

Abstract
The development of preventive corona virus disease (COVID)-19 vaccines is an urgent need, especially for the aging population that is most affected by the ongoing pandemic. The Janssen Ad26.COV2.S vaccine candidate is currently the only one evaluated as a single dose vaccination regimen in Phase 3 clinical studies. While the advantages of single dose vaccines, especially for use during a pandemic, are obvious, multiple doses may potentially improve magnitude and durability of immune responses. Here we assessed the immunogenicity of one- and two-dose Ad26.COV2.S vaccine regimens in adult and aged non-human primates (NHP). A second vaccine dose, administered 8 weeks post the first immunization, induced a significant increase in antigen-specific binding and neutralizing antibody responses in both adult and aged animals as compared to a single dose. In addition, in one-dose regimens neutralizing antibody responses were maintained for at least 14 weeks, providing an early indication of durable immune responses elicited by Ad26.COV2.S. Similar to what we showed previously in adult animals, Ad26.COV2.S vaccination of aged NHP induced a CD8+ T cell response and a Th1 skewed CD4+ T cell response. These data support the initiation of a two-dose Ad26.COV2.S regimen in a Phase 3 clinical trial in adults and elderly.
Introduction
Development of safe and effective vaccines to control the ongoing COVID-19 pandemic (Cucinotta & Vanelli, 2020)(WHO, 2020) caused by severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) (Wu et al., 2020; Zhu et al., 2020) is a global priority. Ideally, especially in the context of a pandemic, a vaccine provides both an early onset of protection and durable protection. The durability of vaccine elicited protection depends on the capacity of the vaccine platform, specific antigen (design) and vaccination regimen to efficiently stimulate the immune system (Cohen, 2019)(Pulendran & Ahmed, 2011) and on several characteristics linked to the recipient of the vaccine (Zimmermann & Curtis, 2019). Age for instance plays an important role, as in the elderly the immune response to vaccines is usually reduced in magnitude and duration, potentially resulting in reduced vaccine efficacy (Wagner et al., 2018)(Crooke et al., 2019) (Gustafson et al., 2020)(Weinberger, 2018). Although people of all ages are at risk of contracting COVID-19 disease, the risk of developing severe or critical illness increases with age (Mallapaty, 2020)(CDC, 2020), warranting the testing of COVID-19 vaccine candidates in different age cohorts.
The Ad26.COV2.S vaccine candidate encodes the stabilized full length SARS-CoV-2 spike protein (Bos et al., 2020). In nonclinical efficacy studies, a single dose of Ad26.COV2.S, provided robust protection against SARS-CoV-2 challenge in both upper and lower airways in rhesus macaques (Mercado et al., 2020) and protected Syrian golden hamsters from severe clinical disease (Tostanoski et al., 2020). Protective efficacy strongly correlated with the presence of virus neutralizing activity in serum of NHP (Yu et al., 2020)(Mercado et al., 2020). These data corroborate previously reported findings on SARS-CoV, which showed that neutralizing antibody responses against the SARS-CoV spike protein, that binds to the same cellular receptor as SARS-CoV-2 for cell entry (Shan et al., 2020), were associated with protection against SARS-CoV challenge in nonclinical models (Chen et al., 2005).
Ad26.COV2.S has been shown to elicit a prompt and strong immune response after a single dose vaccination in both adults and elderly humans in an interim analysis, as measured up to day 29 post-immunization in a Phase 1/2a study (Sadoff et al., 2020). Based on this data, the protective efficacy against COVID-19 is currently being evaluated in humans in a Phase 3 efficacy trial (ENSEMBLE trial, NCT04505722). A second Phase 3 study (ENSEMBLE 2, NCT04614948) is planned to evaluate vaccine efficacy and durability of a regimen based on two doses of Ad26.COV2.S, as durability of immunity and efficacy may potentially be enhanced after two vaccine doses. Indeed, in other programs with Ad26-based vaccines, two doses gave higher and more durable immune responses (Geisbert et al., 2011) (Callendret et al., 2018)(Salisch et al., 2019) (Salisch and Stephenson et al, manuscript submitted). Here we report immunogenicity data after one- and two-dose regimens of Ad26.COV2.S in both adult and aged NHP for a follow-up period up to 14 weeks after the first vaccination.
Results
Immunogenicity of one- and two-dose Ad26.COV2.S vaccine regimes in adult rhesus macaques
Adult rhesus macaques (57 females and 3 males, 3.3 - 5.0 years old) were immunized with either a single dose of 1×1011 vp or 5×1010 vp Ad26.COV2.S (n=14 per group) or with two doses of 5×1010 vp Ad26.COV2.S with a 4- or 8-week interval (n=14 per group). A sham control group (n=4) received saline injection at week 0 and week 8. SARS-CoV-2 spike protein-specific antibody responses were followed up every two weeks up to 14 weeks after the first immunization by enzyme-linked immunosorbent assay (ELISA) and pseudovirus neutralization assay (psVNA). Responses were detected in all vaccinated animals as early as two weeks after immunization and significantly increased by week 4 post-immunization (p≤0.010, ANOVA paired t-test) (Figure 1A and B). Animals that received 1×1011 vp Ad26.COV2.S had 1.6 fold higher binding- and 2.1-fold higher serum neutralizing antibody levels (p=0.008 and p=0.004, respectively, ANOVA t-test) relative to animals immunized with 5×1010 vp Ad26.COV2.S. Similar differences in response levels were maintained throughout the entire observation period, however, at week 14 neutralizing antibody titers were similar between the two one-dose groups (p=0.096, ANOVA paired t-test). Spike protein-specific binding antibody levels declined more rapidly than neutralizing antibody levels, irrespective of the vaccine dose the animals had received.


A) SARS-CoV-2 spike protein binding antibody concentrations were measured over time with an ELISA qualified for human samples, using a trimeric, soluble stabilized spike protein produced in mammalian cells as coating antigen. Individual animal levels are depicted with grey points and paired measurements connected with grey lines. The geometric mean titers (GMT) of binding antibody responses per group is indicated with the red line. The dotted lines indicate the lower limit of detection (LLOD) and lower limit of quantification (LLOQ). B) SARS-CoV-2 spike protein neutralizing antibody titers were measured over time with a psVNA qualified for human samples, using pseudotyped virus particles made from a modified Vesicular Stomatitis Virus (VSVΔG) backbone and bearing the S glycoprotein of SARS-CoV-2. Neutralizing antibody responses are measured as the reciprocal of the sample dilution where 50% neutralization is achieved (IC50). Individual animal levels are depicted with grey points and paired measurements connected with grey lines. The GMT of neutralizing antibody responses per group is indicated with the red line. The dotted lines indicate the LLOD and LLOQ. C) Correlation between S-specific binding antibody concentrations and neutralizing antibody titers per animal for all groups and timepoints except the sham control group and week 0 (baseline). The dotted lines indicate the LLOD for each assay.
A second vaccine dose, given at 4 or 8 weeks post the first vaccination, elicited a significant increase in spike protein-specific antibody responses (p≤0.001, ANOVA t-test) (Figure 1A and 1B) relative to the pre dose-2 timepoint. Compared to the one-dose regimen with 5×1010 vp Ad26.COV2.S, a second immunization given at 4 or 8 weeks post first dose, elicited a 5.7- and 11.8-fold increase (p<0.001, ANOVA t-test) of binding antibody concentrations, and a 7.6- and 15.2-fold increase (p<0.001, ANOVA t-test) of neutralizing antibody titers, respectively, as measured 2 weeks post dose-2. Similar differences were observed when comparing the antibody responses elicited by the two-dose vaccine regimens, to those elicited by the single 1 x 1011 vp vaccine dose.
While the two-dose vaccine regimens with 4- and 8-week interval elicited comparable spike protein-specific binding antibody concentrations two weeks post second immunization (p=0.456, ANOVA t-test) (Figure 1A), the geometric mean of neutralizing antibody titers was 2.2-fold higher (p=0.005, ANOVA with t-test) for the 8-week compared to the 4-week regimen (Figure 1 B). At week 4 and week 6 post second immunization, binding and neutralizing antibody levels declined in both two-dose groups with similar kinetics, maintaining the relative difference in neutralizing antibody titers (2.1- and 2.4-fold higher at 4-and 6-weeks respectively for the 8-week regimen, p=0.021 and p=0.001, respectively, ANOVA t-test).
In spite of the more rapid decline of binding antibody concentrations relative to neutralizing antibody titers in animals that received a one-dose regimen, we observed a good overall correlation between binding and neutralizing antibody levels across timepoints for all tested regimens (R =0.7875, p<0.001, Spearman rank-correlation test) (Figure 1C).
Immunogenicity of one- and two-dose Ad26.COV2.S vaccine regimes in aged rhesus macaques
As COVID-19 severity and mortality is increasing with age, we also analyzed the immunogenicity of Ad26.COV2.S in aged rhesus macaques (20 females, 13.75 - 21.9 years old). An aluminum hydroxide (Al(OH)3) adjuvanted soluble trimeric spike protein stabilized in its prefusion conformation was included as a T helper 2 (Th2) skewing control vaccine. Groups were immunized with a one-dose regimen of 1×1011 vp Ad26.COV2.S (n=6), a two-dose regimen with 5×1010 vp Ad26.COV2.S (n=6) or a two-dose regimen with Al(OH)3-adjuvanted 100 μg spike protein (n=4), 8 weeks apart. A sham control group received an Ad26 vector encoding an irrelevant antigen (Ad26.RSV.gLuc; sham control; n=4) at week 0 and week 8. SARS-CoV-2 spike protein-specific binding and neutralizing antibody levels were measured every two weeks up to 10 weeks post the first immunization and spike protein-specific cellular responses were measured at 4 and 10 weeks.
Spike protein-specific binding antibody concentrations significantly increased for each vaccination regimen from week 2 onwards (p≤0.034, ANOVA paired t-test comparing week 0 versus week 2). At weeks 6 and 8 the Ad26.COV2.S induced antibody concentrations were significantly increased compared to Al(OH)3-adjuvanted spike protein induced concentrations (p≤0.036, ANOVA t-test). No statistically significant differences in antibody responses elicited by the two Ad26.COV2.S dose levels could be detected up to week 8. At week 10, 2 weeks after the second dose, the groups that received a second dose of 5×1010 vp Ad26.COV2.S or Al(OH)3-adjuvanted spike protein had significantly higher antibody concentrations compared to recipients of the single dose 1×1011 vp Ad26.COV2.S (4.4-fold and 5.9 fold for the 5×1010 vp Ad26.COV2.S group and Al(OH)3-adjuvanted spike protein group respectively, p≤0.002, ANOVA t-test). Spike-specific antibody concentrations between the two dose regimens were not significantly different (p=0.482) (Figure 2A).


A) SARS-CoV-2 spike protein binding antibody concentrations were measured over time with an ELISA qualified for human samples, using a trimeric, soluble stabilized spike protein produced in mammalian cells as coating antigen. Individual animal levels are depicted with grey points and paired measurements connected with grey lines. The geometric mean titer (GMT) of binding antibody responses per group is indicated with the red line. The dotted lines indicate the lower limit of detection (LLOD) and lower limit of quantification (LLOQ). B) SARS-CoV-2 neutralization antibody titers over time, as measured by wtVNA. Individual animal levels are depicted with grey points and paired measurements connected with grey lines. The GMT per group is indicated with the red line. The dotted line indicates the LLOD. C) Correlation between S-specific binding antibody concentrations and neutralizing antibody titers per animal for all groups and timepoints except the sham control group and week 0. The dotted lines indicate the LLOD for each assay.
Kinetics of neutralizing antibody responses were determined by a wild-type virus neutralization assay. A single dose of 1×1011 vp Ad26.COV2.S induced neutralizing antibody titers at week 2 which were significantly increased at week 4 in all animals compared to the previous timepoint (p=0.031, sign test), and remained stable thereafter up to week 10. Similarly, the two-dose 5×1010 vp Ad26.COV2.S regimen induced neutralizing antibody titers that significantly increased at week 4 (p=0.031, sign test) and 6 (p=0.008, Tobit ANOVA z-test) compared to previous timepoints. At week 10, 2 weeks after the second dose, antibody titers were increased 8-fold compared to week 8 (p<0.001, Tobit ANOVA z-test). Al(OH)3-adjuvanted spike protein induced only low and transient levels of neutralizing antibodies after the first dose in 2 out of 4 animals only. At week 10 however, 2 weeks after the second dose, all 4 animals had neutralizing antibody titers in the same range as the Ad26.COV2.S groups (no statistical analysis possible due to small group size of the adjuvanted protein group). Pairwise comparison of vaccine groups at week 10 showed that the two dose 5×1010 vp Ad26.COV2.S regimens or Al(OH)3-adjuvanted spike protein induced significantly higher neutralizing antibody titers compared to the single dose 1×1011 vp Ad26.COV2.S group (10- and 5.5-fold for 5×1010 vp Ad26.COV2.S and Al(OH)3-adjuvanted spike protein group, respectively, p≤0.004, Tobit ANOVA z-test). Neutralizing antibody titers between the two dose regimens were not significantly different (p=0.303) (Figure 2B). The spike protein-specific binding antibody concentrations as measured with ELISA strongly correlated with neutralizing antibody titers (R=0.92, p=<0.001, Spearman rank correlation) showing a higher sensitivity of the ELISA (Figure 3C).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
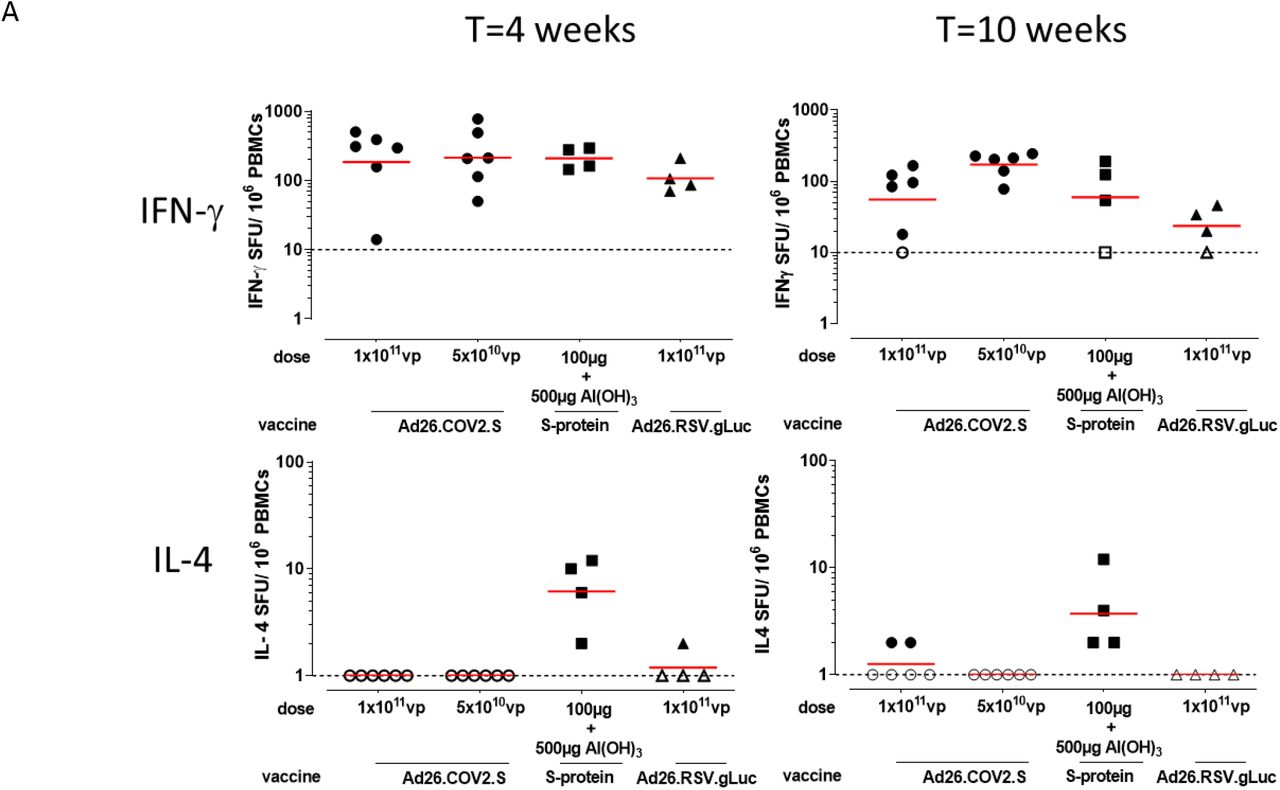
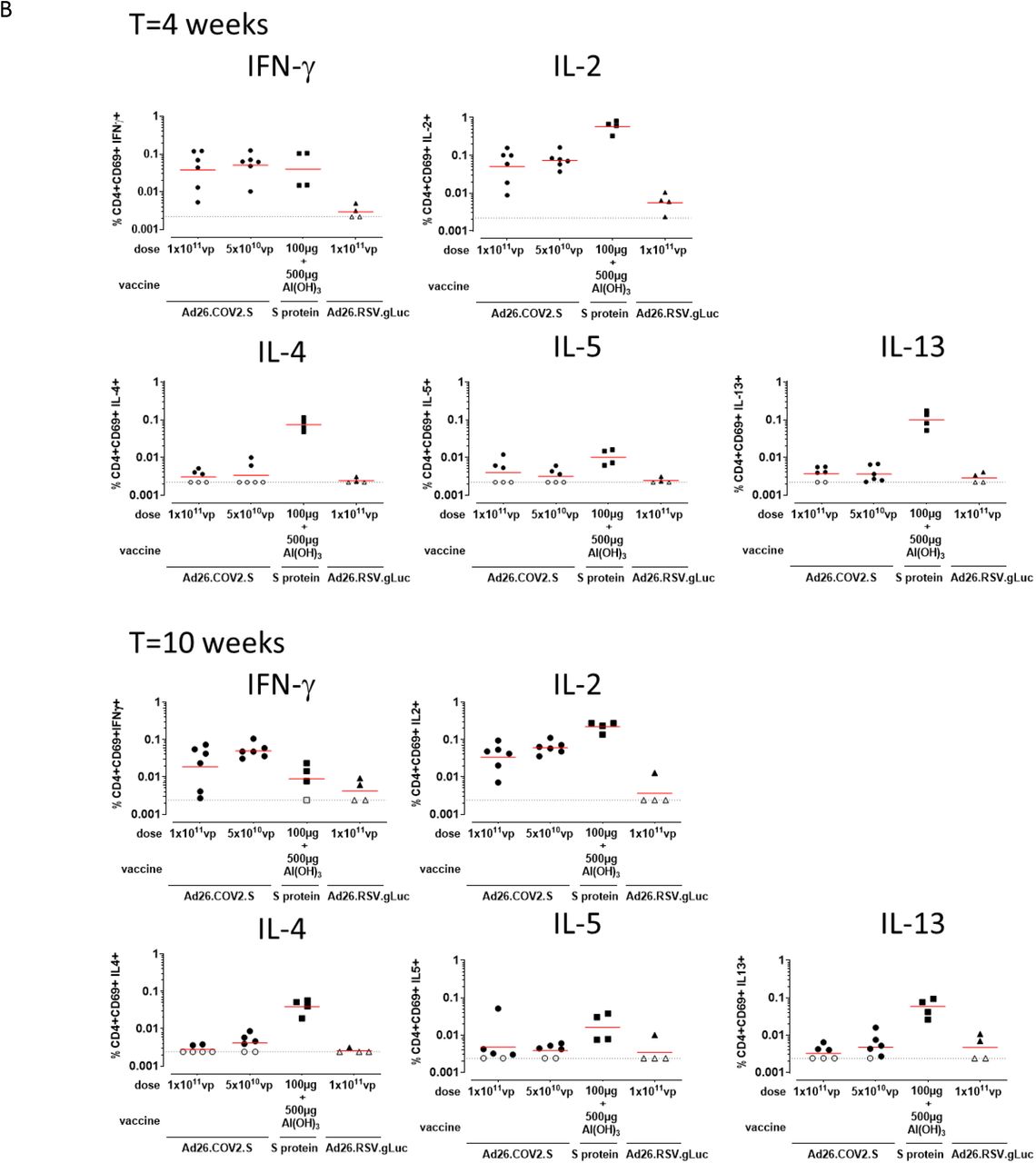
A) Spike protein-specific T cell responses as measured with an IFN-γ/ IL-4 Double-color ELISpot at indicated timepoints. The geometric mean titer (GMT) response per group is indicated with a horizontal line. Samples with background subtracted counts below or equal to 0 were set a 10 and 1. for IFN-γ and IL-4 respectively for visualization purposes and indicated by open symbols and the dotted line. B) Spike protein-specific T cell responses as measured by intracellular cytokine staining at indicated timepoints. Frequency of CD4+CD69+ T cell expressing cytokines. Gating strategy is provided in supplemental figure 2. The geometric mean response per group is indicated with a horizontal line. The dotted line indicates the technical threshold. Open symbols denote samples at technical threshold.
Spike protein-specific T cell responses were measured with ELISpot and intracellular cytokine staining (ICS) using peripheral blood mononuclear cell (PBMC) stimulated with 15-mer peptides overlapping by 11 amino acids and spanning the complete SARS-CoV-2 spike protein. Both Ad26.COV2.S regimens as well as Al(OH)3-adjuvanted spike protein induced IFN-γ responses at 4 weeks after the first immunization (Figure 3A and 3B). At week 10, IFN-γ responses were lower for the 1×1011 vp Ad26.COV2.S and adjuvanted spike protein groups compared to week 4. In animals vaccinated with the two dose 5×1010 vp Ad26.COV2.S regimen IFN-γ responses were comparable to the week 4 time point at week 10, suggesting that a second dose of Ad26.COV2.S maintains spike-specific T cell responses.
Substantial IL-4 responses were observed only for the Al(OH)3-adjuvanted spike protein group at both week 4 and week 10 by ELISpot (Figure 3A). CD4+ and CD8+ T cell cytokine responses were also analyzed by ICS. Spike protein-specific CD4+ T cell IFN-γ and IL-4 expression patterns confirmed the ELISpot responses, with all vaccine groups inducing significantly higher levels of IFN-γ compared to the sham control group at week 4 (p≤0.029, Mann-Whitney-U test) that decreased at week 10, except for the two dose 5×1010 vp Ad26.COV2.S group, which remained significantly higher compared to the sham group (p=0.010, Mann-Whitney-U test). IL-4 expression was only significantly higher for the Al(OH)3-adjuvanted spike protein group compared to the sham control group at both timepoints (p=0.029 Mann-Whitney-U test) (Figure 3B). Additional cytokines IL-2, or IL-5 and IL-13 were measured and reflected the patterns as seen for IFN-γ or IL-4, respectively, with minimal or no boosting of responses for groups that received a second dose, and only Al(OH)3-adjuvanted spike protein inducing robust levels of Th2 cytokines IL-4, IL-5 and IL-13 (Figure 3B). Spike protein-specific CD8+ T cells induced by Ad26.COV2.S mainly produced IFN-γ and IL-2, while Al(OH)3-adjuvanted spike protein only produced IL-2. None of the immunization regimens induced CD8+ T cells producing significant amounts of IL-4, IL-5 or IL-13 (Supplementary figure 1).
Discussion
We here evaluated the immunogenicity of one- and two-dose Ad26.COV2.S regimens in adult and aged rhesus macaques for up to 14 weeks after the first dose, to gain insight both in the durability of immunity after a single dose of the Ad26.COV2.S vaccine candidate and in the impact of a second dose on the magnitude of spike protein-specific immune responses.
In both adult and aged macaques, spike protein-binding and SARS-CoV-2 neutralizing antibody responses were detected as early as two weeks after the first Ad26.COV2.S immunization and had significantly increased by week 4. The kinetics and magnitude of antibody responses appeared similar in adult and aged rhesus macaques for all Ad26.COV2.S vaccine regimens tested. These observations are in agreement with our observations in human adults and elderly at week 4 post single immunization with Ad26.COV2.S (Sadoff et al., 2020). On the contrary, Al(OH)3-adjuvanted spike protein in NHP only induced low and transient levels of neutralizing antibodies after the first dose and required 2 doses to elicit a robust neutralizing antibody response, representing a limitation for a vaccine intended to provide early onset of protection given the ongoing COVID-19 pandemic.
In adult macaques, a single dose of Ad26.COV2.S elicited humoral immune responses, which were maintained at least up to week 14 post-immunization. Binding antibody responses did show some decline over time, while neutralizing antibody responses were more stably maintained, providing an early sign of durable immunity elicited by the Ad26.COV2.S vaccine candidate. Although humoral immune responses were significantly higher in NHP that received the 1×1011 vp dose as compared to recipients of the 5×1010 vp vaccine dose, differences in neutralizing antibody levels decreased over time and do not suggest a clear benefit of the higher dose, in agreement with interim Phase 1/2a clinical data (Sadoff et al., 2020).
A second dose of Ad26.COV2.S given at 8 weeks post the first immunization resulted in a significant increase in spike protein-specific binding and more importantly neutralizing antibody responses, in both adult and aged NHP compared to the one-dose regimen. This is in line with our observations with other Ad26-based vaccines, where a second dose always elicited a higher and more durable immune response in both animal models and humans (Geisbert et al., 2011)(Callendret et al., 2018) (Salisch et al., 2019)(Baden et al., 2013) (Salisch and Stephenson et al, manuscript submitted). Our findings with the two-dose Ad26.COV2.S regimen in NHP, may suggest that a two-dose Ad26.COV2.S vaccine regimen will also result in higher and more durable immune responses in humans, as compared to our one-dose regimen.
The evaluation of the potential impact of length of interval between two vaccine doses in adult NHP demonstrated that the neutralizing antibody responses were higher in animals that received the two doses with an 8-week interval as compared to the 4-week interval, albeit that both two-dose regimens were more immunogenic than one-dose. This confirms that the interval between vaccine doses can significantly affect the magnitude and/or quality of the antibody response, with longer intervals between vaccine doses associating with improved antibody responses (Ledgerwood et al., 2013) (Siegrist, 2018) (Sallusto et al., 2010) (Roozendaal et al., 2020, in press). Thus, from an immunogenicity point of view, a vaccine regimen with an 8-week interval between 2 doses is preferred over a regimen with a 4-week interval. A regimen with two-doses of 5×1010 vp Ad26.COV2.S with an 8-week interval is planned to be tested for efficacy in humans (ENSEMBLE 2, NCT04614948).
In both adult and aged NHP studies we found a high correlation between binding and neutralizing antibody responses, although different neutralization assays were used in the two studies. This suggests that spike protein binding antibody concentrations measured by ELISA could be used as a surrogate readout for neutralizing antibody responses, which is also supported by our earlier observations (Yu et al., 2020)(Mercado et al., 2020).
An important aspect to be evaluated when developing a COVID-19 vaccine is the potential and theoretical risk of Vaccine-Associated Enhanced Respiratory Disease (VAERD)(Lee et al., 2020)(Bottazzi et al., 2020) (Haynes et al., 2020), which is generally considered to be associated with non-neutralizing antibody responses and Th2-skewed cellular immune responses. In this study we show that in aged NHP Ad26.COV2.S elicited CD4+ T cell responses that were Th1 skewed, as previously shown in both NHP as well as in humans (Mercado et al., 2020)(Sadoff et al., 2020). This is similar to what has been found for other genetic vaccine platforms encoding SARS-CoV-2 spike protein in both NHP and humans (van Doremalen et al., 2020) (Yu et al., 2020)(Anderson et al., 2020) (Vogel et al., 2020)(Corbett et al., 2020). In contrast, the Al(OH)3-adjuvanted spike protein induced a more Th2 skewed immune response, as expected with this adjuvant, and confirming that a Th2 skewed response can be elicited in this NHP animal model. The Th1-skewed response in NHP together with the induction of robust and durable neutralizing antibody responses by Ad26.COV2.S, reduce the likelihood of VAERD for this vaccine.
In summary, our data show that a one- and two-dose Ad26.COV2.S vaccine regimens elicit similar antibody responses in adult and aged NHP. Importantly, a second vaccine dose administered 8 weeks post the first immunization, induced a significant increase in (neutralizing) antibody responses in both adult and aged animals compared to a single vaccine dose. In addition, antibody responses in adult macaques were maintained up to 14 weeks post first immunization, providing an early indication of durability of immune responses elicited by a single dose Ad26.COV2.S. Overall, these data support our decision to evaluate a single 5×1010 vp dose of Ad26.COV2.S in our Phase 3 ENSEMBLE study and also to initiate our second Phase 3 study ENSEMBLE 2, to evaluate the protective efficacy of a two dose Ad26.COV2.S regimen.
Author Contributions
Designed studies and reviewed data: L.S., H.K., S.K.R.H., J.E.M.vd.L., L.D., R.R., F.W., R.C.Z. Performed experiments and analyzed data: D.N.C.C., A.I.G., M.R.M.B., J.D., J.V., E.v.H., Y.C, J.V., T.J.D., S.K.M., M.K., E.J.S., G.K., P.M., W.M.J.M.B., L.M., J.T.B.M.T. Drafted the paper: L.S., H. K. Reviewed the paper: all authors.
Competing interests
The authors declare no competing financial interests. L.S., H.K., S.K.R.H., J.E.M.vd.L., L.D., D.N.C.C, A.I.G, M.R.M.B., J.D., J.V., E.v.H., Y.C., J.V., L.M., J.T.B.M.T., R.R., H.S, F.W. and R.C.Z. are employees of Janssen Vaccines & Prevention and may be Johnson & Johnson stockholders.
Methods
Animals
Adult NHP
The NHP study including adult animals was conducted at Charles River Laboratories (CRL) Montreal ULC, Laval Site (CA). Animals were obtained from Kunmings Biomed international Ltd, China. Prior to transfer from test facility colony, all animals were subjected to a health assessment and tested at least once for tuberculosis by intradermal injection of tuberculin. An anthelmintic treatment was administered to each animal by subcutaneous injection. The evaluations were performed in accordance with the standard operating procedures by technical staff. Animal experiment approval was provided by the Institutional Animal Care and Use Committee (IACUC) at CRL Montreal ULC, Laval Site (CA). Animal experiments were performed in compliance with Guidelines published by the Canadian Council on Animal Care and the Guide for the Care and Use of Laboratory Animals published by the National Research Council Canada. The Test Facility is accredited by the Canadian Council on Animal Care (CCAC) and the American Association for Accreditation of Laboratory Animal Care (AAALAC). In addition, the study was conducted according to EMA guideline, ICH M3(R2): Guidance on Non-Clinical Safety Studies for the Conduct of Human Clinical Trials and Marketing Authorization for Pharmaceuticals and FDA guideline, Redbook 2000: General Guidelines for Designing and Conducting Toxicity Studies.
Aged NHP
The study using aged NHP was performed at the Biomedical Primate Research Center, Rijswijk, The Netherlands (an AAALAC-accredited institution). Animals were captive-bred for research purposes and socially housed. Animal housing was according to international guidelines for non-human primate care and use (The European Council Directive 86/609/EEC, and Convention ETS 123, including the revised Appendix A as well the ‘Standard for humane care and use of Laboratory Animals by Foreign institutions’ identification number A5539-01, provided by the Department of Health and Human Services of the United States of America’s National Institutes of Health (NIH)). All animal handlings were performed within the Department of Animal Science (ASD) according to Dutch law. A large, experienced staff is available, including full-time veterinarians and a pathologist. ASD is regularly inspected by the responsible authority (Voedsel en Waren Autoriteit, VWA), and by an independent Animal Welfare Officer. Some animals were seropositive for antibodies to Simian Herpes B Virus, while some animals of the mock-immunized control group were positive for antibodies to Simian T-cell Leukemia Virus and Simian Retro Virus. All animals were classified healthy according to physical examination and evaluation of complete blood count and serum chemistry. The Institutional Animal Care and Use Committee of the Biomedical Primate Research Centre (dierexperimentencommissie, DEC-BPRC), approved the study protocols developed according to strict international ethical and scientific standards and guidelines. The qualification of the members of this committee, including their independence from a research institute, is requested in the Dutch law on animal experiments.
Vaccines
The Ad26.COV2.S vaccine has been generated as previously described (Bos et al., 2020). Briefly, Ad26.COV2.S is a replication incompetent adenovirus serotype 26 (Ad26) vector encoding a prefusion stabilized SARS-COV-2 spike protein sequence (Wuhan Hu1; GenBank accession number: MN908947Replication-incompetent, E1/E3-deleted Ad26-vectors were engineered using the AdVac system (Abbink et al., 2007), using a single plasmid technology containing the Ad26 vector genome including a transgene expression cassette. The codon optimized, prefusion stabilized, SARS-COV-2 spike protein encoding gene was inserted into the E1-position of the Ad26 vector genome. Manufacturing of the Ad26 vector was performed in the complementing cell line PER.C6 TetR (Wunderlich et al., 2018) (Zahn et al., 2012). The negative control vector Ad26.RSV.gLuc encodes the RSV F protein fused to Gaussia firefly luciferase as a single transgene separated by a 2A peptide sequence, resulting in expression of both individual proteins. Manufacturing of the vector was performed in Per.C6 (Sanders et al., 2013).
The full-length spike protein used for immunization (COR200099) (Bos et al., 2020) was produced on Expi293F cells. COR200099 is based on the Wuhan-Hu-1 SARS-CoV-2 strain (MN908947) and stabilized by two point mutations (R682A, R685G) in the S1/S2 junction that knocks out the furin cleavage site, and by two consecutive prolines (K986P, V987P) in the hinge region in S2. In addition, the transmembrane and cytoplasmic regions have been replaced by a fibritin foldon domain for trimerization and a C-tag, allowing the protein to be produced and purified as soluble protein. Adenoviral vectors and protein were tested for bioburden and endotoxin levels prior to use.
Study design animal experiments
Adult NHP
60 (57 females and 3 males. 3 males were allocated to test groups 3, 4 and 5, 1 male in each group) rhesus macaques (Macaca Mulatta) from Chinese origin between 3.3 to 5.0 years old were assigned to five groups by a randomizing stratification system based on body weights, fourteen animals were included in each vaccine group and four animals were included in the sham control group. Group 1 (n=4) is the sham control group and received saline injection at week 0 and week 8, group 2 and 3 (n=14 each group) received one immunization with 1×1011 viral particles (vp) and 5×1010 vp of Ad26. COV2.S, respectively, at week 0, group 4 and 5 (n=14 each group) received two immunizations with 5×1010 vp of Ad26. COV.2 spaced by four (week 0 and week 4) and eight weeks (week 0 and week 8), respectively. All immunizations were performed via the intramuscular route in the quadriceps muscle of the left hind leg. Blood for serum was obtained prior to the first vaccine dose and every 2 weeks subsequently up to week 14 of the study.
Aged NHP
20 female rhesus macaques (Macaca Mulatta), aged between 13.75 and 21.9 years, were distributed over 4 experimental treatment groups and housed in ABSL-III facilities, pair-housed with socially compatible animals. Group 1 (n=6) received 1×1011 viral particle (vp) of Ad26. COV2.S at week 0. Group 2 (n=6) received 5×1010 vp of Ad26. COV2.S at week 0 and 8. Group 3 (n=4) received 100 μg spike protein, adjuvanted with 500 μg Aluminum Hydroxide (Al(OH)3; 2% Alhydrogel, InvivoGen) at week 0 an 8. The sham control group (Group 4, n=4) was immunized with 1×1011 vp Ad26.RSV.gLuc, an Ad26 vector expressing an irrelevant antigen. All immunizations were performed intramuscularly in quadriceps of the left hind leg. Blood for serum and peripheral blood mononuclear cells (PBMC) isolation was obtained as indicated in the text.
Enzyme-linked immunosorbent assay (ELISA)
IgG binding to SARS-CoV-2 spike protein was measured by ELISA using a recombinant spike protein antigen based on the Wuhan-Hu-1 SARS-CoV-2 strain (MN908947). The SARS-CoV-2 spike protein antigen was adsorbed on 96 well microplates for a minimum of 16 hours at 4°C. Following incubation, plates were washed in PBS/0.05% Tween-20 and blocked with 5% skim milk in PBS/0.05% Tween-20 for 1 hour at room temperature. Serum standards, controls and NHP serum samples were diluted and incubated on the plates for 1 hour at room temperature. The plates were washed and then incubated with peroxidase conjugated goat anti human IgG for 1 hour at room temperature, washed, and developed with tetramethylbenzidine (TMB) substrate for 30 minutes at room temperature and protected from light, then stopped with H2SO4. The optical density was read at 450/620 nm. The antibody concentrations were back calculated on the standard and the reportable value were generated based on all passing dilutions, expressed in ELISA units [EU]/mL. The lower limit of detection (LLOD) is 3.4 EU/mL, based on the standard lowest interpolation range concentration multiplied per the dilution factor and is used as an informative LLOD. The lower limit of quantification (LLOQ) is based on qualification performed for human samples and has been set on 50.3 EU/mL.
Pseudovirus neutralization assay (psVNA)
SARS-CoV-2 S neutralizing antibody titers were measured by pseudovirus neutralizing assay. Pseudotyped virus particles were made from a modified Vesicular Stomatitis Virus (VSVΔG) backbone and bear the S glycoprotein of the SARS-CoV-2. The pseudoparticles contain a Luciferase reporter used for detection. Serial dilutions of heat-inactivated NHP serum samples were prepared in 96-well transfer plates. The SARS-CoV-2 pseudovirus was added sequentially to the serum dilutions and incubated at 37°C with 5% CO2 supplementation for 60 ± 5 minutes. Serum-virus complexes were then transferred onto plates, previously seeded overnight with Vero E6 cells, and incubated at 37°C and 5% CO2 for 20 ± 2 hours. Following this incubation, the luciferase substrate was added to the cells in order to assess the level of luminescence per well. The plates were then read on a luminescence plate reader. The intensity of the luminescence was quantified in relative luminescence units (RLU). The neutralizing titer of a serum sample was calculated as the reciprocal serum dilution corresponding to the 50% neutralization antibody titer (IC50) for that sample. The LLOD is 10, which is the first sample dilution (1:10) used as an informative LLOD. LLOQ is based on qualification performed for human samples has been set on 33 IC50.
Wild type virus neutralization assay (wtVNA)
Neutralization assays against live SARS-CoV-2 were performed using the microneutralization assay as previously described (Bos et al., 2020), with the modification of a different strain used. Clinical isolate SARS-CoV-2/human/NLD/Leiden-0008/2020 (Leiden-0008) was isolated from a RT-PCR positive throat swab and passaged twice in Vero E6 cells. The NGS-derived complete genome of this virus isolate is available under GenBank accession number MT705206.1. Isolate Leiden-0008 was propagated and titrated in Vero E6 cells.
ELISpot
IFN-γ /IL-4 Double-Color was performed on freshly isolated PBMCs. PBMC were isolated from ethylene diamine tetraaceticacid (EDTA) whole blood using Ficoll gradient centrifugation (10ml 92% Ficoll-Paque (GE Healthcare) Plus in 1:4 DPBS-diluted blood) The ELISpot was performed using the ImmunoSpot Human IFN-γ/IL-4 Double-Color Enzymatic ELISpot Assay Kit according to the manufacturer’s protocol (Cellular Technology Limited). Ethanol-activated 96-well ELISpot plates were coated overnight with anti-human IFN-γ and IL-4 capture antibodies. Cells were plated at a concentration of 250,000 cells per well and stimulated with either cell culture medium in presence of DMSO, 2 pools of consecutive 15 mer peptides with 11 amino acid overlap (JPT) spanning the entire length of the SARS-CoV-2 spike protein at a peptide concentration of 2 μg/mL, or 1 μg/mL PHA as positive control for 22 hours. Analysis was performed using the CTL ImmunoSpot Analyzer and ImmunoSpot Software (Cellular Technology). Spot-forming units per 1.0 × 106 PBMCs were calculated by subtraction of medium stimulus counts of the individual peptide pools per animal and summed across the 2 peptide pools.
Intracellular cytokine staining (ICS)
For analysis of intracellular cytokine production, 1×106 freshly isolated PBMC were stimulated at 37 °C overnight (approximately 15 hours) with either cell culture medium in presence of DMSO, 2 μg/mL SARS-CoV-2 spike protein peptide pools (as described for ELISpot), or 5 μg/mL PHA in the presence of GolgiStop (BD Biosciences). Stimulated cells were first incubated with LIVE/DEAD Aqua viability dye (Thermo Fisher Scientific), followed by surface staining with anti-human monoclonal antibodies CD3-PerCP-Cy5.5, CD4-APC H7, CD8-BV650, CD14-BV605, CD69-BV786 (BD Biosciences) and CD20-BV605 (Biolegend). Cells were subsequently fixed with Cytofix/Cytoperm buffer (BD Biosciences) and stained intracellularly with anti-human IL-2-PE, IFN-γ-APC (BD Biosciences), IL-5-Vio515 (Miltenyi Biotec), IL-4-PE Dazzle594 and IL-13-BV421 (Biolegend). Sample acquisition was performed on a LSR Fortessa (BD Biosciences) and data were analysed in FlowJo V10 (TreeStar). Antigen-specific T cells were identified by consecutive gating on single cells (FSC-H versus FSC-A), live cells, size (lymphocytes) (FSC-A versus SSC-A), CD3+, CD4+ or CD8+ cells and CD69+ plus cytokine-positive (the gating strategy is shown in Supplementary Fig. 2). Cytokine-positive responses are presented after subtraction of the background response detected in the corresponding medium stimulated sample of each individual animal. Responders were defined by a technical threshold (Bowyer et al., 2018), the theoretical ability to detect at least 1 event in a cytokine gate and here defined as the reciprocal of the average number of CD4 or CD8 T cells of the medium and peptide pool stimulated samples for each assay run. Ratio of Th1 versus Th2 cytokines were calculated by Boolean gating of Th1 (CD4+CD69+ T cells expressing IFN-γ or IL-2) and Th2 (CD4+CD69+ T cells expressing IL-4 or IL-5 or IL-13) subsets.
Statistical analysis
ELISA and psVNA
For binding and psVNA neutralizing antibody data, comparisons between specific vaccine groups were made with the two-sample t-test in an analysis-of-variance (ANOVA). Successive time points have been compared with the paired t-test per vaccine group. P values were calculated on f log10 transformed values.
wtVNA, ELISpot and ICS
Vaccine groups were compared to the negative control group with the Mann-Whitney U-test. Pairwise comparison between vaccine groups was performed using Tobit ANOVA with vaccine as factor if less than 50% of the titers are at LLOD. The pairwise comparisons between vaccines were done with the z-test. If for an assay any vaccine group had 50% censoring or more, then the pairwise comparisons were done with the Mann-Whitney U-test.
The difference in titer between consecutive time points was calculated per animal for each assay.
Depending on the number of censored measurements, the differences were compared with a Tobit ANOVA followed by a post-hoc z test or a sign test.
For all statistical tests the significance level was 5%. No multiple comparison adjustment was applied. All statistical calculations are done in SAS 9.4. (SAS Institute Inc Cary, NC, US).
Correlation analysis
Correlation analysis between binding antibody concentrations and neutralizing antibody titers measured was calculated using two-sided Spearman rank-correlation.
Acknowledgements
This project was funded in part by the Department of Health and Human Services Biomedical Advanced Research and Development Authority (BARDA) under contract HHS0100201700018C. We thank Johan Verspuij for assistance with data processing, Sarah Janssen for ad-hoc statistical support, Janssen colleagues of the Vector Generations and Sub Unit Vaccine Design departments for providing reagents. We thank Gert Scheper, Danielle van Manen and Martin Friedrich Ryser for reviewing the paper and for providing valuable input. We thank Daniella Mortier from the Biomedical Primate Research Centre for logistical and biotechnical support. We thank the team of Charles River Laboratories Montreal ULC, Laval Site (CR-LAV) (CA) and Reno NV Site (US) for their accurate and punctual work on the NHP study with adult macaques, in particular we thank Anne Marie Downey, Roula Salame, Carolyne Dumont, Rajen Patel and Sunjay Sethi. We thank the Nexelis Team of Laval site (CA) for their speed and flexibility in accommodating S ELISA and psVNA sample analysis within short timelines, in particular we thank Luc Gagnon, Helen Diamantakis, Mary Osei-Twum, Greg Kulnis, Steven-Phay Tran, Julien St-Jean, Marcel Dupelle and Akeel Baig.
References




