

Abstract
Tumour-cell detachment is a critical early event in the metastatic cascade. Although several mechanisms have been reported, the role of cell mechanical properties in facilitating cell detachment and migration is not well understood. This exploratory study aimed to assess how intracellular stiffness changes these processes. MDA-MB-231 cells were embedded as 10,000-cell spheroids in 2 and 4 mg/ml collagen matrices. Using mitochondrial-based particle tracking microrheology, the intracellular stiffness was assessed of cells that migrated distances equivalent to four and six times the cell diameter (dC) from the spheroid and compared to cells at the spheroid surface (0dC), representing medium, high and no migration, respectively. The mitochondrial mean square displacement (MSD) and intracellular stiffness decreased during detachment and migration for both collagen concentrations (i.e. rigidities). The MSD of 4dC and 6dC cells was similar, whereas cell stiffness was lower for 4dC than for 6dC cells. The stiffness of 0dC cells decreased with increasing matrix rigidity, whereas matrix rigidity did not affect the stiffness of 4dC and 6dC cells. It is proposed that decreased cell stiffness drives cellular detachment and migration and increased matrix rigidity physically hinders migration where cells need to either decrease their stiffness or remodel the environment to migrate. The absence of an influence of matrix rigidity on the stiffness of migrated cells suggests that cells facilitate migration by remodelling their environment through the cleavage of matrix proteins. Proposed targets for further mechanobiological studies of metastatic cancer cells include the expression of matrix metalloproteinases and transforming growth factor β, and the role of cell volume on detachment and migration in matrices with varying pore sizes.
Insight This study suggests that decreased intracellular stiffness facilitates the detachment of metastatic breast cancer cells from clusters and subsequent migration in three-dimensional extracellular matrices (ECM). The cellular softening during detachment is more prominent in stiffer ECMs, whereas the stiffness of the migrating cells is not affected by the environmental mechanics. These findings, complemented with further investigations into identified biochemical and morphological targets, can offer the potential for therapies inhibiting breast cancer metastasis targeted to the tissues of the secondary tumour sites.
1. Introduction
Metastatic cancer cells alternate between various labour-intensive tasks associated with disease progression, including detachment from the tumour, invasion into the tumour environment and formation of new tumours (Gal and Weihs 2012). Considerable attention has focussed on understanding the role that the mechanical properties of cancer cells play in these processes. It has been shown that the mechanical properties have crucial implications in regulating chemical and mechanical cues and in maintaining cell or tissue architecture in several cell types, including metastatic cancerous cells (Cui et al. 2017, Mak et al. 2014).
Many of these studies have probed cells in two-dimensional environments. However, cells in two-dimensional (2D) and three-dimensional (3D) environments exhibit substantial differences in morphology, gene expression, and mechanical and structural properties. Several studies reported a marked decrease in intracellular fluctuations from 2D to 3D environments (Baker et al. 2009, Mak et al. 2014). Additionally, 3D in vitro systems can recapitulate in vivo conditions more closely than 2D systems (Baker et al. 2009).
Therefore, studies of solid tumours should incorporate 3D models that mimic complex tumour architecture, including cell-cell and cell-matrix interactions (Close et al. 2018). Cell spheroids are powerful in mimicking many aspects of solid tumours, including architecture (i.e. an outer proliferative layer of cells, a middle layer of senescent cells and a core of necrotic cells), enhanced cell-cell and cell-matrix connections, and gene expression (Costa et al. 2016, Fennema et al. 2013).
Tumour spheroid assays have addressed essential questions relating to spheroid morphology, growth, viability, drug development (Close et al. 2018), signalling pathways associated with cell detachment and invasion (Naber et al. 2012), cancer stem cell content (Reynolds et al. 2017). However, few studies have investigated cell mechanics in the detachment and invasion process due to the technical challenges of quantifying the mechanics of live cells in 3D environments during detachment and invasion. Microrheology can offer non-invasive techniques that allow passively probing intra-cellular mechanics to characterise single and collective cell behaviour without introducing cellular and matrix deformation (Mak et al. 2014).
It is generally accepted that high tissue rigidity is a characteristic of solid tumours (Wullkopf et al. 2018) and that cells adapt their stiffness to environmental stiffness (Mitrossilis et al. 2010). It is essential to investigate whether and how cells adapt their stiffness during tumour detachment and invasion as the disease progresses. It has been shown that reduced cell-cell adhesion decreases cell stiffness (Omidvar et al. 2014) and increases migration (Geiger et al. 2019) and that increased migration and decreased cell stiffness are directly correlated (Guck et al. 2005, Remmerbach et al. 2009). However, it is not yet known whether these observations of cellular mechanics translate directly to physiologically relevant environments that mimic the tumour-cell detachment process. Considering that the mechanical properties of cells depend on the properties of the extracellular matrix (ECM), it is essential to clarify how cells regulate their mechanical properties when the properties of the ECM change.
This study aimed to explore the role of intracellular stiffness of metastatic breast cancer cells during detachment from spheroids and invasion, and to identify targets for further research to complement this initial work. Different levels of migratory capacity were established by considering cells increasingly migratory with increasing migration distance from the spheroid.
2. Materials and methods
2.1. Cell culture
Metastatic breast adenocarcinoma cells, MDA-MB-231, were cultured in DMEM (Dulbecco’s Modified Eagle Medium, Life Technologies), supplemented with 10% foetal bovine serum (FBS) and 1% penicillin-streptomycin, and maintained in 25 cm2 tissue flasks (Costar, Corning Life Science, Acton, MA) at 37°C and 5% CO2 until near-confluency. Media was replaced every 3-4 days. Cells were used in passages 6-16.
2.2. Spheroid formation
Cell suspensions were grown in agarose-coated well plates to form spheroids according to the liquid overlay method (Metzger et al. 2011). An agarose solution was prepared by diluting 0.18 g agarose (Sigma-Aldrich, St. Louis, MO) in 12 ml 1XPBS and heated in a microwave oven for 36 s or until the agarose dissolved. The solution was immediately transferred to sterile conditions and kept on a hot plate to prevent premature gelation. A 96-well plate was prepared by pipetting 70 μl of the hot liquid agarose solution onto the flat bottoms of the wells, ensuring the solution covered the entire surface and contained no air bubbles. The agarose-coated 96-well plate was exposed to ultraviolet light for 30 minutes for sterilisation. Separately, cell monolayers were lifted from the culture flask using standard laboratory trypsinisation procedures. Spheroids of 10,000 cells were created by placing 100 μl trypsinised cells (100,000 cells/ml containing 2.5% Matrigel) into the agarose-coated wells, centrifuged at 1,000 rpm for 5 mins, and incubated at 37°C and 5% CO2 for 72 hrs.
2.3. Spheroid collagen embedding
MDA-MB-231 spheroids were embedded in Type I Rat Tail Collagen Solutions (BD Biosciences) at 2 and 4 mg/ml final collagen concentrations. Type I Rat Tail Collagen at a stock solution of 9.61 mg/ml was combined with equal amounts of neutralising solution (100 mM HEPES in 2X PBS with pH 7.3) and further diluted with 1XPBS on ice to create 2 and 4 mg/ml collagen solutions. Spheroids were added to the unpolymerised collagen solutions on 35mm glass-bottom dishes, with each 50 μl collagen solution containing one spheroid to avoid spheroid-spheroid interaction, and incubated at 37°C and 5% CO2 for 1 hr. After 1 hr, the collagen solutions had polymerised, and 500 μl media were added.
2.4. Mitochondrial particle tracking microrheology
The use of mitochondria as tracer particles to determine the viscoelastic response of cells has been validated byMak et al. (2014), who showed that the mean square displacement (MSD) of mitochondria and ballistically injected nanoparticles was similar when used in metastatic breast cancer MDA-MB-231 cells.
Embedded MDA-MB-231 tumour spheroids were prepared for mitochondrial tracking by adding 500 nM Mitotracker Red solution (Life Technologies, Carlsbad, CA) to the supplemented DMEM media 24 hrs before experimentation to allow the Mitotracker solution to sufficiently diffuse through the spheroid. Before imaging, the media was removed from the wells, and the wells were placed in an environmental chamber at 37°C and 5% CO2.
After 24 hrs, cells started detaching from the spheroid and migrating into the collagen gel predominantly as linear branches with a leader cell and follower cells.
To avoid boundary effects of the rigid 2D glass-bottom surface and to ensure full engulfment, cell branches near the middle of the collagen gel were located and imaged. Mitochondrial fluctuations of branching cells at the spheroid surface and varying distances from the spheroid were captured using time-lapse fluorescent imaging for 120 s (exposure time: 50 ms/frame; spinning disk confocal microscope, 63x 1.4 oil immersion objective and CCD camera, Hamamatsu Photonics, Hamamatsu, Japan). Fluorescent images were acquired of surface and migrated cells (branched). The distance migrated (dm) from the spheroid was reported in cell diameter (dC), where dm = 0dC, 4dC and 6dC correspond to the cell at the spheroid surface, intermediately migrated, and furthest migrated, respectively. These experiments were repeated on two independent days (n = 2), with data taken from at least two spheroids, each with at least five branches, resulting in 10 cells per condition. Each branch comprised at least six migrated cells. Cells with at least 60 mitochondria, visible for at least 80% of the total image acquisition time, were retained for analysis.
Post-processing involved constructing mitochondrial trajectories using TrackMate (Tinevez et al. 2017) in Fiji Image J (Schindelin et al. 2012). The tracking assumed particles with a diameter of 1 μm and allowed up to 4 missed frames between subsequent spots to account for missing detections. Mitochondrial trajectories were imported into MATLAB (The MathWorks, Natick, MA), and the ensemble-averaged, time-dependent MSD was calculated using @msdanalyzer (Tarantino et al. 2014) according to:

Here, 〈Δr2 (τ)xy〉 is the ensemble-averaged MSD, τ is the time interval or delay time between the first and last image frame used for the analysis, and x(t) and y(t) refer to the spatial coordinates of particle positions at time t.
For viscoelastic materials, the MSD scales nonlinearly with the delay time τ according to a power-law relationship, 〈Δr2 (τ)〉 ~ τα (Baker et al. 2010). The power-law coefficient α = ∂ln 〈Δr2 (τ)〉/∂ln (τ) represents the slope of the logarithmic MSD-τ curve.
2.5. Statistical analysis
Data are reported as mean ± standard deviation in the text. Error bars in graphs represent the standard error of the mean (SEM). Two-way ANOVA was employed to detect differences in MSD and α of cells at different delay times. ANOVA assessed two main effects, i.e. distance from the spheroid and changes in collagen concentration, and one interaction between main effects. The migrated distance contained three levels, i.e. no migration, medium migration, and large migration, with which posthoc analysis was performed using Tukey’s HSD test. The collagen concentration contained two levels, i.e. 2 mg/ml and 4 mg/ml collagen, and ANOVA was followed up with linearly independent pairwise comparisons among estimated marginal means.
A criterion for statistical significance of p < .05 formed the basis of all evaluations. SPSS Statistics for Windows (Version 25.0, IBM Corp., Armonk, NY) was used for all statistical analyses.
3. Results
To provide a platform for cell detachment from tumours and migration in physiologically relevant environments, metastatic breast cancer cells (MDA-MB-231) were embedded as tumour spheroids in 3D collagen matrices. Mitochondrial particle tracking microrheology experiments (n = 2) were conducted after 24 hrs of incubation when cells started detaching from spheroids.
A low MSD value corresponds to constrained particle fluctuations and indicates a stiffer, solid material, whereas a high MSD indicates greater particle motility and a more fluid-like material. For short delay times, the mitochondrial MSD primarily represents the viscoelastic intracellular properties, whereas active intracellular motor-driven processes dominate the MSD for long delay times (Kim et al. 2018, Mak et al. 2014). The power-law coefficient α helps classify the motion of the tracer particles. An α close to 1 corresponds to diffusive motion such as thermal fluctuations in Newtonian fluids, whereas an α close to 0 indicates constrained, sub-diffusive motion such as thermal fluctuations in an elastic material (Kim et al. 2018, Mason et al. 1997).
Following detachment from the tumour surface, cells invaded the tumour environment predominantly in linear branches. Several cells trailed a leader cell in these branches and migrated away from the tumour (Figure 1).

Illustration of detaching (surface) and migrated (branched) cells as (a) bright-field image (5x magnification) and (b) mitochondrial fluorescent image (63x magnification). The blue dashed line indicates the spheroid border, green dashed circles indicate the approximate diameter of cells migrating away from the spheroid in a branch, and the yellow dashed circle indicates a cell out of focus.
3.1. Effect of detachment and migration: Migrated cells are more deformable than detaching cells
The effect of detachment and migration of cells on mitochondrial fluctuations and cell stiffness was assessed in 2 and 4 mg/ml collagen.
The MSD of 0dC, 4dC, and 6dC cells in 2 and 4 mg/ml collagen increased with increasing delay time (Error! Reference source not found.). Lower and flat MSD-τ curves indicate solid-like intracellular behaviour, whereas higher and increasingly sloped curves indicate fluid-like, intracellular motor-driven mechanics.
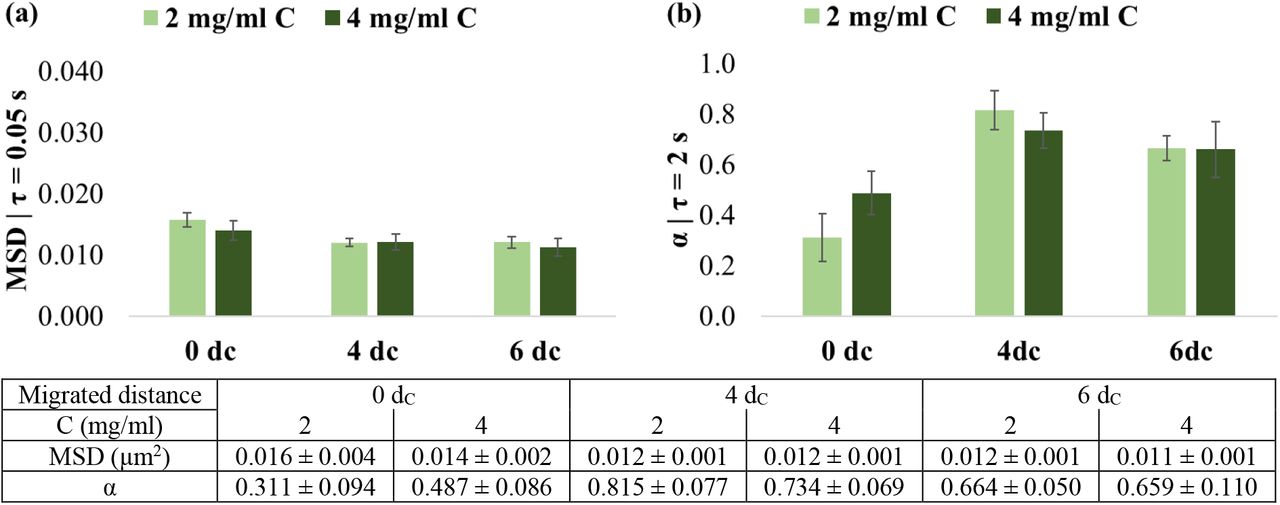
MSD and α were further compared for short delay times of τ = 0.05 s and 2 s, respectively, to determine the viscoelastic properties of cells as the distance from the spheroid increased (Error! Reference source not found.).
In 2 mg/ml collagen matrices, mitochondrial fluctuations (at τ = 0.05 s) were larger, and the power-law coefficient α (at τ = 2 s) was smaller for cells at the spheroid surface, i.e. dm = 0dC, than for cells that had detached from the spheroid and invaded the environment (Error! Reference source not found.).
The MSD in 0dC cells was 0.016 ± 0.004 μm2 compared to 4dC cells with 0.012 ± 0.002 μm2 (p = .012) and 6dC cells with 0.012 ± 0.003 μm2 (p = .010). The MSD of 4dc and 6dc were similar (p = .946).
The α was lower of in 0dC cells (0.311 ± 0.298) than in 4dC (0.815 ± 0.243, p < .001) and 6dC cells (0.664 ± 0.159, p = .003), with a decrease in α in cells from 4dC to 6dC, although non-significant (p = .173). The mitochondrial motion was strongly sub-diffusive in 0dC cells, closely diffusive in 4dC cells, and weakly sub-diffusive in 6dC cells.
In the 4 mg/ml collagen environment, the MSD of the 0dC cells of 0.014 ± 0.005 μm2 was slightly larger than for 4dC (0.012 ± 0.004 μm2) and 6dC cells (0.011 ± 0.004 μm2). These differences were, however, not significant (p = .436). The observations for α were similar to those in the 2 mg/ml collagen, although differences were not significant (p = .151). The 0dC cells exhibited lower α (0.487 ± 0.284) than the 4dC (0.734 ± 0.220) and 6dC cells (0.659 ± 0.349). These α values indicated weaker sub-diffusive motion for the migrated 4dC and 6dC cells than the 0dC cells at the spheroid surface.
3.2. Matrix rigidity affects cell stiffness during detachment but not during subsequent migration
The effect of collagen concentration of the matrix on mitochondrial fluctuations and cell stiffness was assessed for varying migration distances from the tumour spheroids.
The increased collagen concentration and rigidity of the matrix did not affect the mitochondrial fluctuations of the cells. The MSD of 0dC, 4dC and 6dC cells in 4 mg/ml was similar to that in 2 mg/ml collagen (p = .290, p = .773 and p = .426, respectively) (Error! Reference source not found.a).
An increase, although not significant, in intracellular power-law coefficient α with increased collagen concentration was observed for the 0dC cells at the spheroid surface (p = .182) but not for the migrated 4dC (p = .444) and 6dC cells (p = .964) (Error! Reference source not found.b).
4. Discussion
In the present study, metastatic breast adenocarcinoma cells, MDA-MB-231, were embedded as tumour spheroids in collagen gels of two concentrations (2 and 4 mg/ml collagen) to investigate the mechanical properties of the cancer cells associated with tumour-cell detachment and migration in physiologically relevant 3D environments. Mitochondrial-based particle tracking microrheology was utilised to probe intracellular fluctuations. Cells were distinguished between cells attached to the spheroid, i.e. before tumour-cell detachment, and migrated cells that have employed the necessary processes to detach and gain enhanced migratory properties for cell dissemination away from the tumour spheroid. Cells were denoted according to the distance migrated from the spheroid, normalised to cell diameter (dC), where 0dC, 4dC and 6dC represented no, medium and high migration, respectively.
This study extends work on the microrheological characterisation of breast cancer cells in 3D environments. MDA-MB-231 cells were used since their mechanics has been extensively studied as single cells in 2D and 3D environments (Baker et al. 2009, Mak et al. 2014) and with and without drug treatment (Kim et al. 2018, Wullkopf et al. 2018). However, the intracellular mechanics that aid cells in detaching from the tumour and invading the surrounding matrix is not well studied. Here, the microrheology of cells during physiological relevant processes such as tumour detachment and directed migration were characterised in 3D environments.
This study indicates that cells are more solid-like before tumour detachment than after detachment and migration into the surrounding matrix. The cells at the spheroid surface (i.e. 0dC) displayed a lower power-law coefficient α, alluding to increased cell stiffness, than migrated 4dC and 6dC cells in both 2 and 4 mg/ml collagen concentrations. In the context of migration, studies have shown an inverse relationship between migratory potential and cell stiffness, irrespective of whether different grades of cancers were from the same cancer type or identical tumour specimens were used (Swaminathan et al. 2011, Xu et al. 2012).
Wullkopf et al. (2018) reported the stiffness of highly invasive breast cancer (4T1) and pancreatic cancer (KPR172HC) cells in the spheroid centre, branch stalk and branch tip during the invasion in compliant and rigid collagen matrices. Here, the spheroid centre, branch stalk and branch tip represented no, medium and large migration, respectively. Irrespective of the matrix rigidity, tip cells that had migrated furthest exhibited lower stiffness than cells in the spheroid centre (Wullkopf et al. 2018). These results support the findings in the current study. However, the stiffness of 6dC cells was higher than that of 4dC cells that had migrated a shorter distance. In addition, the cells’ diffusivity α increased from 0dC to 4dC and decreased from 4dC to 6dC irrespective of collagen concentration.
The 0dC cells were attached to the spheroid through cell-cell adhesion, whereas 4dC and 6dC cells detached from the spheroid and migrated away. Hence migrated cells are less mechanically affected by the spheroid and adapt to the matrix rigidity. Interestingly, the cell stiffness at 6dC was higher than at 4dC distance from the spheroid. MDA-MB-231 cells require actomyosin-based contractility to invade a 3D environment despite their spherical morphology (Poincloux et al. 2011). Therefore, it is proposed that the farther migration of 6dC cells compared to 4dC cells requires increased actomyosin contractility and, hence, increased cell stiffness. Furthermore, the mitochondrial fluctuations in migrated 4dC and 6dC cells neared or entered the diffusive regime much earlier than in non-migrated 0dC cells, indicating heightened motor activity to contract the cytoskeleton in preparation for migration and invasion (Guo et al. 2014).
In the lower collagen concentration (2 mg/ml), the mitochondrial motion was strongly sub-diffusive (α < 0.3) in 0dC cells, diffusive (α = 1.0) in 4dC cells, and weakly sub-diffusive (0.7 < α < 1.0) in 6dC cells. In comparison, the mitochondrial motion was sub-diffusive in 0dC cells and weakly sub-diffusive in 4dC and 6dC cells in the higher collagen concentration (4 mg/ml). These observations indicate that weakly sub-diffusive to diffusive behaviour is characteristic of cells that have migrated away from their spheroid.
The migrating cells exhibited amoeboid motility and moved in a straight path away from the spheroid. Since amoeboid migration is driven by increased myosin II-mediated contractility (Poincloux et al. 2011), it is suggested that this contractility mechanism underlies the diffusive mitochondrial motion in migrating cells in this study.
Increased matrix rigidity played a role in decreasing the stiffness of cells at the spheroid surface but did not affect the stiffness of migrated cells. The intracellular power-law coefficient α of 0dC cells was lower in 2 mg/ml than in 4 mg/ml collagen, whereas the α of both 4dC and 6dC cells was similar in 2 and 4 mg/ml collagen matrices. Increased matrix rigidity has been reported to increase intracellular activity and decrease the stiffness of isolated cells in 3D environments (Baker et al. 2009, Butcher et al. 2009). The stiffness of 0dC cells at the spheroid surface is consistent with these reports. For migrated cells, the similar stiffness in soft and stiff matrices for 4dC and 6dC cells, respectively, is consistent with Wullkopf et al. (2018). They found that the stiffness of detached and migrated invasive breast cancer 4T1 and pancreatic cancer KPR172HC cells were similar in collagen matrices of high and low rigidity. In conjunction with the effect of migrated distance, this finding suggests that cells minimise their stiffness during migration irrespective of the matrix rigidity.
The results of this study are consistent with several reports that suggest that decreased cell stiffness mediates the escape of breast cancer cells from the initial metastatic niche (Guck et al. 2005, Remmerbach et al. 2009). However, this study should be repeated with different cancers to validate the current findings and provide a more comprehensive understanding of the adjustment of intracellular mechanics during cell detachment and invasion.
A target for further research is the interplay of cellular stiffness, ECM remodelling, and biochemical signalling to promote cancer invasion. Matrix metalloprotease (MMP) expression often increases with cancer progression (Deryugina and Quigley 2006), and MMPs affect the remodelling of the ECM by cancer cells, particularly in dense matrices (Harjanto et al. 2011). Transforming growth factor β (TGFβ) promotes cancer invasion at the advanced disease stage (Massagué 2008), e.g. through upregulation of MMPs in breast cancer (Naber et al. 2012, Wiercinska et al. 2011). Combining cellular microrheology and assessment of MMP and TGFβ expression in spheroid invasion assays will provide more insight into how cancer cells regulate mechanical and biochemical cues to metastasise.
Cell volume is a modulator of cell adhesion and detachment (Yang and Jiang 2018). Wang et al. (2020) found a decrease in volume and motility of metastatic breast cancer cells in a 3D ECM with increasing stiffness but constant adhesion site density. In the current study, the change in collagen concentration affected the pore size and adhesion site density of the collagen fibre network. It is thus of interest to assess the relationship of pore size of the collagen fibre network and cell volume to complement the findings of intracellular mechanics.
5. Conclusion
This exploratory study demonstrated that mitochondrial particle tracking microrheology could detect stiffness changes in metastatic breast cancer cells as they detach and migrate away from their spheroids in soft and stiff collagen matrices. The findings revealed the collective effects of enhanced migratory conditions and increased matrix rigidity on the mechanical properties of the cells. The study identified targets for further mechanobiological investigations, including matrix metalloproteases, TGFβ and cell volume, towards therapeutics that target cancer cells at the initial tumour site to inhibit metastatic spread.

Intracellular fluctuations from spheroids (n = 2) in 2 and 4 mg/ml collagen gels. MSD and power-law coefficient α versus delay time of cells in 2 mg/ml collagen (a, b) and 4 mg/ml collagen (c, d). Error bars omitted for clarity. Arrows indicate the delay time of τ = 0.05 s for MSD and τ = 2 s for α. Trends of MSD and α of cells were similar in 2 and 4 mg/ml collagen. Mitochondrial fluctuations of 0dC were larger than migrated 4dC and 6dC cells. 0dC cells exhibited the largest cell stiffness, followed by 6dC and then 4dC cells.
{kind=link}
{kind=link}
{kind=link}
Comparison of (a) mean square displacement (MSD) and (b) power-law coefficient α of cells at the spheroid surface (0dC) and after migration of a distance of 4dC and 6dC from the spheroid surface in 2 and 4 mg/ml collagen matrices (C) (dC indicates cell diameter). All differences were non-significant. Values are mean ± SEM; error bars are SEM.
Funding
This study was supported financially by the National Research Foundation of South Africa (grant number IFR14011761118 to TF), the South African Medical Research Council (grant number SIR328148 to TF) and the National Institutes of Health [grant numbers U01CA202123 and P01HL120839 to MHZ]. This research was also supported in part by the National Institutes of Health training grant at Boston University, T32 EB006359 that supported the work of JEK. GH acknowledges a Max & Lillie Sonnenberg International Travel Scholarship and an FHS FRC Postgraduate Publication Incentive Award of the University of Cape Town. Any opinion, findings, conclusions, or recommendations expressed in this publication are those of the authors and do not necessarily represent the official views of the funding agencies.
Conflicts of interest statement
The authors declare that they do not have conflicts of interest.
Availability of data
Data presented in this article are available on ZivaHub http://doi.org/10.25375/uct.13359398.
Footnotes
References




