

ABSTRACT
BACKGROUND The application of RNAi to control pests has attracted the attention of researchers. Our results indicated that knockout of PxRdl2 can decrease resistance to fipronil in Plutella xylostella, providing a suitable target gene for RNAi-based pest control.
RESULTS The differences in the sensitivity of two established homozygous knockout strains of P. xylostella. (Rdl1KO and Rdl2KO) and susceptible P. xylostella to a series of compounds were evaluated. Quinazolines and isoxazolines both showed stronger efficacy in the Rdl2KO strain. Therefore, we proposed a method based on the knockdown of the P. xylostella Rdl2 gene as a tactic to enhance the toxicity of quinazolines and isoxazolines. To reduce costs and protect dsRNA against degradation, we applied a bacterial expression system using the L4440 vector to express PxRdl2 dsRNA in HT115 Escherichia coli. Transformed bacteria (dsRNA-Bac) fed through leaves combined with quinazoline and isoxazolines proved to be more effective in both the susceptible and fipronil-resistant P. xylostella.
CONCLUSION Our results provide a strategy for the development of novel insecticide spray formulations containing dsRNA-Bac, which synergize with insecticide toxins by suppressing PxRdl2, reducing the use of pesticides in the field.
1 INTRODUCTION
Since RNA interference (RNAi) was first discovered in the nematode Caenorhabditis elegans1, it has been widely used as a powerful genetic tool for research on various taxa, including plants, insects, nematodes, bacteria, fungi, ect.2,3. RNAi has been considered a new generation of pest control technology because it has great advantages, such as safety for nontarget organisms and the ability to maintain pollution-free conditions. Although dsRNA injected directly into an organism is highly efficient, it is difficult to apply as pest control in the field because of the very high costs of and complexity in preparing dsRNA. Fortunately, ingestion of specific dsRNA through the oral cavity can trigger a silencing response in most body tissues in insects; this paves the way for the development of RNAi-based agricultural pest control if dsRNA can be protected from degradation in the environment and insect intestines by suitable delivery methods.
Currently, wrapping dsRNA in nanomaterials or expressing dsRNA in bacteria are the two main methods of dsRNA-insect delivery. It has been reported that nanomaterials can improve the efficiency of delivery allowing dsRNA to permeate cell membranes directly or by endocytosis4; thus, this has become a new type of delivery agent and has been used in pest RNAi control5. Employing bacteria as delivery vectors of dsRNA molecules is another approach that has been used in recent years and was first carried out in the bacteriophagous nematode C. elegans6. Considering the technology and economic-related issues of dsRNA synthesis in vitro, there have been many reports of bacteria expressing dsRNA to induce insect RNAi to control pests7-9. Other limitations related to the application of dsRNA for pest control include the identification of the appropriate target genes and increasing the insect lethality. For this point, genes targeted by chemical insecticides and bio-insecticides might be ideal sites for dsRNA targeting. It has been reported that silencing Sl102, MvIIS and OfCTP through RNAi increases the toxicity of Bacillus thuringiensis (Bt)10-12.
With the cultivation of cruciferous crops, the diamondback moth (DBM), P. xylostella, has developed into a major pest worldwide13. Following the misuse of insecticides in the field, P. xylostella has developed such severe resistance to most pesticides through multiple mechanisms that it is difficult to control14,15. Although RNAi was reported to be less efficient in lepidoptera16, RNAi in P. xylostella through dsRNA injection or oral administration demonstrated that this approach is for this species17,18. Therefore, the control and resistance management of P. xylostella may be achieved by dsRNA ingestion and interference with specific target genes.
In insects, γ-aminobutyric acid receptors (GABARs) are the major inhibitory receptors widely distributed in the central nervous system19 and are the target of natural compounds (picrotoxin) and synthetic pesticides (such as dieldrin, fipronil, fluxametamide and fluralaner)20-22. According to the classification of the Insecticide Resistance Action Committee (https://irac-online.org/modes-of-action/, IRAC), Compounds that act on GABAR are divided into two categories: blockers (Category 2, represented by fipronil and chlordane) and allosteric modulators (Category 30, represented by broflanilide and fluxametamide) (referred to as GABAR compounds in this paper). Resistance to dieldrin (Rdl) gene, which combines with other GABAR subunits to constitute the target site of cyclodiene and phenylpyrazole pesticides, was cloned from dieldrin-resistant Drosophila melanogaster and named Rdl23. There are two Rdls in P. xylostella24. Our previous research showed that in Xenopus oocytes, the sensitivity of PxRdl2 to fipronil was 40-fold lower than that in P. xylostella Rdl1. In addition, knockout of PxRdl1 reduced the efficacy of fipronil, while knockout of PxRdl2 increased its efficacy25. This may indicate that the PxRdl2 gene plays an important role in resistance to GABAR compounds, which is beneficial for researching toxicology and resistance to insecticides. Based on previous results, we verified whether the knockout of PxRdl2 or PxRdl1 has effects on other GABAR compounds. Furthermore, we considered whether it is possible to develop PxRdl2-based RNAi to improve the efficacy of insecticides.
In this study, two homozygous knockout strains of P. xylostella (Rdl1KO and Rdl2KO) and susceptible strains were established to evaluate the differences in sensitivity of 5 compounds by bioassay. The bioassay results suggested that the newly synthesized quinazolines-4a and the two new isoxazolines (fluxametamide and fluralaner) displayed greater activity in the Rdl2KO strain than in the susceptible strain. Furthermore, PxRdl2 interference by dsRNA, which induced expression by transformed HT115 Escherichia coli (named dsRNA-Bac in this paper), could improve the efficacy of quinazoline and isoxazolines (named RNAi compound in this paper) in both susceptible and fipronil-resistant P. xylostella. In addition, we also verified the effect of UV radiation on dsRNA-Bac to ensure that it could remain stable when exposed to sunlight in field applications. In summary, our study provides new insights for pest control, improving safety for nontarget organisms. Thus, the results may be beneficial for pesticide resistance management and reducing the usage of pesticides in the field, which could be part of an integrated pest management (IPM) approach.
2 MATERIALS AND METHODS
2.1 Insect strain rearing and compounds
Susceptible and fipronil-resistant P. xylostella were generously provided by Dr. Minsheng You (Fujian Agriculture and Forestry University, China). Two homozygous knockout strains (Rdl1KO and Rdl2KO) were previously established from susceptible strains using CRISPR/Cas9 previously25, and were reared on an artificial diet for more than 30 generations in our laboratory. The four strains were kept at 25±1 °C and 65±5% relative humidity under 16:8 h (light: dark) conditions.
Compounds of fipronil (95%), fluxametamide (95%), chloraniliprole (95%), fluralaner (95%) and abamectin (95%) were purchased from TargetMol (Shanghai, China). Compounds 5a26 and 4a27 were synthesized in our laboratory.
2.2 Bioassays
Two bioassays were conducted in this study. To evaluate the sensitivity of the susceptible, RdlKO and fipronil-resistant strains to the compounds, leaf-dip bioassays were carried out as recommended by the IRAC (https://irac-online.org/methods/plutella-xylostella-larvae/). The LCs of the different compounds were determined through the leaf-dip bioassays. To better simulate the outdoor spraying method applied in the field and to detect the impact of gene silencing on the lethality of the compounds, RNAi compounds were subjected to leaf-dip bioassays, in which dsRNA-Bac was evenly applied to the surface of the leaves containing compounds.
For the PxRdl2 RNAi compounds, a sublethal dose (LC20) allowed the assessment of any increase in the mortality rate caused by RNAi-induced gene silencing. Third instar larvae were fed with leaves containing PxRdl2 dsRNA-Bac (or GFP dsRNA-Bac) and compounds. Each treatment used at least 10 larvae with 4 replications. Mortality was recorded every 12 h for 2 days. The theoretically reduced dosage was calculated by the following formula: (LCs – LC20 of the PxRdl2 RNAi compounds)/LCs × 100%. The impact of PxRdl1 dsRNA on the chemical toxicity of P. xylostella was also assessed as described above, with the expectation that the LC50 of each compound dosage was used.
2.3 In vitro synthesis of PxRdls dsRNA
Total RNA was extracted from 3rd instar larvae or bacteria using an RNAsimple Total RNA Kit (TianGen, Beijing, China). The extracted RNA (1 μg) was used to reverse transcribe cDNA using the PrimeScript™ RT reagent Kit with gDNA Eraser (Perfect Real Time) (TaKaRa Biotechnology, Dalian, China).
A 428-bp fragment of PxRdl1 and a 441-bp fragment of PxRdl2 were amplified from cDNA extracted from larvae. The specific fragments were purified using an E.Z.N.A.® Gel Extraction Kit (OMEGA Bio-tek, Guangzhou, China). The PCR products were used as templates for the next PCR and primers containing the T7 polymerase promoter sequence at their 5’ ends were carried out. The PCR products were used as templates to synthesize specific dsRNAs according to the manufacturer’s instructions using the in vitro Transcription T7 Kit (TaKaRa Biotechnology, Dalian, China).
The 289-bp GFP fragment was amplified from the cloning vector pEGFP-N1-ro1GFP (Plasmid #82369, Addgene), and then, a similar method was applied to synthesize specific GFP dsRNA. All the primers in this paper were designed using Primer Premier 5 (Premier, Canada) and are shown in Table 1.
Detailed information of primers used in this study.
2.4 Construction of transformed HT115 Escherichia coli expressing dsRNA and overexpression of dsRNA
The 289-bp GFP, 428-bp PxRdl1 and 441-bp PxRdl2 fragments were cloned into L4440 vectors (Plasmid #1654, Addgene) via homologous recombination28,29. The recombinant L4440 vectors were transformed into HT115 E. coli competent cells. To produce dsRNA, the transformed bacteria were induced by isopropyl β-D-1-thiogalactopyranoside (IPTG). dsRNA was extracted from the bacterial cells for subsequent qRT-PCR absolute quantification.
2.5 qRT-PCR absolute quantification of dsRNA produced by the transformed bacteria
qRT-PCR was carried out using SYBR® Premix Ex Taq™ II (Takara Biotechnology) and a CFX96 Connect Real-Time system (Bio-Rad Laboratories, Inc., Hercules, CA, USA). Based on the absolute quantification method30, the quantity of the dsRNA was confirmed by threshold (CT) values, which relate to an established standard curve. The standard curve for dsRNA was established by plotting the logarithm of a 10-105-fold dilution of the starting solution of template cDNA with inserts against the corresponding CT values.
2.6 Pretreatment of bacteria
Bacteria were collected by centrifugation at 5000×g for 5 min at 4°C and suspended in DEPC-H2O. To inactivate the bacteria and promote the release of dsRNA, sonication treatment was carried out by a low-temperature ultrahigh-pressure cell crusher (Guangzhou Juneng Nano & Biotechnology Co., Ltd, Guangzhou, China) at 1800 bar. LB agar plates containing 100 μg mL-1 ampicillin were used to evaluate whether the bacteria were totally inactivated.
2.7 qRT-PCR relative quantification of PxRdls transcription
dsRNA-Bac (100, 200 or 400 ng μL-1) was evenly applied to the surface of the leaves, and then naturally air-dried. Gene analysis was performed by qRT-PCR, which was carried out by using gene-specific primers, after 2 days of feeding the dsRNA-Bac-covered leaves to 3rd instar P. xylostella larvae. Gene-specific primers were designed to test a segment of the mRNA external to the segment targeted by the dsRNA. The qRT-PCR data were normalized to an internal control (Actin gene, GenBank No. JN410820.1) and analysed by the 2− ΔΔCT method31. Each treatment used 5 larvae with 3 replications. The controls received GFP dsRNA-Bac.
2.8 UV irradiation on dsRNA-Bac
To better simulate the effects of field applications, a UV irradiation experiment was carried out to determine the stability of dsRNA-Bac. PxRdl2 dsRNA-Bac and GFP dsRNA-Bac were placed on the surface of glassware. The glassware was placed on an HD-650 desktop clean bench (Su Jing Purification Equipment, Zhejiang, China). A UV lamp was used to irradiate the glassware for 0 h, 0.5 h, 1 h, 2 h or 4 h (254 nm, 8 W). Before UV irradiation, absolute quantification was carried out on the dsRNA-Bac. The treated dsRNA-Bac was fed to P. xylostella through leaves for gene expression analysis after two days.
2.9 Statistical analysis
For the leaf-dip bioassay data, SPSS 22.0 (SPSS Inc., Chicago, IL, USA) was used to calculate the LCs. The LCs were analysed using regression-probit analysis, and “Log base 10” was selected for transformation. The LC50 values were confirmed to be significantly different if the 95% fiducial limits did not overlap. The other data were analysed using GraphPad Prism, version 5.0 (GraphPad Software Inc., La Jolla, CA, USA). Gene expression was analysed by one-way ANOVA and Tukey’s test (alpha = 0.05). Survival curves of the P. xylostella 3rd instar larvae were compared using log-rank analyses.
3 RESULTS
3.1 Toxicological responses of the two Rdl knockout strains to five compounds
To test roles of the two Rdl homologous genes in the interaction with the five tested compounds, the responses of the two knockout strains and susceptible strain were measured (Table 2 and Fig. 1: data for fipronil were adapted from our previous publication25). The Rdl1KO strain showed greater resistance (2.85-fold) than the susceptible strain to the newly synthesized compound quinazoline (4a). In contrast, the toxicity of 4a to the Rdl2KO strain increased significantly as the resistance ratio (RR) decreased to 0.45. These results revealed that while PxRdl1 and PxRdl2 are both vital determinants of the sensitivity of P. xylostella to 4a, they have adverse effects. Such a tendency was also observed in response to fipronil25 and 5a treatment (unpublished data). The toxicity of fluxametamide to the Rdl1KO and Rdl2KO strains was significantly increased relative to that to the susceptible strain. Unlike fluxametamide, knockout of PxRdl1 had no significant effect on fluralaner susceptibility, but knockout of PxRdl2 increased fluralaner susceptibility (2.50-fold). No difference in toxicity was observed in the three strains for abamectin.
Toxicity of different compounds to susceptible, fipronil-resistant and RdlKO Plutella xylostella.

Resistance levels of the Rdl1KO (PxRdl1 knockout strain) and the Rdl2KO (PxRdl2 knockout strain) to chemical insecticides compared with the susceptible strain of Plutella xylostella. Resistance ratio = LC50 of Rdl1KO or Rdl2KO divided by LC50 of susceptible. * mean their LC50s are significantly different from susceptible.
The negative control, chloraniliprole, exhibited toxicity similar to that of the two Rdl knockout strains compared with that of the susceptible strain. The increased susceptibility to knockout of Rdl2 suggested that it could be an ideal target for RNAi in the development of synergists with GABAR compounds.
3.2 Production of bacteria expressing PxRdls dsRNA
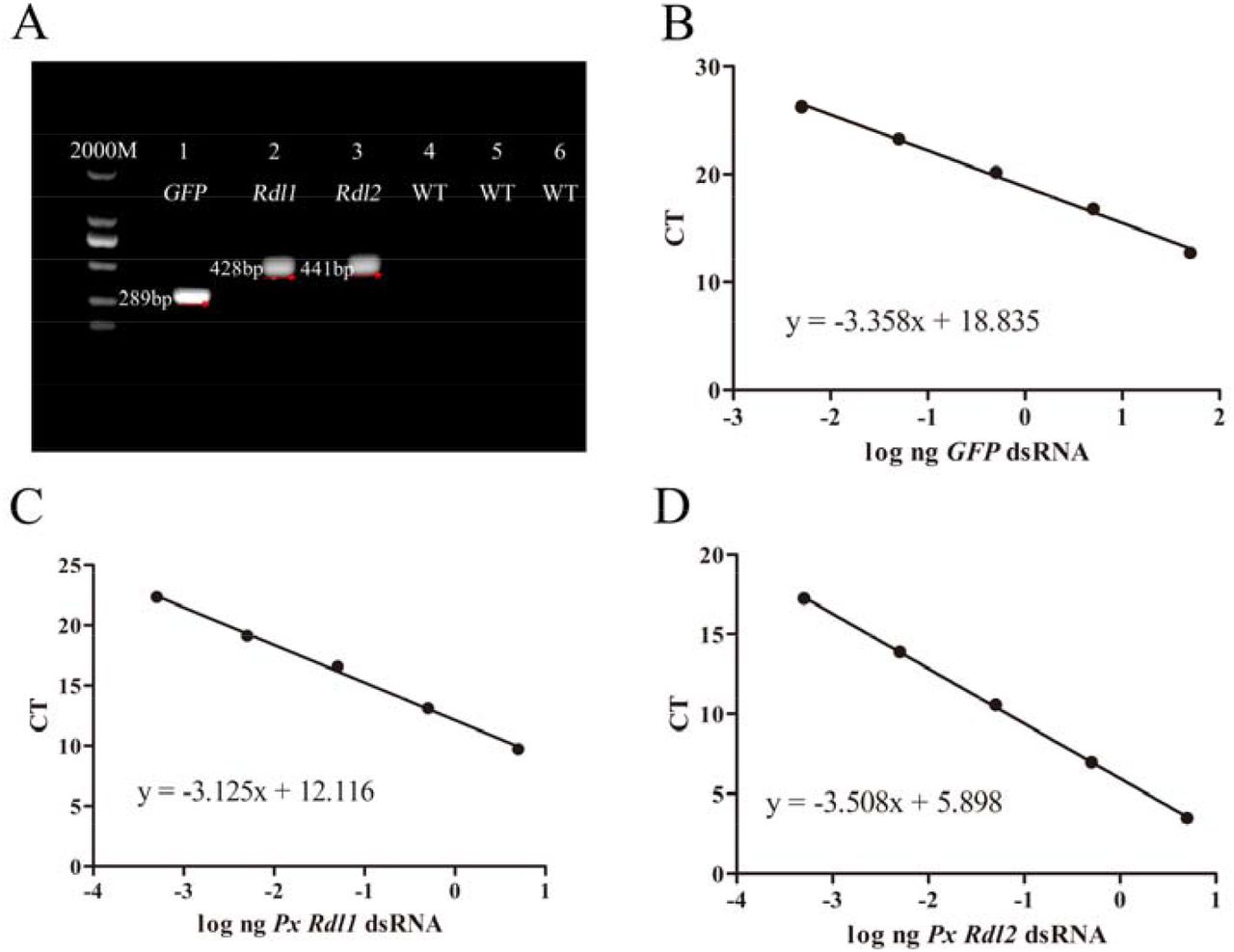
To construct a vector expressing dsRNA, the GFP and PxRdls PCR products were cloned into the L4440 vector, which was induced by the addition of IPTG to overexpress dsRNA under the double T7 promoters and used to transform HT115 E. coli cells. The quantity of dsRNA produced by the HT115 E. coli (Fig. 2a) was quantified by qRT-PCR (Fig. 2b, Fig. 2c, Fig. 2d). For GFP, the PCR efficiency (E = 95.516%) was calculated according to the slope and the coefficient of correlation (R2) of the standard curve (slope = -3.358, y intercept = 18.835, R2 = 0.996). For PxRdl1, the PCR efficiency (E = 108.930%) was calculated according to the slope and the coefficient of correlation (R2) of the standard curve (slope = -3.125, y intercept = 12.116, R2 = 0.998). For PxRdl2, the PCR efficiency (E = 92.780%) was calculated according to the slope and the coefficient of correlation (R2) of the standard curve (slope = -3.508, y intercept = 5.898, R2 = 0.998).

Production of HT115 Escherichia coli cells expressing dsRNA. a Expression of dsRNA by transformed HT115 E. coli; the total RNA were carried out to RT-PCR, and the production of amplification were resolved on 1% agarose gel. The specific primers used for GFP, PxRdl1 or PxRdl2 genes produced amplicons of the expected fragments in transformed HT115 E. coli (lines 1, 2 and 3), whereas the same primers did not produced any fragment from non-transformed HT115 E. coli (WT) (lines 4, 5 and 6). b c and d Standard curves used on qRT-PCR absolute quantification of GFP, PxRdl1 and PxRdl2 dsRNA produced by E. coli suspensions.
3.3 Gene silencing in P. xylostella by feeding individuals leaves containing dsRNA-Bac
Quantitative analysis of the expression of the two PxRdl genes in 3rd instar larvae was performed by qRT-PCR two days after the larvae were fed leaves containing dsRNA produced by HT115 E. coli. The results demonstrated that the transcription levels of the targeted PxRdl1 and PxRdl2 genes were significantly affected by the dsRNA treatment and were positively correlated with the experimental dose used (P < 0.05) (Fig. 3a, Fig. 3b).

mRNA expression of PxRdls gene in Plutella xylostella 3rd instar larvae orally treated for 2 days with dsRNA-Bac. a and b The PxRdl1 and PxRdl2 gene are down-regulated upon ingestion of dsRNA-Bac administered by leaves, which was dose-dependent; c The PxRdl2 dsRNA-Bac treated different UV irradiation times (0h, 1h, 2h, 4h) was fed to P. xylostella 3rd instar larvae for 2 days. UV irradiation for less than 1h had no obvious effect on the activity of dsRNA-Bac. GFP dsRNA-Bac were used as control. The values are the mean ± standard errors (* P < 0.05, Tukey’ s test).
3.4 The effect of UV irradiation on dsRNA-Bac
After dsRNA-Bac was irradiated with UV for different times, the treated dsRNA-Bac was fed to 3rd instar larvae through leaves. The gene expression analysis data showed that dsRNA-Bac had strong stability and could maintain activity under strong UV irradiation for 1 h. The activity of dsRNA-Bac decreased when exposed to UV for more than 2 h (P < 0.05) (Fig. 3c).
3.5 PxRdl2 dsRNA-Bac enhances the toxicity of quinazolines and isoxazolines in susceptible Plutella xylostella
RNAi compounds were used to test the efficacy of the simultaneous administration of PxRdl2 dsRNA-Bac and compounds (fipronil, quinazolines and isoxazolines). This experiment aimed to more closely reproduce the possible effects of the two active ingredients (dsRNA and compounds) applied in the field. The results showed that simultaneous administration of PxRdl2 dsRNA-Bac and compounds led to obviously higher mortality in PxRdl2-silenced larvae than in controls (Fig. 4) (log-rank test for 3rd instar larvae exposed to different compounds: fipronil: chi-square = 4.501, df = 3, P = 0.0339; fluxametamide: chi-square = 14.80, df = 3, P = 0.0001; fluralaner: chi-square = 12.33, df = 3, P = 0.0004; 5a: chi-square = 14.95, df = 3, P = 0.0001; 4a: chi-square = 34.26, df = 3, P < 0.0001).

PxRdl2 RNAi-compounds in susceptible Plutella xylostella 3rd instar larvae Susceptible 3rd instar larvae were treated for 2 days with leaves containing with PxRdl2 dsRNA-Bac (corresponding to 400 ng μL-1 of dsRNA) and with compounds (LC20). a for fipronil, * P = 0.0339; b for fluxametamide, *** P = 0.0001; c for fluralaner *** P = 0.0004; d for 5a *** P = 0.0001; e for 4a *** P < 0.0001 (based on log-rank test).
In addition, to further evaluate the effect of RNAi compound-mediated toxicity enhancement, the theoretical reduction in compound usage was calculated. When the compounds were used alone, the LCsb in Table 3 shows the dose corresponding to mortality, indicating the final mortality of PxRdl2 RNAi compounds after 48 h. According to the calculation results, the enhanced toxicity of the compounds mediated by RNAi could reduce the doses needed of different compounds (fipronil: 45.00%; fluxametamide: 56.82%; fluralaner: 45.45%; 5a: 59.31%; and 4a: 72.36%) (Table 3).
The LCs of 5 compounds in susceptible Plutella xylostella
3.6 PxRdl1 dsRNA-Bac-mediated gene silencing led to reduced toxicity of quinazolines and isoxazolines in susceptible Plutella xylostella
The LC50 values of fipronil and the quinazolines and isoxazolines were used for a PxRdl1 RNAi compound bioassay similar to that for PxRdl2. The data showed that PxRdl1 silencing reduced the toxicity of fipronil and the quinazolines and fluxametamide, except for fluralaner (Fig. 5) (log-rank test of the 3rd instar larvae for different compounds, fipronil: chi-square = 11.11, df = 3, P = 0.0009; fluxametamide: chi-square = 4.672, df = 3, P = 0.0307; fluralaner: chi-square = 0.5698, df = 3, P = 0.4503; 5a: chi-square = 4.159, df = 3, P = 0.0414; 4a: chi-square = 8.844, df = 3, P = 0.0029).

PxRdl1 RNAi-compounds in susceptible Plutella xylostella 3rd instar larvae Susceptible 3rd instar larvae were treated for 2 days with leaves containing with PxRdl1 dsRNA-Bac (corresponding to 400 ng μL-1 of dsRNA) and with compounds (LC50). a for fipronil, *** P = 0.0009; b for fluxametamide, * P = 0.0307; c for fluralaner P = 0.4503; d for 5a * P = 0.0414; e for 4a ** P = 0.0029 (based on log-rank test).
3.7 PxRdl2 dsRNA-Bac enhances the toxicity of quinazoline and isoxazolines in fipronil-resistant Plutella xylostella
Fipronil-resistant P. xylostella are extremely resistant to fipronil, and its resistance has increased 1881-fold. For 4a, the resistance also increased 14-fold. Compound 5a and two isoxazolines had good effects against fipronil-resistant P. xylostella. These 3 compounds were used to the PxRdl2 RNAi compounds. The data showed that these three compounds could increase the toxicity to fipronil-resistant P. xylostella of PxRdl2 RNAi compounds (Fig. 6) (log-rank test of 3rd instar larvae for different compounds, fluxametamide: chi-square = 11.47, df = 3, P = 0.0007; fluralaner: chi-square = 4.964, df = 3, P = 0.0259; 5a: chi-square = 14.65, df = 3, P = 0.0001). In addition, the enhanced toxicity mediated by RNAi could reduce the dose required of different compounds, such as fluxametamide (58.06%), fluralaner (43.14%), and 5a (45.61%) (Table 4).
The LCs of 5 compounds in fipronil-resistant Plutella xylostella

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
PxRdl2 RNAi-compounds in fipronil-resistant Plutella xylostella 3rd instar larvae Fipronil-resistant 3rd instar larvae were treated for 2 days with leaves containing with PxRdl2 dsRNA-Bac (corresponding to 400 ng μL-1 of dsRNA) and with compounds (LC20). a for fluxametamide, *** P = 0.0007; b for fluralaner, * P = 0.0259; c for 5a, *** P = 0.0001 (based on log-rank test).
4 DISCUSSION
Recent studies have shown that the existence of multiple copies of homologous Rdl further enriches the functional library of GABARs in insects24,32,33. These different Rdl subunits may combine to produce GABARs with a series of physiological and pharmacological properties32,33. It has been reported that knockout of Rdl1 or Rdl2 alone does not affect the survival of H. armigera or P. xylostella25,34. These results indicated the functional redundancy of multiple homologous Rdl genes in H. armigera and P. xylostella. We also tried to knockout both Rdl genes in P. xylostella, but unfortunately, no viable homozygous knockouts were obtained (the associated data and process were not shown in this paper). This may further illustrate the functional redundancy of the two Rdls, which cannot be deleted at the same time.
Three strains of the Rdl genotype in P. xylostella were used in bioassays of different compounds to clarify the role of the two Rdls as anti-compounds. Data from our previous bioassays showed that the Rdl1KO strain obtained 10.4-fold resistance to fipronil and that the Rdl2KO strains obtained 4.4-fold sensitivity to fipronil compared to the sensitivity of susceptible strains25. For 4a, knockout of PxRdl1 significantly increased resistance, by 2.85-fold, whereas knockout of PxRdl2 significantly increased susceptibility, by 2.21-fold. 5a is a possible antagonist of GABAR26, our unpublished results also proved that 5a has a similar trend to 4a and fipronil. This suggested that both PxRdl1 and PxRdl2 were involved in mediating fipronil and newly synthesized quinazolines toxicity. Furthermore, PxRdl2 played an important role in anti-quinazolines activities, and PxRdl2 was more tolerant than PxRdl1 to quinazolines. In addition, different pharmacological properties between the two PxRdls may resolve their counter contribution to the sensitivities of P. xylostella to these compounds.
The novel isoxazoline insecticides fluxametamide and fluralaner were also used in the sensitivity tests of the three strains. The toxicity of the two isoxazolines to the three strains showed different trends, which may verify the novel mode of action compared to that of fipronil and two quinazolines. The key mode of action of avermectin in invertebrates is to activate glutamate-gated chloride channels (GluCls), although it has also been proposed that avermectin binds to and activates GABAR35. However, in our study, knockout of PxRdl1 or PxRdl2 did not affect sensitivity to abamectin, confirming that abamectin may not be active on PxRdl1 or PxRdl2 individually. Susceptibility to the negative control, chloraniliprole, was not significantly affected by knockout of either PxRdl1 or PxRdl2. In summary, in pharmacological reactions, our results indicated that PxRdl2 plays an important role in resisting quinazolines and isoxazolines.
Pest control based on RNAi, due to its specificity and minimal or nonexistent impact on nontarget species, provides new opportunities for the development of sustainable IPM plans. Baum et al36 demonstrated that genetically engineered dsRNA expression in maize (Zea mays) could cause larval mortality in the western corn rootworm (WCR), Diabrotica virgifera. This aroused great interest from researchers related to the use of dsRNA produced by transgenic plants as a novel pest control agent. This new type of pest control tool has been widely applied in coleoptera because of its high RNAi efficiency37. Therefore, it should be further studied and expanded to control other important agricultural pests, such as lepidopteran. The reaction efficiency of lepidopteran insects to RNAi is uneven, and the dsRNA delivery mode will affect its interference efficiency. It has been reported that H. armigera and S. littoralis were sensitive to orally delivered dsRNA10,38. Moreover, there were also reports of RNAi in P. xylostella using orally delivered dsRNA39, which could provide us with references to control P. xylostella using RNAi.
In our previous research, PxRdl2 was confirmed to play an important role in the fight against fipronil and the function of PxRdl2 in fighting quinazolines and isoxazolines was verified in this article. This discovery allows the extension of a new pest control method aimed at enhancing the toxicity of insecticides through RNAi-mediated PxRdl2 gene silencing in P. xylostella. However, the development of efficient, safe and economically sustainable RNAi delivery strategies is vital to achieve this goal. In this paper, we probed the utilization of bacteria as potential carriers of dsRNA targeting the PxRdl2 gene and evaluated their effects on quinazolines and isoxazolines for the control of P. xylostella. The recombinant vector L4440 was transformed into HT115 E. coli to induce the expression of a specific dsRNA fragment, which may be the preferred method for the mass production of dsRNA. Expression of dsRNA by bacteria was effective in silencing the target gene PxRdls and showed a quantitative dependence when administering P. xylostella dsRNA-Bac via leaves (Fig. 3a, Fig. 3b). It was previously reported that the dsRNA molecule was protected by the bacterial envelope and was not easily degraded (in both the environment and the insect gut), which could allow dsRNA to exist/release for a longer time38,40. This provides support for the application of RNAi technology in the field, where light (UV) is an important factor affecting the effect of dsRNA. In the laboratory, UV light is an important means for sterilization and digestion of nucleic acids. To better simulate field light factors, UV irradiation was carried out to test the stability of dsRNA-Bac. Compared with in vitro synthesized dsRNA41, our data showed that dsRNA-Bac was resistant to UV exposure and needed to be continuously irradiated with strong UV for more than 2 h obviously to lose part of its activity (Fig. 3c). It was speculated that dsRNA-Bac may be more stable under the protection of the bacterial envelope than dsRNA directly exposed to the external environment. This experiment proved the stability of dsRNA-Bac under light exposure, because UV radiation in the laboratory is several times stronger than the intensity of solar UV radiation in the field, which laid the foundation for the use of dsRNA-Bac for RNAi in the field.
Feeding P. xylostella dsRNA-Bac confirmed that the target gene (PxRdls) was successfully silenced, which prompted us to evaluate the effect of PxRdls gene silencing on insecticidal activity. The data suggested that PxRdl2 gene silencing induced by dsRNA-Bac has a strong synergistic effect with quinazolines and isoxazolines. Significantly, the mortality of larvae increased in the case of simultaneous feeding of compounds and dsRNA-Bac (Fig. 4). In addition, the theoretically reduced doses of the compounds were calculated and ranged from 45.00% to 72.36%. To further verify these results, PxRdl1 dsRNA-Bac was used for RNAi compound treatment. For fipronil and the quinazolines and fluxametamide, the results showed that the influence of PxRdl1 was the opposite of the influence of PxRdl2 (Fig. 5), which further verifies that PxRdl2 is a key gene in the response to the compounds. However, fluralaner did not show the exact opposite results (Fig. 5c), and more research may be required to explore the mechanism driving this response. There was high resistance to fipronil and 4a in the fipronil-resistant P. xylostella (Table 2). Fortunately, 5a and two isoxazolines were very effective against the fipronil-resistant strain. In the subsequent PxRdl2 RNAi compounds, silencing the PxRdl2 gene also increased the toxicity of the three compounds to the fipronil-resistant strain (Fig. 6). The calculated theoretical reduction in the doses of the three compounds was from 43.14% to 58.06% (Table 4).
In summary, oral PxRdl2 dsRNA-Bac combined with GABAR compounds could increase the activity of the compounds against P. xylostella, and these combinations are suitable for both susceptible and resistant strains. Stable dsRNA-Bac can be prepared by bacteria in large quantities, which greatly reduces the associated costs, supporting the development of pesticide sprays. Therefore, a spray may be developed that includes dsRNA-Bac as a synergist of insecticides, which is beneficial for reducing pesticide doses, delaying pest resistance and protecting nontarget organisms.
Competing financial interests
The authors declare no competing financial interests.
ACKNOWLEDGEMENTS
We would like to thank Professor Xiaorong Wang for her support in insect rearing. This work was supported by the National key R&D program of China (2018YFD0200300), the Project of Science and Technology in Guangdong Province (2018A030313188), the Natural Science Foundation of Guangdong Province (2017A030310490) and the Research and Innovation Team of Key Technologies in Modern Agricultural Industry in Guangdong Province (2019KJ130).
REFERENCES




