

Abstract
H-protein, one of the four component proteins (H, T, P and L) of glycine cleavage system (GCS), is generally considered a shuttle protein interacting with the other three GCS-proteins via a lipoyl swinging arm. We report that without P-, T- and L-proteins, lipoylated H-protein (Hlip) enables GCS reactions in both glycine cleavage and synthesis directions in vitro. This apparent catalytic activity is closely related to the cavity on the H-protein surface where the lipoyl arm is attached. Heating or mutation of selected residues in the cavity destroys or reduces the stand-alone activity of Hlip, which can be restored by adding the other three GCS-proteins. Systematic study of the Hlip-catalyzed overall GCS reactions and the individual reaction steps provides a first step towards understanding the stand-alone function of Hlip. The results in this work provide some inspiration for further understanding the mechanism of the GCS and give some interesting implications on the evolution of the GCS.
Significance statement Glycine cleavage system (GCS) plays central roles in C1 and amino acids metabolisms and the biosynthesis of purines and nucleotides. Manipulations of GCS are desired to promote plant growth or to treat serious pathophysiological processes such as aging, obesity and cancers. Reversed GCS reactions form the core of the reductive glycine pathway (rGP), one of the most promising pathway for the assimilation of formate and CO2 in the emerging C1-synthetic biology. H-protein, one of the four GCS component proteins (H, T, P and L) is generally considered a shuttle protein interacting with the other three proteins via a lipoyl swinging arm. Here, we discovered that without P-, T- and L-proteins, H-protein alone can catalyze GCS reactions in both glycine cleavage and synthesis directions in vitro. The surprising catalytic activities are related to a structural region of H-protein which can be manipulated. The results have impacts on engineering GCS to treat related diseases, to improve photorespiration, and to efficiently use C1-carbon for biosynthesis.
Introduction
In the mitochondria of plant and animal cells as well as in the cytosol of many bacteria, the glycine cleavage system (GCS) comprising four (H, T, P and L) proteins catalyzes the reversible decarboxylation and deamination of glycine to yield CO2, NH3 and provide a methylene group for the conversion of tetrahydrofolate (THF) to N5,N10-methylene-tetrahydrofolate (5,10-CH2-THF)1, 2, 3. The overall reaction cycle catalyzed by GCS comprises three steps as illustrated in Figure 1a (hereinafter GCS is used to refer the four-enzyme system regardless of reaction direction). The reaction is first catalyzed by P-protein (glycine decarboxylase; EC 1.4.4.2) to yield CO2 from glycine and methylamine-loaded H-protein (Hint) from the oxidized form (Hox) of the lipoylated H-protein (Hlip). T-protein (aminomethyltransferase; EC 2.1.2.10) then catalyzes the release of NH3 and transfer the methylene group from Hint to THF to form 5,10-CH2-THF, leaving dihydrolipoyl H-protein (Hred). Finally, L-protein (dihydrolipoyl dehydrogenase; EC 1.8.1.4) catalyzes the oxidation of Hred to regenerate Hox in the presence of NAD+. H-protein as a shuttle protein interacts with the other three GCS-proteins via a lipoyl swinging arm and plays central role in the GCS.

Schematic diagrams of the reversible glycine cleavage reaction catalyzed by GCS or stand-alone H-protein (Hlip). (a) The glycine cleavage and synthesis reactions catalyzed by GCS with the complete set of enzymes; (b) Glycine synthesis reactions catalyzed by Hlip alone under the presence of PLP, THF and DTT (DTT as a reductant replacing the functions of FAD and NADH); (c) Glycine cleavage and (d) synthesis reactions catalyzed by Hlip alone under the presence of PLP, THF, FAD and NADH.
The physiological roles of GCS in various organisms have been well studied. In human and most vertebrates, GCS is part of the serine and glycine metabolism pathway. Serine is catalyzed by serine hydroxymethyltransferase (SHMT) to form glycine and 5,10-CH2-THF, and then the product glycine is degraded by GCS. The final product, 5,10-CH2-THF, whose methylene group derived from the β-carbon of serine or the α-carbon of glycine, is one of the few C1 donors in the biosynthesis process, such as the biosynthesis of purine and methionine2. Decrease or loss in the activity of GCS will lead to glycine accumulation in human body, which is linked to glycine encephalopathy4 (also known as nonketotic hyperglycinemia). Relevant studies have shown that most patients with glycine encephalopathy have a P-protein deficiency, and the rest are caused by T-protein or H-protein deficiency5. Moreover, recent studies have shown that glycine metabolism is associated with tumorigenesis, and P-protein as a key factor regulates glycolysis and methylglyoxal production in cancer cells6, 7. In C3 plants, GCS is the key enzymatic system that deals with a large amount of glycine in mitochondria during the photorespiration, and the activity of GCS directly determines the growth rate of plants. Knockout of GCS gene is lethal to plants, which is relevant to impaired one-carbon metabolism8, whereas overexpression of L-protein9 or H-protein10 have been shown to improve photorespiration rates for further increasing biomass yield.
Although GCS in most organisms runs mostly in the direction of glycine cleavage, it catalyzes glycine synthesis in a few anaerobic bacteria such as Clostridium acidiurici11, Eubacterium acidaminophilum12 and Arthrobacter globiformis13, 14. The reversibility of GCS was first discovered in mitochondrial extract of rat liver15, in A. globiformis16 and in cock liver mitochondria17, and most of these studies were already carried out in 1960-1980s. Today, attributing to the reversibility of GCS, GCS gains renewed attention of researchers, because reductive glycine pathway (rGP), with GCS as its key component pathway, is considered to be the most promising synthetic pathway for the assimilation of formate and CO2 to produce pyruvate18, a key precursor that enters the central metabolic pathway for cell growth and biosynthesis. Sánchez-Andrea et al.19 discovered that rGP functions in an anaerobic sulphate-reducing bacterium Desulfovibrio desulfuricans, and stated that it represents the seventh natural CO2 fixation pathway. Recently, rGP has been successfully introduced into E.coli20, 21, 22, 23 for autotrophic growth on formate and CO2. At the same time, part of this pathway was successfully transferred into Saccharomyces cerevisiae24, Cupriavidus necator25 and Clostridium pasteurianum26. However, the flux of rGP is quite low, which limits the growth of microorganism. It has been pointed out that the reaction catalyzed by GCS is the rate-limiting step in rGP21. Therefore, it is particularly important to understand the catalytic mechanism of GCS for increasing the flux of rGP. Substantial progress has been made in understanding the catalytic properties of GCS, and H-protein is so far considered to function merely as a shuttle protein of the cofactor lipoic acid. Lipoic acid is attached by an amide linkage to the conserved lysine residue of H-protein at the 64th position, and the lipoylated H-protein (Hlip) plays a pivotal role acting as a mobile substrate which undergoes a cycle of reductive methylamination, methylamine transfer and electron transfer in the enzymatic cycle of GCS27.
In this work, we discovered that Hlip alone can enable the GCS reaction cycle in both glycine cleavage (Figure 1b) and synthesis directions (Figures 1c and 1d) in the absence of P-, T- and L-proteins. The formation of glycine from C1 compounds in the presence of suitable cofactors was demonstrated by choosing HCHO as the source of α-carbon of glycine. More detailed analyses led to the striking finding that Hlip can apparently “catalyze” all the GCS reaction steps previously believed to be solely catalyzed by P, T and L-proteins, respectively. These findings not only shed new light into the functions of H-protein, but also provide useful hints for engineering H-protein and GCS, either for treating diseases such as hyperglycinemia, for enhancing biomass yield in plants, or for developing synthetic pathways for technical use of C1-carbons. The fact that stand-alone Hlip can catalyze the synthesis of the basic amino acid glycine from inorganic compounds may also have important implications for the evolution of life.
Results
Effects of components of the GCS reaction system on glycine cleavage and synthesis
On the basis of previous studies8, 28, we successfully constructed GCS catalyzed glycine cleavage and synthesis reactions in vitro. Normally, all the four GCS enzymes are included in the reaction system. During kinetic studies, we found that the reactions of glycine cleavage and glycine synthesis can also occur in the absence of certain GCS enzymes and reaction components. This triggered us to systematically examine the effects of missing a certain component or enzyme in the reaction mixture on the reaction rate of both reaction directions. As shown in Table 1, the lack of a certain component or enzyme can cause very different changes of the reaction rate. As expected, the reaction did not occur in the absence of essential substrates (glycine in the cleavage direction and NH4HCO3 in the synthesis direction). The presence of the Hox was also vital, as no reaction was observed in the absence of Hox. However, varied reaction rates (10-76 % of the reference values) were observed when only one of the P-, T- and L-proteins was missing. Compared to the effects of GCS proteins, the missing of substrates and cofactors (THF, PLP, NAD or NADH) showed often stronger effects on the cleavage and synthesis of glycine. In this context, the effects of PLP were surprising: (1) missing of both P-protein and PLP resulted in neither cleavage nor synthesis of glycine; (2) while missing PLP alone resulted in strongly impaired glycine synthesis, it had, however, no negative effect on glycine cleavage. This might be partially explained by the fact that PLP is covalently bound to P-protein29, 30, 31. Therefore, P-protein expressed in E. coli might have PLP covalently bound to it during its expression. The purified P-protein might still have PLP attached to it and can therefore function well in decarboxylation without externally adding PLP. On the contrary, the effect of PLP absence was even worse than the absence of P-protein for glycine synthesis which implied the importance of PLP for the stand-alone catalytic activity of Hlip.
Effects of missing a certain component of the GCS reaction system on the rates of glycine cleavage (determined as HCHO formation from the degradation of 5,10-CH2-THF) and glycine synthesis.
Hlip alone enables glycine synthesis and glycine cleavage reactions
The results in Table 1 suggested that P-protein, T-protein and L-protein are not essential for the functionality of GCS both in glycine cleavage and glycine synthesis directions. This led us to the question if Hlip alone can “catalyze” glycine formation from NH4HCO3 and HCHO, or glycine cleavage in the opposite direction.
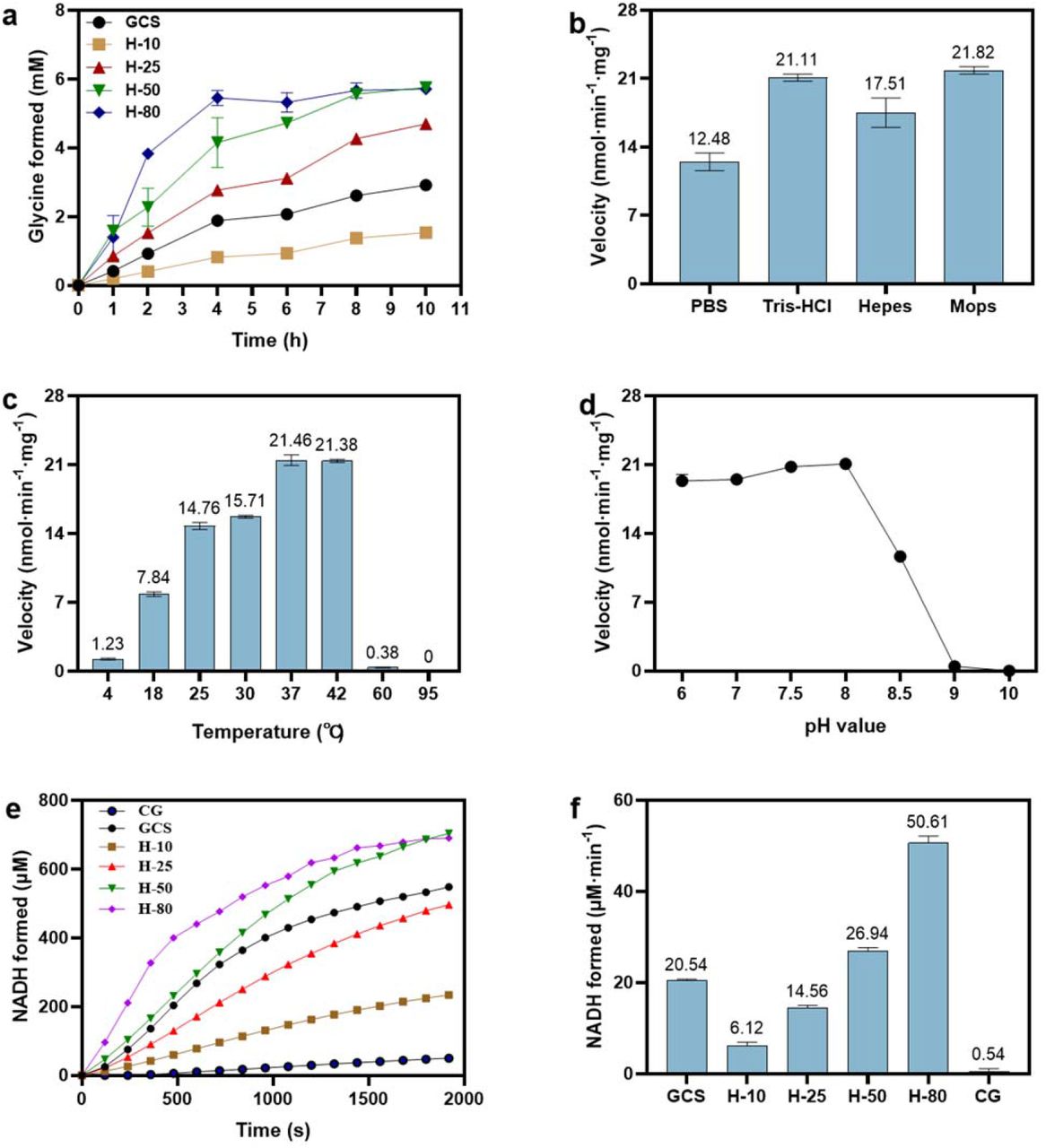
For glycine synthesis, the experimental results with Hlip as the only GCS protein in an array of reaction mixtures are presented in Figure 2a. Compared with the glycine synthesis catalyzed by the all four GCS proteins (GCS as control), the reaction rate catalyzed by Hlip alone at the same concentration of 10 µM (H-10) was somewhat lower, but glycine formation was well detected. With the increase of Hox concentration the initial reaction rate increased and the final concentration of glycine synthesized was higher than that of the control. The above results prove that Hlip alone can apparently catalyze the synthesis of glycine from NH4HCO3 and HCHO in the presence of THF, PLP and DTT. To get the optimum reaction conditions for the glycine synthesis, the reaction rate of glycine synthesis catalyzed by Hlip alone was investigated under conditions of using different buffers, temperatures and pHs. Figure 2b shows the effect of different types of buffer, in which the order of the catalytic ability of Hlip was as follows: Tris-HCl≈Mops > HEPES > PBS. The effect of temperature and pH on the activity were studied by changing temperature from 4 °C to 95 °C (Figure 2c), and pH from 6.0 to 10.0 (Figure 2d). The reaction rate decreased sharply when the temperature was higher than 42 °C or pH was higher than 8.0. The optimum temperature and pH were at 37-42 °C and 7.5-8.0, respectively.

Hlip alone enabled glycine synthesis and glycine cleavage. Effects of Hox concentration (a), buffer (b), temperature (c) and pH (d) on glycine synthesis. “GCS” refers to a reaction mixture for glycine synthesis as specified in “Materials and Methods” without missing any reaction components and enzymes; “H-10”, “H-25”, “H-50” and “H-80” were the same reaction mixture containing no P-, T- and L-proteins but only Hox at 10 µM, 25 µM, 50 µM and 80 µM, respectively. (e) Effects of Hox concentration on glycine cleavage. “CG” refers to no GCS enzymes in the reaction mixture, “GCS” refers to a reaction mixture for glycine cleavage as specified in “Materials and Methods” without missing any reaction components and enzymes; “H-10”, “H-25”, “H-50” and “H-80” were the same reaction mixture containing no P-, T- and L-proteins but only Hox at 10 µM, 25 µM, 50 µM and 80 µM, respectively. (f) NADH formation rate in glycine cleavage catalyzed by different concentrations of Hox.
For glycine cleavage, the reaction could not be observed even at high Hlip concentrations (up to 80 µM) using the same reaction mixture as used for the GCS-catalyzed glycine cleavage reaction but without P-, T- and L-proteins. Later, we found out that when FAD, the coenzyme of L-protein, was added, Hlip alone was indeed able to activate the glycine cleavage, and the reaction rate increased with the increase of Hlip concentration, as shown by the time-courses of NADH formation (Figure 2e) and initial rates of glycine cleavage (Figure 2f). The essentiality of FAD for the glycine cleavage but not for glycine synthesis catalyzed by stand-alone Hlip is due to the presence of DTT which can convert Hox to Hred required in the direction of glycine synthesis (details see below).
H-protein is a small heat-stable protein, so heating does not lead to precipitation. In literature, thermal stability of H-protein is therefore used to terminate the lipoylation of H-protein catalyzed by the enzyme lipoate-protein ligase A (LplA), in which LplA is completely denatured and precipitated 32, 33. We have tried to use heated Hox (at 95 °C for 5 min) to catalyze the reactions of glycine synthesis and cleavage, but no reaction in either direction was observed (the details are discussed in a later section). We therefore speculate that the structure of Hlip was altered by heating at high temperature, which made it lose its catalytic activity shown above for glycine synthesis and cleavage.
In order to explore the reasons behind the function of the stand-alone Hlip observed, we further studied the effect of Hlip alone on the three GCS reaction steps, i.e., the glycine decarboxylation reaction (accompanied by the reductive aminomethylation of Hox to Hint) in the absence of P-protein, the aminomethyl transfer reaction in the absence of T-protein, and the electron transfer reaction without the presence of L-protein, respectively.
Decarboxylation and carboxylation reactions in the absence of P-protein
The results in Table 1 revealed that no activity could be measured for either the cleavage or the synthesis of glycine, when both P-protein and PLP were absent in the reaction mixtures. However, activities were observed when only P-protein was missing. We therefore speculate that the presence of PLP alone might be sufficient to enable Hlip to catalyze the decarboxylation/carboxylation reaction normally catalyzed by P-protein. This was confirmed by the HPLC results shown in Figure 3a for the decarboxylation reaction in the glycine cleavage direction. Hint was formed from Hox without the presence of P-protein. This astonishing result suggests that glycine decarboxylation activated by Hlip alone can occur independent of P-protein, as long as PLP is present (Figure 3a) under the experimental conditions of this study.

Examination of Hlip (Hox) for the function of P-protein. (a) Formation of Hint from Hox during the glycine decarboxylation reaction was determined using HPLC to show that Hlip functions as decarboxylase in the presence of PLP. Chromatograph “With P-protein” refers to a reaction mixture containing 50 mM glycine, 50 µM Hox, 25 µM PLP, and 5 µM P-protein; Chromatograph “Without P-protein” refers to a reaction mixture similar to that “with P-protein” but without the addition of P-protein; Chromatograph “Hox standard” refers to a test solution containing only Hox; (b) PLP-dependent glycine formation was determined to show that carboxylation can take place in the absence of P-protein, albeit at lower reaction rate. “With P-protein” refers to a reaction mixture for glycine synthesis as specified in “Materials and Methods” without missing any reaction components and enzymes, “Without P-protein” refers to a reaction mixture containing all reaction components and enzymes except for P-protein.
For the carboxylation reaction in the glycine synthesis direction, in the absence of P-protein, glycine formation could be still detected and the reaction rate showed a nearly linear increase with the PLP concentration in the low PLP concentration range, albeit that the reaction rate was lower than those determined in the presence of P-protein (Figure 3b). This is understood that CO2 fixation (carboxylation) has a higher energy barrier and P-protein is needed for its activation.
Aminomethyl transfer reaction in the absence of T-protein
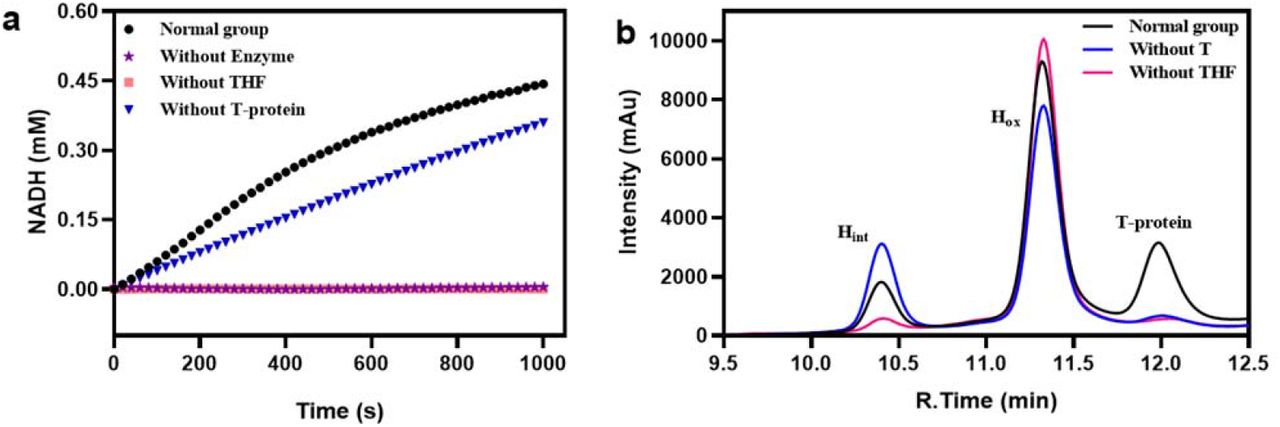
According to the results in Table 1, the overall GCS reaction in both glycine cleavage and glycine synthesis directions could still precede reasonably well in the absence of T-protein in the reaction mixtures. In comparison, it is obvious that the absence of THF had a more significant negative effect on the reaction rate regardless of the reaction directions, i.e. with a reduction of the reaction rate for over 96 % in glycine cleavage and 91 % in glycine synthesis. Figure 4a shows the change of NADH production with time in the direction of glycine cleavage under different conditions. The initial rate without adding T-protein was still more than half of that with adding T-protein, whereas in the experimental group “without THF” the formation of NADH could be hardly detected. For the aminomethyl transfer reaction in the direction of glycine synthesis, Figure 4b shows that Hint could still be generated without adding T-protein in the reaction mixture. It was found by Kochi et al14 that when dihydrolipoic acid, HCHO and NH4+ were mixed together, compounds in the form of -S-CH2NH2 could be obtained. The question then arises: is the aminomethylation of Hred to Hint in the absence of T-protein the result of a complete non-enzymatic reaction due to the presence of HCHO and NH4Cl in which H-protein acts only as the shuttle protein?

Examination of Hlip for the function of T-protein. (a) Effects of THF or T-protein absence on the overall glycine cleavage reaction rate. “Normal group” refers to a reaction mixture containing all reaction components and enzymes required; “Without Enzyme” refers to a reaction mixture containing all reaction components but no GCS enzymes; “Without THF” refers to a reaction mixture containing all reaction components and enzymes except for THF; “Without T-protein” refers to a reaction mixture containing all reaction components and enzymes except for T-protein. (b) Detection of Hint by HPLC proved that the aminomethyl transfer reaction can take place spontaneously. “Normal group” refers to a reaction mixture containing 50 µM Hox, 0.5 mM THF, 20 mM DTT, 50 mM NH4Cl, 10 mM HCHO, and 5 µM T-protein; The reaction mixtures of “Without T” and “Without THF” were the same as that of “Normal group”, but no T-protein or THF was added, respectively.
Interconversion of Hox and Hred in the absence of L-protein
The function of L-protein is to catalyze the interconversion between the Hox and its reduced form dihydrolipoyl H-protein (Hred) involving NAD+/NADH and FAD/FADH2. The experimental results in Table 1 show that in the absence of L-protein both the glycine cleavage and synthesis systems can still work. Previous studies34, 35 have shown that the disulfide bond-reducing agent TCEP allows the reduction of the disulfide bond of the lipoyl group associated with the H-protein during the course of the reaction catalyzed by L-protein. Since a certain amount of DTT was added to prevent the oxidation of THF during the preparation of THF stock solution, we expected that DTT should have the same reduction function as TCEP. Figure 5a schematically shows the interconversion between Hox and Hred through the combination of the reduction of Hox by TCEP (Left) or DTT (Right) with the re-oxidation of Hred catalyzed by L-protein, yielding thereby NADH from added NAD+. The functionality of this combination was demonstrated by the relevant results presented in Figure 5b. Next, we compared the effects of three different reducing agents (TCEP, DTT and β-ME) on the reducibility of Hox to Hred in different buffers (Tris-HCl, PBS, Mops and HEPES) (Figure 5c). It was found that the reducibility of Hox by DTT in four different buffers is PBS > Mops > Tris-HCl > HEPES; TCEP had no reducing effect on Hox in Tris-HCl but showed the strongest reducing power in PBS, even much better than DTT. β-ME was obviously not suitable to reduce Hox in these four buffers. Thus, for glycine synthesis, the functionality of L-protein can be well replaced by DTT or TCEP, as also evidenced by the result shown in Table 1 that the absence of NAD+/NADH did not affect glycine synthesis.

Examination of Hlip for the function of L-protein. (a) Interconversion between Hox and Hred by combining the reduction of Hox to Hred by TCEP (Left) or DTT (Right) with the re-oxidation of Hred to Hox catalyzed by L-protein. (b) Reduction of the lipoamide group of Hox by DTT. “Normal group” refers to a reaction mixture for electron transfer reaction as specified in “Materials and Methods” without missing any reaction components and enzymes. “Without DTT” refers to a reaction mixture similar to “Normal group” but without adding DTT. “Without L-protein” refers to a reaction mixture similar to “Normal group” but without adding L-protein; (c) Comparison of different disulfide reductants on the reduction of Hox in different buffer solutions. (d) Time courses of NADH formation as the result of the redox reaction of Hlip (H) or heated Holip (ΔH) or lipoic acid (LA) in the presence of either L-protein (L) or FAD. (e) NADH formation rates in the redox reaction of Hox, heated Hox and lipoic acid in the presence of either L-protein or FAD.
In order to verify that Hlip has the catalytic function of L-protein, we used FAD, the redox coenzyme of L-protein, instead of L-protein in combination with DTT to observe whether the redox reaction can still occur. To this end, NADH formation as the result of the redox reaction of Hlip (Figure 5a) by either L-protein or FAD was measured. Considering the results shown in Figure 5d and Figure 5e, the following conclusions can be drawn: (1) As long as L-protein is present, the redox reaction of the disulfide bond on the lipoyl group is comparable, with the lipoyl group bound to H-protein (Hlip) showing a slightly higher activity than that not bound (in lipoic acid). (2) In the absence of L-protein, the redox reaction of the lipoyl group bound to H-protein still occurred with the help of FAD, though to a lesser extent, but FAD was not able to replace L-protein in the reoxidation of dihydrolipoic acid, indicating that binding of the lipoyl group on H-protein is the prerequisite for the function of FAD in the absence of L-protein. To find out whether this is simply due to “fixation effect” of lipoyl group bound to H-protein which enables an easier approach of FAD to the lipoyl group, or this is facilitated through an unknown interaction of FAD with H-protein, we also examined to use heat-treated (95 °C for 5 min) Hox, which still had the lipoyl arm linked to it. Interestingly, in the presence of L-protein, there was nearly no difference in the redox reaction of the lipoyl group between the heated Hox and the unheated Hox, however, FAD completely lost its function on the heated Hox, clearly suggesting that an interaction of FAD with H-protein is required for its function in the absence of L-protein, and heating-induced structural change of H-protein destroyed the possibility of such interaction.
Possible reasons of apparent catalytic functions of Hlip in glycine cleavage and glycine synthesis
The above results show that for in vitro GCS reactions, Hlip alone enables both the glycine synthesis and the glycine cleavage without the presence of P-, T-, and L-proteins. It seems that Hlip might functionally replace at least P- and L-proteins and acts as glycine carboxylase and dihydrolipoyl dehydrogenase with the help of PLP and FAD, respectively. It is also suggested by the experiment results shown in Figure 5d and 5e that heated Hox lost the catalytic function of L-protein, obviously because heated Hox cannot interact with FAD for the redox reaction of the lipoyl group bound to H-protein.
We therefore further systematically studied the effects of heating (95 °C for 5 min) on the catalytic activity of Hlip for glycine synthesis, in comparison to unheated Hox as well as to lipoic acid. By either using Hlip alone or combined with other GCS enzymes, the overall reaction rate of glycine synthesis was measured. From the results in Table 2, we can ascertain several interesting observations and conclusions. First, the overall glycine synthesis reaction could be catalyzed by the unheated Hox alone; adding P-protein significantly enhanced glycine synthesis, showing the importance of P-protein; the addition of either T- or L-protein has no positive effect. In fact, the in vitro glycine synthesis could run even better without T- and L-proteins, suggesting that in the presence of Hlip the two individual steps, namely aminomethyl transfer and electron transfer catalyzed by the two proteins respectively, could take place through spontaneous aminomethylation of Hred to Hint in the presence of HCHO and NH4+ and reduction of Hlip to Hred by DTT. Second, compared with the unheated Hox, the heated Hox alone could not catalyze the reaction of glycine synthesis; adding P-protein and L-protein partially revived glycine synthesis to different extents, indicating that heated H-protein lost the catalytic ability but was still functional as the shuttle protein of lipoyl group; the addition of T-protein did not bring any effect. Furthermore, to our surprise, when the heated Hox was added with the other three GCS proteins to form a complete GCS, the rate of glycine synthesis was even higher than the GCS containing the unheated Hox, indicating that heated Hox loses its catalytic function but the heat-induced change is even beneficial for H-protein to exert its role as a lipoyl-carrying protein to work together with the other three GCS proteins. Finally, in the presence of P-, T- and L-proteins, free lipoic acid can act as intermediate substrate to sustain the reversed GCS reaction towards gylcine synthesis under the given experimental conditions, even though to a much lower extent than that observed in the case of lipoyl group bound to H-protein.
Synthesis of glycine using unheated Hox or heated Hox or lipoic acid at the same concentration of 10 µM, either alone or in varied combination with other GCS enzymes
Heating Hlip led to the loss of its catalytic activity regarding glycine synthesis. Although we found no change of the HPLC retention time of Hlip in the reversed-phase HPLC chromatograph after heating, which indicates no obvious change in the overall polarity and size of Hlip, heating may induce structural changes that are vital for the catalytic activity of Hlip. We therefore additionally performed HPLC analysis of Hint and Hox to further determine the catalytic activity of Hlip in the two individual reaction steps normally catalyzed by P-protein and T-protein, respectively. As shown in Figure 6a, in the group of unheated Hox the formation of Hint from Hox clearly demonstrated that Hlip was not only a lipoyl-carring protein but could also replace P-protein in catalyzing the glycine decarboxylation reaction. Hint was not detected in the group of heated Hox, indicating that heated Hox lost the catalytic activity of P-protein. Figure 6b shows the reaction results of aminomethyl transfer from Hred (generated in situ from Hlip) to Hint. The group of unheated Hox clearly exhibited the catalytic activity of T-protein; in the group of heated Hox there was a small amount of Hint formation found, however, this low level of aminomethyl transfer activity was most likely not due to a catalytic activity of T-protein manifested by heated Hox, but due to a spontaneously occurring aminomethyl transfer reaction. Since heated Hox still preserves its function as shuttle protein, the difference between the unheated and heated Hox in the aminomethyl transfer reaction may provide an answer to the question in section 3.4 that in the absence of T-protein, the participation of Hlip in the aminomethyl transfer reaction is not only as shuttle protein.

Effects of heating on the catalytic activity of Hlip as decarboxylase or aminomethyltransferase. (a) Decarboxylation reaction of glycine. “Hox” refers to a reaction mixture containing 50 mM glycine, 50 µM Hox, and 25 µM PLP; “heated Hox” was the same as “Hox” except for using heated Hox. (b) Aminomethyl transfer reaction (started from Hox, which was reduced to Hred by DTT, before Hred is converted to Hint by aminomethyl transfer reaction). “Hox” refers to a reaction mixture containing 50 µM Hox, 0.5 mM THF, 20 mM DTT, 50 mM NH4Cl, and 10 mM HCHO; “heated Hox” was the same as “Hox” except for using heated H-protein.
To explain the above experimental observations we examined the possibility that the catalytic ability of unheated Hox is due to intermolecular interactions of Hlip itself, and such interactions would enable Hlip to catalyze the interconversion of the different forms (Hlip, Hred and Hint) to complete cycle by cycle the electron transfer, aminomethyl transfer, and reductive aminomethylation. In order to verify this hypothesis, we developed a concept to distinguish a possible intermolecular interaction of Hlip from its function as shuttle protein. On the one hand, the lysine residue at position 64 of H-protein for binding lipoic acid was mutated into alanine to obtain HK64A mutant protein, which is unable to be lipoylated and consequently unable to act as shuttle protein, but is still expected to be able to uphold the capability of intermolecular interactions of the wild-type H-protein. On the other hand, as discussed above, heated Hox losing its catalytic activity for glycine synthesis well preserves its function as a shuttle protein. Therefore, if the hypothesis holds true, we would expect that mixing heated Hox with HK64A would “restore” the catalytic activity exhibited by unheated Hox towards glycine synthesis. No glycine formation could be detected which rules out the hypothesis and points out that the interconversion of the three forms of lipoylated H-protein, and consequently the stand-alone catalytic activity of H-protein, is not the result of intermolecular interactions of H-protein itself.
An answer to the question of why heating destroys the catalytic ability of H-protein but still preserves its function as shuttle protein might lie in the special surface structure of H-protein. As shown by the crystal structure of Hint (Figure 7a), following methylamine transfer the lipoamide-methylamine arm enters into the hydrophobic cavity on the surface of the H-protein and prevents thereby it from nucleophilic attack by water molecules28, 36, 37, 38. Based on our previously molecular dynamic simulation study, the lipoamide-methylamine arm may interact with some amino acid residues in the proximity of the cavity39, such as Glu-12, Glu-14, Ser-67, Cys-68 and Tyr-70 (Figure 7b). We speculated that the catalytic activity found for the stand-alone Hlip is related to the structure of the cavity. Therefore, to verify this assumption, the above five residues of the wild-type H-protein were mutated to alanine. Assays of glycine synthesis were then performed with these mutated Hlip in comparison with the wild-type Hlip. As shown in Figure 7c, compared to the wild-type Hlip the glycine synthesis rates were strongly reduced in reaction mixtures containing Hlip mutants. However, as shown in Figure 7d, when these mutants were combined with the other three GCS proteins, the glycine synthesis rates of all the mutants were increased to levels comparable or even better than that of the wild-type Hlip. The results are very similar to what observed with heated Hox (Table 2). It is therefore confirmed that the cavity on the H-protein surface plays a decisive role in the catalytic functions of H-protein, and alterations of the cavity structure (in size or form) either through mutation or heating will reduce or even destroy the stand-alone catalytic functions of H-protein (Figure 7c, Table 2) due to yet unclear mechanisms. One possible reason might be that the lipoamide-methylamine arm cannot properly enter the deformed cavity, resulting in the failure or imbalance of the GCS cycle in the presence of heated Hox alone. By adding other GCS proteins, H-protein is mainly required to act as a shuttle protein and, consequently, GCS reactions can be revived (Table 2). Moreover, under the given in vitro reaction conditions, the lipoamide-methylamine arm may undergo fast and continuous reaction in the GCS cycle that not only minimizes the probability of its hydrolysis, but also maintains or even increases the reaction efficiency by omitting the process of its entry and exit from the cavity (Figure 7d, Table 2).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Study of the essential roles of the H-protein cavity. (a) Three-dimensional structure of E. coli H-protein bearing the lipoamide-methylamine arm protected in the cavity (ecHint). (b) Docking of the lipoamide-methylamine arm in the cavity with highlighting of some surrounding amino acid residues selected for mutation. (c) The rate of glycine synthesis catalyzed by stand-alone Hlip or its mutants. (d) The rate of glycine synthesis catalyzed by GCS comprising Hlip or its mutants and the other three GCS proteins.
Discussion
In this work, we show for the first time that stand-alone lipoylated H-protein (Hlip) has the catalytic functions so far believed to be carried out by the P-, T- and L-proteins of GCS. It enables glycine cleavage reactions, as well as the reversed reactions towards glycine synthesis with NH4HCO3 and HCHO as the substrates. The Kcat value for the overall synthesis reaction is about 0.01 s−1 for GCS catalyzed reaction and 0.0057 s−1 for H-protein alone catalyzed reaction. After purification of the GCS proteins, we used the most commonly used methods SDS-PAGE and HPLC to verify that there was no obvious residual of other GCS proteins in the purified H-protein solutions, though we did not confirm this by using more precise methods like mass spectroscopy. Through calculations, we can state that even if other GCS proteins would exist in the H-protein solution (e.g. up to 10%), it will not qualitatively affect the main conclusions drawn in our work (see Supplementary Materials for detailed explanation). Therefore, the purity of H-protein meets the requirement needed for this study.
The stand-alone catalytic activity of Hlip is closely related to the cavity on the H-protein surface, where the lipoyl swing arm is bound. Both heating Hlip and mutating cavity-related amino acid residues result in complete loss or strong reduction of the stand-alone catalytic activity of Hlip, because they may cause deformation of the cavity, resulting in failure of the lipoamide-methylamine arm to properly enter the cavity and consequently failure of GCS reactions. Cohen-Addad et al.36 suggested that the lipoamide-methylamine arm is locked into a very stable configuration within the hydrophobic cavity and therefore highly stable against the non-enzymatic hydrolysis (which leads to the release NH3 and HCHO) due to nucleophilic attack by water molecules.
However, our experiments surprisingly show that when heated Hox or a Hox mutant was combined with the other GCS proteins, the rate of glycine synthesis was recovered or even increased (Table 2 and Figure 7). This implies that under the given in vitro reaction conditions, heated Hox truly acts only as shuttle protein in the presence of P-, T- and L-proteins, it is unnecessary and even disadvantageous for the lipoamide-methylamine arm to enter the cavity in order to undergo GCS reactions. Indeed, there was no obvious decrease of the peak area of Hint on HPLC even after hours of waiting, indicating that the hydrolysis of Hint is very slow (data not shown). This is in consistence with our recent finding that mutations of the key residue Ser-67 which reduce the residence time of the lipoamide-methylamine arm in the cavity can significantly increase the in vitro GCS activity39. The molecular dynamic simulations of H-protein carried out by our group39 also implies that the lipoamide-methylamine arm can leave the cavity of unheated H-protein even without the interaction with T-protein. This is confirmed by further molecular dynamic simulations of H-protein (results not published yet). These results therefore raise the question why the lipoamide-methylamine arm should enter the cavity and be protected in the GCS system? Based on the fact that the cavity is closely related to the stand-alone catalytic activity of H-protein, perhaps we may make a bold speculation for the interpretation of these results from a perspective of evolution. In vivo, GCS may have evolved from a simple system for glycine cleavage catalyzed by H-protein alone at the early time of evolution to a sophisticated system, in which H-protein is assisted by specialized P-, T-, L-proteins for more effective catalysis of the GCS reactions to meet the growing metabolic requirements of organisms. However, the cavity structure and the stand-alone catalytic functions of H-protein have been retained till now.
Oliver40 discovered that the H-protein and the small subunit of ribulose-1,5-bisphosphat-carboxylase/-oxygenase (RuBisCo) have obvious similarities in plants. The two proteins are not only about the same size, but also have similar mechanism in terms of transcriptional control of the corresponding genes. It has been reported that there are striking sequence and structure similarity between H-protein and the E2 protein of pyruvate dehydrogenase complex (PDC)27. Therefore, further research on the catalytic mechanism of H-protein may give useful hints for understanding the evolution and function of PDC and other 2-oxoacid dehydrogenase multi-enzyme complexes (e.g. alpha-ketoglutarate dehydrogenase complex) which are all of fundamental importance in cellular metabolism, governing the synthesis of C1-C4 metabolites for life.
Hlip alone catalyzes the glycine cleavage and synthesis in vitro with the help of the cofactors PLP, THF and FAD. This seems in contradictory to the results reported in the previous literature that deletion in gcvP or gcvT is lethal for organisms8, 41. By comparing the differences between the in vivo and in vitro conditions, it is conceivable that in vivo these cofactors are stoichiometrically linked to P-, T and L-proteins, respectively, to play their dedicated catalytic roles in a concerted action with H-protein. The in vivo concentrations of these cofactors are much lower than what we used in our in vitro experiments to facilitate the cleavage or synthesis of glycine catalyzed by H-protein alone. Thus, at one hand, it may not be feasible for cells to maintain high concentration pools of these cofactors; at the other hand, it might be beneficial for cells to have the sophisticated complete GCS system for fast and fine tuning and adapting of this important system to the changes in metabolic demands. This may be one reason why P-, T-, L-proteins are necessary in vivo. This study shows that for the stand-alone catalytic functions of Hlip an interaction of FAD with Hlip is required for its function in the absence of L-protein, and heating-induced structural change of Hlip destroyed the possibility of such interaction (Figure 5d). It is most likely that PLP also needs to interact with Hlip to exert its function in the absence of P-protein. This thus raises questions: How do PLP and FAD interact with Hlip? Is it similar to the binding of PLP to P-protein and FAD to L-protein? To answer these questions, more experiments will be carried out in the future studies.
In addition, the overall reaction rate of GCS with T-protein deficiency was only reduced to 52 % in the direction of glycine cleavage and 76.5 % for glycine synthesis, compared to those of the complete GCS, indicating that T-protein had the least effect on the catalytic activities of GCS. These results are in agreement with the results of Timm et al.8 who showed that knockdown mutants of Arabidopsis containing very low T-protein expression under physiological conditions were able to grow and propagate in normal air, only showing some minor changes. Meanwhile, their study also found that the knockout mutation without T-protein expression was lethal even in non-photorespiratory environment of air enriched to 1 % CO2. This result may indicate that THF needs weak binding with T-protein to participate in glycine cleavage and synthesis reaction in vivo. When the T-protein is completely knocked out, THF cannot participate in the reaction, resulting in slow reaction rate and plant death. The deficiency of THF is far more detrimental and has the greatest impact on the reaction rate of GCS in vitro (Table 1).
Although we have identified the cavity of the H-protein as the key structural region that determines the catalytic activities of stand-alone Hlip, the specific catalytic mechanism is not explored from the perspective of structure and molecular interaction. By studying the structures and their dynamics of heated and unheated H-proteins, it will certainly help to better understand the mechanism. This in turn will help to better engineer H-protein and GCS, leading to new possibilities to improve the growth and physiology of cells, organisms and plants and to design industrial microbes for utilizing C1 compounds for biosynthesis.
Materials and Methods
Materials
The substrates glycine, nicotinamide adenine dinucleotide (NAD+, NADH), Tris (2-carboxyethyl) phosphine (TCEP) and the derivatization reagents dansyl chloride were purchased from Yuanye Bio-Technology (Shanghai, China). Dithiothreitol (DTT), β-mercaptoethanol (β-ME), pyridoxal 5’-phosphate monohydrate (PLP) and flavin adenine dinuleotide (FAD) were obtained from Aladdin (Shanghai, China). 6-(RS)-Tetrahydrofolate (THF) was obtained from Sigma-Aldrich (St. Louis, MO, USA). Other chemicals in this study were of analytical grade and purchased from Solarbio (Beijing, China) or Sinopharm (Shanghai, China), unless otherwise noted. Escherichia coli Top10 and BL21 (DE3) were used for plasmid construction and the overexpression of recombinant proteins, respectively. Ni2+-NTA resin was purchased from Genscript (Nanjing, China). Amicon® Ultra-15 filtration devices (molecular size cut-off 10 KDa for H-protein, 30 KDa for T-protein and L-protein and 100 KDa for P-protein) were purchased from Millipore (Billerica, MA, USA). Mut Express II Fast Mutagenesis kit V2 was purchased from Vazyme (Jiangsu, China). BCA protein assay kit and sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) gels were purchased from SolarBio (Beijing, China). Luria-Bertani (LB) medium containing tryptone (10 g/L), yeast extract (5 g/L) and NaCl (10 g/L) were used for cloning and expression, and tryptone and yeast extract were purchased from Oxoid.
Enzyme preparation
The plasmids and bacterial strains used in this experiment were given in Table 3. Oligonucleotide sequences of primers used for cloning target proteins were given in Table 1 of Supplementary Materials. The genes coding for P-protein, T-protein, L-protein and H-protein were amplified from E. coli K12 genomic DNA, then cloned into the expression vector pET28a (NdeI and XhoI). E.coli BL21 (DE3) harboring the resulting constructs (pET28a-P, pET28a-T, pET28a-L and pET28a-H) were cultured in LB medium supplemented with 50 mg/L of kanamycin at 37°C until the OD600 of the culture reached 0.6-0.8, Isopropyl-beta-D-thiogalactopyranoside (IPTG) was added to a final concentration of 0.2 mM to induce protein expression for 12 h at 30 °C.
The plasmid pET28a-H was used as a template to generate mutations using Mut Express II Fast Mutagenesis kit V239. Lipoylation of H-protein was performed during its over-expression in vivo. To this end, the strain containing the plasmid pET28a-H or a H-protein mutant were added with lipoic acid (200 μM, pH 7.0), prior to starting the cultivation to directly obtain lipoylated H-protein (Hlip). Following the overexpression, enzymes were purified as described previously42. The purified enzymes were checked by SDS-PAGE (shown as Figure S1 in Supplementary Materials). In the lane of the H protein no residuals of T, P and L proteins were found except for the existence of low amount of the inactive H apo-protein (Hapo). In addition, HPLC analysis also confirmed that there was no obvious residual of other GCS proteins in the purified H-protein solutions. Hlip obtained in such a way is considered to primarily exist in the oxidized form of H-protein (Hox). Whereas Hlip is mostly used to refer to lipoylated H-protein in general, it is numerically equivalent to Hox in this work when concrete reactions involving Hlip are referred to and vice versa.
Enzyme activity assays
The overall reaction of glycine cleavage
(1) Glycine cleavage catalyzed by GCS
The reaction mixture contained Tris-HCl (50 mM, pH 7.5), 0.5 mM THF, 20 mM DTT, 25 μM PLP, 5 mM NAD+, 5 µM P-protein, 5 µM T-protein, 5 µM L-protein and 10 µM Hox. After premixing and centrifugation, the reactions were initiated by the addition of 50 mM glycine, and carried out for 30 min at 37 °C. The overall GCS activity was determined by measuring either NADH formation at 340 nm using an Enspire multimode plate reader (PerkinElmer, USA) or formaldehyde formation according to our previous reported method17.
(2) Glycine cleavage enabled by Hlip alone
The glycine cleavage reaction enabled by Hlip alone was monitored by determining the formation of NADH at 340 nm. The reaction mixture contained Tris-HCl (50 mM, pH 7.5), 0.5 mM THF, 20 mM DTT, 25 μM PLP, 5 mM NAD+, 40 μM FAD and 10 µM Hox. The reaction was initiated by adding 50 mM glycine.
The overall reaction of glycine synthesis
(1) Glycine synthesis catalyzed by GCS
The reaction mixture containing Tris-HCl (50 mM, pH 7.5), 0.5 mM THF, 20 mM DTT, 10 mM HCHO, 25 μM PLP, 5 mM NADH, 5 µM P-protein, 5 µM T-protein, 5 µM L-protein and 10 µM Hox. The reaction was initiated by adding 50 mM NH4HCO3 to the reaction mixture and carried out for 2 h at 37 °C. The amount of glycine formed was determined using HPLC. One unit of glycine synthesis activity was defined as the amount (in mg) of H-protein that catalyzed the formation of 1 nmol of glycine per minute.
(2) Glycine synthesis enabled by Hlip alone
The reaction mixture contained Tris-HCl (50 mM, pH 7.5), 0.5 mM THF (added with β-ME to prevent its oxidative degradation), 10 mM HCHO, 25 μM PLP, 50 mM NH4HCO3, 40 μM FAD and 5 mM NADH, and different concentration of Hox (10-80 μM). Alternatively, 20 mM DTT can be used to replace FAD and NADH for the reduction of Hox to Hred. The reaction condition and enzyme activity calculation were the same as stated above in 2.3.2 (1).
Individual GCS reaction steps in the presence of Hlip with or without the corresponding enzymes
(1) Glycine dcarboxylation reaction catalyzed by P-protein
The reaction mixture contained Tris-HCl (50 mM, pH 7.5), 50 mM glycine, 50 µM Hox and 25 µM PLP. 5 µM P-protein was added to the reaction mixture as the control group. The reaction was carried out for 2 h at 37 °C. The substrate Hox and the product Hint were measured using HPLC.
(2) Aminomethyl transfer reaction catalyzed by T-protein
In this reaction of converting Hred to Hint through aminomethyl transfer, Hred required was generated by reducing Hox with DTT (see 2.3.3 below), and 5,10-CH2-THF was derives from the condensation of HCHO and THF. Therefore, the reaction mixture contained Tris-HCl (50 mM, pH 7.5), 50 µM Hox, 0.5 mM THF, 20 mM DTT, 50 mM NH4Cl, and 10 mM HCHO. 5 µM T-protein was added to the reaction mixture as the control group. The reaction was carried out for 2 h at 37 °C. The substrate Hox and the product Hint were measured by HPLC.
Electron transfer reaction between Hox and Hred with or without the presence of L-protein
The interconversion of Hox and Hred was performed according to a reported enzymatic assay using an excess amount of a reductant (8 mM TCEP or 20 mM DTT) for the reduction of the H-protein-bound lipoic acid34, 35, and then the produced Hred is re-oxidized by L-protein in the presence of NAD+. For the assay, the reaction mixture contained different types of buffer (50 mM, pH 7.5), 8 mM TCEP or DTT, 5 µM Hox and 0.2 µM L-protein. In order to prove that Hox can still undergo redox reaction without L-protein, 40 µM FAD is used instead of L-protein. The reactions were initiated by the addition of 5 mM NAD+. The rate of NADH formation was determined spectrophotometrically at 340 nm.
Analytical methods using HPLC
Hox and Hint proteins were analyzed based on the HPLC method previously developed in our lab33. The analysis was performed with a Inertsil WP300 C4 column (5 μm, 4.6×150 mm) and monitored at 280 nm using a diode array detector (DAD).The mobile phase consisted of acetonitrile (A) and 0.1 % trifluoroacetic acid aqueous solution (B). The volumn percentage of buffer B was varied as follows: linearly increased from 30 % to 50 % (0-13.4 min), sharply increased from 50 % to 90 % (13.4-13.41 min), held at 90 % (13.41-14.2 min), and then sharply decreased to 30 % (14.2-14.21 min), held at 30 % to 18 min. The flow rate was 1.0 mL·min−1.
Glycine concentration in the reaction mixture was determined by pre-column dansyl chloride derivatization. To this end, 40 µL of a reaction mixture was mixed with 160 µL of 0.2 M NaHCO3 and 200 µL of 5.4 mg · mL−1 dansyl chloride in acetonitrile. Derivatization occurred at 30 °C for 30 min. After the reaction, 600 µL of 0.12 M HCl was added to adjust the pH of the sample to weak acidic. After centrifuged at 10,000 rpm the supernatant was filtered with 0.22 µm membrane. The dansyl derivative of glycine was measured using HPLC (Shimadzu LC-2030C system) on a Shim-pack GIST C18 column (5 μm, 4.6×150 mm) at 30 °C, with a mobile phase composed of acetonitrile and 20 mM potassium phosphate buffer pH 6.0 (25:75 v/v) at a flow rate of 0.8 mL/min. The effluent was monitored at 254 nm using a diode array detector (DAD). The HPLC results of glycine were given in Supplementary materials Figure 2.
Statistics and Reproducibility
Enzyme activities and reaction rates were measured by three independent experiments and averaged for report. Individual data points are added in the graphs, and error bars are defined by the standard deviation.
Data availability
Major data generated and analyzed during this study are included in the article. The source data underlying the graphs and charts presented in the main figures are available as Supplementary Data. Other datasets generated and analyzed during the study are available from the corresponding author on reasonable request.
Author Contributions
Y.X. designed and performed the experiments, wrote the initial manuscript. Y.L. assisted in experiments and data analysis. H.Z. provided H-protein mutants and participated in data analysis. J.N. assisted in experiments and preparing the figures. J.R. involved in experimental design, data analysis and drafting the manuscript. W. W. involved in data analysis and revised most of the manuscript content. A.-P.Z. supervised the project, involved in experimental design, data analysis and discussion, reviewed and revised the paper.
Acknowledgments and funding
This work was financially supported by the Beijing Advanced Innovation Center for Soft Matter Science and Engineering, Beijing University of Chemical Technology.
Reference




