

Abstract
Human Cytomegalovirus (HCMV) can infect a variety of cell types by using virions of varying glycoprotein compositions. It is still unclear how this diversity is generated, but spatio-temporally separated envelopment and egress pathways might play a role. So far, one egress pathway has been described in which HCMV particles are individually enveloped into small vesicles and are subsequently exocytosed continuously. However, some studies have also found enveloped virus particles inside multivesicular structures but could not link them to productive egress or degradation pathways.
We used a novel 3D-CLEM workflow allowing us to investigate these structures in HCMV morphogenesis and egress at high spatio-temporal resolution. We found that multiple envelopment events occurred at individual vesicles leading to multiviral bodies (MViBs), which subsequently traversed the cytoplasm to release virions as intermittent bulk pulses at the plasma membrane to form extracellular virus accumulations (EVAs). Our data support the existence of a novel bona fide HCMV egress pathway, which opens the gate to evaluate divergent egress pathways in generating virion diversity.
Introduction
Human Cytomegalovirus (HCMV) is a ubiquitous betaherpesvirus of high clinical importance that establishes lifelong latent infection in humans. It is the leading cause of congenital disabilities in the developed world and a significant cause of disease in immunocompromised patients, such as transplant recipients, AIDS, or cancer patients (reviewed in [1]). HCMV has been ranked highest priority for vaccine development by the Institute of Medicine for over 20 years [2]. Despite continuing efforts, no approved vaccine exists so far, and antiviral therapy is currently the only treatment option, with the development of viral resistance being a significant inherent concern [3]. As HCMV causes disease affecting various tissue types and organs, understanding how HCMV can infect different cell types is essential for developing novel antiviral strategies.
HCMV infected cells release distinct virus populations that differ in their glycoprotein content [4, 5] to target specific cell types. While the pentameric complex consisting of gH/gL/UL128/pUL130/pUL131 guides tropism for endothelial and epithelial cells, primarily through cell-to-cell spread, the trimeric complex consisting of gH/gL/gO is needed to mediate infectivity of cell-free virions. Factors that mediate the abundance of these complexes on virions have been recently identified [5–9], but it is unclear how distinct glycoprotein concentrations on individual virus particles are achieved. Spatio-temporally separated envelopment or egress pathways could explain how distinct virus populations are generated, but little is known about these aspects during HCMV assembly.
The virions of herpesviruses assemble in the host cytoplasm using a culminating step called secondary envelopment. During secondary envelopment, viral capsids bud into host-derived membranes, resulting in enveloped viral particles inside transport vesicles (reviewed in [10, 11]. These transport vesicles subsequently release mature virions by fusing with the plasma membrane. Compared to the morphogenesis of the alphaherpesviruses Herpes simplex virus 1 (HSV-1) and Pseudorabies virus (PRV), HCMV morphogenesis is much more involved, taking not only hours but several days. During this time, the virus extensively remodels the host cell’s secretory apparatus leading to the formation of the assembly compartment (AC) [12]. The AC is a dense, donut-shaped, perinuclear structure consisting of convoluted and interconnected membranes centered around the microtubule-organizing center [13]. It contains many cellular proteins traditionally used as organelle-specific markers, including proteins originally associated with Golgi, trans-Golgi, endosomes, and lysosomes [14–17]. However, the extensive viral transformation of the cell’s secretory pathways during AC formation and the short-circuiting of established cellular pathways through viral factors renders the origin of membranes and their identity less clear. Consistently, proteomics analyses indicate that the virus-induced reorganization of the secretory apparatus during AC formation leads to the mixing of membranes from different provenance as targets for secondary envelopment [18].
Currently, secondary envelopment events have only been shown to occur as individual events at small vesicles in the center of the AC [19, 20]. This is consistent with data from alphaherpesviruses where individual virions are continuously released through the fusion of diffraction-limited, virion-containing exocytic vesicles with the plasma membrane [21, 22]. In the case of HCMV, however, studies also found enveloped particles in large multivesicular structures of unknown origin. These large multivesicular structures, containing HCMV progeny, have been called endosomal cisternae or multivesicular bodies (MVBs) in the literature, even though conclusive evidence regarding their biogenesis has been lacking [14,23–25]. A recent study from the Wilson and Goodrum labs suggested that virus-containing MVBs in HCMV fibroblasts and endothelial cells are derived from membranes of different cellular origins [26].
Currently, it is unclear if virus particles in multivesicular structures represent a productive egress pathway or are instead targeted for degradation since secondary envelopment at them or subsequent release of virus progeny by exocytosis could not yet be demonstrated. Interestingly, it has been shown that the deletion of the viral protein UL135 leads to an abrogation of virus-filled MVB-like structures, and mutating UL71, a viral protein likely being involved in membrane scission [27], led to an enlargement of these MVBs [24], possibly indicating their productive role. In addition, a number of other publications implicate MVBs in HCMV morphogenesis [24,25,28–30], and data from the related human herpesvirus 6A (HHV-6A) suggest MVB-like structures as targets for egress [31].
To provide an unbiased view on HCMV envelopment and identify potential alternative HCMV egress routes, we here employed an integrative approach based on volumetric live-cell imaging and three-dimensional correlative light and electron microscopy (3D-CLEM). It provided an unprecedented, spatio-temporally highly resolved view into whole HCMV infected fibroblast cells. We found large transient accumulations of enveloped virions in MVB-like structures that were positive for CD63 in line with previous reports. We dubbed these structures multi-viral bodies (MViBs) as it is unclear at this point if their biogenesis parallels bona fide MVBs. Importantly, we identified secondary envelopment events at MViBs, strongly suggesting that they are targets for the viral envelopment machinery. Live-cell lattice light-sheet microscopy (LLSM) showed that MViBs were transported to the plasma membrane, where they relaxed, and live-cell confocal microscopy illustrated that these events lead to intermittent pulses of virus bulk release. Moreover, a pH-sensitive biosensor functionally confirmed that the observed pulses were indeed due to membrane fusion events. This intermittent virus bulk release led to the formation of extracellular viral accumulations (EVAs) at the plasma membrane. Our data argue for a model in which a large fraction of HCMV capsids envelope at MViBs, which subsequently are transported to the plasma membrane where they fuse intermittently and release bulk pulses of viral particles. We propose that this pathway likely constitutes a so-far neglected HCMV egress route. Future work is needed to dissect its role in generating virion diversity.
Results
HCMV-infected fibroblasts accumulate viral material at specific extracellular sites
To get an overview of HCMV envelopment and egress routes, we initially used live-cell spinning-disk fluorescence microscopy and followed the fate of capsids and viral membranes with an HCMV mutant expressing EGFP-labeled capsid-associated tegument protein pp150 and mCherry-labeled viral glycoprotein gM (HCMV-TB40-pp150-EGFP-gM-mCherry) [32] by single-particle tracking. Despite considerable effort and computational filtering of thousands of analyzed capsid tracks, we could not identify more than a few instances in which diffraction-limited capsid and membrane signals merged and were subsequently co-transported. While we assumed that this was due to the high signal background in the viral AC, it made us look for alternative HCMV egress and envelopment routes that we missed by focusing on trackable small individual events.
Surprisingly, we found that between 72 and 96 hpi pp150-EGFP and gM-mCherry positive virus material accumulated at defined sites in the extracellular space (Fig. 1).

1A Overview indicates the subfigures’ positions in relation to the whole cell. 1B Quantification of EVA occurrence. HFF cells were infected with HCMV-pp150-SNAP-gM-mScarlet- I or HCMV-TB40-WT at an MOI of 1 and fixed at 120 hpi. HCMV-TB40-WT infected cells were stained for gB. Late-infected cells were counted, and the rate of EVAs was quantified. Borders show the 95% confidence interval of the mean. N=269 from 11 replicates for HCMV-pp150-SNAP- gM-mScarlet-I and N=750 from 8 replicates for HCMV-TB40-WT. No significant difference could be found. 1C Spinning-disk confocal section of HFF-wt cells infected with HCMV-pp150-EGFP-gM- mCherry (MOI = 3) at 4 dpi, showing EVAs positive for pp150-EGFP (green) and gM-mCherry (magenta) close to the plasma membrane. Scale bar represents 10 μm. 1D/E CLEM of the area marked in 1C. 1D Rendering of SBF-SEM data depicting the area close and below the plasma membrane. Scale bar represents 3 μm. 1E Correlative overlay of SBF-SEM data from 1D with the corresponding fluorescence data from 1C indicating that pp150-SNAP and gM-mScarlet-I positive EVAs are located outside the cell. Scale bar represents 3 μm. 1F Two z-slices are depicting invaginations (white arrowheads) next to an EVA that can be found in SBF-SEM data along the cell surface. Scale bars represent 700 nm.
These sites were reminiscent of exocytosis hotspots described for HSV-1 [33] (Fig. 1C) and we dubbed them extracellular viral accumulations (EVAs). At 120 hpi, 80-90% of late-infected cells had EVAs at their plasma membrane (Fig. 1B).
To investigate the nature and genesis of EVAs in HCMV-infected human foreskin fibroblasts, we developed a novel three-dimensional correlative light and electron microscopy (3D-CLEM) workflow that combines spinning-disk fluorescence microscopy with serial block-face scanning electron microscopy (SBF-SEM). This approach allowed us to correlate specific labels for capsids (pp150) and viral membranes (gM) with volumetric EM data of whole infected cells at high resolution (Fig. 1D-E).
We identified EVAs below infected cells by fluorescence microscopy (Fig. 1C) and analyzed them by 3D-CLEM (1D-F, Sup. Vid. 1). The EM workflow is based on aldehyde fixation, followed by an adapted reduced-osmium, thiocarbohydrazide, osmium tetroxide (rOTO) regimen, as well as final uranyl acetate and lead aspartate contrasting, which resulted in high contrast volumetric stacks with well-defined membrane and capsid morphology. As depicted in Sup. Fig. 1, HCMV particles in different stages of viral assembly could be clearly identified. DNA-filled virus capsids were easily recognizable as dark contoured, round to hexagonal objects, with their condensed DNA visible as a dark dot or a short line inside depending on orientation. We found that infected cells accumulated virions, dark stained enveloped bodies, which likely represented dense bodies (capsid-free, tegument only containing particles), and a plethora of vesicles of different sizes in pp150 and gM positive EVAs (Fig. 1D-F). We also observed large invaginations at the plasma membrane that might have resulted from either endo- or exocytosis of EVAs (two slices from a volume are shown in Fig. 1F).
HCMV particles bud into and accumulate in MVBs
Next, we sought to investigate the source of EVAs. To our surprise, we found large intracellular bodies positive for both capsid and viral membrane markers at four days post-infection (dpi). A z-projection of a 3D spinning-disk microscopy stack from two adjacent cells is shown in Fig. 2B, and a merge of these light and their respective EM data in Fig. 2C illustrates the correlation of the datasets. Correlation of the large pp150 and gM positive spots identified in the fluorescent light microscopy data with the respective EM volumes confirmed that they represented multivesicular structures filled with significant numbers of virions, dense bodies as well as other structures (Fig. 2D,2F-G, Sup. Fig. 2, Sup. Vid. 2). To distinguish them from bona fide MVBs in the cell-biological sense, we dubbed them multiviral bodies (MViBs). The MViBs in our data resembled structures described in other studies [24, 28], but it has been unclear if they result from a so-far unrecognized egress pathway or a degradation pathway. To elucidate if MViBs could lead to EVAs, we quantified their contents and compared the fractions of virions, dense bodies, and other vesicular material. We found no significant difference between the content of the MViBs and EVAs (Fig. 2E), supporting the hypothesis that EVAs are the result of MViB release. In general, MViBs were very heterogeneous in size and content. Some contained only a few particles, others up to several hundred; see also Sup. Fig. 3 for an overview of a complete AC and Sup. Vid. 3A-B for representative whole-cell datasets. Sup. Vid. 2 depicts a representative MViB that we rendered and where we segmented its contents and color-coded them as done in Fig. 2E. Importantly, we found non-enveloped capsids on the surface of these MViBs (Sup. Vid. 4). Smaller virus-containing vesicles described in previous EM-based studies [19] were often not as prominent in fluorescence microscopy but could also be found in the SBF-SEM data (Sup. Fig. 4). We also regularly found MViBs in cells infected with wild-type HCMV (Fig. 2H-I), confirming that MViBs are not an artifact of the fluorescently-tagged mutants. We concluded that HCMV envelopment can lead to MViBs and that EVAs had very similar contents.

2A Overview indicates the subfigures’ positions in relation to the whole cell. CLEM of HFF-cells infected with HCMV-pp150-EGFP-gM-mCherry (MOI = 3) at 4 dpi. 2B Maximum z-projection of a 3D spinning-disk confocal microscopy stack. pp150-EGFP is colored in green and gM-mCherry signals in magenta. N marks nuclei. Scale bar indicates 10 μm. 2C Correlative overlay of the fluorescence data shown in 2B and corresponding SBF-SEM data. Scale bar indicates 7 μm. The white frame marks MViBs. See also Sup. Vid. 3B. 2D Correlative rendering of MViBs highlighted in 2C. The white frame marks one MViB detailed in 2F 2E Quantitative comparison of MViB and EVA contents. Statistical analysis was performed with a 2-way ANOVA and Šídák’s multiple comparisons test. No significant differences in the contents of MViBs and EVAs could be found. 2F Section from the rendered SBF-SEM stack shown in 2D. Image signals were inverted to facilitate comparison with TEM images. An HCMV capsid budding into an MViB is highlighted (white arrow). Also, refer to Sup. Vid. 3A-B for a 3D rendering of the presented data. Scale bar indicates 1 μm. 2G Insert from 2G. Arrow marks inwards budding viral capsid. Scale bar indicates 0.2 μm. 2H-I HFF cells were infected with HCMV-TB40-WT at an MOI of 5, fixed, and processed for EM as described for SBF-SEM at 120 hpi with the modification that the cells were embedded for classical sectioning in Epon without conductive fillers. Filled arrowheads indicate MViBs filled with virus progeny. All scale bars represent 0.2 µm.

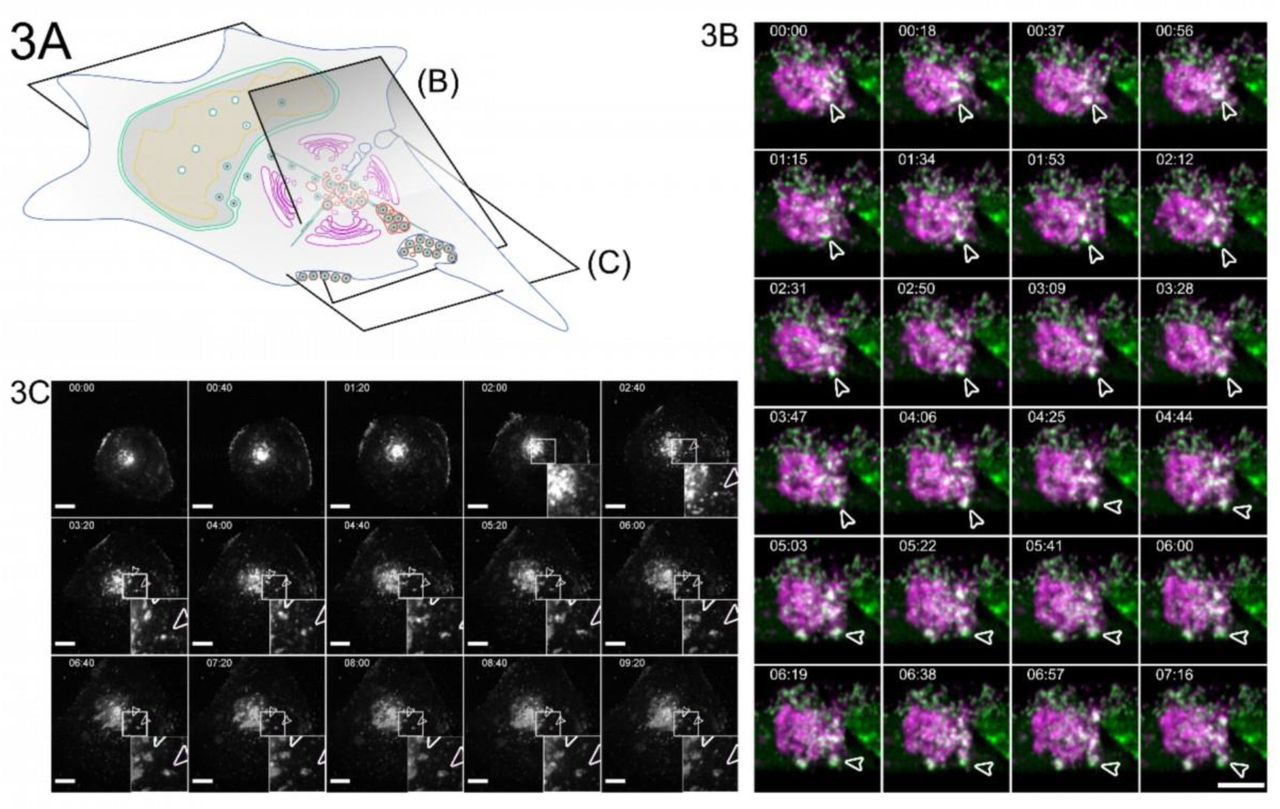
3A Overview indicates the subfigures’ positions in relation to the whole cell. 3B HFF cells were infected with HCMV-pp150-EGFP-gM-mCherry at an MOI of 1. At 96 hpi, the cells were imaged by lattice light-sheet microscopy, taking whole-cell volumes every 2.11 seconds at a 30° angle to the growth substrate. Maximum projections of 20 slices with a total depth of 2 µm of an area under the viral AC and incorporating the plasma membrane are shown. White arrowheads highlight an MViB positive for pp150-EGFP (green) and gM-mCherry (magenta) that approaches the plasma membrane and relaxes at it. Also, refer to Sup. Vid. 5 for a rendering and several side views. 3C HFF cells were infected with HCMV-pp150-SNAP-gM-mScarlet-I at an MOI of 1. At 72 hpi, cells were imaged live with confocal spinning-disk microscopy. Only the gM-mScarlet-I channel is shown. Both channels can be seen in Sup. Vid. 7. The formation of two EVAs is highlighted with white arrowheads. Scale bar indicates 10 μm. The time format is hh:mm. Also, refer to Sup. Vid. 6 and 7.
Pulses of bulk release lead to viral extracellular accumulations at the plasma membrane
To illuminate the fate of MViBs, we used two live-cell fluorescence microscopy modalities. First, we utilized inverted lattice light-sheet microscopy to acquire 3D volumes of infected cells at high temporal resolution for 15-45 minutes for minimal phototoxicity and photobleaching. We found that MViBs traveled from the assembly complex to the plasma membrane, where they seemed to relax, possibly indicating their fusion with the plasma membrane (Fig. 3B, Sup. Vid. 5). In a second approach, we imaged longer timespans in the infection cycle using time-lapse live-cell microscopy with less temporal coverage in 2D. To this end, we used a modified HCMV mutant with more photostable fluorescent tags (HCMV-TB40-pp150-SNAP-gM-mScarlet-I) for imaging z-stacks over several days. We imaged HFF cells between 72 and 96 hpi for 18 to 60 hours every 40 minutes. Strikingly, we observed MViBs coming close to the observation plane at the plasma membrane, where they relaxed into patches of viral material (Fig 3C, Sup. Vid. 6-7). These patches were identical in their phenotype to the EVAs shown in Fig. 1 and were positive for pp150 and gM. The EVAs did not diffuse away but were often left behind when cells moved away, indicating that most of the exocytosed material did not stay cell-associated. EVA formation generally occurred as intermittent pulses as MViBs came into the observation plane near the plasma membrane and relaxed (Sup. Vid. 6-7). The release events varied in their fluorescence intensity, consistent with our observation that MViBs were very heterogeneous in size and content. Based on these observations, we concluded that MViB exocytosis leads to EVA formation.
MViBs release their cargo through fusion with the plasma membrane and result in EVAs
To confirm that the observed bulk release events were indeed induced by fusion of MViBs with the plasma membrane, we used the pH-sensitive fluorescent protein super-ecliptic pHluorin as a biosensor to detect exocytosis events. We created a cell line stably expressing a CD63-pHluorin fusion construct [34] as our data (presented in the next paragraph) indicated that CD63 is enriched on MViBs membranes but not on virions. In this construct, pHluorin is inserted into an extracellular loop of CD63, such that it points towards the luminal side in multivesicular structures and to the extracellular environment after fusion. Accordingly, pHluorin is quenched by the acidic pH inside this luminal space of MVBs, rendering the construct almost non-fluorescent. However, upon fusion with the plasma membrane, pHluorin gets exposed to the pH-neutral extracellular milieu, and fluorescence recovers rapidly. The increase in fluorescence intensity provides an easily detectable and quantifiable indicator of fusion with the plasma membrane. Imaging of fixed, permeabilized cells in which intracellular pHluorin was dequenched confirmed that the fluorescence signal from the CD63-fusion marked gM and gB positive bodies (Sup. Fig. 5A-C, Sup. Vid. 8).
For imaging of potential fusion events, we picked cells that had not yet accumulated EVAs on the outside of the basolateral cell surface and used live-cell total internal reflection microscopy (TIRF) to image fusion events for several hours without phototoxicity. We took images every 1.5-2 seconds for 60 minutes since we predicted that actual membrane fusion and pH equilibration might be very rapid. We found that MViBs came into the TIRF-field and relaxed into EVAs shortly after arrival at the plasma membrane. MViB fusion resulted in EVAs positive for pp150 and gM (Fig. 4B, arrows). Quantification of the gM-mScarlet-I and pp150-SNAP signals showed that as the vesicular bodies arrived at the plasma membrane, their fluorescence intensities increased until they peaked and subsequently fell to stable plateaus of continuously elevated signals (Fig. 4C). Strikingly, these events were accompanied by flashes of green fluorescence between the MViBs arrival and the relaxation event, indicating that the membranes had fused (Fig. 4C). The reduction of green fluorescence indicated that most of the CD63 diffused away from the fusion site. The gM and pp150 signals increased directly before the fusion event and decreased as MViBs relaxed into a flattened patch. The exocytosed material emitted a continuously elevated signal. These results indicated that MViB fusion with the plasma membrane led to EVA formation.

4A Overview indicates the subfigures’ positions in relation to the whole cell. HFF-CD63-pHluorin were infected with HCMV-pp150-SNAP-gM-mScarlet-I at an MOI of 0.6 and imaged at 72 and 96 hpi by fluorescence microscopy under TIRF conditions for 1h at an average frame rate of 0.57 frames per second (fps). 4B TIRF images of a cell before (upper row labeled with before) and after (lower row of images labeled “after”) bulk release events from MViBs occurred. Positions of EVA formation are marked by the white arrows (black in the merge). Scale bar indicates 10 µm. 4C Quantification of fluorescence signals during EVA formation events over time. Solid lines are averages from 14 EVA formation events extracted from 5 different cells chosen from 4 replicates of infections. Grey areas show the standard error of the mean.
MViBs carry markers of the endocytic trafficking system and the exosome pathway
Intermittent bulk release of vesicles is a functional hallmark of exosomal pathways. Therefore, we used immunofluorescence combined with mass spectrometry to approximate possible overlaps between virus composition and exosome generation. To this end, we performed a mass spectrometry analysis of gradient-purified extracellular virions. Gradient-purified virus particles contained markers of Golgi-to-endosome trafficking (syntaxin 12, Rab14, VAMP3), early endosomes (Rab 5C, syntaxin 7), as well as exosomes (HSP70, HSP90, GAPDH, enolase 1, 14-3-3, and PKM2)[35], suggesting that HCMV might use a mix of membranes originating from Golgi- and endosomal membranes for secondary envelopment to generate MViBs (Sup. Table 1, Sup. Fig. 6). Our findings are consistent with a recent study, concluding that HCMV hijacks parts of the exosome pathway for egress [36].
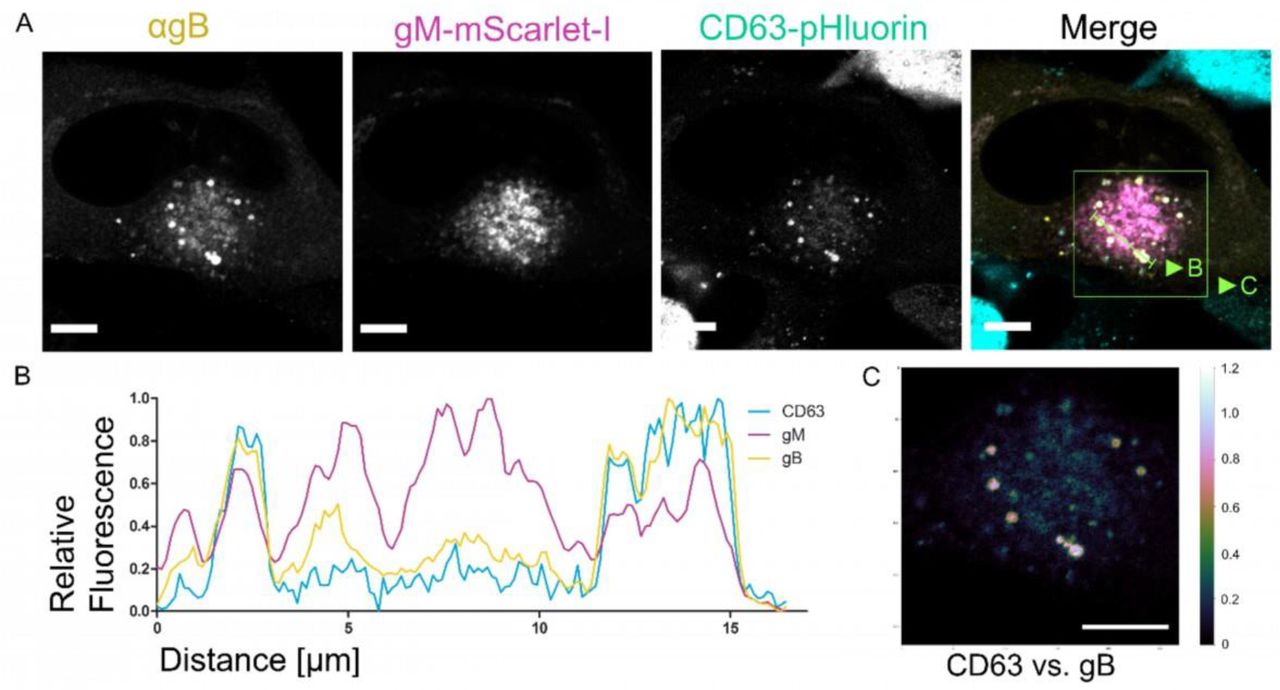
Other classical markers for membranes used in the exosomal pathway are the tetraspanins such as CD9, CD63, and CD81. The role of CD63 in HCMV infection has been investigated before, however, with conflicting results [37, 38]. Using immunofluorescence, we tested if the tetraspanins are localized to MViBs (Fig. 5A-C, Sup. Fig. 7A-B). The density of protein signals in the AC complicated the analysis, yet we could identify CD63 colocalizing with large vesicles containing pp150 and gM (Fig. 5A-C). We also performed EM with immunogold labeling against CD63 to investigate its presence on MViBs at high spatial resolution. Although the content of large bodies was often poorly retained after processing for immunogold staining, we regularly found HCMV particles in large bodies that also were positive for CD63, indicated by the presence of nanogold particles (Sup. Fig 8). However, CD9 and CD81, in our hands, localized to the AC but not specifically to MViBs (Sup. Fig. 7A-B). Besides being present on the MViB limiting membrane, exocytosed material in EVAs did not show any significant CD63 signal, implying that CD63 is unlikely to be incorporated into virions (Fig. 5A-C). This observation is supported by the absence of CD63 in our virion proteomics data (Sup. Tab. 1) and is in line with previous studies [39].

5A HFF cells were infected at an MOI of 1 with HCMV-pp150-SNAP-gM-mScarlet-I, fixed at 4 dpi, stained for CD63, and whole cells were imaged using confocal laser scanning microscopy. From a representative cell, two slices are shown. One slice depicts the middle of the cell (cell middle), and one depicts the plasma membrane level (cell bottom). The fluorescence pattern of CD63 (⍺CD63) was compared to gM (gM-mScarlet-I), and pp150 (pp150-SNAP). In the cell’s center, CD63 localized to the assembly complex’ center and marked MViBs in the cytoplasm, which were pp150 and gM positive. At the plasma membrane, EVAs were positive for pp150 and gM signals but lacked CD63. 5B Spatially weighted colocalization analysis shows areas in the assembly complex where CD63 colocalization with MViBs is especially pronounced (cell middle). No significant colocalization between CD63 and pp150 or gM is present in EVAs (cell bottom). All scale bars (5A and 5B) indicate 10 µm. 5C Line plots for the indicated areas in 5B.
To gain further insight into the biological identity of MViBs, we tested if HCMV bulk release was susceptible to inhibitors of MVB biogenesis or exosome release. Out of an initial panel of ten drugs (Bexin-1, Simvastatin, Climbazole, GW4869, Ketotifen, Manumycin A, Nexinhib20, Suphisoxazole, Tipifarnib, U18666A) that were described to influence MVB or exosome biogenesis, we characterized the effect on HCMV for three of them (Ketotifen, Tipifarnib, U18666A) in more detail. U18666A is an inhibitor of cholesterol trafficking [40, 41], Tipifarnib is a farnesyl transferase inhibitor with high activity against exosome production [42], and Ketotifen is a mast-cell stabilizing agent, currently under investigation for its ability to block exosome release from cancer cells [43, 44]. We tested these drugs at concentration ranges between 0.1x and 2x of reported active concentrations from the literature during virus infection. Only Tipifarnib was able to significantly reduce viral titers at 4 dpi (Sup. Fig. 9C). Tipifarnib was also able to reduce the number of EVAs present at 5 dpi (Sup. Fig. 9A-B, D). Moreover, we found no significant cytotoxicity of Tipifarnib in our HFF cells compared to the vehicle control (Sup. Fig. 9E). While Tipifarnib had no pronounced effect on the expression of the immediate-early genes IE1/2 or the early gene UL44, it had a significant effect on the abundance of the late protein pp150, being a peripheral part of the viral capsid (Sup. Fig. 9F) in total cell lysates. Using Ketotifen, Tipifarnib, and U18666A, we were, therefore, unable to delineate the biogenesis of MViBs.
While our data indicate a novel functional egress pathway for HCMV in which MViBs are targeted for secondary envelopment and subsequently exocytosed, leading to the intermittent bulk release of a large number of viral particles into EVAs, it is unclear at this point how MViBs relate to MVBs. Future studies are needed, importantly also to illuminate the potential role of MViBs in releasing virions of specific cell-tropism.
Discussion
Little data exist on the spatio-temporal organization of HCMV egress at the subcellular level. Previous studies have mostly reported single-virion/single-vesicle envelopment events, which have shaped our current picture of HCMV secondary envelopment [19, 24]. These data are consistent with a study by Hogue et al. [45], which shows that individual alphaherpesvirus virions are released at the plasma membrane. Still, data suggests that virus-filled multivesicular structures can form in HCMV-infected cells as well as HHV-6A and Murine Cytomegalovirus (MCMV) infected cells [14,24,28,31,46] and that Golgi- and endosome-derived membranes are targeted by HCMV [14–17,23,39]. While the previous literature often called these virus-containing multivesicular structures “MVBs”, we decided to dub them multi-viral bodies (MViB) as we could not untangle their descent clearly. A recent study from the Wilson and Goodrum labs suggested that virus-containing structures in HCMV fibroblasts and endothelial cells are derived from membranes of different cellular origins [26]. Importantly, a functional role for these “virus-containing MVBs” or “MViBs” in egress has lacked so far [14,23–25].
Here, we started by investigating EVAs. We were intrigued that most infected cells were positive for EVAs late in infection and investigated their formation. Using a novel 3D-CLEM workflow that combines dynamic information from spinning-disk fluorescence microscopy with high-resolution information from serial block-face scanning electron microscopy, we found that HCMV can form virus particles by budding into MViBs. By time-lapse and functional live cell imaging, we provide evidence that MViBs can fuse with the plasma membrane and intermittently release tens to hundreds of virus particles in bulk, resulting in plasma membrane-associated EVAs. Finally, proteomics of purified virions, functional imaging, and correlation of CD63 localization with MViBs suggested that MViB-mediated HCMV egress might use features of the cellular exosomal pathway; however, drugs inhibiting MVB formation and exosome release showed no or inconclusive effects.
While EVAs represented static endpoints, MViBs were highly dynamic and transient. Integrating imaging technologies that can cover large spatio-temporal ranges of HCMV infection proved to be instrumental in analyzing the role of MViBs in HCMV egress. Our live-cell imaging indicates that MViBs form relatively quickly between 72 and 96 hpi and are rapidly released asynchronously, leading to pulses of EVA formation. This mechanism is in contrast to studies that have been performed in alphaherpesviruses, where single PRV virus particles have been shown to travel to the plasma membrane and be released by fusion [21]. Our data, however, does not exclude the existence of a separate egress pathway, analog to the mechanisms shown for alphaherpesviruses (reviewed in [47]). Compared to previous studies, our new correlative 3D-CLEM workflow provides a major technological advancement permitting us to observe whole cells in a defined infection state without the need for serial sectioning [19]. This has allowed us to analyze transient MViBs, which would have been otherwise hard to catch at high resolution.
From our data, it remains unclear if HCMV uses bona fide cellular MVBs for envelopment and transforms them into MViBs or if they are generated de novo. Cellular MVBs produce similar bulk pulses of extracellular vesicles (EVs) or exosomes by fusion with the plasma membrane [48–50]. EVs form through budding into the lumen of late endosomes. This process generates MVBs characterized by the presence of the late endosomal markers CD63, LAMP1, LAMP2, Rab4, and Rab5 (reviewed in [50]). Budding at MVBs is catalyzed by the endosomal sorting complex required for transport (ESCRT) [50]. While some parts of the ESCRT machinery play a role in the secondary envelopment of alphaherpesviruses [27,51–53], they likely do not play a role in HCMV infection [54–56]. However, it was recently shown that HSV-1 proteins pUL7 and pUL51 form a complex that might constitute a mimic of an ESCRT-III complex. HCMV homologs pUL103 and pUL71 are predicted to be structurally very similar to their HSV-1 counterparts and might likewise perform ESCRT functions for the virus during infection [27]. A recent proteomics study supports this notion by showing that HCMV utilizes parts of the exosome biogenesis machinery independently of classical ESCRT-pathways [36].
Members of the tetraspanin family, such as CD9, CD81, and CD63, have also been described to be enriched on EV membranes [48]. Tetraspanins are known to form microdomains called tetraspanin-enriched microdomains on the cell surface [57] and are active in the organization of the plasma membrane, recycling, and cellular signaling [57, 58]. Tetraspanins are involved in sorting and targeting cargo to MVBs and, in cooperation with the ESCRT machinery, into EVs [59, 60]. While it has been shown that HCMV-infected cells release EVs that contain viral surface proteins such as gB [61], the role of exosomal pathways in HCMV particle envelopment and release are broadly not defined. Although inhibitors of exosome biogenesis can slow HCMV spread, they do not significantly influence viral titers [37, 62], possibly arguing for an involvement of the MVB/exosome-pathways in cell-to-cell spread. Contradictory evidence exists for the role of CD63 in HCMV virus production. While one study did not find a significant effect of siRNA-mediated CD63 knock-down on HCMV titers [37], another recent study found a substantial reduction of HCMV titers upon CD63 siRNA knock-down [38]. The reason for this discrepancy is difficult to determine since the experimental settings in which each of the datasets was acquired varied drastically. This is especially true for the virus strains used in these studies. While Hashimoto et al., as well as Turner et al., used the lab-adapted AD169 strain, Streck et al. investigated the more clinical TB40/E strain [36–38]. AD169 is adapted to release large amounts of supernatant virus from in vitro cultured fibroblast, while strains like TB40/E, which resemble clinical isolates more closely, produce both cell-associated virus and cell-free virus [63, 64]. It is, therefore, tempting to speculate that release pathways that rely on CD63 are used mainly by HCMV to produce cell-free virus, whereas other pathways responsible for cell-associated spread might not be impaired in the absence of CD63.
We found colocalization between the tetraspanin CD63 and the viral envelope glycoproteins gB, gM, and the tegument protein pp150. However, CD81 and CD9, which are also associated with exosomes, did not colocalize with the viral markers as strongly. Since EVAs were negative for CD63, this marker might be excluded during the budding process at the MVB surface. However, this idea contradicts a previously published study showing that CD63 is incorporated in the virion envelope [55]. Importantly, we and others did not find significant enrichment of CD63 in proteomic analyses of purified HCMV virions [36]. CD63 possibly plays a role in the sorting of viral glycoproteins to sites of secondary envelopment, as tetraspanins are known to be involved in sorting plasma membrane-bound molecules into MVBs [59, 60]. HCMV gB is known to localize to the plasma membrane and be sorted through endocytic and recycling pathways by an acidic cluster in its cytoplasmic domain [65, 66]. For HSV-1, it was reported that disrupting the endosome-to-MVB trafficking pathway leads to the mislocalization of HSV-1 gB [67]. More recently, it has also been shown that HSV-1 replication leads to an increase in the exocytosis of CD63-containing extracellular vesicles, leading the authors to hypothesize that HSV-1 modulates exosome biogenesis for its benefit [68]. Taken together, these reports indicate that endocytic pathways can be involved in the trafficking of viral factors to sites of herpesvirus secondary envelopment. Our observation that HCMV gB strongly localized with CD63 might support this hypothesis and fits a recent report that gB is enriched in exosomes [61]. Moreover, a recent proteomics study focusing on exosome release from HCMV infected cells aligns with this interpretation [36]. This study further identified several additional viral proteins that likewise appear in exosomes. The data provided by the authors strengthen the overall idea that HCMV exploits endocytic trafficking and exosome biogenesis pathways for the assembly and egress of virus particles. However, how much of the host factors involved with exosome generation in the absence of virus infection are involved in virus particle production remains unclear. In our hands, the MVB inhibitor U18666A does not influence virus production. In contrast, Tipifarnib, an inhibitor of exosome biogenesis, significantly reduces virus titers 4 dpi and EVA generation. Tipifarnib has been shown to reduce Rab27a, nSMase2, and Alix levels, which might result in an effect on trafficking of viral components to assembly sites or membrane remodeling during secondary envelopment [42]. Alternatively, inhibition of the cellular farnesyltransferase by Tipifarnib might also act on Ras signaling pathways, which have been reported to positively influence HCMV, HSV-1, and other herpesvirus infections [69–71]. An effect of Tipifarnib on transcription would be the simplest explanation for the reduced pp150 levels at 3 and 4 dpi. On the other hand, inhibition of downstream HCMV assembly processes might also result in the degradation of structural proteins such as pp150. Moreover, it is conceivable that the host farnesyltransferase is directly involved in the post-translational modification of virus proteins and that its inhibition by Tipifarnib has a negative effect on viral protein levels and replication.
Instead of being the result of an altered MVB pathway, MViBs might originate from the fusion of individual virus-filled transport vesicles as described for the related betaherpesvirus HHV-6A [31]. This model fits reports that MViBs were mostly found in the AC periphery while most capsid budding into individual vesicles is observed in the center of the AC, where early endosomal markers and Golgi-markers merge [15, 16]. However, in the work we present here, we regularly found budding events at MViBs but could not identify intracellular vesicle fusion events leading to MViB formation in entire 3D-EM volumes of infected cells. We, therefore, conclude that MViB-mediated HCMV egress is a novel spatio-temporally separated egress pathway.
HCMV produces cell-free virus in addition to cell-associated virus in fibroblasts and predominantly cell-associated progeny in endothelial cells [4]. These different particle populations vary in their trimeric to pentameric glycoprotein complex composition, resulting in their different cell tropism. It is plausible to hypothesize that these virus populations might undergo different envelopment processes in the cell and are exocytosed with a different spatio-temporal profile [4]. A recent study from the Wilson and Goodrum labs suggests that virus-containing MVBs in fibroblasts and endothelial cells are derived from different cellular membranes, which would add another potential HCMV egress pathway that could result in different virus populations; however, it is unclear if these pathways are functional in egress [26].
Future work needs to focus on characterizing the particle populations exocytosed by these different pathways regarding their glycoprotein content and define their role in potentially divergent egress routes. We used the HCMV strain TB40, which can produce two virus populations on HFF cells which are endothelial-cell and fibroblast-topic [4]. The EVAs that we found were largely static during live-cell imaging and might represent a cell-associated viral population. We found EVAs not only trapped between the cell and the cell support but also on the upper side of infected cells, as well as between cells. This observation would support the idea of cell-to-cell spread. However, our proteomics data and a recent study [36] found that soluble, purified virions showed markers of the exosome pathway. If the virions released through EVAs are the only ones that carry exosome-markers, this would suggest that it is unlikely that they stay cell-associated and play a role in cell-to-cell spread.
In summary, our data, combined with published studies, suggest a model in which membranes originating from a fusion of both the endosomal and trans-Golgi network are used for either individual envelopment of capsids or to generate MViBs in two spatio-temporally separated processes. MViBs are then transported to the plasma membrane, where fusion results in bulk pulses of virus particle exocytosis and the formation of EVAs (Fig. 6). Future work is needed to delineate the biogenesis of MViBs and, importantly, their potential role in producing specific virus populations.

1-3 Membranes of late-endosomal and trans-Golgi origin are trafficked to the center of the assembly complex and subsequently utilized for secondary envelopment. 4 After egress from the nucleus, capsids are trafficked to the assembly complex where they either bud individually into single vesicles or into MViBs to acquire their membrane envelope. 5-6 Virus-containing vesicles and MVBs are transported towards the plasma membrane, and 7 fuse with it to release their content to the extracellular space. 8 MViB fusion leads to EVA formation.
Materials and Methods
Cells and Viruses
HFF-1 cells (ATCC-SCRC-1041, ATCC) were cultivated in Dulbecco’s Modified Eagles Medium Glutamax (Thermo Fisher Scientific), supplemented with 5% FBS superior (Merck) and 2*105 units/ml Recombinant Human FGF-basic (PeproTech Inc.). HCMV-pp150-EGFP-gM-mCherry was a kind gift by Christian Sinzger [32]. The HCMV-TB-40-BAC4 was a kind gift by Wolfram Brune [72]. Different multiplicities of infection (MOIs) were used for the infection experiments. In general, low MOI infections were used to avoid artifacts generated by high virus doses. Therefore, whenever possible, we used MOIs between 0.5 and 1. However, for particular experiments, such as bulk assays or electron microscopy, we used MOIs of up to 5. The used MOI is indicated for each experiment.
Spinning-disk Fluorescence Microscopy
Spinning-disk microscopy was carried out on a Nikon TI2 (Nikon) based spinning-disk system equipped with a Yokogawa W2, a Nikon 1.49 NA Apo-TIRF objective, and an Andor iXON888 EMCCD (Andor Technology). The resulting pixel size was 130nm, and image acquisition was done with NIS-Elements. Further, the setup was equipped with 405, 488, 561, and 640 laser lines and corresponding filter sets. Life cell experiments were carried out with a humidified incubation chamber heated to 37°C and 5% CO2 controlled by a gas mixer. For fluorescence microscopy, cells were grown in Ibidi 35mm glass-bottom dishes (Ibidi GmbH), for CLEM in Ibidi 35mm grid polymer bottom dishes. SNAP labeling before live-cell imaging with SNAP-Cell 647-SIR (New England Biolabs GmbH) was done according to the manufacturer’s instructions. Image processing and analysis were performed in ImageJ/FIJI.
Serial Block Face Scanning Electron Microscopy (SBF-SEM)
For SBF-SEM, cells were fixed at the indicated time-points with 2% Paraformaldehyde (PFA/ Science Services) and 2.5% Glutaraldehyde (GA/ Science Services GmbH) in Dulbecco’s phosphate-buffered saline (D-PBS, Sigma-Aldrich) for 5 minutes at room temperature (RT) and 55 minutes on ice. Subsequently, the sample was processed with the following procedure: Postfixation with 2% Osmium Tetroxide (OsO4/ Science Services) and 2.5% GA in D-PBS on ice, staining with 2% OsO4, 1.5% potassium ferrocyanide (Sigma-Aldrich), 2mM CaCl2 (Sigma-Aldrich) in water, incubation in 0.5% thiocarbohydrazide (Sigma-Aldrich) in water, staining with 2% OsO4 in water, incubation in 1% gallic acid (Sigma-Aldrich) in water, staining with 2% uranyl acetate (Merck KGaA) overnight in water. On the next day, the sample was stained with freshly prepared Waltons lead aspartate [73] (Pb(NO3)2 (Carl-Roth), L-Aspartate (Carl-Roth), KOH (Merck)), and subsequently subjected to a PLT dehydration series to Ethanol Rotipuran (Carl-Roth). Finally, the samples were infiltrated with 70% Epon in Ethanol before two incubations with 100% Epon and the final polymerization was carried out in Epon supplemented with 3% silver flakes (Sigma-Aldrich) and 3% (w/w) Ketjen Black (TAAB). Sample blocks of 0.5x0.5 mm were cut, mounted, and inserted into a Gatan 3View stage (Gatan) built in a Jeol JSM-7100F scanning electron microscope (Jeol). For imaging, the sample stage was biased with a 500V positive charge to account for sample charging during the scanning process. For the acquisition, 3x3 nm pixel size images were scanned, followed by the repeated ablation of 50 nm sections. The acquisition was controlled by the Gatan Digital Micrograph software, which was also used for stack alignments. Further processing of the datasets was performed in FIJI, and the volumes were rendered in Imaris 8 (Bitplane). To quantify MViB and EVA compositions, subvolumes of those structures were randomly chosen and extracted. Subsequently, particles were manually identified and counted. The image handling tasks were performed in ImageJ/FIJI. Statistical analysis was performed in GraphPad Prism 8.
Transmission Electron Microscopy (TEM)
For TEM, cells were fixed and processed as described for SBF-SEM up to the embedding step. The cells were embedded in Epon without fillers, sectioned to 50 nm on a Leica Ultracut Microtome (Leica), and transferred to copper mesh grids. Electron microscopy was performed on an FEI Tecnai G20 (FEI/ Thermo Fisher Scientific), and images were acquired on an Olympus Veleta side-mounted camera (Olympus).
Lattice Light Sheet Microscopy
Lattice light-sheet microscopy was performed on a Zeiss Lattice Light Sheet 7 (Carl Zeiss) as part of an early adaptor program, controlled with Zeiss Zen Blue software. The device is equipped with 488, 561, and 640 laser lines and multi-bandpass filters. Live-cell experiments were carried out on Ibidi 35mm glass-bottom dishes at 37°C with 5% CO2 in a humidified atmosphere. Images were acquired on a pco.edge (PCO AG) sCMOS camera with a final pixel size of 145nm. Images were deconvolved after acquisition in Zen Blue using the built-in constrained-iterative algorithm. 2D image processing was done in Zen Blue, arrangements and montages were made in FIJI. 3D image processing was done in Arivis 4D (arivis AG); videos were cut and arranged in Adobe Premiere Pro (Adobe Inc).
BAC Mutagenesis
BAC mutagenesis was performed as described before by en-passant Red Recombination [74]. The creation of HCMV-TB40/BAC4-pp150-SNAP-gM-mScarlet-I was done in two steps. At first, UL32 (gene locus of pp150) was mutated by the C-terminal insertion (after K1045) of the SNAP-Tag-SCE-I-KanR shuttle sequence with a nine amino acid linker (HTEDPPVAT) and subsequent second recombination to clear the Kanamycin resistance and restore the SNAP-Tag sequence (NEB; for complete insertion sequence see Table 1). This was followed by the insertion of the mScarlet-I [75] sequence in the UL100 gene between the codons for amino acids V62 and M63 of gM by amplifying the mScarlet-I-SCE-I-KanR shuttle construct with the primers shown in Table 2, with the second recombination as described for the first step. The virus was reconstituted by electroporation of the BAC DNA into HFF cells.
Gateway Cloning and Lentivirus Transduction
Plasmid pCMV-Sport6-CD63pHluorin was a gift from DM Pegtel through Addgene (Addgene plasmid # 130601 ; http://n2t.net/addgene:130901 ; RRID:Addgene_130901, Addgene). For Gateway (Thermo Fisher Scientific) cloning, the pCMV-Sport6-CD63pHluorin was recombined with pDONR-221 (Thermo Fisher Scientific) to produce the pENTR-CD63pHluorin vector that was further recombined with pLenti-CMV-Puro-DEST (w118-1), a gift from Eric Campeau & Paul Kaufman through Addgene (Addgene plasmid # 17452; http://n2t.net/addgene:17452; RRID: Addgene_17452).
The resulting pLenti-CMV-CD63pHluorin-Puro was then transfected with polyethyleneimine (Polysciences) together with 3rd generation Lentivirus vector helper plasmids, gifts by Didier Trono, RRID: Addgene_12253, Addgene_12251, Addgene_12259) into 293XT cells (Takara Holdings). Lentivirus containing supernatant was harvested at 48, 72, and 96 hours post-transfection, filtered through 0.2 µm syringe filters, and used to transduce HFF-1 cells. 72hpi, the HFF-cells were selected with Puromycin (Thermo Fisher Scientific) at 5 µg/ml. Furthermore, the cells were sorted by fluorescence-activation (FACS), using a FACS Aria Fusion (BD Biosciences), for the 10% strongest fluorescent cells, further cultivated and used for the experiments.
Immunofluorescence
For immunofluorescence experiments, cells were grown in 35mm glass-bottom Ibidi dishes and fixed at the indicated time-points with 4%PFA in D-PBS. SNAP labeling with SNAP-Cell 647-SIR was done as described in the manual for SNAP-Cell 647-SIR (NEB). Afterward, the samples were permeabilized with TritonX-100 at 0.1% in D-PBS with subsequent blocking with 3% Bovine Serum Albumin (Sigma-Aldrich) in D-PBS. Primary antibodies used in this study were Ultra-LEAF™ Purified anti-human CD63 H5C6 (Biolegend), Anti-Cytomegalovirus Glycoprotein B antibody [2F12] (ab6499) (Abcam), Purified anti-human CD9 HI9a (Biolegend), Purified anti-human CD81 (TAPA-1) 5A6 (Biolegend). Secondary antibodies used were Alexa 647 goat anti-mouse (Thermo Fisher Scientific) and Alexa 488 goat anti-mouse (Thermo Fisher Scientific).
Quantification of the frequency of extracellular viral assemblies
HFF-WT cells were infected with HCMV-pp150-SNAP-gM-mScarlet-I or HCMV-TB40-WT at an MOI of 1 and fixed at 120hpi. HCMV-TB40-WT infected cells were stained for gB as described for the other immunofluorescence experiments. Late infected cells were identified in WT-infected cells by a well identifiable gB-positive assembly complex. In the HCMV-pp150-SNAP-gM-mScarlet-I infected cells, late infected cells were identified by three conditions: 1) Well identifiable gM-positive assembly complex. 2) Nuclear signal of pp150-SNAP. 3) Significant pp150-SNAP signal in the assembly complex.
Confocal Scanning Imaging
Confocal Laser Scanning Microscopy was carried out on a Nikon TI2 microscope equipped with an A1 confocal laser scanning unit, a 1.4 NA 60x Plan Apo objective, PMT, and GaAsP detectors, standard 404, 489, 561, and 637 laser lines, and corresponding filter sets (Nikon). Imaging conditions were optimized for each sample. Scan sizes were adapted to fulfill the criteria for Nyquist-sampling, resulting in a pixel size of 118 nm. The acquisition was run in NIS-Elements, post-processing and image analysis were performed in FIJI.
Weighted Spatial Colocalization Analysis
For the weighted colocalization heatmaps, pixel intensities were calculated, taking into account the absolute intensities in both channels and the ratio between the intensities. The calculation was performed by first normalizing each channel to relative intensity. In the following, the relative intensities of each pixel in both channels a and b were interpreted as a vector  Describing the vector to the position of that pixel in a classical scatter plot. The length of the vector was then multiplied by 1 − | sin(α) − cos(α)| while α is the angle between the vector and the x-axis. This multiplication emphasizes pixels where the two colors colocalize with similar relative intensities. The product then was plotted back to the original pixel position in the image resulting in the heatmap shown in the figures. With this strategy, we could put the information of a 2-channel scatter plot back into the image’s spatial context.
Describing the vector to the position of that pixel in a classical scatter plot. The length of the vector was then multiplied by 1 − | sin(α) − cos(α)| while α is the angle between the vector and the x-axis. This multiplication emphasizes pixels where the two colors colocalize with similar relative intensities. The product then was plotted back to the original pixel position in the image resulting in the heatmap shown in the figures. With this strategy, we could put the information of a 2-channel scatter plot back into the image’s spatial context.
The Jupyter notebook for this analysis is available on GitHub: (https://github.com/QuantitativeVirology/2D-Colocalization)
Gradient purification of HCMV
A 15 cm dish of HFF cells was infected with HCMV-TB40-WT at MOI 0.05. Seven dpi, the infected cells were trypsinized and split onto 16x 15 cm dishes of HFF cells. 7 days after subculturing, the supernatant was harvested and clarified by centrifugation at 1200 xg for 5 min. The virus was pelleted by centrifugation at 14000xg for 1.5 h at 4°C and then resuspended in 1% FBS/PBS overnight on ice. The resuspended virus was centrifuged at 18000xg for 1 min at 4°C to remove large aggregates and then loaded over a continuous gradient made from 15% sodium tartrate with 30% glycerol (w/w) and 35% sodium tartrate (w/w) in 40 mM sodium phosphate pH 7.4 [76]. The gradient was made with a Gradient Master (BioComp Instruments) for an SW41 rotor. After centrifugation at 65000xg for 1.5 h at 4°C, the bands were isolated, diluted 10-fold in PBS, and pelleted at 14000xg for 1.5 h at 4°C. The purified virus pellet was resuspended overnight in PBS and stored at -80°C.
Mass Spectrometry
The purified virus was mixed with 3 volumes of lysis buffer (100 mM Tris, 50 mM DTT, 8 M Urea pH 8.5) and incubated at room temperature for 15 min. Samples were digested using the FASP protocol, employing 30 kDa molecular weight cut-off filter centrifugal units (Amicon, Merck, [77]). Briefly, the lysed virus was added to the centrifugal unit and washed with TU buffer (100 mM Tris, 8 M Urea pH 8.5). Next, 8 mM DTT in TU buffer was added and incubated at 56°C for 15 min. After two further washes, 50 mM iodoacetamide (IAA) in TU buffer was added and incubated for 10 minutes at room temperature. The centrifugal units were washed twice, treated again with DTT, washed once further with TU buffer, and twice with 50 mM ammonium bicarbonate solution. MS grade trypsin (Promega) was added in a 1:100 enzyme:protein ratio, and the sample was incubated overnight at 37°C. The flow-through containing trypsinized peptides was collected and pooled, and the sample was lyophilized with a SpeedVac (Thermo Fisher Scientific). The resulting peptides were enriched with C18 stage tips prepared in-house and eluted with 80% acetonitrile containing 0.5% acetic acid. The samples were dried down by SpeedVac (Thermo Fisher Scientific) and resuspended in 97% water, 3% acetonitrile with 0.1% formic acid, and 10 fmol/µL E. coli digest (Waters Corporation) for analysis by LC-MS/MS.
Peptides resulting from trypsinization were analyzed on a Synapt G2-Si QToF mass spectrometer connected to a NanoAcquity Ultra Performance UPLC system (both Waters Corporation). The data acquisition mode used was mobility enhanced MSE over m/z range 50-2000 with the high energy collisional voltage in the transfer region ramped from 25 to 55 V. Mobile phases used for chromatographic separation were water with 0.1% formic acid (A) and acetonitrile with 0.1% formic acid (B). Samples were desalted using a reverse-phase SYMMETRY C18 trap column (100 Å, 5 µm, 180 µm x 20 mm, Waters Corporation) at a flow rate of 8 µl/min for 2 minutes. Peptides were separated by a linear gradient (0.3 µl/min, 35 °C; 97-60% mobile phase A over 90 minutes) using an Acquity UPLC M-Class Reversed-Phase (1.7 µm Spherical Hybrid, 76 µm x 250 mm, Waters Corporation).
LC-MS data were peak detected and aligned by Progenesis QI for proteomics (Waters Corporation). Proteins were identified by searching against the Human and HCMV proteomes in Uniprot. The database search was performed with the following parameters: mass tolerance was set to software automatic values; enzyme specified as trypsin; up to two missed cleavages; cysteine carbamidomethylation as a fixed modification, with the oxidation of methionine, S/T phosphorylation, and N-terminal acetylation set as variable modifications. Abundances were estimated by Hi3-based quantitation [78].
For comparison with the Turner et al. (2020; [36]) dataset, protein accession was converted to UniParc codes. Raw MS data have been deposited to PRIDE with accession code PXD023444.
Live TIRF Microscopy
For live-cell TIRF imaging, infection experiments were carried out in 35 mm glass-bottom Ibidi dishes. SNAP labeling with SNAP-Cell 647-SIR was done as described in the manual for SNAP-Cell 647-SIR (NEB) before imaging. Microscopy was performed on a Nikon TI equipped for TIRF microscopy and equipped with standard 488, 561, and 640 laser lines, corresponding filter sets, and an incubation chamber with a heating system. The illumination angle was determined experimentally by manually adjusting for TIRF illumination, and image acquisition was performed with NIS-Elements using an ANDOR iXon Ultra 897 EMCCD camera. Live-cell experiments were carried out at 37°C. Intensity measurements in the time courses were done with FIJI by manually placing ROIs. The data analysis and visualization in the graphs were performed in GraphPad Prism 8.
Immunogold labeling
For immunogold labeling of HCMV infected cells, 1x 10 cm cell culture dish of HFF-WT cells were infected with HCMV-WT at an MOI of 0.5. At 4 dpi, the cells were fixed with a mixture of 2% PFA and 0.5% GA (both Science Services) in PBS (Sigma-Aldrich) for 10 minutes at 37°C and 5% CO2. The cells were washed once in PBS and subsequently scraped in 1% gelatin (food grade brand). The cells were pelleted, resuspended in 10% gelatin, and pelleted again while letting the gelatin cool to solidify. The gelatin with the embedded cell pellet was cut into small (1-3 mm) chunks, immersed in 2.3M Sucrose solution, and stored overnight at 4°C. The next day, the pieces were mounted on a sample holder and flash-frozen by immersion in liquid nitrogen. Afterward, the pieces were trimmed and sliced into 70 nm thin sections on a Leica EM FC7 cryo-microtome (Leica) using a diamond knife (Diatome). The sections were recovered by picking them up with a drop of 2.3M Sucrose and transferring them to Formvar and carbon-coated nickel grids, letting the sections thaw in the process. In the following, the sections were immunogold labeled by the following protocol: removal of residual gelatin by incubation in PBS for 20 minutes at 40°C. 3x 2 minutes incubation with PBS, 3x 2 minutes incubation in 0.1% Glycine (Sigma-Aldrich) in PBS, and blocking for 3 minutes in Aurion donkey serum (Aurion/ Science Services). After blocking, the samples were incubated with Ultra-LEAF™ Purified anti-human CD63 H5C6 antibody (Biolegend), diluted 1:5 in Aurion donkey serum, for 30 minutes. Afterward, the samples were washed 5x 2 minutes in PBS and incubated with 10 nm gold coupled donkey-anti-mouse IgG (Aurion/ Science Services), diluted 1:20 in Aurion donkey serum, for 1 hour. Subsequently, the samples were washed 5x 2 minutes in PBS, followed by fixation with 1% GA in PBS for 5 minutes. Afterward, the samples were incubated 10x 1 minute in distilled water and stained, first with uranyl acetate (Merck) in water and secondly with uranyl acetate in 1% methylcellulose. Finally, the grids were air-dried after blotting the uranyl acetate-methylcellulose solution and observed by transmission electron microscopy.
Inhibitor Treatments
U18666A was acquired from Merck, Ketotifen-fumarate, and Tipifarnib were bought from Sigma-Aldrich. The substances were dissolved in DMSO to produce stock solutions (U18666A at 4 mg/ml, Ketotifen at 20 mM, and Tipifarnib at 5 mM), which were subsequently aliquoted and frozen at - 80°C. The drugs were added to the complete growth medium at the indicated time points and at the concentration indicated for each experiment. The medium containing the inhibitor was renewed every 24 hours.
Titrations
To assess the virus titers in supernatant from HCMV infected cells. HFF-WT cells were seeded in 24-well dishes to reach 90-100% on the next day. Tenfold dilutions of the harvested infectious supernatants were made in complete growth medium from 10-1 – 10-4. The medium from the HFF cells was removed and replaced by 100 µl of one the dilutions per well. The plates were rocked gently every 15 minutes for 1 hour to ensure even distribution. After one hour of incubation, an overlay of DMEM containing 2% FCS and 0.6% Methylcellulose (Sigma-Aldrich) was added to the cells, and the plates were incubated at 37°C and 5% CO2 for 14 days. Afterward, the cells were fixed, and fluorescent virus plaques were counted.
Cytotoxicity Assays
HFF-WT cells were seeded on a black 96-well plate to reach confluency on the next day. Then the cells were treated with the indicated substance and concentration for 24 hours. Afterward, the cell viability was measured using the CellTiter-Glo® luminescent cell viability assay (Promega) and a FLUOStar Omega plate reader (BGM Labtech), both according to the instructions from the manufacturer.
Western Blotting
HFF-WT cells were infected with HCMV-WT (MOI = 3), cells were harvested and lysed at 1 hpi (input) and every 24 hours until 96 hpi. SDS-PAGE was performed on Bio-Rad Mini-PROTEAN TGX 4-15% gels (Bio-Rad). Separated protein was blotted on Amersham Protran 0.45 µm nitrocellulose membranes (Cytiva). The membrane was cut, and the sections were subsequently stained with one of the primary antibodies against HCMV pp150 (kind gift by Eva-Maria Borst and Stipan Jonjic), anti-CMV ICP36 monoclonal antibody 10D8 (Virusys), and anti-IE1/2 (hybridoma supernatant [79], kind gift by Wolfram Brune) followed by a secondary antibody Goat Anti-Mouse IgG StarBright Blue 700 (Bio-Rad). The stained blots were imaged using a ChemiDoc MP imager (Bio-Rad).
Acknowledgments
The authors declare no competing interests.

HFF cells were infected with HCMV-pp150-EGFP-gM-mCherry (MOI 3) and processed for EM 4 dpi. Image signals were inverted to facilitate comparison with TEM images. N marks nucleoplasm, C indicates cytoplasm, and Ex the extracellular space. Highlighted in the panels are examples of B-capsids in the nucleus (unfilled black triangles), DNA-filled nuclear C- capsids (white triangles with black contour), cytoplasmic non-enveloped C-capsids (black triangles), intracellular, enveloped virus particles (black filled arrowheads) as well as enveloped, released particles (empty arrowhead with black contour). Scale bar lengths are specified in each image.

S2A Single SBF-SEM section of an infected HFF cell. HFF cell infected with HCMV- pp150-EGFP-gM-mCherry (MOI 3) at 4dpi. Image signals were inverted to facilitate comparison with TEM images. The white frame indicates the area cropped and enlarged in B, showing the surface of an MViB in the periphery of the assembly complex. Scale bar indicates 10 µm. S2B A detail showing a single particle budding into an MViB (white arrowhead). Scale bar indicates 200nm.

HFF cell infected with HCMV-pp150-EGFP-gM-mCherry (MOI 3) at 4dpi. Shown is the assembly complex in a resliced section through an SBF-SEM stack. Scale bar indicates 1.5 µm. Image signals were inverted to facilitate comparison with TEM images.

S4A Single SBF-SEM section of an infected HFF cell. HFF cell infected with HCMV-pp150-EGFP-gM-mCherry (MOI 3) at 4dpi. The white frame indicates the area cropped and enlarged in B. Scale bar indicates 10 µm. S4B A detail showing a single capsid budding into a single vesicle (white arrowhead). Scale bar indicates 200 nm. Image signals were inverted to facilitate comparison with TEM images.
S5A HFF-CD63- pHluorin cells were infected at an MOI of 1 with HCMV-pp150-SNAP-gM-mScarlet-I. Cells were fixed at 4 dpi and stained for gB. The images show a representative cell and the localization pattern of the cellular MVB marker CD63 in relation to the viral glycoproteins gB and gM. CD63 localizes to large vesicles positive for gB and gM. Scale bars indicate 10 µm. The green line indicates the section quantified in S5B. S5B Line plot for the indicated areas in S5A. CD63 signal correlates with gM and gB signals in two MViBs. S5C Spatial weighted colocalization analysis highlights the specific areas for CD63 and gB colocalization. Scale bar indicates 10 µm.

Pathway analysis of the mass spectrometry data from purified virions, done with string-db.org. The color of the dots indicates factors from either GO-term associated pathways or publications related to their functionality. The colors of the connections indicate the type of evidence for the interactions and are filtered for the highest interaction confidence (0.900) as provided by the database.

S7A-B HFF cells were infected at an MOI of 1 with HCMV-pp150-SNAP-gM- mScarlet-I. Cells were fixed at 4 dpi and stained with specific antibodies for CD9 (⍺CD9) and CD81 (⍺CD81). The images show representative cells and the localization pattern of the CD molecules relative to gM (gM-mScarlet-I) and pp150 (pp150-SNAP). Scale bars indicate 10 µm.

HFF cells were infected with HCMV-TB40-WT at an MOI of 0.5. After 4 dpi, the cells were fixed and processed for immunogold labeling against CD63. Note that membranes appear white in this preparation method, and low GA concentrations used to preserve epitopes might lead to less preservation of MViB contents. S8A-G Shown is large bodies containing virus particles (black triangles), dense bodies (white-filled triangles), and 10 nm gold particles (arrowheads). S8H A body with the classical phenotype of an MVB decorated with 10 nm gold particles (arrowheads). All scale bars indicate 0.2 µm.

S9A-B HFF cells were infected with HCMV-TB40-pp150-SNAP-gM- mScarlet-I at an MOI of 2 and treated with the indicated substance at the indicated concentrations until 5 dpi. The medium containing the inhibitors was refreshed every 24 hours. 5 dpi cells were fixed, labeled with SNAP-Cell-SiR, and imaged by spinning-disk microscopy. White triangles indicate EVAs. Scale bars indicate 20 µm. S9C HFF cells were infected with HCMV-TB40-pp150- SNAP-gM-mScarlet-I at an MOI of 2 and treated with the indicated inhibitors at the indicated concentrations until 4 dpi. The medium containing the inhibitors was refreshed every 24 hours. At 4 dpi, the supernatant was collected and titrated on HFF cells. Bars show mean, and error bars indicate standard deviation. Statistical significance was probed using one-way ANOVA (p-values: Tipifarnib: <0.0001, U18666A: 0.4154, Ketotifen: 0.8364) and Dunnet’s multiple comparisons tests (shown in the figure). S9D HFF cells were treated as described for S9 A-B. Large overviews were created by spinning-disk microscopy, and EVAs were quantified. Bars show mean, and error bars indicate standard deviation. Statistical significance was calculated using a one-way ANOVA (p = 0.0179; in total, 687 late infected cells from triplicates were counted) and Dunnet’s multiple comparisons test (shown in the figure). S9E HFF were treated with the indicated substance at the indicated concentration for 24 hours. Cell viability was measured with an ATP assay at 24 hpi. The apoptosis inducer Staurosporine was used as a positive control. Bars show mean, and error bars indicate standard deviation. Statistical analysis by a 2-way ANOVA confirmed statistically significant differences in the viabilities of the three groups (p-value < 0.0001). The cytotoxicity of Tipifarnib was not significantly different from the vehicle control, as determined by Tukey’s multiple comparisons test. In contrast, the change in cell viability of Staurosporin was significant in the same analysis. S9F Western blot of HFF cells infected with HCMV-TB40-WT (MOI = 3) and treated with 1 µM Tipifarnib or DMSO (0.01%; vehicle control). At each indicated time point (input = 1 hpi), the cells were harvested and lysed, and the blot was probed for pp150 as a late protein, IE1/2 as immediate-early gene products, or UL44 as an early gene. GAPDH served as the loading control.
Supplementary Video 1. SBF-SEM of an area between the cell surface and the growth substrate. The video shows a subset of planes from the dataset described in Fig. 1 rendered as a video. Infection conditions are as described before. The signal was inverted to resemble TEM contrast. Shown is a large invagination below the cell at the growth substrate.
Supplementary Video 2. 3D-rendering of an MViB from SBF-SEM data. In this video an MViB from the SBF-SEM dataset described in Fig. 2 is rendered in 3D. The yellow surface marks the limiting membrane of the multivesicular structures. The contents are rendered as surfaces in different colors to show the heterogeneity of the MViB cargo. Virions are rendered in dark green, dense bodies in cyan, and other vesicular material in magenta. Scale bar indicates 600 nm.
Supplementary Videos 3A and 3B. SBF-SEM rendering of infected HFF cells. The video shows an excerpt from the dataset described in Fig. 1. HFF cells were infected with an MOI of 3 and fixed 4dpi. 3A Overview rendering of the whole SBF-SEM dataset of the cells shown in Fig. 1 and 2. 3B A group of prominent virus-filled MVBs is highlighted by a surface rendering. Several more MVBs are present in the cell.
Supplementary Video 4. Stack of an MViB and associated immature particles. This video shows an MViB from the SBF-SEM dataset shown in Fig. 1. Cells were infected and treated as described before. White triangles indicate immature, non-enveloped particles in close proximity or directly associated with the multivesicular structure next to the nucleus. Scale bar indicates 0.2 µm.
Supplementary Video 5. Multi-perspective 3D rendering of volumetric time-lapse microscopy data of HCMV release. HFF cell, infected with HCMV-pp150-EGFP-gM-mCherry as described in Fig. 3A. The video shows several perspectives on how a large MViB positive for pp150-EGFP (green) and gM-mCherry (red) traverses the cytoplasm and fuses with the plasma membrane. The first seconds show the 3D video, followed by a split-screen part of three different perspectives. A spotlight effect (circle) highlights the same body in all three parts. In the left third, the MViB is followed by a moving section parallel to the growth substrate, through the volume on its way downwards to the lower cell surface. In the middle part, the body is followed as a 3D rendering through the cell. The camera angle moves to keep the body visible as well as possible. The last third shows how the MViB fuses with the plasma membrane in a static cross-section. Due to the optical setup of the lattice-light-sheet microscope (See Fig. 3A), the grid added by Arivis 4D (Arivis AG, Rostock, Germany) is tilted 30° respective to the real physical orientation of the cell in the microscope.
Supplementary Videos 6 and 7. Live-cell long time-lapse spinning-disk microscopy videos. HFF cells were infected with HCMV-pp150-SNAP-gM-mScarlet-I at an MOI of 1. At 72 hpi, cells were stained for pp150-SNAP and imaged live by spinning-disk microscopy. 8-micrometer stacks in 1-micrometer increments were acquired every 40 minutes. The plane shown is the section of the cell closest to the coverglass. Cells can be seen to release virus particles in short intermittent bursts over several hours, indicated by the white arrowheads. pp150-SNAP labeling is shown in green and gM-mScarlet-I label in magenta. The time format is hh:mm.
Supplementary Video 8. 3D rendering of immunofluorescence data. In this video, the IF dataset from Sup. Fig. 6 is rendered in 3D. The coloring scheme is the same as in Sup. Fig. 6. The 3D rendering shows the 3-dimensional correlation between the molecules.
Supplementary Table 1 Mass Spectrometry Results
Acknowledgments
We thank Wolfram Brune, Christian Sinzger, and Kerstin Sampaio for their generous gift of viruses HCMV-TB-40-BAC4, HCMV-pp150-EGFP-gM-mCherry, reagents, and their support.
This study was funded by the Wellcome Trust through a Collaborative Award (209250/Z/17/Z) to KT, KG, and JBB. KG and JBB are funded by the Deutsche Forschungsgemeinschaft (DFG, German Research Foundation) under Germany’s Excellence Strategy – EXC 2155 – project number 390874280. We thank the DFG for funding the lattice light sheet system through a large equipment grant to KG and JB, project number 413831413. We thank Zeiss for including us in their lattice light-sheet early adaptor program. FJF is holding a graduate student fellowship by the Studienstiftung des deutschen Volkes. The Leibniz Institute for Experimental Virology is supported by the Free and Hanseatic City of Hamburg and the Federal Ministry of Health. KG is further funded by the Free Hanseatic City of Hamburg (grant LFF-FV 71-2019). This study is part of the Leibniz ScienceCampus InterACt (Grant Agreement No. W6/2018). The mass spectrometer used in this study was funded by a Wellcome Trust instrumentation grant 104913/Z/14/Z to KT.
Footnotes
The manuscript has been significantly revised due to suggestions from peer reviewing experts. Experiments were added and the figures updated accordingly. All sections have been updated.
References
- 1.↵
- 2.↵
- 3.↵
- 4.↵
- 5.↵
- 6.
- 7.
- 8.
- 9.↵
- 10.↵
- 11.↵
- 12.↵
- 13.↵
- 14.↵
- 15.↵
- 16.↵
- 17.↵
- 18.↵
- 19.↵
- 20.↵
- 21.↵
- 22.↵
- 23.↵
- 24.↵
- 25.↵
- 26.↵
- 27.↵
- 28.↵
- 29.
- 30.↵
- 31.↵
- 32.↵
- 33.↵
- 34.↵
- 35.↵
- 36.↵
- 37.↵
- 38.↵
- 39.↵
- 40.↵
- 41.↵
- 42.↵
- 43.↵
- 44.↵
- 45.↵
- 46.↵
- 47.↵
- 48.↵
- 49.
- 50.↵
- 51.↵
- 52.
- 53.↵
- 54.↵
- 55.↵
- 56.↵
- 57.↵
- 58.↵
- 59.↵
- 60.↵
- 61.↵
- 62.↵
- 63.↵
- 64.↵
- 65.↵
- 66.↵
- 67.↵
- 68.↵
- 69.↵
- 70.
- 71.↵
- 72.↵
- 73.↵
- 74.↵
- 75.↵
- 76.↵
- 77.↵
- 78.↵
- 79.↵





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}