

Abstract
AROMATASE, encoded by the CYP19A1 gene, is the cytochrome enzyme responsible for the synthesis of estrogens in vertebrates. In most mammals a peak of expression of the CYP19A1 gene occurs in the fetal XX gonad when sexual differentiation starts up. To elucidate the role of this peak, we produced 3 lines of TALEN genetically edited CYP19A1 KO rabbits, that were void of any production of estradiol. All KO XX rabbits developed as females, with aberrantly small sized ovaries at adulthood, an almost empty reserve of primordial follicles and very few large antrum follicles. Ovulation never occurred. Our histological, immunohistological and transcriptomic analyses showed that the surge of estradiol in the XX fetal rabbit gonad is dispensable for its determination as an ovary, or for meiosis. However, it is mandatory for the high proliferation and differentiation of both somatic and germ cells, and consequently for the establishment of the ovarian reserve.
In Vertebrates, AROMATASE is the unique enzyme responsible for the synthesis of estrogens. It converts specifically three androgenic substrates (androstenedione, testosterone and 17alpha-testosterone) to estrone, estradiol, and estriol respectively. It is the product of the CYP19A1 gene, which is the unique member of its gene family. Although the CYP19A1 gene is commonly known to be mainly expressed by gonads in adults, a physiologically significant expression has already been reported in numerous tissues or organs such as bones, adipose tissue, brain or placenta (the expression in the latter depending on the species, see for review in (Conley and Hinshelwood 2001) and references herein).
In mammals, the expression of the CYP19A1 gene begins early at around implantation in the embryo in rabbits (Dickmann et al. 1975; George and Wilson 1978b), pigs and ruminants (Gadsby et al. 1980). A second burst of expression was reported later in fetal life in the ovaries shortly after the sex determination process. This was described in rabbits (George et al. 1978; George and Wilson 1978b; Gondos et al. 1983; Daniel-Carlier et al. 2013), sheep (Mauleon et al. 1977; Payen et al. 1996; Torley et al. 2011), goats (Pannetier et al. 2006), bovines (Shemesh 1980; Ross et al. 2009; Garverick et al. 2010) and humans (George and Wilson 1978a) by RNA measurements or by estrogen assays in gonads or gonadal culture media. This peak is transitory and spans until meiosis initiation. Later on, CYP19A1 gene expression increases progressively with follicle differentiation. It culminates in antrum follicles, the mural granulosa cells being the site of expression. By the past, the physiological significance of this fetal peak of expression has been searched in the rabbit species. Namely, George et al (George et al. 1978) have supposed that the local production of estrogens in the XX gonad was a key event for the initiation of the differentiation of the female fetal gonad but this hypothesis has still not been proven. In birds (Elbrecht and Smith 1992; Vaillant et al. 2001) and fishes (Guiguen et al. 2010), it was shown that estrogens are major determinants of the ovary orientation of the undifferentiated gonad in fetal or larval life. The question was thus to investigate the role of the fetal peak of estrogen in the mammalian gonad and more specifically to search whether in mammals it is mandatory for ovary differentiation.
To reach this aim, we decided to suppress the expression of the CYP19A1 gene in a mammal species other than the mice. Indeed, rodents represent some of the few mammalian species without detectable peak of expression at this developmental stage in the fetal XX gonad i.e. before meiosis. Thus, thanks to the TALEN methodology already shown to be efficient for gene targeting in the rabbit species (Lee et al. 2018; Peyny et al. 2020), we produced CYP19A1 knocked-out (KO) rabbits completely devoid of estrogens secretion. Here we report the phenotype of the XX CYP19A1 KO (ARO−/−) female rabbits, which are completely infertile at adulthood due to ovarian developmental defects detectable as soon as the first stages of their differentiation.
Results
Three aromatase mutant rabbit lines which do not produce estradiol were generated
The TALEN subunits were designed to generate InDel mutations through non-homologous end joining near the initiation site of translation of the CYP19A1 gene as described in Figure 1. After transferring 67 injected embryos in 3 recipient female rabbits, 6 founders (2 males and 4 females) harboring mutant alleles were characterized from 11 neonates. Each founder F0 was mated with a wild type counterpart in order to transmit a mutant allele to the descendants and generate mutant lines carrying one mutated allele per line.

(A): structure of the rabbit aromatase gene, with the various tissue specific promoters i.e. the I.1 placental promoter at around 80kb upstream of the transcription start point (tsp, marked by squared arrows) and the two proximal ovarian promoters I.r and II as previously described (Bouraïma et al. 2001). The enlargement shows the sequence of the exon II with the initiation site of translation (ATG in green letters). The two subunits (left and right arm) of the TALEN are shown as green symbols and the targeted sequence is written in red. The horizontal black arrows point the primers used for PCR detection of mutants in founders (F0/R0 set) and for routine qPCR genotyping (F1/R1 set).
(B): sequences of the mutant alleles Δ339, Δ498 and Δ829. Capital letters report to exon sequence, lowercase letters to intron sequence. The deletion spans over 339 nucleotides (nt) in the Δ339 mutant allele (149 nt in the 3’ end of exon II and 190 nt of intron 2) and over 498 nt in the Δ498 mutant allele (148 nt in the 3’ end of exon II and 350 nt of intron 2), both with insertion of few nt at the repair position. In the Δ829 allele, the mutation consisted in the elimination of 829 nt encompassing 250 nt upstream of the beginning of exon II (transcription start site of the gene as regard to the ovarian promoter), the totality of the second exon (263 nt) and 316 nt in the second intron. All mutations suppressed the ATG codon and the splice donor site at the 3’ extremity of exon II.
Three mutant lines harboring each one a specific mutant allele were generated and studied. Homozygous (ARO−/−) rabbits were generated by crossing heterozygous (ARO+/−) rabbits. Mating was performed between rabbits within each line in order to avoid mixing the various mutant alleles.
Mutations are shown in Figure 1. A 339, 498 or 829 nucleotide-long (nt) deletion was observed in each of the alleles respectively named Δ339, Δ498 and Δ829. Insertion of a small number of nucleotides was also detected in Δ339 and Δ498 alleles. Briefly, mutations Δ339 and Δ498 eliminated the 3’ end of exon II including the translation initiation site (ATG codon) plus a large part of the second intron; the mutation Δ829 eliminated a larger fragment encompassing the proximal promoter upstream of exon II, the whole exon II and a large part of the second intron. Thus, in the absence of the translation initiation site, no protein was expected to be translated from any of these mutant alleles. Moreover, the deletion of the 3’ part of the exon II theoretically should lead to the elimination of 48 amino acids in the N-terminus of the AROMATASE enzyme. As previously published (Kaur and Bose 2014), these amino acids encompass the signal anchor responsible for the translocation of the protein to the endoplasmic reticulum. It is thus expected that if a protein is translated from the mutant mRNA, it has a good chance to be not correctly embedded in the endoplasmic reticulum bilayer, and consequently not functional.
Finally, to confirm the absence of any active AROMATASE enzyme, we searched for estradiol by Gas Chromatography / Mass Spectrometry (GC/MS) in rabbit serum samples collected after birth and in fetal gonads. The concentrations of estradiol in sera from ARO−/− XX females after birth were under the limit of quantification (LOQ = 0,2pg/ml) whatever the age of the animals, when in wild type (ARO+/+) and heterozygous (ARO+/−) females the estradiol concentration increased progressively with the age of the animal (supplementary Figure 1). The concentration of testosterone was measured in the same samples. In ARO−/− sera, the values ranged in similar way than in WT or ARO+/− XX rabbits.
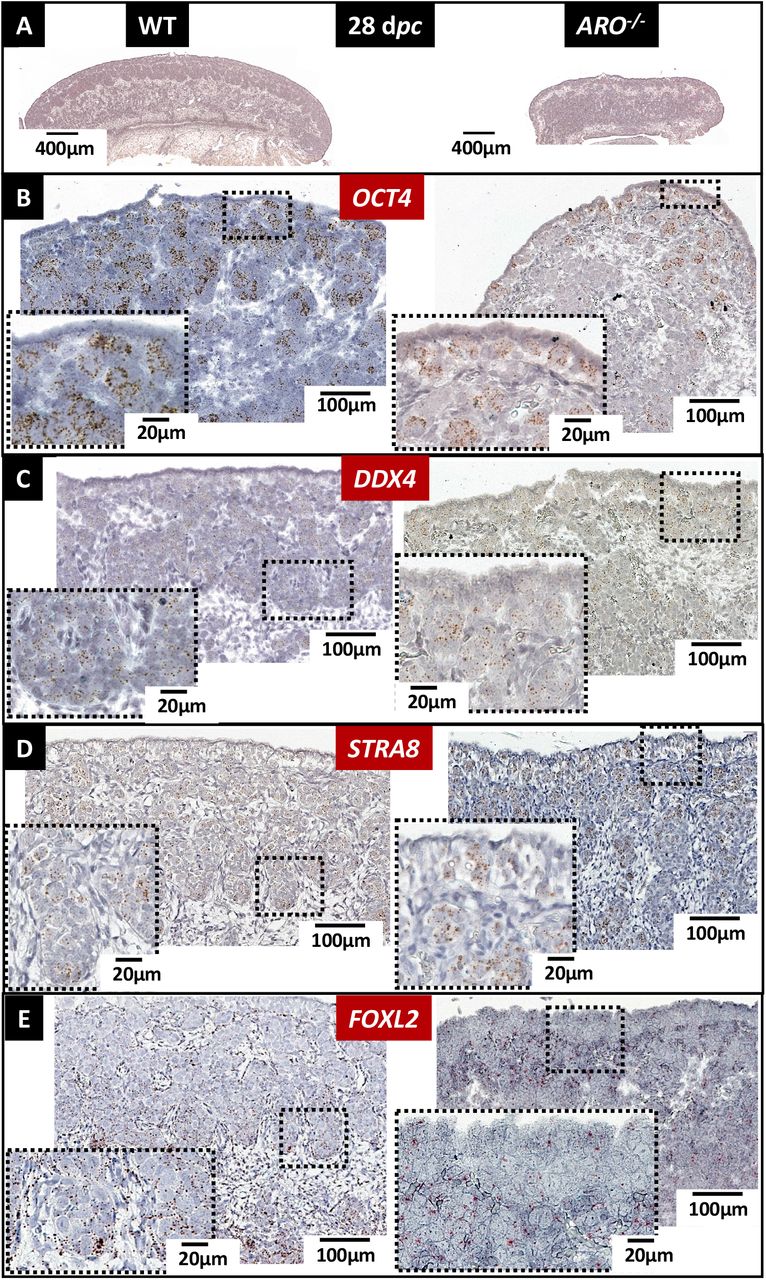
We measured also by GC/MS the steroid content of the gonads in fetuses aged of 28 days post coitum (dpc). The estradiol content was undetectable in gonads from all ARO−/− XX fetuses and in all XY gonads, when it was always detected, even sometimes in very low amounts, in XX gonads from ARO+/− and WT genotypes (supplementary figure 2). Testosterone was not detected in any XX gonads whatever their genotype at the CYP19A1 locus. This shows that androgens were not over-synthesized nor accumulated in XX ARO−/− gonads. Finally, these data also show that neither estradiol nor testosterone from maternal or placental origin was detected in the 28dpc fetal gonads.
XY and XX ARO+/− rabbits were fertile, with normal genital tracts and normal gonads morphology
In the three mutant lines, all ARO+/− and XY ARO−/− rabbits were viable until adults and did not suffer apparently from any metabolic or postural disease at least until 2 years after birth. No apparent modification of external genitalia was visible. All were fertile and gave birth to progeny. The sexual behavior was not studied. The number of neonates per litter from heterozygous mating (♀ARO+/− × ♂ARO+/−) was similar to that of wild type mating (♀WT × ♂WT) (supplementary Figure 3). The percentage of mating without pregnancy was not different: 37,9% (11 null per 29 mating) in WT x WT mating and 35,3% (23 null per 65 mating) in heterozygous x heterozygous mating. All this suggests that the fertility of heterozygous animals was not affected.
At puberty (5-6 months after birth), genital tracts and gonads were not different in heterozygous and wild type XX rabbits (supplementary Figure 4). Since all measured parameters did not differ in heterozygous and wild type females (similar estradiol and testosterone serum levels, similar prolificacy, similar overall structure of genital tracts and ovaries), we used ovary samples indifferently from wild type or heterozygous females as reference ones. Nevertheless, the genotype has always been distinctly reported in the legends of all figures.
XX ARO−/− rabbits developed small female genital tracts, ovaries with almost no follicular reserve and very few antral follicles
All XX ARO−/− rabbits were viable without any metabolic or postural disease at least until 1.5 years after birth. No apparent modification of external genitalia was visible. The sexual behavior of XX ARO−/− rabbits was not studied in detail.
Obvious modifications of the genital tracts and ovaries were observed in XX ARO−/− rabbits from the three lines with similar characteristics. After puberty, a typical phenotype was observed, with under-developed uterus horns, oviducts and pavilions resembling those of neonatal females. Their size was clearly smaller than in wild type or heterozygous females (supplementary Figure 4). In all three lines, XX ARO−/− gonads developed like ovaries (Figure 2, supplementary Figure 5) but with a very small size. As the phenotype was identical in Δ339, Δ498, and Δ829 lines, it is hardly attributable to any off-target alteration of the genome. Ovaries contained almost no primordial follicles, scarce primary and secondary follicles, and were filled with numerous remnants of atretic follicles. The abundant follicle reserve composed of primordial follicles observed at the cortex of wild type ovaries (Figure 2A and B) was absent in ARO−/− gonad mutants (Figure 2C and D). Nevertheless, some follicles developed to reach the large antrum stage in ARO−/− ovaries, showing granulosa and theca cell layers, oocyte and cumulus. The number of large antrum follicles was high in some females (see the ovary of the Δ829 female in supplementary Figure 5) possibly reflecting the surge of gonadropins that occurs at early puberty (around 5-6 months in the rabbit). Numerous Call-Exner bodies (spherical deposits of basal lamina matrix surrounded by granulosa cells, observed in rabbits and some mammals) were observed in the granulosa of growing follicles in ARO−/− ovaries as classically observed in wild type ones (Lee et al. 1996). The number of pyknotic cells or of cells with dense colored nuclei was low.

Ovaries from wild type female rabbits (ARO+/+, A and B) and from the mutant ARO−/− line Δ339 (C and D) were fixed in PAF then HES colored. Females were 5,5 months old. Two distinct sections of the same ovary are presented for the mutant ARO−/− to show the follicles that were otherwise not visible. Enlarged zones point the different types of follicles. Black arrows = Call-Exner bodies. Black stars = accumulation of fibrous tissue. Red stars = remnants of degenerated follicles. “p” = primordial follicle; I = primary follicle; II = secondary follicle; IC = interstitial cells. The phenotype of the ARO−/− ovary was similar in the other lines Δ498 and Δ829 (supplementary Figure 5).
The expression of the marker genes RSPO2 and FOXL2 specific respectively for the oocyte and for the granulosa cells in differentiating follicles was studied by in situ hybridization (ISH, Figure 3). In the ARO−/− ovary, the rare detected oocytes were positive for the RSPO2 labelling (Figure 3A), and granulosa cells were positive for FOXL2 (Figure 3B). In wild type ovaries, AROMATASE was immuno-detected in a small number of follicles with antrum, thus characterizing the pre-ovulatory ones (Figure 3C). The Anti Müllerian Hormone (AMH) was immuno-detected in granulosa cells of many follicles in wild type ovaries except those positive for AROMATASE. In ARO−/− ovaries, granulosa cells of all follicles were AROMATASE negative and AMH positive (Figure 3D). However, the AMH serum level was lower in ARO−/− females, reflecting the small number of follicles and/or their poor synthesis capacity.

In Situ Hybridization to localize mRNA of RSPO2 and FOXL2 genes and immunodetection of AROMATASE and AMH in adult ovaries. Ovaries were collected in wild type (WT) or ARO−/− Δ329 females aged 5-6 months. The expression of RSPO2 (A) and FOXL2 (B) genes was detected by ISH probes. AROMATASE (C) and AMH (D) were detected by immunohistochemistry on two adjacent sections. The RSPO2 and FOXL2 probes labelled respectively the cytoplasm of oocytes (brown dots) and of granulosa cells (red dots) from all follicles, from primordial to antrum stages in wild type and ARO−/− rabbits as well. Some thecal cells were also positive for the FOXL2 labelling in both genotypes. Mural granulosa cells from pre-ovulatory follicles were positively labelled by the anti-AROMATASE antibody in wild type ovaries (brown colored); no cells were positive in ARO−/− ovaries. The anti-AMH antibody labelled most granulosa cells of growing follicles in wild type ovaries (brown colored), excepted those positive for AROMATASE. In ARO−/− ovaries, all follicles with antrum were positively labelled. Similar staining were observed in females from the two other strains. The graph represents the AMH serum level in WT (black points), ARO+/− (green points) and ARO−/− (red points) females.
Intense modifications affected the medulla of the ovary. In wild type rabbits, the medulla consisted mostly in interstitial tissue composed of cells with large clear cytoplasm and round regular shape nuclei (Figure 2B); besides, in ARO−/− ovary, the interstitial tissue appeared as a mixture of cells with heterogeneous shape and angular nuclei. More, several areas filled with fibrous material rich in collagen and devoid of cells were detected (Figure 2C and D; supplementary Figure 5).
Some of the heterozygous (ARO+/−) and homozygous (ARO−/−) 10 months old XX rabbits were hormonally treated to induce ovulation, mated and sacrificed to collect gonads for histological analysis. Ovulation rupture points were observed in ovaries from ARO+/− rabbits, but not in ARO−/− ovaries (supplementary Figure 6). While several primordial, primary, pre-antral and antral follicles were detected at the cortical periphery of ovaries in ARO+/− rabbits after super-ovulation treatment, no primordial or primary follicles were seen in the treated ARO−/− ovarian cortex. Nevertheless, some large antrum type follicles were observed in ARO−/− ovaries, with developed granulosa, theca cells, and oocyte.
The mutation impacted the XX gonad from the first stages of their differentiation
Our goal was to search when and how ovaries were impacted by the lack of functional AROMATASE enzyme and of estradiol during fetal life. In the rabbit species, pregnancy spans for 31 days. The present study analyzed the fetal gonads from 18 dpc onwards. At this stage in rabbits, the gonadic sex is already determined, the multiplication of germ cells and somatic cells is intense (Chretien 1966) and the peak of CYP19A1 gene expression starts in the ovary, concomitantly with the increase of FOXL2 gene expression (Daniel-Carlier et al. 2013). At 20 dpc, we detected a specific labelling with a CYP19A1 ISH probe in cells of the surface epithelium and cells forming invaginated structures issued from this epithelium (supplementary Figure 7). Simultaneously, we observed that these same cells were labelled by an ESR1 specific ISH probe and we detected a surge of the expression of the estrogen receptor alpha gene (ESR1) by RT-qPCR (supplementary Figure 7). The FOXL2 gene was mainly expressed in somatic cells underneath the surface epithelium and those surrounding invaginated structures (ISH labelling, supplementary Figure 7). Only few cells of the surface epithelium were clearly positive for both the CYP19A1 and FOXL2 ISH probes. The expression of the ESR2 gene coding for the estrogen receptor beta was low at these stages compared with the levels determined later at birth (34 dpc). In agreement, a faint labelling was observed by ISH labelling (supplementary Figure 7).
Therefore, the morphology of the gonads and the location of specific markers were investigated at various fetal stages and compared between wild type or heterozygous gonads versus ARO−/− ones from the stage of gonadal determination onwards. At 18 dpc, the gonads from all XX fetuses, ARO−/− and ARO+/−, were differentiated as ovaries with the same morphology except a thinner coelomic epithelium in ARO−/− gonads (Figure 4A). Regardless of the developmental stages, XX ARO−/− gonads showed no evidence of testicular appearance (see for comparison the 18 dpc testis in the supplementary Figure 8). A similar density of OCT4 positive germ cells was detected in ARO−/− and ARO+/− gonads, as shown by the labelling with specific antibodies directed against OCT4 (Figure 4B). Mitotic activity was detected by KI67 immunostaining (Figure 4C). KI67 positive cells were mainly present in the ovarian surface facing the abdominal cavity and in invaginated structures below in both ARO+/− and ARO−/− ovaries. Interestingly, the KI67-positive ovarian surface appeared thinner in KO gonads than in the ARO+/− controls (Figure 4C).
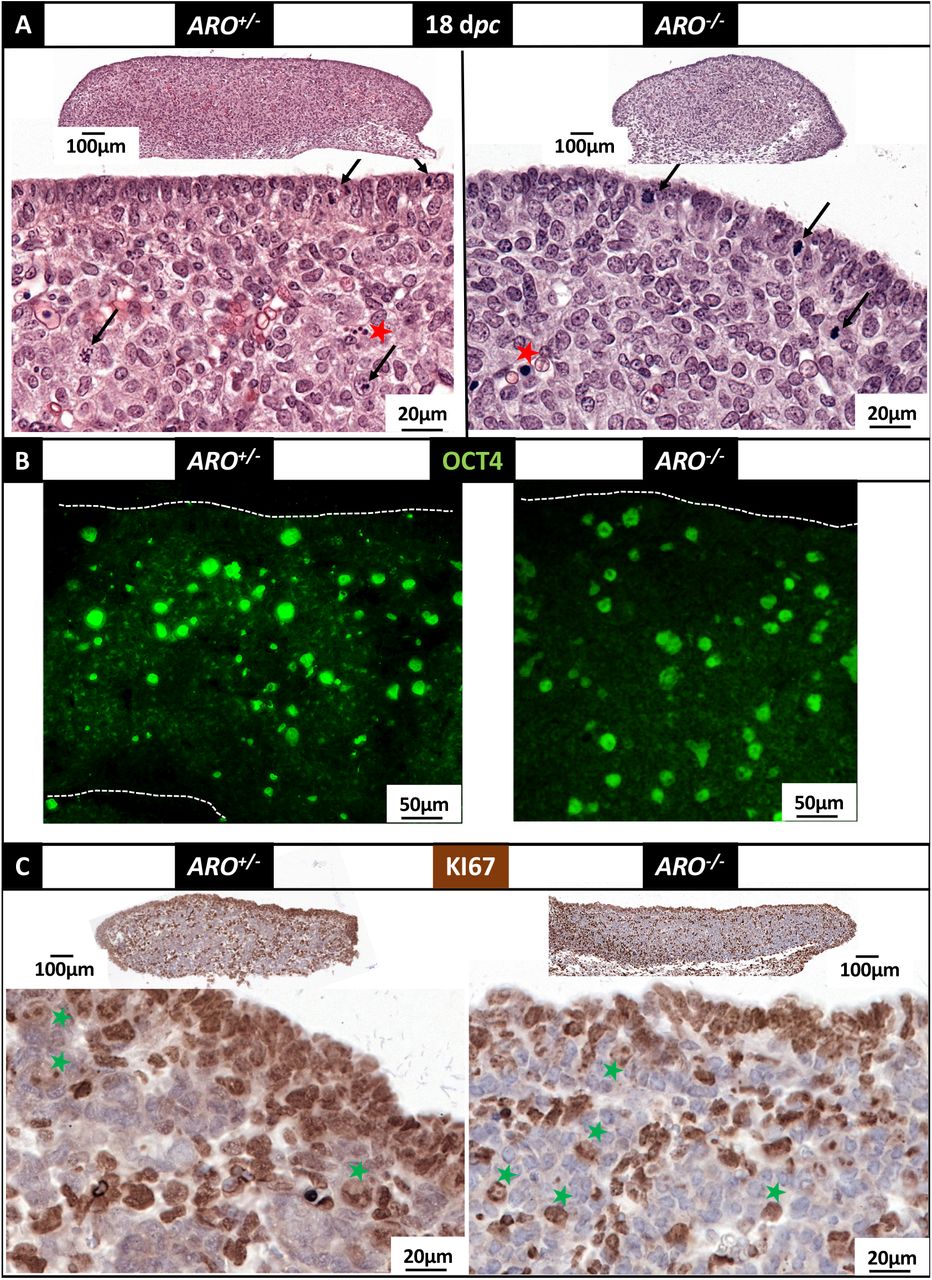
Gonads were collected at 18 dpc in ARO+/− and ARO−/− fetuses, fixed in Bouin’s then HES colored (A) or fixed in PAF then treated for antibody labelling (B and C). (A): HES coloration. The surface epithelium appeared as a continuous layer of epithelial cells. Red stars point dense nuclei that could correspond to pyknotic nuclei. Arrows point figures of mitoses. (B): detection of OCT4 positive cells. The immune-labelling was visualized using a fluorescent secondary antibody. Nuclei of positive cells (large round green-labelled) were dispersed through the gonad in both ARO+/− and ARO−/− ovaries. (C): detection of KI67 positive cells. The immune-labelling was visualized using a peroxidase coupled secondary antibody. Positive cells with brown colored nuclei were found at the surface epithelium, and in rows of cells delimiting cell clusters. Green stars point brown light-colored large nuclei, possibly corresponding to germ cells.
From 20 dpc onwards, more important alterations progressively appeared in ARO−/− ovaries. The cortex was thinner, composed of a single layer of epithelial cells, connected to small size nascent ovigerous cords, by opposition to the thick cortex observed in ARO+/− ovaries, connected to numerous large size ovigerous cords (Figure 5A and 5D). A discontinuous layer of cells with small and dense colored nuclei was observed underneath the surface epithelium, forming at 22 dpc (Figure 5D) and later on (34 dpc, Figure 9) a loose connective tissue between the thin cortex and the inside of the ovary.

Gonads were collected from ARO+/− and ARO−/− fetuses, fixed in Bouin’s then HES colored (A, D). Ovigerous nests and cords are underlined by a dotted white line in A-F. The red double arrow points the thickness of the nascent cortex. Blue stars point the connective tissue. In situ hybridization was performed to visualize simultaneously two RNA targets to show the ESR1 (blue points) and FOXL2 (fast red points) RNAs (B, E), and the ESR1 (blue) and OCT4 (fast red) RNAs (C, F). The lower panel is the fluorescence observation of the fast red OCT4 ISH labelling to show the overall density of the OCT4 positive cells (G, H).
The cells of the surface epithelium expressed the ESR1 gene in both genotypes (Figures 5B and 5E). The intensity of the ISH ESR1 labelling varied greatly at the surface of the ARO+/− ovaries as shown in figure 5 (compare 5B, E, C and F, which are different sections from the same gonad). Nevertheless, in all treated sections, clusters of labelled ESR1 cells connected to the surface epithelium were easily observed. Besides, in ARO−/− ovaries, labelled ESR1 cells were mainly dispersed in the inner of the gonad. Similarly, the FOXL2 ISH probe labelled evenly dispersed cells in the inner of the ARO−/− gonad, which differed from the strong FOXL2 labelled cell cords of the ARO+/− ovary. OCT4 positive germ cells were detected in both genotypes but the hybridization signals were lighter in ARO−/− germ cells (Figures 5C and 5F-H). Numerous were found in the surface epithelium, interestingly in close contact with somatic cells expressing the ESR1 gene.
KI67 positive cells were overall detected in both types of ovaries at 22 dpc (Figure 6A) but there was still a clear difference of mitotic activity in the coelomic epithelium between ARO+/− and ARO−/− ovaries. Indeed, while many KI67 positive cell layers were detected in the coelomic epithelium of ARO+/− gonads, only one layer was visible in the ARO−/− one. The γH2AX-antibody, marker of double strand breaks of the DNA, labelled mostly germ cells in both types of ovaries as suggested by their large round nuclei (Figure 6B). The density of γH2AX positive cells (probably apoptotic cells) was apparently not different in ARO+/− and ARO−/− ovaries.

The marker of mitotic activity KI67 (A), and that of double strand breaks γH2AX (B) were localized by immunohistochemistry in sections from PAF fixed ovaries of ARO+/− and ARO−/− fetuses at 22 dpc. The KI67 antibody labelled nuclei of most cells of the coelomic epithelium, and numbers of cells inside the ovary in both genotypes. Cells with flat nuclei KI67 positive (red arrowheads) surrounded from place to place the ovigerous cords. Cells with large round nuclei positive for KI67 labelling (probably germ cells) were found mainly at the inside of the cords (green stars). The γH2AX antibody labelled large round shape nuclei corresponding probably to germ cells. Note the disconnection between the coelomic epithelium and the new forming ovigerous cords in ARO−/− ovaries.
The relative expression levels of key marker genes were determined at these developmental stages on total RNAs extracted from the whole gonads (Figure 7). The expression levels of the OCT4 and FOXL2 genes were reduced in ARO−/− ovaries, reflecting a reduction of the number of OCT4 positive germ cells and FOXL2 positive somatic cells respectively. Interestingly, the expression level of the DDX4 gene, known to be its lowest at this ovarian developmental stage (Daniel-Carlier et al. 2013), did not differ between WT and ARO−/−. It increased between 20 and 22 dpc, which reflects the early differentiation of germ cells from the pluripotent stage. The SOX9 gene expression level was 5- to 10 times lower in the ovaries from all genotypes than in testes at the same age. The SOX9 gene was not detected by immunohistology either in wild type or in ARO−/− ovaries, while a clear labeling was observed in the fetal testis at 22 dpc (supplementary Figure 8). This reinforces the demonstration that ARO−/− ovaries did not show any sex-reversal phenotype. The expression levels of the ESR1 and ESR2 genes did not change significantly.

Ovaries were collected from wild type and mutant fetuses. The diagrams show the relative mRNA expression levels. Each point represents one RNA sample extracted from both gonads of one animal at the indicated developmental stage. The same RNA samples were analyzed in the 6 diagrams. Red triangles refer to mutant from Δ339 line, and red point from Δ829 line. No samples from line Δ498 were analyzed. The blue point in the SOX9 diagram shows the SOX9 mRNA level measured in the two testes of one 20 dpc WT fetus. Horizontal bars represent the medians. At these stages DDX4 and ESR2 gene expression levels were very low, but significantly measured.
Thus, from 20 dpc onwards, the differentiation of the ARO−/− gonad was profoundly modified, with a decrease of proliferation of the somatic cells of the coelomic epithelium and a reduction of the number of germ cells. Finally this leads to a clearly underdeveloped small organ with small ovigerous nests, an abnormal distribution and differentiation of the FOXL2 positive cells and the spread of connective tissue underneath the surface epithelium.
Impact of estrogens deprivation on germ cell meiosis and follicular differentiation
In the rabbit ovary, the first signs of meiosis are histologically detected at 30-31 dpc, i.e. around birth. Later on, typical meiotic germ cells (prophase I) are observed for less than 2 weeks. After meiosis arrest at diplotene stage, follicles begin to differentiate. The peak of expression of the STRA8 gene that starts at 24-28 dpc (Daniel-Carlier et al. 2013) marks the commitment of germ cells to meiosis. Thus, we sought to analyze at 28 dpc (meiosis commitment), 34 dpc (beginning of meiosis) and few days or weeks after birth (folliculogenesis) whether initiation and arrest of meiosis I and early follicular formation were impacted in ARO−/− ovaries.
In wild type and ARO−/− 28 dpc ovaries, numerous germ cells were positively labelled by the OCT4, DDX4 and STRA8 ISH specific probes (Figures 8B, 8C, 8D). The STRA8 labelling proves the overall commitment of germ cells in meiosis in WT and ARO−/− ovaries. However, in these later, the number of positive germ cells for the three probes was low. More, they were clustered in small ovigerous-like structures, while they were all observed in large ovigerous cords in wild type ovaries. Positive FOXL2 labelled cells were also much less numerous in ARO−/− ovaries, as shown by the few numbers of hybridization labels. Nevertheless, some of these FOXL2 positive cells harbored a flattened shape and surrounded the small ovigerous-like structures embedding germ cells, as in wild type ovaries (Figure 8E).

Ovaries collected at 28 dpc were fixed in PAF, then HES colored or analyzed by ISH. (A): morphology of the ovaries. The pictures show the difference of the overall size of the gonad and the different thickness of the cortex. The OCT4 (B) and DDX4 (C) ISH probes labelled respectively pluripotent germ cells and differentiating ones. The STRA8 ISH probe (D) labelled germ cells committed to meiosis. The FOXL2 ISH probe (E) labelled the differentiating somatic cells. Note that the latter were mainly distributed around and inside the ovigerous cords in the WT ovaries, and in contrast were dispersed throughout the gonad in the ARO−/− rabbit. As in all ISH pictures, positive labeling appears as colored dots.
At 34 dpc, in wild type ovaries, most ovigerous cords were filled with numerous zygotene germ cells, thus showing that meiosis had begun in a majority of germ cells (Figure 9A and a1). In contrast, in the ARO−/− ovaries, germ cells at zygotene were sparse (Figure 9B and b1, b2). The differentiation of the ARO−/− ovaries was further altered as shown by the low staining observed with DDX4, STRA8, SPO11 and FOXL2 probes and by the qPCR low expression of these genes (figure 9 C-F). The ratios of the STRA8/DDX4 and SPO11/DDX4 mRNA levels were lower in ARO−/− ovaries, suggesting that germ cells expressing STRA8 and SPO11 gene expression, and thus undergoing meiosis, were less abundant in ARO−/− ovaries.
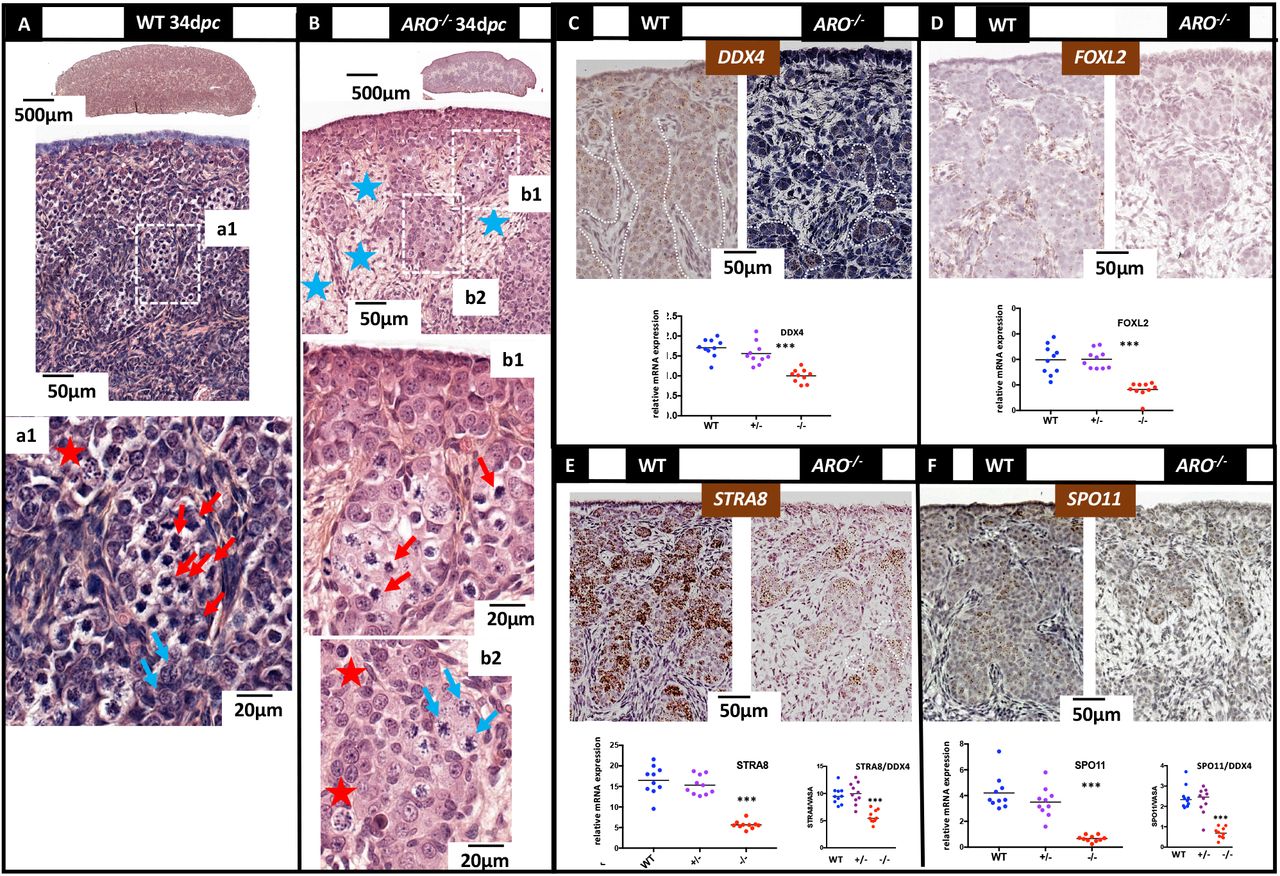
Ovaries collected at 34 dpc (3-4 days after birth) were fixed in Bouin’s then HES colored (A-B) or PAF then ISH treated (C-F). The stages of meiosis were identified as previously published (Peters et al. 1965). (A): In wild type ovaries, large ovigerous cords were filled with numerous germ cells in zygotene (a1, red arrows). Few pachytene stages were observed at the inner part of the cords (a1, blue arrows). (B): In ARO−/− ovaries, the rare ovigerous cords were filled with a few numbers of germ cells in zygotene (b1). As in wild type ovaries, some germ cells in pachytene were found at the inner part of the cords (b2). In both ovaries, aberrant pictures of cell nuclei were characterized by punctiform or condensed and dark colored chromatin (red stars). Blue stars show the connective tissue already observed mostly in ARO−/− ovaries. (C-F): ISH was performed using DDX4 (C), FOXL2 (D), STRA8 (E) and SPO11 (F) genes specific probes. In wild type ovaries, numerous germ cells positive for DDX4, STRA8 and SPO11 filled the ovigerous cords; pre-granulosa cells FOXL2 positive were found around and inside the cords. In ARO−/− ovaries, germ cells positive for these same ISH probes were also found but much less numerous; the FOXL2 labelling was very faint. The diagrams show the results of a quantitative analysis of gene expression. Values are relative expressions measured using total RNA extracted from the two gonads of each animal from wild type (WT), ARO+/− (+/−) and ARO−/− (−/−) rabbits; the same RNA samples were analyzed in the four diagrams. Horizontal bars represent the medians. The STRA8/DDX4 and SPO11/DDX4 diagrams represent the ratio between the expression of the genes for each sample. Stars represent the statistical significance (p<0.01).
These differences intensified further (supplementary Figure 9). As meiosis progressed (10 days post partum, dpp), zygotene, pachytene and diplotene stages were simultaneously observed in wild type ovaries. At the interface between the cortex and the medulla, the large ovigerous cords broke down, forming numerous early primordial follicles encompassing diplotene stage germ cells surrounded by a ring of flat pre-granulosa cells. This was far to be observed in ARO−/− ovaries. Indeed, at 7 days after birth, almost no germ cells in zygotene or pachytene stages were detected; a small number of diplotene-like stage cells were nonetheless detected, some surrounded by a discontinuous ring of somatic cells that could be assumed to be pre-granulosa cells.
Later on at 16-19 dpp, the gonad size was approximately twice as small in ARO−/− rabbits, harboring a thin cortex with a small number of primordial follicles (Figure 10A). All ovigerous cords broke down both in wild type and ARO−/− ovaries and follicles were isolated. However, in ARO−/− ovaries, the ring of pre-granulosa cells surrounding the oocytes was frequently discontinuous and rare well-developed primordial follicles were detected. The FOXL2 ISH labelling was very weak reflecting a low level of well-differentiated granulosa cells (Figure 10B). Nevertheless, oocytes of primordial follicles were positive for the RSPO2 ISH specific probe in ARO−/− follicles, as in wild type ones (Figure 10C).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Ovaries were collected at 16 or 19 dpp then treated with Bouin’s or PAF fixative and processed for HES staining or ISH labelling respectively. (A): HES staining of 19 dpp sections. Blue stars point the abnormal persistence of connective tissue, void of differentiating oocytes or follicles, at the outer part of the cortex and in the medulla of ARO−/− ovaries. The green arrows point some of the primordial follicles in formation with a discontinuous wall of granulosa cells. Early formed primordial follicles surrounded by a wall of regular shape granulosa cells are found in wild type and ARO−/− ovaries (white arrows). (B) and (C): ISH labelling using FOXL2 and RSPO2 gene specific probes. The FOXL2 and RSPO2 probes labelled granulosa cells and oocytes respectively in both wild type and ARO−/− ovaries. Note the huge decrease of FOXL2 positive cells in ARO−/− ovaries compared to the WT control ones.
To conclude, in the ARO−/− ovaries, a small number of oocytes had differentiated after birth; these oocytes were surrounded by a ring of granulosa cells. Despite the probably abnormal differentiation of the latter (shown by the weak expression of the FOXL2 gene), a small population of primordial follicles could differentiate. At the same time, the ovary was invaded by the connective tissue filled with fibers rich in collagen as previously observed from 20 dpc onwards; this latter was not replaced by the ovigerous nests as normally occurs in non-mutant ovaries. The ARO−/− ovaries may have failed to fill-in this connective tissue due to the disturbed formation of ovigerous nests.
Discussion
The already long-standing demonstration of a peak of CYP19A1 gene expression, AROMATASE activity and estradiol production by the fetal ovary in several mammal species between the stage of sex determination and the initiation of meiosis has raised questions on the role of locally produced estrogens on the fetal differentiation of this organ. The mouse model was for a long time the sole available model to study the function of genes in vivo. So, strains of Cyp19a1 gene or estrogen-receptor genes KO mice have been created and a series of papers have reported their phenotype (Fisher et al. 1998; Couse et al. 1999; Britt et al. 2000; Britt et al. 2001; Findlay et al. 2001; Hamilton et al. 2014). All XX mice developed ovaries, but follicle differentiation was abnormal after birth. However, this model could not answer to our question since the mouse is one species where Cyp19a1 gene expression is detected after meiosis initiation in the “late” fetal ovary (Dutta et al. 2014) and not before (Greco and Payne 1994). Thanks to the recent possibility to target genome modifications in numerous other species, we created a rabbit model harboring a mutation in the CYP19A1 gene (ARO−/−), a species with a peak of CYP19A1 gene expression in fetal ovary before meiosis. The ARO−/− rabbits were totally void of any estrogen production. Particularly, the amount of estradiol in gonads of all ARO−/− fetuses was undetectable, which eliminates the possibility of any compensatory mechanism through estradiol from maternal or placental origins. More, no testosterone was detected in the ARO−/− fetal gonads, thus eliminating any possibility of interference due to the local accumulation of androgens in response to the lack of androgen aromatization. The key results of this study regarding XX ARO−/− ovary phenotype were: 1) no XX sex reversal in ARO−/− rabbits, 2) an abnormal differentiation of FOXL2 positive cells, leading to the poor differentiation of ovigerous nests and the reduction of the number of meiotic germ cells and 3) a quasi-absence of follicular reserve, also linked to early folliculogenesis failure.
AROMATASE is not an ovarian determining enzyme in mammals
The question of the role of estrogens in sex determination in mammals was prompted by the finding that in goats, the FOXL2 gene was a main actor for ovarian determination, since the FOXL2 gene loss of function caused an ovary-to-testis sex reversal of the fetal gonad (Boulanger et al. 2014). Previously, FOXL2 was shown to be a crucial transcriptional activator of the CYP19A1 gene in goats (Pannetier et al. 2006) and fishes (Govoroun et al. 2004; Pannetier et al. 2006; Guiguen et al. 2010), and possibly in chicken (Govoroun et al. 2004). Thus, the question was raised to know whether FOXL2 could act on sex determination through CYP19A1 gene transcriptional activation. This question was all the more important that in non-mammalian vertebrate species, anti-aromatase drugs induced female to male sex-reversal (see (Vaillant et al. 2001) in the chicken and (Yin et al. 2017) in fish). The rabbit was possibly an interesting model since, as in goats, the surge of expression of the FOXL2 gene in the fetal ovary occurs when starts the peak of CYP19A1 gene expression at the gonadal sex determination (Diaz-Hernandez et al. 2008; Daniel-Carlier et al. 2013). The results of the present study demonstrate that estrogens play no role on the female/male determinism of the rabbit gonad. A similar finding had already been described in humans, where several cases of total CYP19A1 gene inactivation were reported (Akcurin et al. 2016; Zhu et al. 2016; Mazen et al. 2017; Unal et al. 2018; Praveen et al. 2020). In spite of numerous symptoms affecting the development of gonads, of external genitalia and of breast in the girl, with multiple disorders in energy and bone metabolisms, there was no sex reversal of the human ovary as regard to the genetic sex (Zhu et al. 2016). In contrast, opposite data were reported in the mouse species, describing an increase in the serum testosterone levels in Cyp19a1−/− females (Fisher et al. 1998) and the presence of seminiferous-like structures in the gonads of adult Esr1/Esr2 double knock out females (Couse et al. 1999; Amano et al. 2017). However, this has been controversial and not found by all authors.
Estrogens are involved in early ovarian development
The fetal ovarian differentiation is known to be intimately linked to mechanisms that involve cellular interactions and cell migration, depending on autocrine and paracrine signaling pathways. These processes involve genes coding for the components of the extracellular matrix and cell-cell adhesion molecules. Estrogens have been reported as potent actors in these mechanisms (Juengel et al. 2002; Gentilini et al. 2007; Wilhelm et al. 2007; Zalewski et al. 2012; Hummitzsch et al. 2013). The reduced size of the gonads together with the reduced number of somatic and germ cells is one of the major characteristics of the ARO−/− rabbit ovaries. It was detected shortly after gonadal sex determination (18 dpc), and still clearly visible around birth with a dramatic decrease of ovarian nests. We have also shown that the estrogen receptor α gene (ESR1) was strongly expressed by the ovarian cortical cells specifically by cells located within the coelomic epithelium (Figure 5B). This is worth to be pointed out because it resembles to what has been described in chicken. Indeed, in this species the female to male sex reversal induced by the AROMATASE inhibitor fadrozole induced a reduction of FOXL2 expression (Hudson et al. 2005). More, the experimental down-regulation of the epithelial Esrα gene was sufficient to severely affect the ovarian cortex differentiation (Guioli et al. 2020), thus suggesting that the natural absence of Esrα expression in the cortex of the right ovary of the hen was at least partially responsible for the asymmetric degeneration of the right ovary (Guioli et al. 2014). We can therefore hypothesize that in rabbit ARO−/− ovaries, the absence of estrogens can lead to a dramatic decrease of proliferative activity in the coelomic epithelium, evidenced by the KI67 staining (Figure 7A) and by the decrease of FOXL2-expressing somatic cells from 20-22 dpc (Figure 6).
Anyway, without estrogens, the proliferation of ovarian cells seemed to be affected but not abolished since some nests were clearly visible in ARO−/− gonads at birth although under-developed. So, given that estrogens are involved in ovigerous nests formation, other factors are likely to play a role in this process as well. It has been shown in mice that the Rspo1/Wnt4/β-catenin pathway is involved in cell proliferation of the coelomic epithelium of both sexes during the gonadal switch (Chassot et al. 2012). This pathway could thus be responsible at least in part of the formation of the few ovigerous nests in ARO−/− rabbit ovaries. Transcriptomics (even at the single cell level) and epigenetics studies are underway on WT rabbit ovaries in our team to search molecular targets of estrogens in early ovarian development by comparison with ARO−/− gonads.
The number of germ cells was affected by the lack of estrogens
The reduced number of germ cells is one of the major characteristics of the ARO−/− rabbit ovaries. One of the first sign was the low expression level of the pluripotency marker gene OCT4 and its reduced ISH labelling (see Figures 5 and 6). In mammals, germ cells colonize early the genital crests (Juengel et al. 2002; Wilhelm et al. 2007; Hummitzsch et al. 2013). In the rabbit species, it starts at 9 dpc and is over at around 18 dpc (Chretien 1966). Since the CYP19A1 gene expression starts at 18 dpc, it is likely that the germ cell colonization occurs independently of CYP19A1 expression. Thus, in the ARO−/− ovary, in spite of the absence of estradiol, the initial stock of germ cells was probably not altered. In mammalian ovaries, once germ cells enter the genital ridges, they multiply intensely by mitosis until meiosis. Then, the number of germ cells does not increase any more. Numerous articles have shown that in mammals estrogens stimulate the mitotic activity in the fetal ovary (see review in (Auersperg et al. 2001; Chou and Chen 2018)). During fetal life in rabbits, the mitotic activity of germ cells, which is maximum around 16-18 dpc, remains high up to 26 dpc (Chretien 1966), when the CYP19A1 and ESR1 genes are strongly expressed (Supplementary Figure 7). It can be suggested that the estradiol produced by AROMATASE in the fetal ovary could regulate the mitotic activity of germ cells. This hypothesis is supported by the fact that the germ cells, labelled at this stage by the OCT4 HIS probe, were co-localized in the surface epithelium with cells expressing the ESR1 gene. Germ cell death has previously been reported to occur continuously in the ovary from early fetal stages until puberty and later on (Hussein 2005; Myers et al. 2014). In ARO−/− ovaries, the number of γH2AX positive germ cells (highlighting DNA fragmentation, and at this stage dying germ cells) was similar to that in wild type ones (Figure 7). Thus, our data suggest that this is probably not germ cell death but rather the lack of germ cell divisions that was responsible for the low number of germ cells in the early ARO−/− ovary before meiosis.
Germ cells were committed in meiosis in spite of the lack of estrogens
The commitment of germ cells in meiosis marks the first step of their differentiation as oocytes. In the rabbit species, it occurs at the third part of pregnancy (at around 28 dpc), and is characterized by the surge of the STRA8 gene expression in germ cells specifically. Interestingly, in the ARO−/− ovaries, numerous germ cells were committed to meiosis as shown by the significant surge of STRA8 gene expression; more, the various steps of the meiotic prophase were distinguishable, and the meiosis specific maker gene SPO11 was significantly expressed as well. Thus, it suggests that nor the estradiol neither the AROMATASE enzyme were necessary for meiotic induction and progression. As discussed previously, the number of germ cells possible to be committed in meiosis was reduced in ARO−/− ovaries. More, it should be pointed out that the ratio of gene expression STRA8/DDX4 and SPO11/DDX4 was low in ARO−/− ovaries, showing that an abnormal loss of germ cells occurred at commitment or during meiosis. Thus, although estrogens are not mandatory for meiosis commitment, the fact remains that they are important for limiting meiotic germ cell losses.
Estrogens deprivation impacts follicle formation
Another key feature of the phenotype of the ARO−/− females concerns the abnormal genital tract and ovarian follicle differentiation. At puberty, the genital tract of the XX ARO−/− rabbit was profoundly altered, with massive atrophy, as already reported in Cyp19a1−/− mice (Fisher et al. 1998; Britt et al. 2000), and in agreement with the fact already demonstrated that the development of the female genital tract is highly dependent on estrogens. The follicle differentiation that starts 12-15 days after birth in rabbit at the arrest of meiosis prophase is a continuous process that involves the surrounding of oocytes with flat pre-granulosa cells and the breakdown of ovigerous nests thus giving rise to primordial follicles. Then, the differentiation of pre-granulosa cells from flat- to cuboidal-form cells signs the differentiation of primary follicles. Of course, the size of the population of primordial follicles is linked to the number of oocytes, the latter being the consequence of all the events which occurred during fetal and neonatal life as discussed previously. In the ARO−/− rabbit ovary, this is of particular importance since the pool of oocytes and that of FOXL2 expressing pre-granulosa cells are extremely reduced. More, it has been reported that the nests breakdown is an event specifically sensitive to apoptotic losses (Myers et al. 2014). This could explain why the pool of primordial follicles was very low in ARO−/− rabbit ovary few days after birth (Figure 10) and almost null at puberty (Figure 2). The poor expression of the FOXL2 marker gene by the granulosa cells 16 days after birth signs the abnormal differentiation of the primary follicles, the number of which was extremely low (Figure 2). Surprisingly, controversial results were reported in Cyp19a1 KO mice, where the number of primary follicles was significantly higher than in wild type mice (Britt et al. 2000). To date, we have no explanation for such differences. Furthermore, we observed that despite the absence of estradiol, large antral follicles with oocytes developed in the ARO−/− ovary, however these follicles were very few. This proves that estrogens are not the only actors in follicular growth and differentiation at puberty and onwards. Many previous studies have shown that these events depend on the interactions between several signaling pathways, among which are estrogens but also the anti-Müllerian hormone (AMH) and gonadotropins (Grynberg et al. 2012; Dewailly et al. 2016; Chou and Chen 2018). Additional studies are underway to further analyze the mechanisms of differentiation of these follicles in the absence of estradiol.
To conclude, the present work shows that in the rabbit species, the CYP19A1 gene mutation and the absence of estrogens from early embryo life was at the source of a series of main disorders of the ovarian differentiation process. It did not disturb the sex determination of the early fetal gonad but impacted profoundly the number of germ cells and the early differentiation of somatic cells. It then had repercussions on the subsequent follicular growth and differentiation and finally led to female infertility. This paper brings major and original data on the role of estrogens in species where the CYP19A1 gene is expressed in the fetal ovary before meiosis. Deciphering the gene pathways sensitive to estrogens is currently underway in our team in order to better understand specifically the role of endocrine disruptors such as xeno-estrogens on female fertility.
Materials and methods
Animals
New Zealand rabbits (NZ1777, Hypharm, Rousssay, France) were bred at the SAAJ rabbit facility (Jouy-en-Josas, France). All experiments were performed with the approval of the French Ministry MENESR (accreditation number APAFIS#6775-2016091911407980 vI) following the recommendation given by the local committee for ethic in animal experimentation (COMETHEA, Jouy-en-Josas). All researchers working directly with the animals possessed an animal experimentation license delivered by the French veterinary services. Hormonal treatments for superovulation and surgery for embryo transfer were performed as previously described (Peyny et al. 2020).
Design of TALEN sequences and plasmid constructions
Exon II of rabbit AROMATASE gene was targeted to create InDel mutations near the site of initiation of translation (ATG site). The target sequences (left arm: 5’-TGCTTCATCTGAAGCCA-3’ (sense); right arm: 5’-TGGGTTCAGTATTTCCA-3’ (antisense); 16 bases spaced) were chosen with the ZiFiT Targeter sofware (http://zifit.partners.org). No homology was identified at any other location in the rabbit genome that could represent a potential off-target site (supplementary table 1). The TALEN were constructed as described (Sander et al. 2011), see in supplementary methods). Each TALEN RNA was diluted (100 ng/μl) in injection buffer (Millipore, France) and stored at −80°C until used.
Generation of mutant rabbits
Embryos produced from superovulated females were injected at single-cell stage with an equimolar mixture of the left and right arm TALEN mRNAs (50ng/μl each). Injected embryos were implanted 3-4 hours after injection into the oviducts of anesthetized recipient rabbits via laparotomy. For all details concerning handling of females and embryos, see in (Peyny et al. 2020).
Offspring were screened for the presence of InDel mutations using genomic DNA extracted from ear clips (Jolivet et al. 2014). First detection of founders and characterization of mutation were performed using one set of primers located far upstream and downstream the targeted ATG (F0/R0, Figure 1 and supplementary table 2 for all primer sequences). The amplified fragments were sequenced (Eurofins Genomics, Courtaboeuf, France) and the extent of the mutation was deduced by comparing with the sequence of a wild type rabbit.
For further routine screening of mutants, a quantitative PCR (Fast SYBR Green Master Mix, Applied Biosystems, ThermoFisher, France) was performed using two sets of primers, the CYP19A1 specific one (CYP19A1 gene specific primers) flanking the position of the characterized InDel mutations (F1/R1, supplementary table 2) and another set to amplify a 2 copies reference gene (the rabbit beta GLOBIN gene, ENSOCUG00000000568). By using the Δ(ΔCt) method with a wild type rabbit DNA as reference DNA, we deduced the copy number of the CYP19A1 allele in each DNA sample (no amplification = the two alleles are mutant (ARO−/− genotype); one copy = one mutant allele (ARO−/+ genotype); two copies = wild type (WT or ARO+/+ genotype).s
The presence/absence of the Y chromosome was deduced from the amplification of the SRY gene through PCR analyses with the rabbit beta GLOBIN gene as positive control of the PCR and SRY specific primers (supplementary table 2). All rabbits were tested to determine the genomic sex in parallel to the external and histological observations. In the present paper, mentions of the XY or XX genotype always refer to the PCR determination.
Histological and immunohistological analyses
Immediately after sampling, gonads were immersed in Bouin’s fixative or paraformaldehyde (4% PAF in PBS 1x), fixed for 24 to 72 hours then paraffin embedded. Microtome sections of 5μm thickness were processed. Hematoxylin-eosin-saffron (HES) colorations were performed by the @Bridge platform (INRAE, Jouy-en-Josas) using an automatic Varistain Slide Stainer (Thermo Fisher Scientific). Sirius Red-Fast Green colorations were performed manually.
In situ hybridization (ISH) was performed using the RNAscope ISH methodology (ACD, Bio-Techne SAS, Rennes, France). Briefly, probes at around 1000nt long were designed and produced by the manufacturer in order to match to the full-length cDNA of interest, taking care to reduce any cross-hybridization with non-specific targets. The list of all synthesized probes is given in supplementary table 3. Hybridization was performed on 5μm sections from PAF fixed tissue using labelling kits (RNAscope 2.5HD assay-brown or -red, or RNAscope 2.5HD duplex chromogenic assay blue (conjugated to horse radish peroxidase) and red (conjugated to alkaline phosphatase)) as recommended by the manufacturer. The red labelling (Fast Red) was observed as visible or fluorescent signals. Hybridization was considered as positive when at least one dot was observed in one cell.
Immunohistological analyses were performed on 5μm sections from PAF fixed tissue. The supplementary table 4 recapitulates the antibodies used.
All colored sections (visible, or fluorescent ones) were scanned using a 3DHISTECH panoramic scanner at the @Bridge platform (INRAE, Jouy-en-Josas).
RNA extraction and RT-qPCR analyses
Gonads from rabbit fetuses were collected and immediately frozen at −80°C. Total RNA from each gonad was extracted using the RNeasy® MicroKit (Qiagen, France). Quantitative PCR was performed on reverse transcribed RNAs (High Capacity Reverse cDNA Transcription kit with the included set of random primers, Applied Biosystems, ThermoFisher, France). Several PCR sets of primers and analyses were those published previously (Daniel-Carlier et al. 2013). Other sets of primers are given in the supplementary table 2. Data from significant and specific amplifications only were reported, thus excluding all amplifications with too high or variable Ct values, and non-specific or multiple amplifications.
Measurement of estradiol, testosterone and anti Mülllerian hormone levels in serum samples and fetal gonads
Estradiol and testosterone were assayed by GC/MS according to the protocol described by Giton et al. (Giton et al. 2015) with modifications (Devillers et al. 2019). Female rabbit serum from ARO−/− genotype was used as the matrix for calibrators and quality control (QC) standards after twice charcoal dextran treatments. Sample extraction and purification, derivatization and determination of estradiol and testosterone levels in serum samples and fetal gonads are described in details as supplementary information (see also supplementary Table 5) or can be provided upon request. Anti Müllerian hormone levels were determined in 50 μl aliquots of serum samples by using an ELISA kit (AMH GenII ELISA, with AMH Gen II calibrators and controls, Beckman Coulter, Villepinte, France) as previously described (Bourdon et al. 2018).
Statistics
The statistical analyses were performed using the GraphPad Prism 7 Software (GraphPad Software Inc., La Jolla, CA, USA). Because of the small number of samples in groups, comparisons between values were made by the Mann-Whitney test for non-parametric values. A probability lower than 0.05 was required for significance.
Acknowledgements
The authors would like to thank the staff of the facility (SAAJ, INRAE, Jouy-en-Josas) for the care of the rabbits and Julie Rivière and Marthe Vilotte (UMR GABI, INRAE, Jouy-en-Josas) for their assistance on the histological plateau (@Bridge platform) and for the access to the virtual slide scanner (MIMA2 platform). Special thanks go to Danielle Monniaux (UMR PRC, INRAE, Nouzilly) for the time she spent in enriching discussions and highly valuable comments. This study was supported by ANR grants (GENIDOV:ANR-09-GENM-009; ARGONADS: ANR-13-BSV2-0017; ARDIGERM: ANR-2020-CE14). The authors declare that they have no competing interests.
References




