low sinking speed but also a narrow turning radius to efficiently ascend through the thermal. A ‘circling envelope’ curve visualizes this performance and the required minimum updraft wind speed. This curve is a graph of the minimum sinking speed (i.e., the required minimum upward wind speed for ascent) against the radius of turn when a bird glides in a steady circle. Circling envelopes have been used to quantify the soaring up ability of extant bird species (16, 17).
For sustainable dynamic soaring, a certain amount of horizontal wind speed is essential. With respect to the performance of dynamic soaring, we used two metrics. For soaring animals, traveling fast enables them to save energy costs. Hence, the first dynamic soaring performance we measured was the maximum travel speed averaged over one dynamic soaring cycle under a given wind speed (Fig. 2B). In addition, the ability to travel in a desirable direction irrespective of unfavorable wind conditions is also necessary. In particular, traveling upwind is the most challenging situation for flying animals. Accordingly, the second performance we measured was the maximum value of the upwind speed, that is, the parallel component of the travel speed to the upwind direction averaged over one soaring cycle (Fig. 2B). We previously lacked an established framework to quantify the dynamic soaring performances and minimum required wind speed. In previous studies, the glide polar has been used to quantify the dynamic soaring ability of animals (1, 13). However, the glide polar assumes that birds glide in a straight line, which is not the case during the complicated process of dynamic soaring. Hence, the glide polar may be an insufficient metric to describe dynamic soaring performance. A numerical optimization method developed in the engineering field was recently proposed to quantify the dynamic soaring performance of birds, requiring wind conditions (18, 19). However, despite its effectiveness, the only animal to which this technique has been applied is the wandering albatross (18, 19); it has never been applied to extinct giant flyers.
The soaring performances and required wind conditions have not been evaluated comprehensively for the four giant extinct species summarized in Table 2. Table 2 shows three knowledge gaps. First, the wind condition and the performance of dynamic soaring have rarely been evaluated. This is due to the lack of a framework to assess dynamic soaring ability, as mentioned above. Second, the thermal soaring performance in the soaring up phase and the minimum required updraft wind speed have not been evaluated for Pelagornis sandersi, Pteranodon, and Quetzalcoatlus. Finally, recent studies estimated that the body masses of Pteranodon and Quetzalcoatlus were about three times heavier than previously expected (5, 10,20, 21), and the soaring abilities of these new heavy body masses have rarely been evaluated.Based on these three gaps, we summarize the previous studies on these four species’ soaring styles.
Pelagornis sandersi
Pelagornis sandersi is predicted to be a dynamic soarer rather than a thermal soarer as its glide polar (and glide ratio that can be derived from its glide polar) is more similar to those of living dynamic soarers than those of living thermal soarers (1). However, this means our understanding of this species’ soaring style has been based on just one metric, that is, its glide ratio (Table 2). Hence, evaluating other metrics of this species, i.e., the required wind conditions in dynamic and thermal soaring, and performance in dynamic soaring and in the soaring up phase of thermal soaring is important for a more accurate estimate of the soaring style of this species. A previous study cautiously calculated Pelagornis sandersi’s glide polars for 24 combinations of estimates (body mass = 21.8 and 40.1 kg, wingspan = 6.06, 6.13, 6.40, and 7.38, and aspect ratio = 13,14, and 15) to deal with morphological uncertainty. Hence, we also employed these estimates in this study.
Argentavis magnificens
Argentavis magnificens is expected to be a thermal soarer. A previous study reported that the thermal soaring performance and required wind conditions of this species are comparable to living thermal soaring species based on glide polars and circling envelopes (4). This result is consistent with the fact that an Argentavis specimen was found on the foothills and pampas of Argentina, far from coastlines (4).
Pteranodon and Quetzalcoatlus
Although assessments of the soaring abilities of Pteranodon and Quetzalcoatlus have been a long-standing issue, we still lack a comprehensive understanding of their soaring style due to many uncertainties in the estimates of their morphology, especially because of the significant changes in weight estimates around 2010. Previously, it was estimated that Pteranodon had a wingspan of around 7 m and a body mass of 16 kg, while Quetzalcoatlus had a wingspan of around 11 m and a body mass of 50–70 kg. Based on these estimates, previous studies argued that they were adapted to thermal soaring (22, 23) and others argued that they could also employ dynamic soaring (13). Around 2010, however, many studies with different approaches suggested that pterosaurs were much heavier than previously expected (5, 10, 20, 21). For example, Witton estimated that Pteranodon was 36 kg with a 5 m wingspan, and Quetzalcoatlus was 259 kg with a 10 m wingspan (5). Other studies also reported similar estimated values (10, 20, 21). Few studies have quantified the soaring performance of these species based on these new heavy body mass estimates. For example, Witton and Habib argued that Pteranodon was a dynamic soarer and Quetzalcoatlus was a thermal soarer by comparing the aspect ratios and the relative wing loadings of these species with those of extant soaring bird species (5, 14). Conversely, a recent study quantified the cost of transport and sinking speed in the gliding of 128 pterosaur species and showed that azhdarchoid pterosaurs, including Quetzalcoatlus, had lower flight efficiency than the other pterosaurs (24). However, despite these studies, we still have not been able to comprehensively quantify the performance and wind requirements of dynamic and thermal soaring of these species.
Furthermore, pterosaurs have a wing morphology that is completely different from that of birds and bats. Some studies reported that the wings of pterosaurs would have been associated with high-profile drag (drag stemming from the wings) (15, 25). Palmer experimentally measured profile drag in a wind tunnel experiment using reconstructed pterosaurs wings (15). With the experimentally derived profile drag, Pteranodon’s glide polars with the heavy body mass estimates were determined. Palmer concluded that Pteranodon adopted thermal soaring with a slow flight speed.
In this study, we aimed to address the knowledge gaps in the soaring abilities of these four extinct giants. First, we calculated the dynamic soaring performances and required wind speeds using a physical model and a numerical optimization method. This method has been developed in the engineering field and provides a framework to quantify dynamic soaring performances and required wind conditions. We applied this framework to the four giant extinct species and three extant dynamic soaring bird species with various sizes ranging from 1 to 9 kg [i.e., the wandering albatross (Diomedea exulans), the black-browed albatross (Thalassarche melanophris), and the white-chined petrel (Procellaria aequinoctialis)]. As the exact shape of the wind gradient is still poorly understood, we conducted the calculation under seven different wind conditions (Fig. 3A–C). In addition, we added an important modification to the previous models: the animal’s wings do not touch the sea surface during their flight (Fig. 3D; and see Eq. 16 and its description in Materials and Methods for details). Second, we quantified the thermal soaring performances and the required upward wind speeds for the four extinct species, five extant thermal soaring species [the magnificent frigatebird (Fregata magnificens), the black vulture (Coragyps atratus), the brown pelican (Pelecanus occidentalis), the white stork (Ciconia ciconia), and the California condor (Gymnogyps californianus)], and kori bustard (Ardeotis kori), the heaviest extant volant bird that does not soar. These values were calculated using the established framework (i.e., glide polars and circling envelopes). Third, to quantify the soaring performances of Pteranodon and Quetzalcoatlus, we used the recent heavy body mass estimates (5). The profile drag coefficients of the pterosaurs (CDpro) were explored for two different cases: the same value as for birds (CDpro = 0.014) (16) and a higher value based on reconstructed pterosaur wings (CDpro = 0.075) (15, 25).
Results
Dynamic soaring
Our computation results indicate that Argentavis magnificens and Quetzalcoatlus could not have employed dynamic soaring (Fig. 4 A–C). Both species showed lower dynamic soaring performances and higher required wind speeds for dynamic soaring than the extant dynamic soaring species under all wind conditions tested in this study.

The dynamic soaring performances and required wind speeds of Pelagornis sandersi and Pteranodon varied substantially with the assumed morphology and shape of the wind gradient, especially the height where the steep wind speed change occurs (Fig. 4 A–C).
When Pteranodon was analyzed with a high-profile drag coefficient it showed poor performance and required strong winds compared with extant species, suggesting that Pteranodon did not employ dynamic soaring. When Pteranodon was analyzed with a low-profile drag coefficient value, its performance outperformed that of extant birds when the wind change was located high from the sea surface (sigmoidal wind condition with hw = 5 and 3). When the wind speed change was located close to the sea surface (logarithmic model and sigmoidal wind condition with hw = 1), Pteranodon showed a poor flight performance and required a high wind speed. It required a stronger wind speed than the extant dynamic soaring species, except for in sigmoidal wind conditions with hw = 5. Hence, Pteranodon could not employ dynamic soaring when a high-profile drag was assumed. Conversely, when a low-profile drag was assumed, Pteranodon was capable of dynamic soaring but required strong wind conditions.
For Pelagornis sandersi, the results were highly dependent on body mass. With heavy body mass estimates (40.1 kg), Pelagornis sandersi required higher wind speeds than extant dynamic soaring species, irrespective of the wind conditions. The performance was superior to extant species for some morphology estimates when the shear height was high from the sea surface (sigmoidal wind condition with hw = 5 and 3 m), but inferior when the wind speed change was located close to the sea surface (logarithmic model and sigmoidal wind condition with hw = 1). When lower body mass estimates were used (21.8 kg), Pelagornis sandersi required lower wind speeds, but its performance (maximum speed and maximum upwind speed) was distinctively lower than that of extant species. Hence, Pelagornis sandersi required harsh wind conditions for dynamic soaring when a 40.1 kg body mass was assumed, and it was poor at dynamic soaring when a 21.8 kg body mass was assumed.
The performances of all species varied with the value of hw, and the variation was especially distinct for large species in contrast to that of white-chinned petrels (Fig. 4 B and C). This variation was due to wingtip boundary conditions. Animals can attain more energy when passing through large wind speed gradients, but when large gradient changes are close to sea level, large animals are unable to use the wind speed gradient efficiently because their wings limit the altitude available to them. In other words, our results show that it is not enough to discuss the ability of dynamic soaring in terms of morphology and glide polars alone. Although the long, thin wings that reduce drag in extant dynamic soaring birds are suited for dynamic soaring (16, 26), detailed dynamic models have shown that excessively long wings can also inhibit efficient dynamic soaring.
Thermal soaring
Extinct species showed high gliding performances with maximum glide ratios ranging from 11 to 22 (Fig. 5A), which are comparative to those of extant species (from 11 to 18); nevertheless, Quetzalcoatlus had the lowest soaring efficiency (i.e., 8) among the other species when a high drag coefficient was assumed (CDpro = 0.075).

With respect to the soaring up phase, all of the extinct giant flyers, except for Quetzalcoatlus, had performances equivalent to or better than the extant species (Fig. 5B). As a previous study has already shown, the circling up performance of Argentavis magnificens was comparable to that of the California condor, one of the largest living thermal soarers (4). The performance of Pteranodon was comparable to that of living thermal soarers, irrespective of the value of the profile drag coefficient (CDpro = 0.014 or 0.075). With a higher drag coefficient (CDpro = 0.075), a narrower circling radius was achieved by Pteranodon, and its circling envelope was very similar to that of the black vulture. The thermal soaring ability of Pelagornis sandersi when a light mass was assumed (21.9 kg) was outstanding. It outperformed many extant thermal soaring species in soaring up ability, and was even comparable to the magnificent frigatebirds, the champion of thermal soaring among extant species. Even with a heavier body mass estimate (40.1 kg), Pelagornis sandersi still outperformed or was comparable to many other species.
Among the four extinct giant animals investigated in this study, the soaring up performance of Quetzalcoatlus was exceptionally low. It required the strongest upward wind speed and the widest circle radius. Its performance was even lower than that of the kori bustard, one of the heaviest volant extant birds that spend most of their time on land and only fly in emergencies, such as when they are under predation risk.
Discussion
Using detailed physical models of soaring birds, we computed and compared the dynamic and thermal soaring performances and the required wind conditions for soaring of four extinct giant flyers with those of extant dynamic and thermal soaring species. Our results indicate that Argentavis magnificens and Pteranodon were thermal soarers, confirming previous studies (4, 15). However, our results also indicate that Quetzalcoatlus could not efficiently perform dynamic nor thermal soaring. In addition, although Pelagornis sandersi was considered a dynamic soaring species in a previous study (1), our results suggest that it was a thermal soaring bird. In the following, we discuss our results in detail for Quetzalcoatlus and Pelagornis sandersi and then describe future issues that need to be addressed for a better understanding of the soaring styles of extinct giant species.
Quetzalcoatlus
There has been a heated debate about the flight capability of Quetzalcoatlus. The focal issues have been whether or not Quetzalcoatlus could take off and if it was capable of sustained flapping flight. Researchers are divided between the opinion that it was too heavy to take off (10, 13, 27) and the opinion that it was able to take off by using quadrupedal launching, like some bats (11, 14). In addition, detailed observations of fossils are also presented as evidence that the giant azhdarchids, including Quetzalcoatlus, were capable of flight; for example, a huge deltopectoral crest on their humeri, which would have anchored muscles for flapping flight (28).
While there is some debate as to whether or not giant pterosaurs could have taken off, it has been widely accepted that if they were able to take off their primary mode of travel would have been thermal soaring rather than flapping flight. Witton and Habib applied a model of bird flap-gliding flight (16) to Quetzalcoatlus, and they found that this species’ flapping flight required anaerobic movement and was difficult to sustain for long periods of time; therefore, it must have relied on wind energy for long-distance travel (14). They also concluded that Quetzalcoatlus used thermal soaring, based on a comparison of its morphology with that of birds (5, 14).
Our results revealed that Quetzalcoatlus had a poor ability to use thermals to ascend. It required a larger turning radius and stronger updraft than the terrestrial kori bustard, let alone species that use thermal soaring. Whether Quetzalcoatlus, with the soaring performance shown in Fig. 5, could routinely travel long distances by thermal soaring is beyond the scope of this study because of the need to examine in detail the wind conditions in their habitat at the time they lived. However, the results of this study alone suggest that Quetzalcoatlus performed poorly at thermal soaring compared with modern and other extinct species, and that the wind conditions under which thermal soaring was possible were limited. This poor thermal soaring performance was due to the large wing loading associated with the large body size. As shown in the Materials and Methods, the turning radius was proportional to the wing loading to the power of one half (eq. 19), and the descent speed during the turn was also proportional to the wing loading to the power of one half, if the effect of the organism’s wing length adjustment was ignored (eqs. 18 and 20, and see also (26)). Since the wing loading is approximately proportional to body size, a giant Quetzalcoatlus required thermals with wider radius and stronger updraft for thermal soaring.
The wing loading also explains why the results of the present study are not consistent with the claims of previous studies that Quetzalcoatlus was adapted to thermal soaring (5, 14). Previous studies compared aspect ratios and relative wing loadings (wing loading divided by body size) between birds, bats, and pterosaurs and concluded that Quetzalcoatlus was adapted to thermal soaring because it was in the domain of thermal soaring birds (and, using the same procedure, it was concluded that Pteranodon was adapted to dynamic soaring). Interspecies comparisons of aspect ratios and relative wing loadings have been made for birds and bats for the purpose of examining the relationship between wing morphology and ecology (29, 30). However, caution should be taken in evaluating soaring ability with this size-independent variable (i.e., relative wing loading) because the thermal soaring ability is inevitably size-dependent, as explained above. When evaluating soaring performance from animal morphology, performance and wind requirements calculated from morphology based on laws of physics (as conducted in this study and (24)) should be more desirable, as taking an inappropriate morphology as a variable may lead to erroneous results. For example, a fictional bird that is 10,000 times larger than but geometrically similar to frigatebirds (i.e., 230 km wingspan and 1.5 × 107 t body mass) needs, in theory, a turning radius and upward wind speed approximately 100 times larger than that of frigatebirds to conduct thermal soaring; however, the aforementioned size-independent metrics method outputs the unrealistic estimation that this bird was as efficient as frigatebirds in thermal soaring.
Recent anatomical studies of the azhdarchid pterosaurs have reported that their skeletal structure shows adaptations to terrestrial walking and suggested that they were terrestrial foragers (31, 32). Furthermore, recent phylogenetic analysis showed that the azhdarchoid pterosaurs differed from other pterosaurs in that they had evolved in the direction of increasing the cost of transport of flapping flight and the sinking speed of gliding (24). Taking into account the adaptations for walking (31, 32), the humeri feature indicating flapping flight capability (28) but not sustainable flapping flight (14), phylogenetic tendency of decreasing flight efficiency (24), and the low thermal soaring ability, we suggest that the flight styles of Quetzalcoatlus and other similar-sized azhdarchid species were similar to those of the bustard or hornbill that spent most of their time on land and rarely flew, except in critical emergencies. Alternatively, they may have routinely flown in early stages of their life history, but as they matured, wing loading would increase, and they would spend most of their time on land.
Pelagornis sandersi
Previously, it was reported that Pelagornis sandersi was a dynamic soarer like the albatross, rather than a thermal soarer like frigatebirds (1). However, we argue that this species is highly adapted for thermal soaring rather than dynamic soaring. The conclusion of the previous study was based on the glide polars of Pelagornis sandersi, which were more similar to those of the wandering albatrosses than those of frigatebirds; glide performance was the only criterion used to evaluate its soaring style. In this study, we quantified other performances and the required wind conditions, which enabled us to evaluate the soaring style of Pelagornis sandersi from multiple perspectives.
Our results indicated that the dynamic soaring performance of Pelagornis sandersi was generally inferior to that of extant dynamic soaring species, although there were substantial variations depending on wind conditions and morphology estimates. One of the factors contributing to the poor dynamic soaring ability of this species was an inability to efficiently exploit the wind speed gradient due to long wings limiting the height above sea level at which the bird could fly. Note that this effect could not be assessed by the glide polars.
Conversely, the thermal soaring ability of Pelagornis sandersi was outstanding regardless of the morphology estimates used. It could circle in narrow thermals with a low sinking speed, as in extant species. Its performance to soar up thermals was even comparative to that of the frigatebird, the champion of extant thermal soaring species. In addition, its glide ratio was also high, outperforming most of the extant thermal soaring species. Accordingly, Pelagornis sandersi showed high performances both in the ascent and gliding phase of thermal soaring, indicating that this species was highly adapted to thermal soaring. Considering that Pelagornis sandersi was found close to the coast, this result indicates that Pelagornis sandersi was well adapted to capture a weak updraft above the sea and could stay aloft for a long time with limited flapping and traveled long distances like frigatebirds.
Future issues
In this section, we discuss some of the simplifications used in this study and issues that we believe need to be examined in the future. The first issue is flight stability. In this study, a steady wind environment was assumed, but actual wind environments fluctuate. In such a fluctuating real-world environment, stability is an important factor that determines the success or failure of flight (33, 34). To simplify our calculations, we did not address stability, but it is important to examine the flight stability of these extinct and extant birds using more detailed morphological information in the future.
The next issue is that the actual wind environment experienced by animals is still largely unknown. For dynamic soaring, the specific form of the wind speed gradient experienced by birds is unknown—for example, whether there is a logarithmic or sigmoidal gust in the shadows of the waves—and information on the height and thickness of the wind speed gradient is not yet known. For this reason, we evaluated performance under various wind conditions (Fig. 3). For thermal soaring, it is also unknown how much updraft animals experience at a given turning radius or the distance between thermals. Recent advances in tracking technology have made it possible to record details of the motion of birds in dynamic and thermal soaring (35–39). We believe that these data will provide information that will help us to understand our results and improve our model, such as the real wind environment experienced by animals (39–43). In addition, it is also important to consider the paleoenvironmental aspects of the wind environment at the time of the extinct species’ inhabitation. For example, we have shown that Quetzalcoatlus had a lower thermal soaring capacity than the extinct species. Paleoclimatic estimates may help us to understand whether the species had a favorable wind environment that allowed it to use thermal soaring as its primary mode of transport, even with their poor soaring up ability.
Finally, we would like to emphasize the importance of a comprehensive assessment of soaring performance and wind requirements of each soaring style, as we have done in this study. Although a great deal of previous research has been done on the soaring performance of extinct species, there have been several evaluation gaps. Our approach filled these gaps and allowed us to examine the soaring style of extinct giants from multiple perspectives, reaching different conclusions about the soaring styles of Quetzalcoatlus and Pelagornis sandersi. Future analyses that take into account the above issues will be necessary to more accurately estimate the soaring ability of extinct giants and thus their paleoecology, which may lead to different conclusions than those of this study. However, the importance of the framework presented here for a comprehensive assessment of soaring flight performance and wind requirements remains unchanged. We hope that this study will serve as a springboard for future discussions of soaring capabilities of extinct species.
Materials and Methods
The dynamics of soaring animals are described using the equations of motion (EOM). We first describe the EOM and parameters therein. Using the EOM, we can calculate soaring performances and the wind speeds required for sustainable soaring. We then describe the calculation procedure for dynamic soaring and thermal soaring, respectively.
Aerodynamic forces and parameters
We regard an animal as a point of mass. The dynamics of the animal’s three-dimensional position X(t) and velocity V(t) are represented by the following EOMs:


When an animal is soaring, three forces—gravitation (mg), lift force (L), and drag force (D)—act on it. Gravitation mg is a product of the constant of gravitation (g) and mass of the bird (m kg), and its direction is towards the ground. The direction of the lift force L is dorsal and perpendicular to the air velocity. Drag force D is against the air velocity. For the analysis of dynamic soaring, we assumed that wind blow is along the y-axis and represent these EOMs in a different way by transforming the ground velocity to pitch γ, yaw ψ, bank angle ϕ, and airspeed V using the following equations (19):




 and
and
 where W(z) represents the wind gradient. A specific form is given in the latter subsection. L represents the strength of the lift force, and D represents that of the drag force. The aerodynamic theory asserts that these values are
where W(z) represents the wind gradient. A specific form is given in the latter subsection. L represents the strength of the lift force, and D represents that of the drag force. The aerodynamic theory asserts that these values are
 and
and

Here, ρ is air density and was set to ρ = 1.225 kg m−3 as in a previous study (1). This is the International Standard Atmosphere values for sea level and 15°C. CL represents the lift coefficient. SW represents the wing area. The drag is composed of three terms. The first term is the profile drag that stems from friction on the body. CDpro is the profile drag coefficient. For birds, a CDpro of 0.014 was employed following previous studies (1, 16). However, based on the reconstruction of pterosaur wings and a wind tunnel experiment, some studies have argued that the pterosaur profile drag coefficient is much higher (approximately 0.05–0.1) (15, 25). Hence, for the profile drag of Pteranodon and Quetzalcoatlus, we explored two cases: a bird-like low-profile drag of 0.014 and an experimentally based high-profile drag of 0.075 (we note that, although the experiments in the previous study assumed a pterosaur with a wing span of approximately 6 m (15), we assumed that the result would not vary significantly with the size of Quetzalcoatlus). The second term is the parasite drag stemming from friction on the wing, where the CDpar is the parasite drag coefficient, and SB is the body frontal area. We used the following recently recommended formula
 on the practical basis that neither CDpar nor SB is exactly known (26). The third term is the induced drag that stems from the lift force. Ra represents the aspect ratio (Ra = b2 / SW, where b is the wingspan). k is the induced drag factor; we set k to 1.1, as in previous studies (1, 14, 16). The lift coefficient has a maximum value; for birds, we set CL to ≤ 1.8 (16). As the aerodynamic properties of pterosaurs can differ from those of birds, and the wind tunnel experiment indicated that CL reached more than 2.0 (15), we set the pterosaurs’ lift coefficient to ≤ 2.2, the same value used in a previous study on pterosaur gliding ability (14).
on the practical basis that neither CDpar nor SB is exactly known (26). The third term is the induced drag that stems from the lift force. Ra represents the aspect ratio (Ra = b2 / SW, where b is the wingspan). k is the induced drag factor; we set k to 1.1, as in previous studies (1, 14, 16). The lift coefficient has a maximum value; for birds, we set CL to ≤ 1.8 (16). As the aerodynamic properties of pterosaurs can differ from those of birds, and the wind tunnel experiment indicated that CL reached more than 2.0 (15), we set the pterosaurs’ lift coefficient to ≤ 2.2, the same value used in a previous study on pterosaur gliding ability (14).
The remaining parameters in the EOMs are body mass (m), wingspan (b), and wing area (Sw). For these morphological parameters of extant birds, we used values reported in previous studies. For Pelagornis sandersi, we used 24 combinations of estimates proposed in a previous study (1). For Argentavis magnificens, we used the estimates in (4). For Pteranodon and Quetzalcoatlus, we used the recent estimates for heavy pterosaurs, as reported in (5). These values are shown in Table 1.
The EOMs include variables that soaring animals can control, that is, bank angle ϕ(t) and lift coefficient CL. Although these variables are time-dependent, for simplicity, we assumed that the animals keep their lift coefficients at a constant value. Hence, using a time series for bank angle, a constant value of CL, and values of parameters, the dynamics of the soaring animals were determined with EOM.
Quantification of the dynamic soaring performance and the required minimum wind speed
Wind gradient model
We explored two types of wind gradients. The first was the logarithmic model represented as
[Logarithmic wind gradient model]

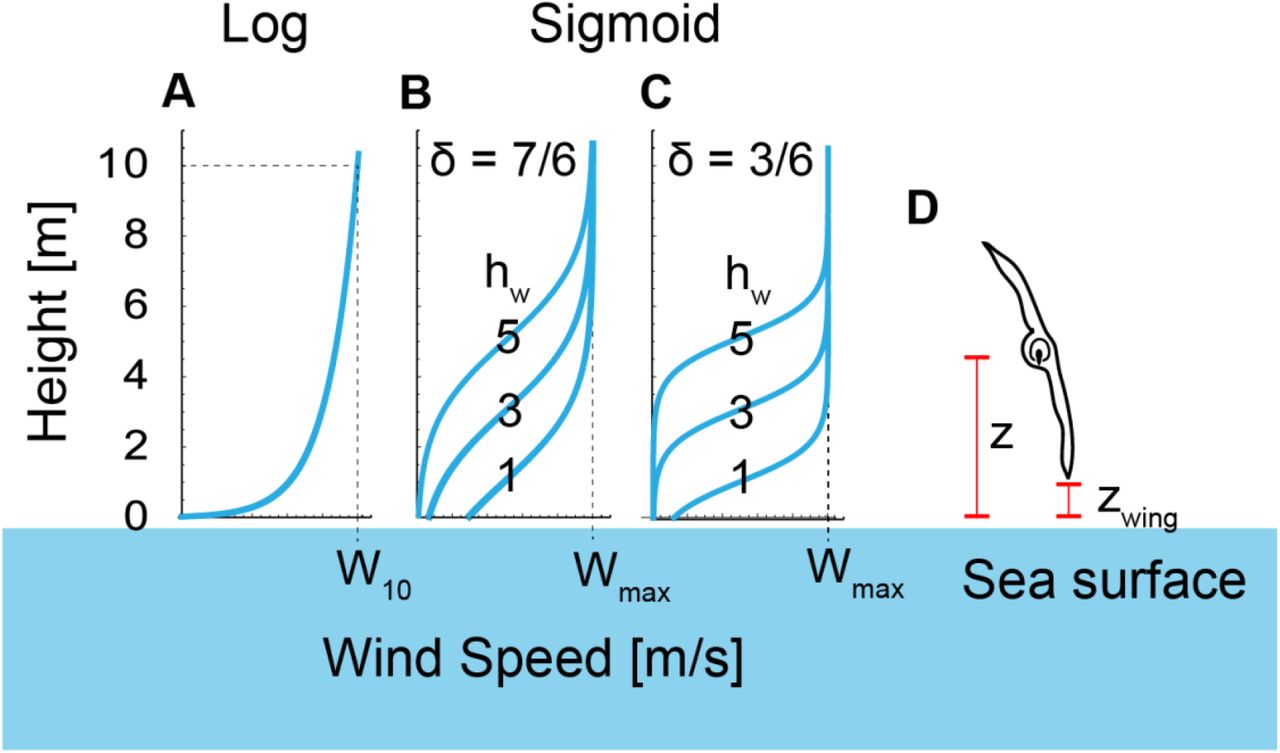
This function is defined at z > hmin. We set hmin to 0.03 [m], following a previous study (18). W10 is the wind speed at height z = 10 m. This model is deemed to be a good model of the average wind field in the first 20 m above the sea surface, assuming a flat sea surface, and has been a popular approach in dynamic soaring modeling. However, recent studies have argued that the real sea surface is not flat, and wind separations in ocean waves may occur more often than expected (44). To describe wind-separation-like wind profiles, a sigmoidal model has been proposed (19, 45). We also employed the sigmoidal wind model with a minor change, represented as
[Sigmoidal wind gradient model]

hw determines the height of wind separation, as shown in Fig. 3. In this study, we set hw to 1, 3, and 5. δ is the thickness parameter. The wind speed changes with height (|z − hw| ≲ 3δ m). In a previous study, the wind shear thickness was speculated as approximately 1.5–7 m. Here, we set δ to 3/6 with a steep wind change, and 7/6 with a gentler change (Fig. 3).
Formulation to numerical optimization
The numerical computation of dynamic soaring performance and minimum wind speed boiled down to the restricted optimization problem. That is, a mathematical problem to find the values of (ⅰ) a certain variable Y that maximizes (ⅱ) an objective function f(Y), satisfying (ⅲ) equalities h(Y) = 0 and (ⅳ) inequalities g(Y) ≦ 0. In the following, we describe the variables, object functions, equalities, and inequalities for dynamic soaring.
(i) Variables
The dynamics of dynamic soaring animals are described by the 3D position (x(t), y(t), z(t)), pitch angle γ(t), yaw angle ψ(t), airspeed V(t), bank angle ϕ(t), lift coefficient CL, and the period of one dynamic soaring cycle τ. Among these variables, 3D position, pitch, yaw, bank, and airspeed are functions of time t (0 ≤ t ≤ τ). Optimization problems that include functions as variables are difficult to be directly solved. Therefore, we employed a collocation approach (19, 46). The collocation approach discretizes the variables in time, such as X(t) (0 ≤ t ≤ τ) to variables Xi = X((i-1)N/τ) (i = 1, N), and converts the EOM to the equalities between those discretized variables. Hereafter, we use X1:N = {X1, X2,…, XN}. In this study, we set the number of discretization points to N = 51 in order to perform computations with reasonable accuracy within a reasonable amount of time. Accordingly, the variables of this optimization problem are position x1:N, y1:N, z1:N, pitch angle γ1:N, yaw angle ψ1:N, airspeed V1:N, bank angle ϕ1:N, lift coefficient CL, and a period of one soaring cycle τ. In addition, when computing the minimum wind speed required for sustainable dynamic soaring, W10 (log model) or Wmax (sigmoid model) were also treated as variables. Hence, the total number of variables were 7 × 51 + 2 (+1) = 359 (or 360).
(ii) Object function
First, we computed (1) the minimum wind speed required for sustainable dynamic soaring for each wind gradient model. As the objective function to minimize, we set W10 for the logarithmic model and Wmax for the sigmoidal model. Then, we computed (2) the maximum speed averaged over one dynamic soaring cycle by maximizing the object function to  . Finally, we computed (3) the maximum upwind speed averaged over one dynamic soaring cycle by maximizing the object function yN/τ. With respect to the maximum speed and maximum windward speed, we computed these values for different wind speeds, that is, from the minimum required wind speed of the species to the highest minimum required wind speed among the examined species (i.e., Quetzalcoatlus) +2 m/s. In this wind speed range, the maximum speed reached an unrealistically high value and/or the optimization calculation did not converge for some species. Thus, we stopped the computation of the maximum speed at the wind speed where the maximum speed exceeded 40 m/s (144 km/h).
. Finally, we computed (3) the maximum upwind speed averaged over one dynamic soaring cycle by maximizing the object function yN/τ. With respect to the maximum speed and maximum windward speed, we computed these values for different wind speeds, that is, from the minimum required wind speed of the species to the highest minimum required wind speed among the examined species (i.e., Quetzalcoatlus) +2 m/s. In this wind speed range, the maximum speed reached an unrealistically high value and/or the optimization calculation did not converge for some species. Thus, we stopped the computation of the maximum speed at the wind speed where the maximum speed exceeded 40 m/s (144 km/h).
(iii) Equalities
The first equalities to be fulfilled for dynamic soaring animals are given in Eq. 3–8. The collocation approach converts the EOM into the equalities between the variables listed in the above section. As the number of original EOM was six and the number of discretization was 51, the EOM were converted into 6 × 51 = 306 equalities (see (19, 46) for the specific representations of these equalities).
The second type of equalities to be fulfilled were periodic boundary conditions of dynamic soaring: at the beginning and end of one dynamic soaring cycle, the state of the animal (i.e., pitch, yaw, airspeed, bank, height) is the same, represented as

(iv) Inequalities
First, we assumed that there was a maximum limit of physical load on the animal. This is because dynamic soaring entails dynamic maneuvering, which results in a corresponding acceleration. We employed the approach of a previous study (18) that restricted the load factor (L/mg) to less than 3,

The second inequality was an important modification of the previous models. The height of the animal’s wingtip (zwing) was calculated as above the sea surface and was represented as

Previous studies discarded the existence of the sea surface (19) or restricted birds to only flying higher than a given height (1.5 m) from the sea surface (18). However, the height a bird can fly depends on the wing length and the bank angle (e.g., with a shorter wing length and a lower bank angle, a bird can fly at a lower height). When dynamic soaring birds fly, they adjust their wingtips close to, but avoid touching, the sea surface (Fig. 3D). Dynamic soaring birds can exploit more flight energy when they pass through stronger wind speed differences. As the wind speed difference is strong close to the sea surface, how close to the sea surface a bird can fly is crucial for dynamic soaring birds. Accordingly, long wings may restrict the minimum height at which the bird can fly and disturb efficient dynamic soaring. Hence, considering the effect of wings is crucial for evaluating dynamic soaring performances.
Third, we also assumed that the height of the animal was higher than 0.5 m, that is,

The optimization problem described here is a restricted non-linear optimization problem. We used the SQP method to solve the problem with the “fmincon” function in MATLAB® Ver R2019a.
Quantification of the thermal soaring performance and the required minimum upward wind speed
For the computation of glide polars and circling envelopes, we followed the same procedure as the Flight software developed for evaluating bird flight performance, as described in (16). In the following, we outline the procedure and parameters employed in this study.
First, before computing the glide polars, we determined how gliding animals adjust their wing area with respect to their airspeed. We assumed that the wingspan decreases as a linear function of speed, based on observations in real birds (47). We employed Flight’s default setting, which assumes that wingspan, wing area, and thus aspect ratio linearly decrease with factor β = (Bstop - V/VS)/ (Bstop - 1); i.e., we replaced b, SW, and Ra with βb, βSW, and βRa, respectively. In this equation, VS is the stall speed, the airspeed of the animal at the highest lift coefficient (i.e.,  ); and Bstop is a constant that determines the degree of wing reduction. For birds, we set Bstop to 5, the default value in Flight. For pterosaurs, we set Bstop to 6, following a previous study (14). Then, the glide polars were derived from the EOM, setting bank angle to 0, assuming that the pitch angle was small enough (γ ≪ 1) and considering the gliding animal was at kinematical equilibrium (
); and Bstop is a constant that determines the degree of wing reduction. For birds, we set Bstop to 5, the default value in Flight. For pterosaurs, we set Bstop to 6, following a previous study (14). Then, the glide polars were derived from the EOM, setting bank angle to 0, assuming that the pitch angle was small enough (γ ≪ 1) and considering the gliding animal was at kinematical equilibrium ( , sin γ = D/L ≃ L/mg). Sinking speed was represented as a function of airspeed V (16),
, sin γ = D/L ≃ L/mg). Sinking speed was represented as a function of airspeed V (16),

This relation gives a glide polar. Note that we used eq. 11 (CDpar SB = 0.01Sw; in this equation, SW was not replaced with βSW) to derive the above equation. The horizontal speed is  . Thus, the maximum glide ratio is the maximum value of
. Thus, the maximum glide ratio is the maximum value of  The circling envelope is given with the radius of the circle (r), and the sinking speed in a circling glide (VSink,Circle) is represented by the bank angle (ϕ),
The circling envelope is given with the radius of the circle (r), and the sinking speed in a circling glide (VSink,Circle) is represented by the bank angle (ϕ),
 and
and
 where
where  and VSink,min are the lift coefficient and the sinking speed at the minimum sinking speed of the glide polar, respectively (16).
and VSink,min are the lift coefficient and the sinking speed at the minimum sinking speed of the glide polar, respectively (16).
Notes
The authors declare no competing interest.
Acknowledgments
The authors thank Chihiro Kinoshita for illustrating Fig. 1. We thank Fujiwara Shin-ichi for reading the draft of this paper and for providing valuable comments. This study was financially supported by Grants-in-Aid for Scientific Research from the Japan Society for the Promotion of Science (16H06541, 16H01769), and JST CREST Grant Number JPMJCR1685, Japan.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}