

ABSTRACT
Background To address the dire need for a safe and effective vaccine to protect individuals from and reduce transmission of SARS-CoV-2, we developed a COVID-19 vaccine that elicits not only robust humoral responses but also activates T cells. Our bivalent vaccine expresses both an optimized viral spike (S) protein (S-Fusion) and the viral nucleocapsid (N) protein with an Enhanced T-cell Stimulation Domain (N-ETSD) that directs N to the endo/lysosomal subcellular compartment to enhance MHC class II responses. The vaccine antigens are delivered by the second-generation adenovirus serotype 5 [E1-, E2b-, E3-] platform (hAd5) that has been safely administered and found to be effective in generating tumor-specific T cells even in the presence of pre-existing adenovirus immunity. Here, we report our findings on the safety and efficacy of our hAd5 S-Fusion + N-ETSD subcutaneous (SC) prime and thermally-stable oral boost vaccine in generating SARS-CoV-2-neutralizing antibodies, eliciting N- and S-specific T-cell responses, and providing complete protection with the clearing of virus after challenge in Non-Human Primates (NHP). A key objective of the study was to explore the efficacy of a novel thermally-stable oral hAd5 S-Fusion + N-ETSD to serve as a booster dose following an SC prime.
Methods Group 1 NHP received the hAd5 S-Fusion + N-ETSD vaccine on Days 0 and 14 by SC injection (1011 VP), and on Day 28 by a single oral boost (1010 VP); Group 2 received vaccination on the same schedule, but with an SC prime and two oral boosts. Group 3 placebo NHP were dosed with vehicle-only SC-oral-oral. Blood for the isolation of sera and PBMCs was collected throughout the study. ELISA was used for determination of anti-S IgG levels, cPass™ for presence of neutralizing antibodies, and ELISpot for interferon-γ(IFN-γ) and interleukin-4 (IL-4) secretion by T cells.
On Study Day 56, all NHP were challenged with intratracheal/intranasal 1 × 106 TCID50/mL SARS-CoV-2. Bronchoalveolar lavage (BAL) samples were collected on Day 42 pre-challenge and at several time points post-challenge; nasal swabs were collected daily post-challenge. NHP were euthanized on Study Day 70 and tissue collected for histopathological analyses. Viral load and active viral replication were determined in BAL and nasal swab specimens by RT qPCR of genomic and subgenomic RNA, respectively. Safety was determined by cage-side observations such as weight as well as hematology and clinical chemistry analyses of blood.
Results The hAd5 S-Fusion + N-ETSD vaccine, both SC and oral, elicited no apparent toxicity seen in clinical chemistry, hematology, or cage-side observations. Neutralizing antibodies were induced in 9 of 10 vaccinated NHP and anti-S IgG positive titers in 10 out of 10. Th1 dominant T-cell responses were elicited by both S and N antigens, with responses being greater for N. Viral replication was inhibited from Day 1 post-SARS-CoV-2 challenge with complete protection in all (10/10) primates within 7 days of challenge from both nasal passages and lung. Replicating SARS-CoV-2 dropped immediately and was undetectable as soon 3 days post-challenge. There was a rapid decline in lymphocytes from the periphery on Day 1 post-challenge and a rebound within 3 days following challenge that was significantly higher by Day 14 post-challenge in Group 1 as compared to Group 3 placebo NHP.
Conclusions In the rhesus macaque NHP model, the bivalent hAd5 S-Fusion + N-ETSD subcutaneous and oral vaccine provided complete protection of nasal passages and lung against SARS-CoV-2 challenge by eliciting neutralizing antibodies plus Th1 dominant N- and S-specific T-cell responses. Inhibition of viral replication within the first 24 hours post-challenge, in vaccinated NHP compared to placebo NHP, suggests the presence of SARS-CoV-2-specific cytotoxic T cells that rapidly cleared infected cells. The rapidity of clearance implies that shedding of live viruses may be attenuated as a result of vaccination and thus the vaccine has the potential to prevent transmission of virus by infected individuals. Clinical trials of hAd5 S-Fusion + N-ETSD are ongoing. The hAd5 S-Fusion + N-ETSD subcutaneous prime/boost vaccine has completed Phase 1 clinical trials and Phase 2/3 trials are actively recruiting. The thermally-stable oral vaccine will enter Phase 1 trials as a prime and boost, as well as explored to provide a boost to subcutaneous vaccination.
INTRODUCTION
To address the ongoing COVID-19 pandemic, particularly in the face of the high efficiency of SARS-CoV-2 transmission that puts vulnerable persons and front-line workers at risk (1–4), we have developed a vaccine to protect individuals from and prevent transmission of SARS-CoV-2 that elicits not only robust humoral responses but also activates T cells. This bivalent hAd5 S-Fusion + N-ETSD vaccine (5) expresses both an optimized viral spike (S) protein (S-Fusion) and a nucleocapsid protein with an Enhanced T-cell Stimulation Domain (N-ETSD) that directs N to the endo/lysosomal subcellular compartment (6) to enhance MHC class II responses. The vaccine antigens are delivered by the second-generation adenovirus serotype 5 [E1-, E2b-, E3-] platform (Fig. 1A) that is safe and effective even in the presence of pre-existing adenovirus immunity (7, 8).

(A) The human adenovirus serotype 5 vaccine platform with E1, E2b, and E3 regions deleted (hAd5 [E1-, E2b-, E3-]) is shown. (B) The SARS-CoV-2 virus displays spike (S) protein as a trimer on the viral surface. S protein comprises the N-terminal (NT), the S1 region including the Receptor Binding Domain (RBD), the S2 and transmembrane (TM) regions, and the C-terminal (CT); other function regions not labeled. (C) The bivalent vaccine comprises both S-Fusion and N-ETSD under control of cytomegalovirus (CMV) promoters delivered by the hAd5 [E1-, E2b-, E3-] platform.
We previously developed this attenuated hAd5 viral vector platform that can be used to rapidly generate vaccines against multiple agents, allowing production of high numbers of doses in a minimal time frame. The hAd5 platform has unique deletions in the early 1 (E1), early 2 (E2b) and early 3 (E3) regions (hAd5 [E1-, E2b-, E3-]), which distinguishes it from other adenoviral vaccine platform technologies under development (9, 10), and not only allows it to be effective in the presence of pre-existing adenovirus immunity but has a very low risk of generating de novo vector-directed immunity. Genes encoding target antigens are cloned into the viral genome, which once administered in vivo infect antigen presenting cells that express the inserted antigen gene and induce immune responses to the pathogenic target. The platform induces both antibodies and cell-mediated immunity (CMI). We have utilized this platform to produce vaccines against viral antigens such as Influenza, HIV-1 and Lassa fever and COVID-19 (5, 11–15). In 2009, we employed the hAd5 [E1-, E2b-, E3-] vector platform to express hemagglutinin (HA) and neuraminidase (NA) genes from the H1N1 pandemic viruses (11). Inserts were consensus sequences designed from viral isolate sequences and the vaccine was rapidly constructed and produced. Vaccination induced H1N1 immune responses in mice, which afforded protection from lethal virus challenge. In ferrets, vaccination protected from disease development and significantly reduced viral titers in nasal washes. H1N1 CMI as well as antibody induction correlated with the prevention of disease symptoms and reduction of viral replication (11).
The overwhelming majority of other SARS-CoV-2 vaccines in development target only the S antigen (Fig. 1B) and are expected to elicit SARS-CoV-2 neutralizing antibody responses. In the development of our vaccine, we have paid specific attention to the generation of T cells which is predicted to enhance the breath and duration of the protective immune response against the two antigens; the addition of N in particular affords a greater opportunity for T cell responses (16–18). Importantly, we have previously shown that the hAd5 S-Fusion + N-ETSD vaccine elicits T helper cell 1 (Th1) dominant antibody responses to both S and N as well as T-cell activation after vaccination of a murine (CD1) pre-clinical animal model (5). We have also shown that the SARS-CoV-2 antigens expressed by the hAd5 S-Fusion + N-ETSD construct are recognized by T cells from previously SARS-CoV-2 infected individuals when expressed by autologous monocyte-derived dendritic cells (6). These studies provide evidence that vaccination with the hAd5 S-Fusion + N-ETSD vaccine (Fig. 1C) will re-capitulate natural infection that will then generate protective antibodies and memory T cells. As described in Canete and Venuesa’s “COVID-19 makes B cells forget, but T cells remember” (19), T cells provide protection even in the absence of antibody responses. This is supported by Sekine et al. (18), who characterized T cell immunity in COVID-19 convalescent patients, finding SARS-CoV-2-specific T cells in most convalescent individuals (including asymptomatic cases) with undetectable antibody responses (18).
A key objective of the NHP study design, described below, was to assess the efficacy of a subcutaneous (SC) prime followed by a thermally-stable oral boost. An oral boost provides several advantages in SARS-CoV-2 vaccination, including a greater potential for generating mucosal immunity. SARS-CoV-2 is a mucosal virus and is only rarely detected in blood (20, 21), therefore vaccines that specifically target mucosal immunity are of interest (22).
Compelling additional advantages of a thermally-stable oral boost are that it would likely transform the global distribution of vaccines, especially in developing nations; potentially enable patients to self-administer the boost(s) at home; and - because hAd5 S-Fusion + N-ETSD induces both humoral and CMI responses to both antigens - has the possibility of serving as a ‘universal’ heterologous booster vaccine to the multitude of SARS-CoV-2 vaccines under development.
Here, we report our findings from our NHP study to evaluate the safety and efficacy of the novel second-generation hAd5 S-Fusion + N-ETSD vaccine in rhesus macaques as well as the efficacy of a thermal-stable oral boost.
METHODS
The study, formally titled “Evaluating the Efficacy of Novel SARS-CoV-2 Vaccines in the NHP Model” (Study Number B05856) was performed at Battelle Biomedical Research Center (West Jefferson, Ohio) and was sponsored by the Biomedical Advanced Research & Development Authority (BARDA), Office of the Assistant Secretary for Preparedness and Response (ASPR), Department of Health and Human Services (DHHS) and the National Institutes of Health/National Institute for Allergy and Infectious Diseases (NIH/NIAID) (Washington, DC). This was a non-GLP study.
Dosing and sample collection
In our study design, we compared two SC injections administered in the center of the back just caudal to the scapular region of 1 × 1011 viral particles (VP) on Days 0 and 14 followed by an oral capsule delivered via a feeding tube after a minimum of 4 hours of fasting of 1 × 1010 infectious units (IU) on Day 28 (Group 1) to one prime SC injection and two oral boost doses with the same dosages and timing (Group 2), as shown in Figure 2.

Group 1 NHP (n = 5) were vaccinated by subcutaneous (SC) injection of 1 × 1011 VP on Days 0 and 14, and received an oral boost of 1 × 1010 IU on Day 28. Group 2 NHP (n = 5) were vaccinated similarly, but both boosts were oral. Sera were collected as indicated (blue arrows). On Day 42, NHP were transferred to a BSL-3 facility for viral challenge with 1×106 TCID50 VP of SARS-CoV-2 intranasally/intratracheally. Nasal swab (yellow arrows) and bronchoalveolar lavage (BAL) samples (red arrows) were collected as indicated. Animals were euthanized on Day 70 and tissues collected for pathological analyses.
Active vaccination groups comprised 5 male (3) and female (2) randomized NHP, and the placebo Group 3 NHP, one male and one female.
On Day 42, NHP were transferred to a BSL-3 facility and on Day 56 they were challenged with SARS-CoV-2 1 × 106 TCID50/mL delivered to anesthetized animals both intratracheally and intranasally. Nasal swabs (buccal swabs, nasal washes, oropharyngeal swabs) were taken daily after challenge, and bronchoalveolar lavage (BAL) was performed every other day until the end of the study of Day 70.
Assessment of safety, determination of immune responses, and quantification of post-challenge viral load and replication
Methods for safety assessments, including cage-side observations, animal weights, hematology and clinical chemistry; and efficacy assessments including ELISA for anti-S IgG, viral neutralization in cPass™, T-cell responses by ELISpot during the pre-challenge, and RT qPCR of viral genomic and subgenomic RNA for determination of viral load and viral replication in the post-SARS-CoV-2 challenge period are detailed in Supplementary Methods.
RESULTS
Safety
The hAd5 S-Fusion + N-ETSD vaccine was found to be safe
NHP that received the hAd5 S-Fusion + N-ETSD in either Group 1 or 2 showed no signs of toxicity as assessed by cage-side observations and recorded weight (Supplementary Fig. S1). No serious adverse events or animal deaths occurred prior to the pre-determined necropsy date.
Hematological findings (Supplementary Table S1) and clinical chemistry analyses (Supplementary Table S2) are reported in the Supplementary Results.
Immunological Responses to Vaccination
The immunological responses including generation of anti-S IgG, SARS-CoV-2 neutralization, and T-cell activation for Groups 1 and 2 are presented below.
Group 1 NHP produced neutralizing, anti-spike antibodies
All Group 1 (SC, SC, Oral) S-Fusion + N-ETSD vaccinated NHP produced anti-S antibodies (Fig. 3A), with levels increasing from Day 0 to Day 42. Anti-S IgG (geometric mean, Fig. 3B) climbed steadily following prime-boost with a peak at Day 42. Sera from all five NHP in Group 1 demonstrated inhibition of S receptor binding domain (RBD) binding to angiotensin converting enzyme 2 (ACE2) in cPass™ (23), reflecting the ability to neutralize the SARS-CoV-2 virus (Fig. 3C). Anti-S IgG levels correlated with inhibition in cPass™ (R2= 0.5166) as shown in Figure 3D.
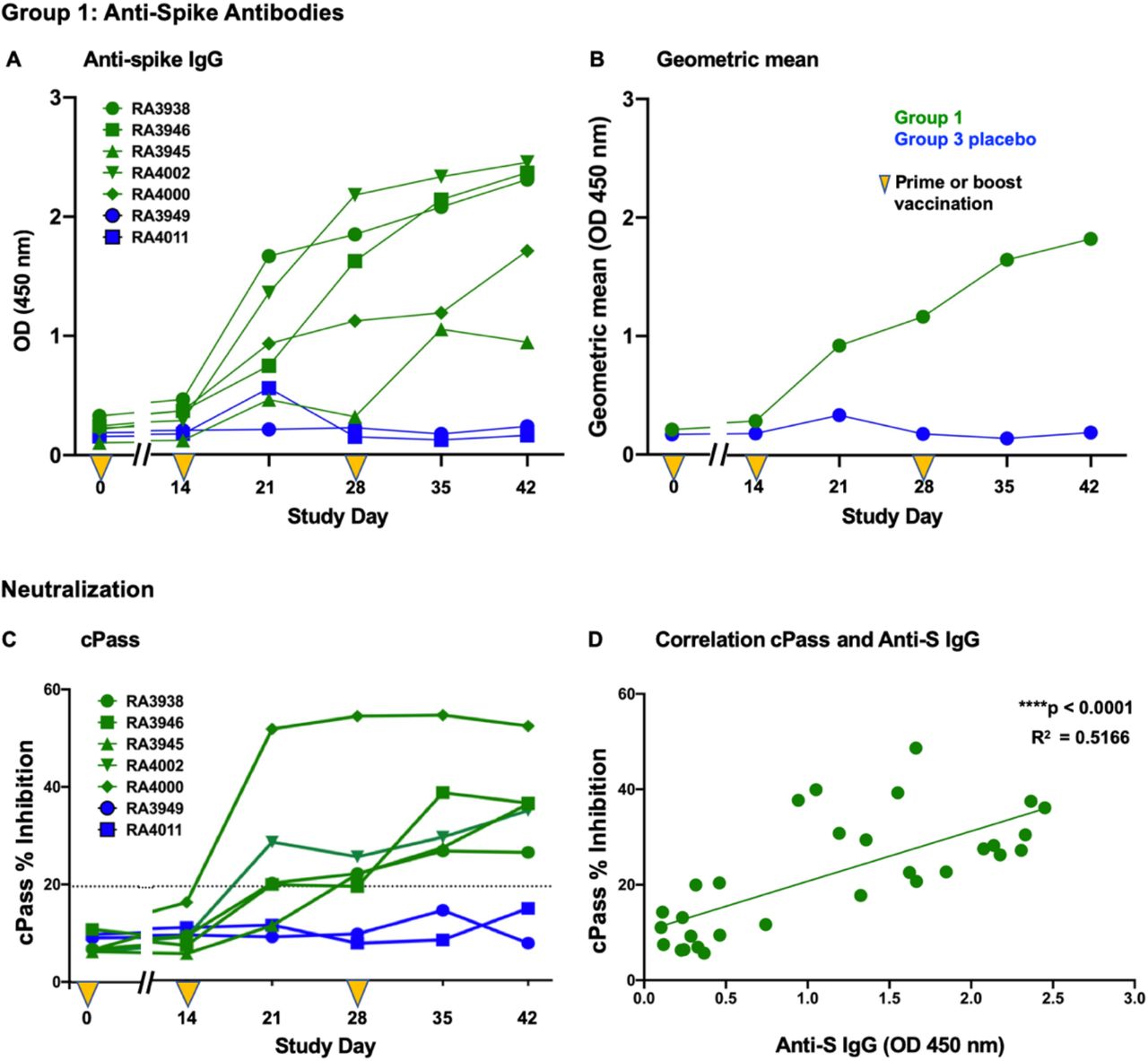
(A) Anti-spike IgG levels in individual NHP and the (B) geometric mean of Group 1 and Group 3 NHP are shown. The results from (C) cPass™ for individual NHP and (D) the correlation of those results to anti-spike IgG levels are shown; p < 0.0001 and R2= 0.5166.
Th1 dominant S- and N-specific T cells were activated following vaccination in Group 1 NHP
As shown in Figure 4A, PBMC-derived T cells secreted IFN-g in response to pulsing by either spike (S1 plus S2) and nucleocapsid (N) peptide pools, with responses being greater on Days 14 and 35 to the N antigen. Conversely, the number of IL-4-secreting T cells in response to either S or N peptide pools was very low (Fig. 4B). The IFN-g/Il-4 ratio demonstrates Th1 dominance, with some ratios being ‘infinity’ due to IL-4 being below the level of detection in some cases.

Secretion of (A) interferon-γ (IFN-γ) and (B) interleukin-4 (IL-4) as well as (C) the ratio of IFN-γ/IL-4 from PBMC-derived T cells from Group 1 and 3 NHP is shown. Ratios of ‘infinity’ due to undetectable IL-4 are represented at the top of the graph as open circles. Cells pulsed with PMA-ionomycin were used as positive controls as described in Methods(Supplementary Materials) but those data are not graphed here. Data graphed with mean and SEM.
Group 2 NHP (single SC prime with oral boost) produced neutralizing, anti-spike antibodies
All Group 2 (SC, Oral, Oral) S-Fusion + N-ETSD vaccinated NHP produced anti-S IgG that increased with time, as shown for individual NHP (Fig. 5A) and reflected by the geometric mean (Fig. 5B). Sera from 4 of 5 NHP in Group 2 demonstrated inhibition in cPass™ (Fig. 5C) and anti-S IgG levels correlated with inhibition (R2 = 0.4242) (Fig. 5D). As will be noted later in results of viral clearance, the NHP not showing inhibition in cPass™ nonetheless successfully cleared the virus from nasal and lung passages to undetectable levels, implying a role for T cells and/or the existence of non-neutralizing antibodies.

(A) Anti-spike IgG levels in individual NHP and the (B) geometric mean of Group 2 and Group 3 NHP are shown. The results from (C) cPass™ for individual NHP and (D) the correlation of those results to anti-spike IgG levels are shown; p < 0.0001 and R2= 0.4242.
Th1 dominant S- and N-specific T cells were activated following vaccination in Group 2 NHP
T-cell responses for Group 2 NHP were very similar to those for Group 1 NHP, increasing with time and being greater for N. Overall, Group 2 T-cell responses were higher than those in Group 1 (Fig. 6A) and peaked at Days 14 and 35. T-cell responses were also Th1 dominant, with IL-4 secretion being very low (Fig. 6B and C).

Secretion of (A) interferon-γ (IFN-γ) and (B) interleukin-4 (IL-4) as well as (C) the ratio of IFN-γ/IL-4 from PBMC-derived T cells from Group 2 and 3 NHP is shown. Ratios of ‘infinity’ due to undetectable IL-4 are represented at the top of the graph as open circles. Cells pulsed with PMA-ionomycin were used as positive controls as described in Methods (Supplementary Materials) but those data are not graphed here. Data graphed with mean and SEM.
Immunological Responses Post-Challenge
Lymphocytes decreased in the periphery immediately post-challenge, then rebounded in vaccinated NHP
On Day 56 of the study (or challenge Day 0), NHP were challenged with SARS-CoV-2 infection. Peripheral lymphocyte levels were determined in blood samples drawn just before challenge on challenge Day 0 and on Days 1, 3, 5, 7 and 14 post-challenge. On the Day of challenge, the lymphocyte level was similar amongst groups (Supplementary Fig. S2). To compare the changes in lymphocytes as a result of challenge, the difference between lymphocyte values at Day 0 and subsequent post-challenge days for each individual NHP was calculated, and the delta values for each group graphed for each day. As seen in Figure 7, on Day 1 (24 hours after initiation of infection) peripheral lymphocyte levels dropped dramatically in all groups, suggesting potential lymphocyte recruitment to the site of SARS-CoV-2 infection - the nose and lung (24, 25). By Day 3 post-challenge, lymphocytes in all groups had increased and continued to increase up to Day 14 for Group 1 (Fig. 7A) and to a plateau in Group 2 by Day 7 (Fig. 7B).

(A) Lymphocyte levels for Groups 1 and 3 post-challenge calculated as the difference from Day 0 for each NHP. On Day 14, the difference between Group 1 and Group 3 was significant (p= 0.0404). (B) Lymphocyte levels for Groups 2 and 3. On Days 3 and 5, differences between Group 2 and Group 3 placebo were significant, p= 0.0252 and p= 0.0141, respectively. The value of ‘0’ (no change) was used for each NHP on Day 0. Statistical analyses performed using a two-tailed, unpaired Student’s t-test.
On Day 14 post-challenge, lymphocyte levels were significantly higher (p= 0.0404) for hAd5 S-Fusion + N-ETSD vaccinated Group 1 NHP compared to Group 3 placebo control NHP (Fig. 7A) and were higher in Group 2 versus Group 3, but not significantly (Fig. 7B). The relatively greater rebound of lymphocytes in vaccinated NHP could be due to proliferation of a subset of T lymphocytes in response to SARS-CoV-2-specific infection (16, 26, 27), a hypothesis that awaits confirmation.
Protective Immunity Following Viral Challenge
Following SARS-CoV-2 challenge, viral replication was immediately inhibited from Day 1 post-challenge in both nasal passages and lung with no further growth and complete clearance within 7 days of challenge in all (10/10) animals. Replicating SARS-CoV-2 was undetectable as early as 3 days post-challenge in some NHP. This represents complete protection of nasal passages and lung against viral challenge afforded by antibody plus Th1 dominant N- and S-specific T-cell responses in both Group 1 and 2 primates receiving SC prime and thermally-stable oral boost vaccination.
Group 1 nasal passages: complete clearance of replicating, infectious SARS-CoV-2 virus in nasal passages following hAd5 S-Fusion + N-ETSD vaccination in Group 1
To determine the viral load, RT-qPCR analysis of genomic RNA was performed on nasal swab samples. Vaccination of Group 1 NHP reduced SARS-CoV-2 viral load in the nasal swab samples as compared to placebo control NHP from Day 1 (Fig. 8A and B). The viral load in Group 1 vaccinated NHP continued to diminish to very low or undetectable levels of SARS-CoV-2 in all vaccinated animals. Placebo controls had moderate to high levels (range 2E+09 - 8.4E+03 gene copies/mL) of SARS-CoV-2 present in nasal swab samples for the duration of the study.

(A) Individual viral load (RT qPCR) and the (B) geometric mean; (C) individual replicating virus (sgRNA) and the (D) geometric mean for nasal samples are shown. Values below the detection level are highlighted in yellow.
Replicating virus in nasal swab samples was determined by RT qPCR of sgRNA, as shown in Figure 8C and D. The dynamics of viral replication inhibition were rapid, with a 2 log reduction of viral replication by Day 2, with no further replication of challenge inoculum, and a reduction to below the level of detection in 4 of 5 NHP by Day 5 post-challenge. By Day 7, replicating virus in all primates was below the level of detection in nasal passages, demonstrating complete protection conferred by the vaccine.
Group 1 lung: complete clearance of replicating, infectious SARS-CoV-2 virus in lung following hAd5 S-Fusion + N-ETSD vaccination in Group 1
Viral load and replicating virus in BAL samples from Group 1 NHP were both ~2 logs lower than in placebo Group 3 NHP starting on Day 1 after challenge with SARS-CoV-2 (Fig. 9B and D), and levels of replicating virus were below the level of detection in in all Group 1 NHP by Day 7 post-challenge (Fig. 9C).
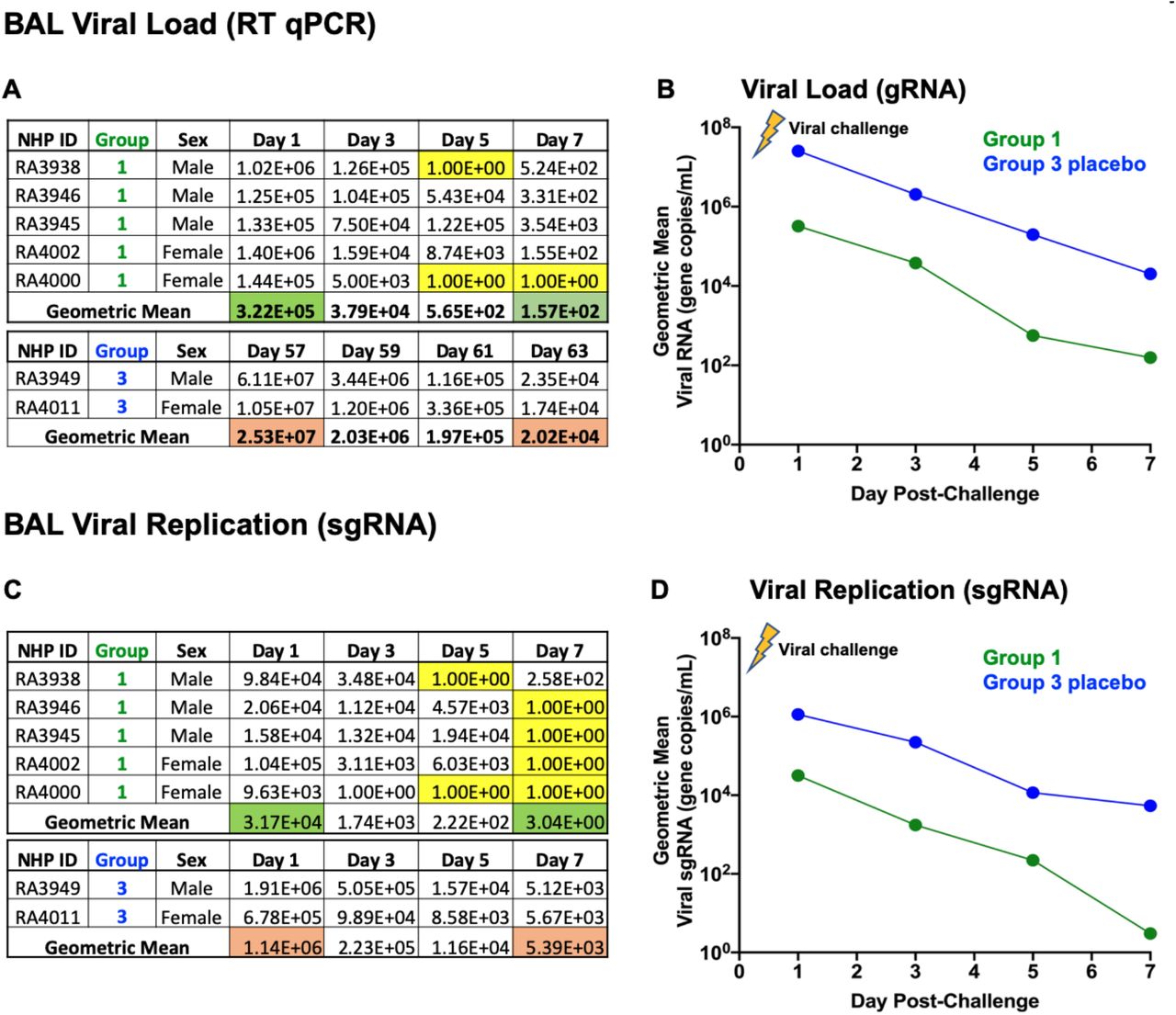
(A) Individual viral load (RT qPCR) and the (B) geometric mean; (C) individual replicating virus (sgRNA) and the (D) geometric mean for lung are shown. Values below the detection level are highlighted in yellow.
Group 2 nasal passages: complete clearance of replicating, infectious SARS-CoV-2 virus from nasal passages following hAd5 S-Fusion + N-ETSD vaccination in Group 2
Similar to Group 1 NHP that received two SC doses and one oral boost, Group 2 NHP receiving one SC and two oral boosts showed both a reduction in viral load and replicating virus in nasal passages (Fig. 10A-D). Strikingly, replicating virus levels in nasal passages of Group 2 NHP were all below the level of detection by Day 5 post-SARS-CoV-2 challenge. (Fig. 10C)

(A) Individual viral load (RT qPCR) and the (B) geometric mean; (C) individual replicating virus (sgRNA) and the (D) geometric mean for nasal samples are shown. Values below the detection level are highlighted in yellow.
Group 2 lung: complete clearance of replicating, infectious SARS-CoV-2 virus from lung following hAd5 S-Fusion + N-ETSD vaccination in Group 2
As with Group 1, Group 2 NHP had >1 log lower viral load (Fig, 11A and B) and replicating virus in lung (Fig, 11C and D) even on Day 1 post-challenge. By Day 7, replicating virus in all Group 2 NHP was below the level of detection (Fig. 11C).

(A) Individual viral load (RT qPCR) and the (B) geometric mean; (C) individual replicating virus (sgRNA) and the (D) geometric mean for lung (BAL samples) are shown. Values below the detection level are highlighted in yellow.
DISCUSSION
We designed our hAd5 S-Fusion + N-ETSD vaccine with the objective of generating vigorous T-cell responses so that the immune protection provided would be both broad and durable. Our vaccine is distinguished from other vaccines in development by (i) the inclusion of N, a SARS-CoV-2 antigen known to be highly effective in eliciting T cell responses (16–18); (ii) the use of an hAd5 [E1-, E2b-, E3-] platform that is effective even in the presence of existing adenoviralimmunity; (iii) the modification of N to target it to the endo/lysosomal pathway to maximize CD4+ T-cell responses (6); and (iv) the development of a thermally-stable oral formulation to overcome cold-chain limitations.
In the rhesus macaque NHP model, this bivalent hAd5 S-Fusion + N-ETSD subcutaneous and oral vaccine provided complete protection of nasal and lung airways against SARS-CoV-2 challenge by eliciting neutralizing antibodies plus Th1 dominant N- and S-specific T-cell responses. Following SARS-CoV-2 challenge, viral replication was immediately inhibited by Day one post-challenge in both nasal passages and lung with no further growth and complete clearance within 7 days of challenge in all (10/10) animals. The immediate inhibition of viral replication by Day 1 post-challenge in vaccinated NHP compared to placebo suggests the presence of SARS-CoV-2-specific cytotoxic T cells that rapidly cleared infected cells. This rapid viral clearance by the vaccine that confers comprehensive B- and T-cell immunity also implies that shedding of live viruses may be attenuated as a result of vaccination with hAd5 S-Fusion + N-ETSD, pointing to its potential to reduce transmission of virus by already infected individuals.
The striking, immediate inhibition of viral replication by the hAd5 S-Fusion + N-ETSD vaccination is in contrast to the reported findings for other adenovirus-vectored S-only vaccine NHP studies (9, 10), wherein there was evidence of continued viral replication. Even when challenged with a high titer of 1 × 106 TCID50/mL, vaccinated animals in our study appeared to be protected from the earliest time point assessed. On Day 1, this rapid protection and clearance was particularly evident in the lung, where both viral load and viral replication were ~1-2 logs lower than placebo in both vaccinated groups.
These findings present the intriguing possibility that the hAd5 S-Fusion + N-ETSD vaccine may be useful not only as a prophylactic to prevent infection and disease, but also as a therapeutic for individuals already infected. This hypothesis remains to be tested in ongoing clinical trials.
Our data demonstrate that all NHP immunized with the hAd5 S-Fusion + N-ETSD vaccine generated anti-S IgG and that 9 of 10 showed inhibition in cPass™ and thus the presence of anti-S RBD neutralizing antibodies (nAbs). Of interest, the one macaque that did not show inhibition in cPass™ still demonstrated complete control of viral replication in the nasal passage 4 days postchallenge and clearance in the BAL one week post-challenge. Although cellular correlates of immune protection against SARS-CoV-2 have not been fully elucidated, this single finding of viral clearance in the absence of high levels of neutralizing antibodies suggests the presence of other vaccine-generated SARS-CoV-2 specific antibodies or T cells that may have controlled the infection in the absence of anti-S RBD nAbs. This possibility is consistent with the findings of the critical role of T cells in protection from infection (19) in COVID-19 convalescent patients where SARS-CoV-2 specific T cells were identified even in the absence of antibody responses (18). Further characterization of the vaccine-elicited cellular and antibody responses is needed to fully understand the role of each immune compartment in reduction of viral load and attenuation of viral replication.
The generation of humoral and CMI responses that conferred protection of the respiratory tract from viral challenge indicate the efficacy of the hAd5 S-Fusion + N-ETSD vaccine when delivered by an SC prime followed by thermal-stable oral capsule boosts and reveal the potential for this vaccine to be developed for global distribution. The oral hAd5 S-Fusion + N-ETSD would not require ultra-cold refrigeration like many COVID vaccines currently under review for approval or in Phase 3 studies. Dependence on the cold-chain for distribution to geographically remote or under-developed areas causes shipping and storage challenges and will likely reduce the accessibility of the soon-to-be distributed COVID-19 vaccines. The possibility of the thermally-stable oral hAd5 S-Fusion + N-ETSD vaccine to act as a boost to other previously administered vaccines (a universal boost) warrants further exploration. The hAd5 S-Fusion + N-ETSD subcutaneous prime/boost vaccine has completed Phase 1 clinical trials and Phase 2/3 trials are actively recruiting. The thermally-stable oral vaccine will enter Phase 1 trials as a prime and boost, as well as explored to provide a boost to subcutaneous hAd5 S-Fusion + N-ETSD vaccination.
SUPPLEMENTARY MATERIAL
Methods
Safety assessments
NHP in all groups were observed twice daily from study Day −7 until the end of the study on Day 70 for clinical signs, including but not limited to anorexia (weights were taken), hunched posture, lethargy, respiratory distress, activity (recumbent, weak, or unresponsive), convulsions, and other abnormal clinical observations. Blood was collected from a femoral artery or vein, saphenous vein, or appropriate vessel of anesthetized animals at baseline, and Days 14, 21, 28, 35, 42, 56 (challenge Day), 57, 59, 61, 63, and 70 (End Study). Collected blood was used for clinical chemistry and hematological analyses as well as isolation of PBMCs.
Statistical analysis
For comparison of animals in groups, One-Way ANOVA was used with an appropriate post-hoc analysis (for example, Tukey’s or Dunnett’s) for specific comparisons. For head-to-head analyses, a two-tailed unpaired Student’s T-test was used. For all statistical analyses, *p <= 0.05, **p < 0.01, ***p<0.001, and ****p<0.0001. All statistical analysis was performed using GraphPad Prism 9 software.
ELISA for anti-Spike IgG
IgG against recombinant spike protein in NHP sera or plasma was determined using an Enzyme-Linked ImmunoSorbent Assay (ELISA) wherein 96 well EIA/RIA plates (ThermoFisher, Cat# 07-200-642) were coated with 50 μL/well by a 1 μg/mL solution of purified recombinant SARS-CoV-2-derived Spike protein (S-Fusion; ImmunityBio, Inc.) suspended in coating buffer (0.05 M carbonate-bicarbonate, pH 9.6) and incubated overnight at 4°C. Plates were washed three times with 150 μL of TPBS solution (PBS + 0.05%Tween 20) then 100 μL/well of blocking solution (2% non-fat milk in TPBS) was added and incubated for 1 hour at room temperature (RT). Plasma and serum samples were heat-inactivated at 56°C for 1 hour before use and 1:50 dilutions were prepared in 1% non-fat milk in TPBS. Plates were washed as described above and 50 μL/well of each dilution was added to the plate and incubated at RT for 1 hour. Plates were washed three times with 200 μL of TPBS before addition of 50μL/well of a 1:6000 dilution of HRP-conjugated, cross-absorbed goat anti-monkey IgG (H+L) secondary antibody (ThermoFisher, Cat# PA1 - 84631) in 1% non-fat milk/TPBS and incubation for 1 hour at RT.
Plates were then washed three times with 200 μL of TPBS and 50 μL of 3,3’,5,5’-tetramethylbenzidine (TMB) substrate (VWR, Cat# 100359-156) was added to each well and incubated at RT for 10 minutes. The reaction was stopped by addition of 50 μL/well of 1N sulfuric acid (H2SO4). The optical density (OD) at 450 nm was measured using a Synergy 2 plate reader (BioTek Instruments, Inc) and data is analyzed using Prism 8 (GraphPad Software, LLC).
cPass™ for determination of the presence of neutralizing antibodies
The presence of neutralizing, anti-spike antibodies in sera from all NHP was determined by assay of sera collected on Days 0, 14, 21, 28, 35 and 42 using the surrogate virus neutralization assay, cPass™ (23). The assay is based on assessment of inhibition of binding of the spike receptor binding domain (RBD) to its human host receptor (in the assay, recombinant) angiotensin converting enzyme 2 (ACE2), with inhibition above 20% being correlated with a level of anti-S antibody that is predicted to be neutralizing. All sera samples were diluted 1:30.
ELISpot for assessment of cytokine secretion
ELISpot assays were used to detect cytokines secreted by fresh peripheral blood mononuclear cells (PBMCs) isolated from the blood of NHP study animals. PBMCs were isolated from whole blood by standard density gradient centrifugation and frozen in liquid nitrogen until use. PBMCs were thawed and re-suspended in RPMI 10% human AB serum, then pulsed with 2 μg/ml of SARS-CoV-2 S or N peptide pools (JPT Peptide Technologies catalogue # PM-WCPV-S and PM-WCPV-NCAP-1 respectively). Negative and positive controls were cells cultured with media alone or phorbol myristate acetate (PMA, 50 ng/ml) and ionomycin (1 μg/ml), respectively. For IFN-γ assessment, PBMCs were cultured (17 hours at 37°C) in a microtiter plate (Millipore catalogue # MAIPS4510) containing an immobilized primary antibody to capture NHP-specific IFN-γ (MabTech catalogue # 3421M-3-1000). IFN-γ was detected by a secondary antibody to human IFN-γ conjugated to biotin (MabTech catalogue # 3420-6-250). A streptavidin/horseradish peroxidase conjugate (Thermo Fisher catalogue # 21126) was used detect the biotin-conjugated secondary antibody. IFN-γ spot development was completed using a peroxidase substrate kit (Vector Labs catalogue # SK-4200). The number of spots per well (3.5 × 105 PBMCs) was counted using an ELISpot plate reader. IL-4 was assessed using a commercial ELISpot kit (MabTech catalogue # 3410-APW-2) using the manufacturer’s instructions. Numbers for graphing were adjusted to spot-forming cells per 106 PBMCs.
Determination of viral load and viral replication post-challenge
RT-qPCR assays were performed to quantify total SARS-CoV-2 RNA copies including genomic RNA using the nucleocapsid protein gene as a target or subgenomic RNA copies that are replication intermediates of the virus using the envelope protein (E) gene as a target. These assays were performed to quantify viral loads following SARS-CoV-2 challenge. RNA was isolated from swabs and bronchioalveolar lavage fluid using the Indispin QIAcube HT Pathogen Kit (Indical Bioscience, Germany) on the QIAcube HT instrument (Qiagen, Germany). The isolated RNA was then evaluated in RT-qPCR using the TaqMan Fast Virus 1-step Master Mix (Thermo Fisher Scientific) on a QuantStudio Flex 6 Real-Time PCR System (Applied Biosystems; Foster City, CA). The primers and probe for total SARS-CoV-2 RNA quantitation were specific to the nucleocapsid protein gene, corresponding to the N1 sequences from the Centers for Disease Control and Prevention (CDC) 2019-Novel Coronavirus (2019-nCoV) Real-Time RT-PCR Diagnostic Panel (https://www.cdc.gov/coronavirus/2019-ncov/lab/rt-pcr-panel-primer-probes.html) except that the probe quencher was modified to Non-Fluorescent Quencher-Minor Groove Binder (NFQ-MGB) (Thermo Fisher Scientific). The primers and probe for the subgenomic RNA quantitation were specific to the E gene subgenomic RNA (Integrated DNA Technologies, Iowa) (28). A standard curve comprised of synthetic RNA containing the corresponding target sequence from SARS-CoV-2 isolate WA1 sequence (GenBank Accession Number MN985325.1) (Bio-Synthesis, Inc.; Lewisville, TX) was included on each PCR plate for absolute quantitation of SARS-CoV-2 RNA copies in each sample. Thermocycling conditions were: Stage 1 - 50°C for 5 min for one cycle; Stage 2 - 95°C for 20 sec for one cycle; Stage 3 - 95°C for 3 sec and 60°C for 30 sec for 40 cycles. Data analysis was performed using the QuantStudio 6 software-generated values (total copies per well of each sample) and additional calculations to determine SARS-CoV-2 RNA or subgenomic RNA copies per mL of fluid.
Supplementary Results
No clinical signs of toxicity were seen
No clinical signs were noted during the twice daily observations for clinical signs of toxicity due to vaccination and no animals died during the two weeks after one subcutaneous immunization of 1x 1011 VP or a week after an oral booster of 1×1010 IU of hAd5-S-Fusion+N-ETSD. No gross pathological effect or adverse events were observed and there were no notable changes in body weight (Figure S1).

Body weights are shown in Kg; note y-axis range is 4-6 Kg.
Hematology
Hematological parameters evaluated in Group 1 and 2 hAd5 S-Fusion + N-ETSD and Group 3 placebo control animals are presented in Table S1. Only hemoglobin was consistently slightly lower in Group 2 NHP as compared to Group 3 placebo for all post-vaccination time points.
Hematological parameters in hAd5 S-Fusion + N-ETSD vaccinated Group 1 and Group 2 NHP versus placebo Group 3.
Lymphocytes
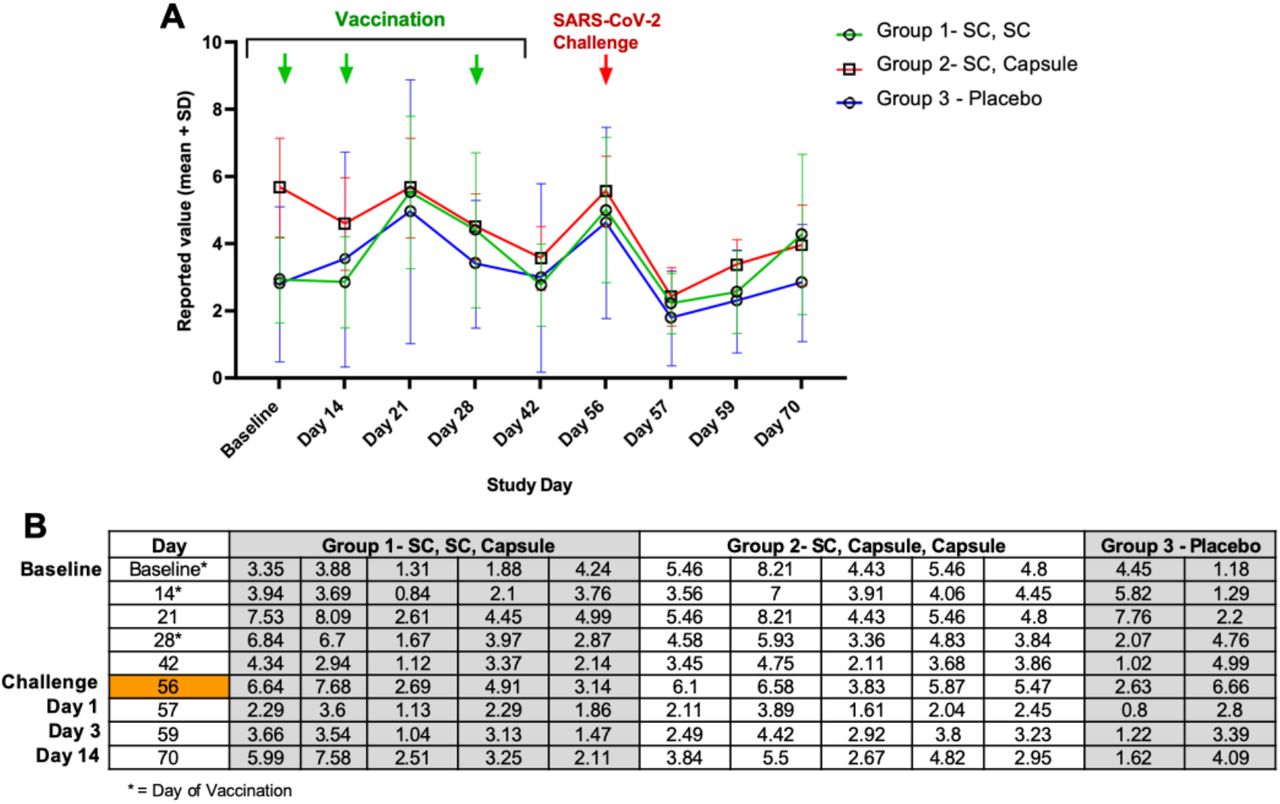
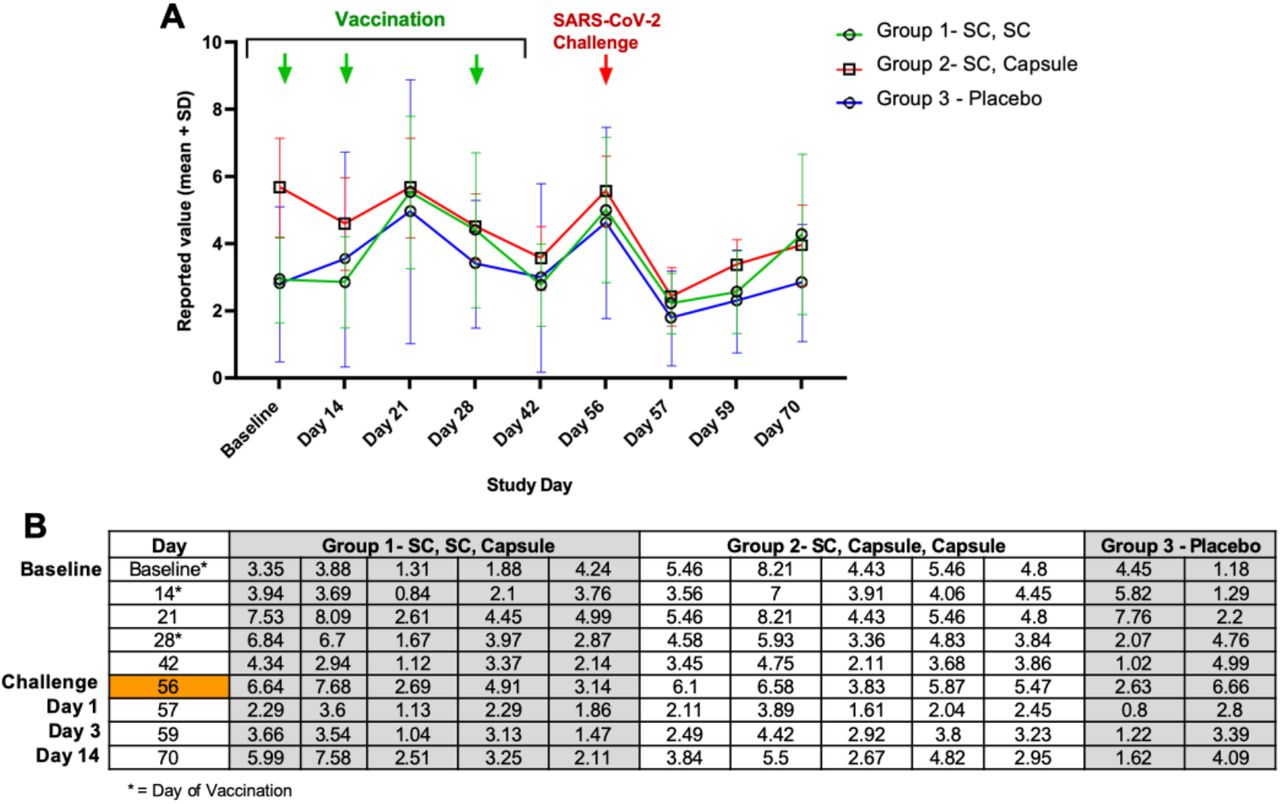
The reported values for lymphocytes from Study Day 0 to 70 are shown below in Figure S2. Lymphocyte levels were varied at baseline, being higher in NHP entered into Group 2, but lymphocytes dropped in Group 2 animals by Day 14 (a vaccination Day). Lymphocytes showed a trend to increase in all groups between Day 14 and Day 21, and then became lower on Days 28 and 42. NHP were moved to the BSL3 facility on Day 42 and on Day 56 (the day of challenge) mean lymphocyte levels were similar all groups. By Day 57 (also referred to as Day1 postchallenge in the main text), lymphocyte levels decreased in all groups. Overall, lymphocytes increased on Days 59 and 70, at which time mean levels were higher in vaccinated NHP.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
(A) Mean lymphocyte levels are graphed for Groups 1, 2, and 3. (B) Table of lymphocyte levels for individual NHP. NHP were vaccinated at Baseline after collection of blood used for lymphocyte determination. NHP received boosts on Days 14 and 28. On Day 42, NHP were transferred to the BSL-3 facility and on Study Day 56, were challenged with SARS-CoV-2 after collection of blood. All animals were euthanized on Day 70.
Clinical Chemistry
Only creatinine was consistently lower in Group 1 NHP on five time points and at two time points for Group 2 (Table S2), but it was lower on Day 0 baseline, indicating this was not vaccine-related.
Clinical chemistry for hAd5 S-Fusion + N-ETSD vaccinated Group 1 and Group 2 NHP as compared to placebo Group 3.
ACKNOWLEDGEMENTS
We would like to thank the Battelle group who performed the study. We would also like to thank the DHHS, NIH/NIAID and ASPR/BARDA for funding the study.
REFERENCES




