

Abstract
Wild relatives of domesticated Capsicum represent substantial genetic diversity and thus sources of traits of potential interest. Furthermore, the hybridization compatibility between members of Capsicum species complexes remains unresolved. Improving our understanding of the relationship between Capsicum species relatedness and their ability to form hybrids is a highly pertinent issue. Through the development of novel interspecific hybrids in this study, we demonstrate interspecies compatibility is not necessarily reflected in relatedness according to established Capsicum genepool complexes. Based on a phylogeny constructed by genotyping using single sequence repeat (SSR) markers and with a portion of the waxy locus, and through principal component analysis (PCA) of phenotypic data, we clarify the relationships among wild and domesticated Capsicum species. Together, the phylogeny and hybridization studies provide evidence for the misidentification of a number of species from the World Vegetable Center genebank included in this study. The World Vegetable Center holds the largest collection of Capsicum genetic material globally, therefore this may reflect a wider issue in the misidentification of Capsicum wild relatives. The findings presented here provide insight into an apparent disconnect between compatibility and relatedness in the Capsicum genus, which will be valuable in identifying candidates for future breeding programs.
The genus Capsicum is comprised of about 35 species including five domesticated species: C. annuum (L.), C. baccatum (L.), C. chinense (Jacq.), C. frutescens (L.), and C. pubescens (Ruiz & Pav.) (Khoury et al., 2020). The diversity of Capsicum species represents a valuable genetic resource for crop improvement (Barchenger and Bosland, 2019). The primary limitations to improving productivity and quality of Capsicum are abiotic and biotic stresses, many of which lack sources of host tolerance or resistance (Barchenger et al., 2019). Furthermore, as a widely consumed crop with cultural and culinary value across global cuisines, there is high demand for Capsicum (Bosland and Votava, 2012). There is therefore significant incentive to overcome challenges to cultivation, and one means of doing so being the introgression of resistance to the various stresses that limit production of Capsicum species.
Understanding interspecies compatibility and identifying barriers to hybridization is essential to the design of introgression breeding programs. Capsicum species are divided among 11 clades (Bosland and Votava, 2012; Carrizo García et al., 2016) and grouped into three complexes - Annuum, Baccatum and Pubescens - based on their relative reproductive compatibility (Tong and Bosland, 1999; Pickersgill, 1971; Emboden Jr., 1962). There is understood to be relatively low reproductive compatibility between species complexes (van Zonneveld et al., 2015). However, a number of cross-complex hybridizations have been achieved (Eggink et al., 2014; Costa et al., 2009; Kamvorn et al., 2014; Yoon et al., 2006; OECD, 2006; Pickersgill, 1991), and the pre- and post-zygotic barriers to hybridization between genetic complexes are not fully understood (Yoon et al., 2006). This suggests isolation between complexes is not total, and there is therefore potential for introgression breeding, or design of genetic bridge strategies in order to best exploit this genetic variation.
In contrast to other Solanaceae crops, including tomato (Solanum lycopersicum) (Lin et al., 2014), potato (S. tuberosum) (Hirsch et al., 2013) and to a lesser extent eggplant (S. melongena) (Gramazio et al., 2017) introgression breeding using wild species has been relatively underutilized in Capsicum; (Mongkolporn and Taylor, 2011). The wild progenitor, C. annuum var. glabriusculum is a potential source of disease resistance, with reported resistance to Beet curly top virus (BCTV: Curtovirus) (Bosland, 2000; Jimenez, 2019). Members of the wild species C. chacoense and C. rhomboideum have been identified as being resistant to powdery mildew (Leveillula taurica) (McCoy and Bosland, 2019). Recently, an accession of C. galapagoense has been proposed to be a potential source of resistance to the insect pest, whitefly, based on trichome density and type (M. Rhaka, pers. comm.). However, despite extensive hybridization no successful progeny have so far been developed (Lin et al., 2020). These results are surprising, because C. galapagoense has been reported as part of the C. annuum clade, and readily hybridize with C. annuum accessions (Carrizo García et al., 2016; Pickersgill, 1971). One reason for unsuccessful hybridization attempts may be misidentification; several genebanks have incorrectly reported accessions identified as C. galapagoense which are, in fact, C. frutescens (P.W. Bosland, pers. comm.). Such misidentification presents a challenge to utilizing knowledge of the relatedness of Capsicum species and their ability to hybridize. Although the genetic diversity and variation within wild populations of Capsicum has been studied (Carrizo García et al., 2016; Cheng et al., 2016; Aguilar Meléndez et al., 2009; Oyama et al., 2006; Votava et al., 2002; Loaiza-Figueroa et al., 1989), the pool of phenotypic data for wild Capsicum species remains limited (Barchenger and Bosland, 2019). There also remains a lack of access to publicly available germplasm representing the diversity of wild Capsicum (Khoury et al., 2020). There is therefore an immediate need to better understand the role of wild Capsicum species in future breeding programs.
The objectives of this study were to elucidate the relationship between interspecies compatibility and relatedness through extensive interspecific hybridization and the construction of a phylogeny. We aimed to clarify the relationships among the wild and domesticated Capsicum species included in the study, and confirm the identities of several World Vegetable Center genebank accessions.
Materials and Methods
Thirty-eight accessions of 15 species of Capsicum were chosen for this experiment (Table 1). The accessions were provided to the World Vegetable Center, having been collected from diverse locations and deposited into collections at either the World Vegetable Center Genebank, the World Vegetable Center Pepper Breeding Collection in Tainan, Taiwan, the United States Department of Agriculture - Agriculture Research Service National Plant Germplasm System, or the Chile Pepper Institute, New Mexico State University, Las Cruces, NM USA. Of each accession, two biological replications were used wherever possible for phenotyping and genotyping, although due to poor germination, four accessions (NMCA50034, PBC556, PBC1892, NMCA50064) did not have a biological replicate.
Capsicum accessions included in this study
All experiments were conducted at the World Vegetable Center, Shanhua, Tainan, Taiwan (lat. 23.1°N; long. 120.3°E; elevation 12 m). Prior to sowing, all seed was treated with trisodium phosphate (TSP) and hydrochloric acid (HCl) following the methods of Kenyon et al. (2017), which has been observed to reduce germination rates. Seeds were sown into 72-cell plastic trays of sterilized peat moss. Trays were placed in a climate-controlled greenhouse for germination at 28 ± 3 °C with a 12-hour photoperiod and ≈95% relative humidity. At the 4-6 true leaf stage, the seedlings were transplanted into pots and moved to a greenhouse without climate control. Plants were irrigated twice daily and regularly fertilized with Nitrophoska (Incitec Pivot Fertilisers, Victoria, Australia) during the experimental period.
The accessions were morphologically characterized according to the Descriptors of Capsicum Manual (IPGRI et al., 1995) for the following characteristics: mature leaf length, mature leaf width at widest point, leaf color, density (if present) of leaf pubescence, leaf shape, lamina margin, stem color, stem shape, density (if present) of stem pubescence, nodal anthocyanin color, node length, anther color, anther length, filament length, corolla color, corolla spot color, corolla shape, corolla length, stigma exsertion, flower position, tillering, leaf density, fruit length, fruit width, fruit pedicel length, neck at base of fruit. Quantitative traits were the mean of 10 values measured across replicates. Qualitative traits were scored according to the IPGRI Descriptors of Capsicum manual based on observations of both plant replicates. Accessions with incomplete data were excluded from analysis of phenotypic data. To identify trends in traits between species, the quantitative traits were used for principal component analysis (PCA) using the R packages, ‘factoextra’ (Kassambara and Mundt, 2020) and ‘ggfortify’ (Horikoshi et al., 2020) for PCA analysis with scaling. The scores of qualitative traits were analyzed using an unweighted pair group method with arithmetic mean (UPGMA) hierarchical cluster analysis. Bootstrap resampling was applied to clustering with 1,000 iterations.
Reciprocal hybridizations were attempted among all combinations of accessions throughout the experimental period. Ability to hybridize in reciprocal was used to confirm previous reports of relatedness and ability to hybridize species across clades and complexes. The fruits of successful hybridizations were collected upon ripening. Within three days of harvest, the seeds were extracted from the fruits and dried for at least 1week. Five seeds each of 112 crosses of interest were sown into 70-cell plastic trays containing sterilized peat moss. The trays were placed in a greenhouse without climate control and irrigated twice daily and observed daily for 12 weeks to assess germination. A chord diagram was produced in R using the package ‘circlize’ (Gu et al., 2014) to visualize successful crosses for which seed was obtained. A heat map was produced in R using the package ‘ggplot2’ (Wickham et al., 2020) to visualise the percentage of seeds germinated after 12 weeks.
For genotyping, DNA was isolated from young, actively growing leaves from plants of each accession using the modified cetrimonium bromide (CTAB) extraction method (Meyer and Buchta, 2020). Using 27 Simple Sequence Repeat (SSR) markers, DNA was amplified by PCR, for which each well of a 96-well microtiter plate contained 2 μl of template DNA, 0.4 μl of primer (0.2 μl each forward and reverse), 0.1 μl of AmpliTaq Gold DNA polymerase, 0.4 μl of deoxyribonucleotides, 1.5 μl of 10× PCR Buffer II Gold buffer (Thermo Fisher Scientific, Waltham, MA, USA), and sterile water to a final volume of 15 μl. The reactions were carried out in a thermal cycler (Single Block Alpha Unit, DNA Engine®, Bio-Rad Laboratories, Berkeley, CA, USA) with an annealing temperature of 55 °C. The electrophoresis of amplified products was performed on 6% acrylamide gels at 160 Volts for 30 minutes (Thermo Electron Electrophoresis EC250-90, Thermo Fisher Scientific). The results were visualized under UV light using UVITEC Imaging Systems (Cleaver Scientific, Warwickshire, UK) following staining with ethidium bromide. Electrophoresis was repeated whenever the clarity of the bands or their exact size was uncertain.
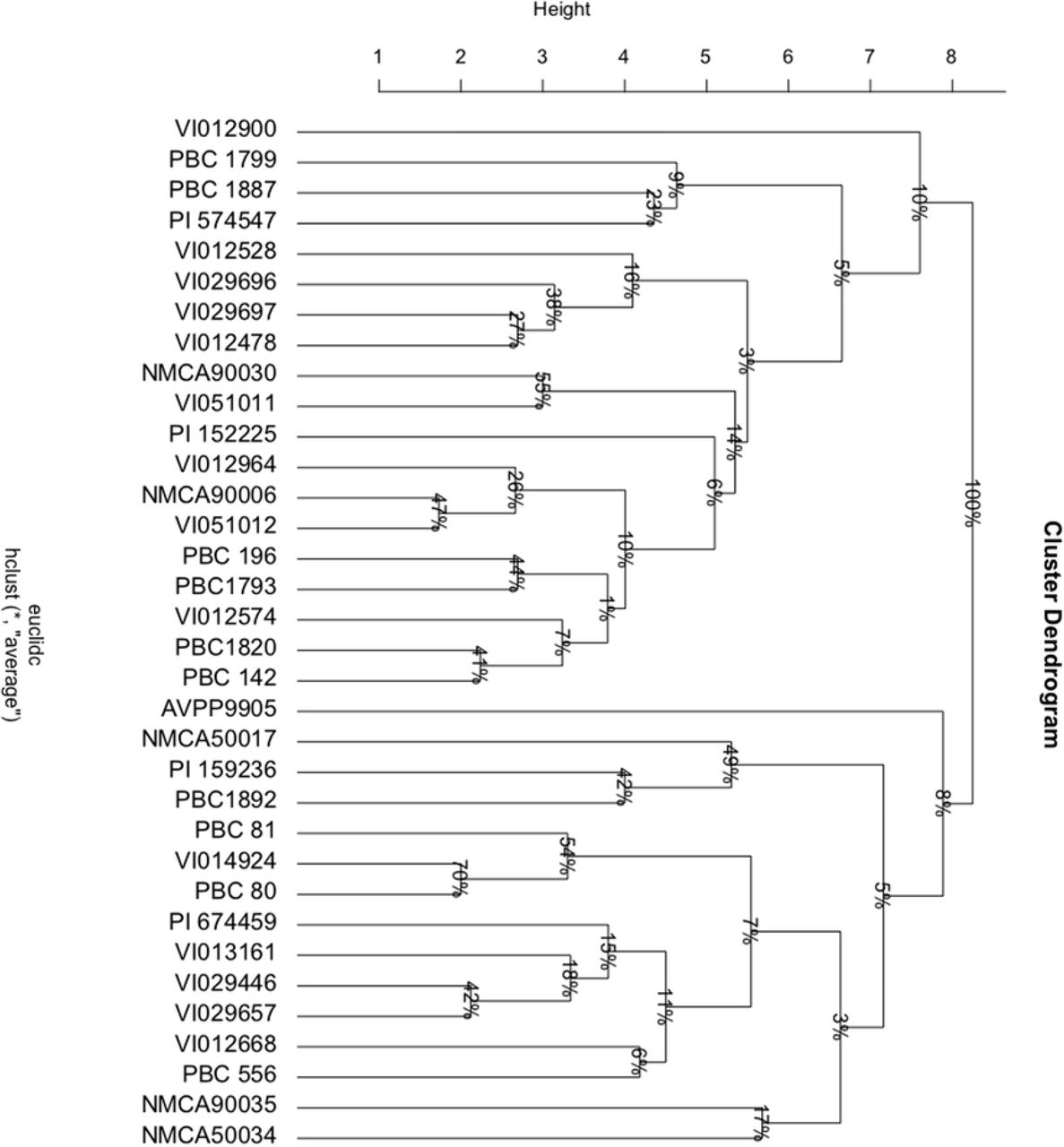
Gels were scored for each primer pair using a binary method: each accession was scored for presence (1) or absence (0) of amplicons of each size. The data were processed in R using the packages, ‘proxy’ (Shipunov et al., 2020) and ‘shipunov’ (Altschul et al., 1990) to produce a dendrogram with bootstrapping for the assessment of the relatedness between the individual accessions. A distance matrix was produced using the Dice index, and an unweighted pair group method with arithmetic mean (UPGMA) hierarchical cluster analysis was carried out. Bootstrap resampling was applied to clustering with 1,000 iterations.
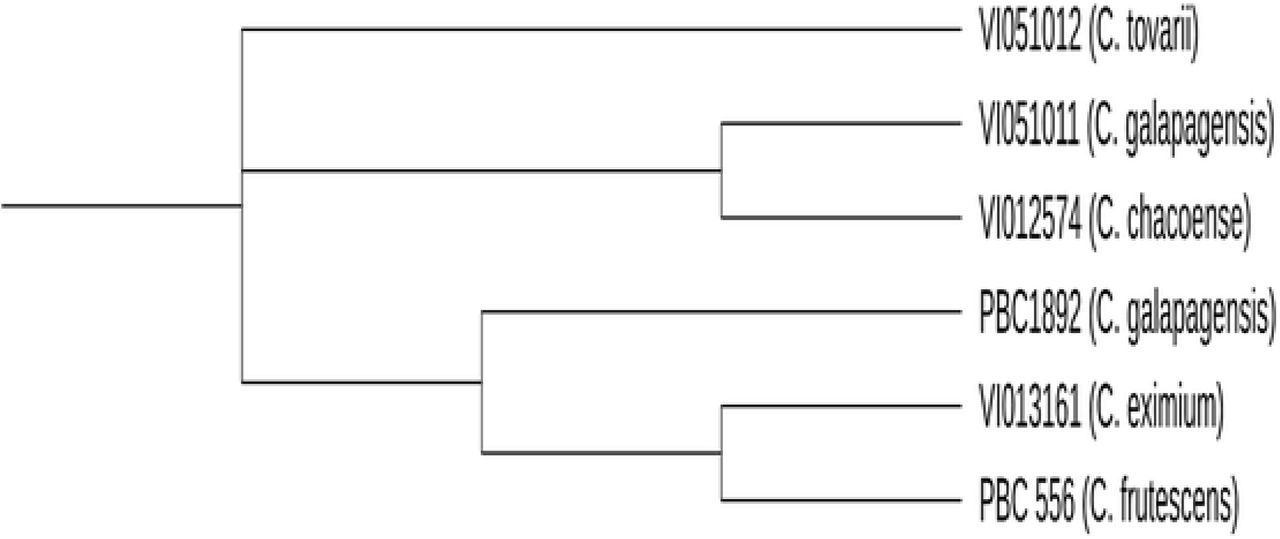
Further molecular analysis to clarify the identification of some accessions included the study of the waxy gene region of six accessions, VI051012 (C. tovarii); VI051011 (C. galapagensis, potentially C. annuum); VI012574 (C. chacoense, potentially C. annuum); PBC1892 (C. galapagensis); VI013161 (C. eximium); and PBC 556 (C. frutescens), using the primer pair, 860F and 2R (Carrizo García et al., 2016). The chosen accessions were those expected to need clarification due to possible misidentification, based on molecular and morphological data. The waxy region was amplified by PCR as before, with an annealing temperature of 60 °C. The quality of the products were evaluated by running on a 2% agarose gel with EtB‘out’ (Yeastern Biotech Co. Ltd., Taipei, Taiwan) at 100 Volts for 50 minutes, then visualized using a Microtek Bio- 1000F gel imager (Microtek International Inc., Hsinchu, Taiwan). The PCR products were sequenced by Genomics Biotechnology Co., Ltd. (New Taipei City, Taiwan) by the Sanger sequencing method. Low-quality nucleotides were manually removed throughout the resulting sequence, including approximately the first and last 60 nucleotides. The sequences were aligned using NCBI nucleotide BLAST (Hall, 1999) and a consensus sequence constructed using the CAP contig assembly program from BioEdit (Katoh and Standley, 2013). The sequences were aligned using multiple sequence alignment tool, Clustal MAFFT (Katoh and Standley, 2013), to produce a dendrogram.
Results
To clarify the phylogeny of the wild and domesticated Capsicum species in the sample, UPGMA clustering was carried out using the SSR molecular markers. The C. baccatum and C. chinense group accessions were distinct from the C. annuum group, with 76% bootstrap support (Fig. 1). The C. baccatum accessions made up a significant group, being closely clustered with the C. praetermissium accessions, as expected. This grouping was adjacent to a large group comprised of closely clustered C. chinense accessions with the C. galapagensis accession PBC1892, as well as C. eshbaughii, C. eximium and C. frutescens, separated from the C. baccatum group with a relatively low confidence interval. Within this grouping, C. chinense accession, PBC1820, was distinct from its counterparts, supported by 92% bootstrap support. Furthermore, the C. chinense species accessions were relatively separate from the accessions at the periphery of this grouping, the C. galapagensis accession PCB1892, the C. frutescens accession PBC556, and the C. eximium accession VI013161. The grouping of the C. eximium accession VI013161 with C. frutescens was similar in clustering from the waxy gene sequence (Fig. 2). These accessions were thus more similar to each other than they were similar to the C. galapagensis accession PBC1892, and this was a distinct grouping from other sequenced accessions.

Unweighted pair group method (UPGMA) clustering of Capsicum species according to single sequence repeat (SSR) markers. ‘Height’ represents dissimilarity, derived from ‘dice method’. Bootstrap resampling applied to clusters, represented as percent confidence interval. Numbers following the hyphen indicate replicates.

Clustering of selected Capsicum species according to their waxy gene sequences.
To better understand the crossing relationship between species, reciprocal hybridizations were performed between each combination of accessions (Fig. 3). Members of C. baccatum and C. praetermissium hybridized as either the female or male parent with at least one accession of each other species with the exception of C. rhomboideum (Fig. 3). However, of the sample of seeds selected for sowing, only the cross between VI014924 and PBC1969 germinated (Fig. 4). Hybridizations were not achieved between C. galapagoensis as either parent with accessions of C. tovarii, C. flexuosum, C. minutifolium, C. cardenasii, C. eshbaughii, and C. rhomboideum species. Capsicum eshbaughii hybridized more readily as the female parent, but failed to hybridize in either direction with accessions of C. eximium, C. frutescens, C. galapagoensis, C. tovarii, C. flexuosum, and C. rhomboideum. The majority of C. frutescens hybrids were achieved with C. annuum accessions, but successful hybridizations were found across a broad species range. Of the sample of seeds sown, 80% of the PBC556 × PBC1970 cross seeds germinated (Fig. 4).

Reciprocal hybridizations achieved between accessions of Capsicum species. Direction of arrow represents successful hybridizations in the male-female direction from which fruit was harvested.

Percent germination of selected hybrid seeds 12 weeks after sowing. Grey indicates unviable seeds.
The C. annuum group, which was adjacent to C. baccatum, consisted of the closely clustered C. annuum species accessions: PBC1799, AVPP9905, PBC1899, PBC1867, and PBC196, along with C. annuum var. glabriusculum PI574547, and C. chacoense VI012574 (Fig. 1). Neighboring this group was a cluster comprised of the C. eximium accession VI012964, the C. eshbaughii accession NMCA90006, the C. annuum accessions PBC142 and VI029657, the C. annuum var. glabriusculum accession PI674459, the C. tovarii accession VI051012, and the C. galapagoensis accession VI051011. Based on clustering of the waxy gene sequence, we found C. tovarii to be less similar to C. galapagoensis and C. chacoense than they were to one another (Fig. 2). Furthermore, the C. galapagoensis accession VI051011 and the C. chacoense accession VI02574 were similar in the waxy gene region, supporting their grouping together adjacent to the C. annuum group.
Accessions of C. annuum hybridized in both directions with one or more accessions of all species except C. rhomboideum and C. minutifolium (Fig. 3). Thirteen of the hybrids sown germinated well (Fig. 4). Capsicum annuum var. glabriusculum hybridized in either direction with at least one accession of every species except C. rhomboideum, and 7 out of 10 of those sown germinated (Fig. 4). Accessions of C. chacoense also hybridized broadly, but not with C. frutescens × chinense, C. minutifolium or C. rhomboideum, and seven of the 25 hybrids sown germinated. More than one cross was achieved between C. tovarii and an accession of every species except C. eshbaughii, C. eximium, C. galapagoensis, C. minutifolium and C. rhomboideum. Of these crosses, VI012574 × VI051012 and NMCA90030 × VI051012 germinated with 40% and 100% efficiency, respectively.
With 80% confidence interval, C. chacoense accession VI012900 and C. cardenasii accession NMCA90030 were clustered separately from the C. baccatum and C. annuum groups (Fig. 1). NMCA90035 clustered distinctly from its C. cardenasii counterpart, with bootstrap support of 57%. The C. flexuosum accessions NMCA50034 and NMCA50030 clustered closely together, with high bootstrap support (99%); adjacent was the C. minutifolium accession NMCA50053, and in a separate cluster, the C. rhomboideium accession NMCA50064, which was the most distinct grouping, separated from its neighbors with 100% confidence.
Capsicum cardenasii species hybridized with every species except C. frutescens, C. flexuosum, C. galapagensis and C. rhomboideum (Fig. 3), and the five of the nine hybrids sown germinated well (Fig. 4). No hybrids were achieved with C. flexuosum or C. minutifolium as female parents, however C. flexuosum hybridized as the male parent with C. chacoense, C. annuum var. glabriusculum, C. baccatum, C. tovarii, C. annuum and C. frutescens, while C. minutifolium hybridized with C. cardenasii, C. eshbaughii, and C. annuum var. glabriusculum, and of the crosses sown, only VI012574 × PBC124 germinated (Fig. 4). No successful hybrids were achieved with C. rhomboideum in either direction (Fig. 3).
We applied principal component analysis to the quantitative phenotypic data collected to understand phenotype across Capsicum species (Fig. 5). The first two components account for 59.9% of the total variation. The C. baccatum accessions made up a group along with PBC196 and VI01668, due to their correlated fruit and flower characteristics (pedicel length, fruit width, fruit length, anther length, filament length, corolla length) (Fig. 5). Capsicum annuum accessions made up a less distinct group, along with the wild progenitor C. annuum glabriusculum, and the other domesticated species C. chinense, C. frutescens, C. frutescens × chinense along with C. eximium (Fig. 5).

First two principal components of accessions in the wild and domesticated Capsicum species based on the quantitative phenotypic data.
The UGMA clustering of the accessions’ qualitative phenotypic data more closely mirrored the genetic relatedness based on SSR molecular markers, especially for the C. baccatum and C. praetermissum accessions (Fig. 6), which formed two groupings (VI012528, VI029696, VI029697; and PBC81, VI014924, PBC81). However, we found C. annuum did not form a unique clade, highlighting the phenotypic diversity of this domesticated species (Fig. 6).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Unweighted pair group method (UPGMA) clustering of wild and domesticated Capsicum species based on the qualitative phenotypic data, scored according to IPGRI Descriptors of Capsicum scoring method. Bootstrap resampling applied to clusters, represented as percent confidence interval.
Discussion
Understanding the relatedness between accessions of Capsicum species, and the extent to which they hybridize is key in identifying candidates for the introgression of traits of interest into commercial varieties. Our results mirror the widely accepted species phylogeny, with clustering based on SSR markers centered around C. annuum, C. baccatum and C. chinense complexes (Bosland and Votava, 2012; Pickersgill, 1971; Emboden Jr., 1962). Similarly, our results of phenotyping the accessions also support previously described genetic complexes (Carrizo García et al., 2016). Generally, accessions from the domesticated species (C. annuum, C. chinense, and C. frutescens) were nearer the origin of the score plot, with the exception of members of the domesticated C. baccatum, which clustered further away from the other domesticated species (Fig. 5). Conversely, members of the wild species were further away, indicating greater diversity (Fig. 5). Although we measured different phenotypic traits, our findings contradict those of Luna-Ruiz et al. (2018), who found greater levels of diversity among domesticated species for capsaicinoids. Interestingly, based on hybridization success rates, there was a weak relationship between relatedness and crossibility, which is in contrast with previous understanding that compatibility between complexes is low (van Zonneveld et al., 2015). This suggests potential for crop improvement with wild relatives of domesticated species with genetic bridge strategies.
When phylogeny, interspecific compatibility and phenotype are considered in concert, the identity of a number of accessions included in this study may be questioned. The issue of misidentification of Capsicum species has been raised previously, with several genebanks incorrectly reporting accessions C. frutescens as C. galapagoense (P.W. Bosland, pers. comm.). Thorough characterization is important in supporting conservation of genetic material and identifying gaps in genebank collections (FAO, 2010). Only 12% of national vegetable germplasm collections have been characterized biochemically, while 65% have been characterized morphologically (FAO, 2010). Thorough characterization is therefore key in understanding the reproductive relationships between Capsicum species.
Clustering based on SSR genotyping revealed a close relationship between C. baccatum accessions (Fig. 1), as expected for this well-accepted domesticated species. Capsicum praetermissium accessions are also grouped within this complex, which supports previous findings (Albrecht et al., 2012) and suggestions that C. praetermissium in fact comprises a subgroup of C. baccatum (Albrecht et al., 2012). Both PCA and UPGMA analysis of C. baccatum and C. praetermissium morphologies also show their close clustering, further demonstrating their relationship. Capsicum praetermissium is thought to have diverged prior to domestication of C. baccatum, and has not been utilized in breeding domestic C. baccatum accessions (Albrecht et al., 2011). We found C. praetermissium readily hybridized with C. baccatum (Fig. 3) in line with the findings of Emboden Jr. (1961), and thus offers potential as a genetic resource.
Capsicum chinense species accessions comprised a significant cluster, which included C. chinense, C. frutescens, C. eshbaughii, and C. galapagoensis (Fig. 1). The grouping of C. chinense adjacent to C. baccatum was in line with a recent study that also used SSR molecular markers to characterize Capsicum species (Guzmán et al., 2020). However, this is contrary to findings of Pickersgill et al. (1991) and Ince et al. (2010), who grouped C. chinense within the C. annuum complex. Furthermore, in this study, a total of 20 crosses were achieved between C. annuum (including the wild progenitor C. annuum var. glabriusculum), and C. chinense, 13 of which had a C. chinense female parent (Fig. 3). Seeds from two of these crosses were sown (VI029446 × PBC1969 and PI152225 × NMCA90030) and germinated well (Fig. 4). This contrasts to previous work that reports a barrier to reproduction between C. annuum and C. chinense (Campos et al., 2004). However, Costa et al., (2009) found that crosses between C. chinense and C. annuum accession were possible. This highlights the variability in compatibility, and its dependence on accession selection.
The grouping of C. frutescens in the C. chinense complex was in line with previous findings of the close relationship of these species (Fig. 1) (Shiragaki et al., 2020). A number of researchers argued their identities as sister species within the annuum clade (Albrecht et al., 2012; Tam et al., 2009; Walsh and Hoot, 2001). Furthermore, we found C. frutescens hybridized readily with both members of the C. baccatum and C. annuum clades, as well as with C. chinense (Fig. 3). Of the three C. frutescens hybrids selected for sowing, 80% of the PBC556 × PBC1970 hybrid seeds germinated (Fig. 4). The relationship of C. eshbaughii to this clade, and its pairing with C. eximium was consistent with its previous placement in the ‘Purple Corolla clade’ (Carrizo García et al., 2016). Capsicum chinense formed hybrids with other members of this grouping (Fig. 3), and 100% of seeds from the C. chinense and C. eximium cross germinated (Fig. 4).
Interestingly, C. eximium and C. cardenasii appeared distantly related in our analysis (Fig. 1), which contradicts previous reports of these species being within the C. pubescens complex (Ibiza et al., 2012; McLeod et al., 1983). Furthermore, their phenotypes correlated closely with C. annuum accessions (Fig. 5). This raises the question of the validity of the identification of accessions VI013161 and VI012964 as C. eximium.
The C. annuum accessions comprise a major grouping adjacent to the C. baccatum group (Fig. 1). A sample of C. annuum accessions (PBC1799, PBC196, PBC1867 and AVPP9905) formed a tightly clustered group, indicating genetic similarity. They also display highly correlated phenotypes, forming a cluster along with accessions from other domesticated species (Fig. 5). The C. annuum accessions PBC142 and VI029657 were in an adjacent group (Fig. 1), therefore may be considered part of the wider C. annuum complex, along with C. chacoense accession VI012574, C. galapagensis, VI051011, and C. tovarii accession VI051012. The presence of C. chacoense (VI012574) in this group, distant from the second C. chacoense accession (VI012900) included in this study, highlights its possible misidentification. Sequencing clustered VI012574 with C. galapagensis accession VI051011, which may be considered a member of the C. annuum complex (Fig. 2). Principal component analysis (Fig. 5) revealed VI012574 was grouped with C. annuum accessions, away from its counterpart, while UPGMA analysis further highlights this disparity. Direct observation of the phenotypes emphasises the similarity between the morphology of VI012574 and typical C. annuum features. This includes upright growth, elongated fruits, and relatively large flowers with blue anthers.
The C. galapagoensis accession, PBC1892 was grouped with the wider C. baccatum cluster (Fig. 1), conflicting previous findings that C. galapagoensis is derived from a C. annuum progenitor population (Choong, 1998). No successful hybridizations were achieved between PBC1892 and PBC556 (C. frutescens), which clustering suggested were closely related. The second C. galapagoensis accession included in the study, VI051011, was grouped within the C. annuum complex, and displayed a distinctly different phenotype to that of PBC1892. PBC1892 had a compact growth habit, very small fruits, flowers and leaves, and densely pubescent stems and leaves, typical of C. galapagoensis descriptions. Conversely, VI051011had a morphology similar to that of C. annuum, reflected in its close proximity to the PCA origin, along with C. annuum accessions. Eight hybridizations were achieved between VI051011 and C. annuum accessions, and of the selected hybrid seeds sown, 20% germinated. The close clustering of VI051011 with the C. annuum complex, their similar morphology and their ability to hybridize suggests likely misidentification of this accession.
The grouping of VI051011 (C. galapagoensis) in the C. annuum complex (Fig. 1) supports previous findings that C. galapagoensis is derived from a C. annuum progenitor population (Choong, 1998). Eight hybridizations were achieved between VI051011 and C. annuum accessions, and of the selected hybrid seeds sown, 20% germinated. The second C. galapagoensis accession in this study, PBC1892, was grouped with the wider C. baccatum cluster (Fig. 1). No successful hybridizations were achieved between PBC1892 and PBC556 (C. frutescens), which clustering suggested were closely related. The phenotypes of these accessions were distinctly different; PBC1892 having a compact growth habit, very small fruits, flowers and leaves, and densely pubescent stems and leaves, which is typical of C. galapagoense descriptions. VI051011 had morphology similar to that of C. annuum (Fig. 5) and its grouping within the C. annuum complex along with its ability to hybridize with C. annuum suggested likely misidentification.
There were five further clusters consisting of C. chacoense, C. cardenasii, C. flexuosum, C. minutifolium, and C. rhomboideum respectively, which had increasingly distant relation to the three major species complexes (Fig. 1). Although C. chacoense has been previously grouped within the C. baccatum clade (Carrizo García et al., 2016; McLeod et al., 1983), this wild species has an apparently distant relationship with C. baccatum. Capsicum cardenasii was similarly distantly related to other clades. Other studies (Ibiza et al., 2012; Ince et al., 2010; McLeod et al., 1983) also found C. chacoense and C. cardenasii not to be closely related to any major clade. Furthermore, VI012900 hybridized readily with members of both C. annuum and C. baccatum clades (Fig. 3). Furthermore, both C. chacoense and C. cardenasii accessions (with the exception of PI159236 and PI15225) lay on the periphery of the PCA plot, clustering with neither C. baccatum or C. annum groups. . This suggests C. cardenasii and C. chacoense accessions are not members of either C. baccatum or C. annuum clades.
Capsicum flexuosum, C. minutifolium and C. rhomboideum were distantly related to the major clades, consistent with the body of literature (Carrizo García et al., 2016; Guzmán et al., 2020; Choong, 1998). The C. flexuosum accession (NMCA50034) also had a distant relationship with more domesticated species (Fig. 5). A small number of hybridizations were achieved between C. flexuosum and C. minutifolium with members of both C. annuum and C. baccatum clades. However, no hybridizations were achieved between C. rhomboideum and any other accession.
The results reported here highlight the extent of phenotypic diversity in Capsicum species, the complexity of Capsicum phylogeny, and the similarly complex reproductive relationships between Capsicum species. The evidence suggesting the incorrect identification of VI013161, VI012964, VI012574, and VI051011 may highlight a broader issue of misidentification of Capsicum in genebanks. Thorough characterization of Capsicum genetic material taking a multifaceted approach is therefore important for the development of future breeding programs. Furthermore, the generation of diverse hybrids among accessions of all species included in this study (with the exception of C. rhomboideum) demonstrates the possibility for introgression of a diverse range of traits of interest directly or through the design of bridge crossing strategies. Wild relatives of domesticated Capsicum species therefore represent significant potential for future breeding programs, and should not be discounted on the basis of their assumed relatedness to domesticated species.
Copyright
© 2021 Parry et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Data Availability
The data used in this study are available in the World Vegetable Center repository, HARVEST, doi:10.22001/wvc.73914, https://worldveg.tind.io/record/73914
Funding
Funding for this research was provided by the Ministry of Science and Technology (MOST) of Taiwan (Project ID:107-2311-B-125 -001 -MY3) as well as long-term strategic donors to the World Vegetable Center, Taiwan; UK aid from the UK government; U.S. Agency for International Development (USAID); Australian Centre for International Agricultural Research (ACIAR), Germany, Thailand, Philippines, Korea, and Japan.
Competing interests
The authors have declared that no competing interests exist.
Acknowledgments
We thank Dr. Paul Bosland of Chile Pepper Institute, New Mexico State University, USA for providing Capsicum accessions.
Literature Cited




