

ABSTRACT
The functional human immunodeficiency virus (HIV-1) envelope glycoprotein (Env) trimer [(gp120/gp41)3] is produced by cleavage of a conformationally flexible gp160 precursor. Gp160 cleavage or the binding of BMS-806, an entry inhibitor, stabilizes the pre-triggered, “closed” (State-1) conformation recognized by rarely elicited broadly neutralizing antibodies. Poorly neutralizing antibodies (pNAbs) elicited at high titers during natural infection recognize more “open” Env conformations (States 2 and 3) induced by binding the receptor, CD4. We found that BMS-806 treatment and crosslinking decreased the exposure of pNAb epitopes on cell-surface gp160; however, after detergent solubilization, crosslinked and BMS-806-treated gp160 sampled non-State-1 conformations that could be recognized by pNAbs. Cryo-electron microscopy of the purified BMS-806-bound gp160 revealed two hitherto unknown asymmetric trimer conformations, providing insights into the allosteric coupling between trimer opening and structural variation in the gp41 HR1N region. The individual protomer structures in the asymmetric gp160 trimers resemble those of other genetically modified or antibody-bound cleaved HIV-1 Env trimers, which have been suggested to assume State-2-like conformations. Asymmetry of the uncleaved Env potentially exposes surfaces of the trimer to pNAbs. To evaluate the effect of stabilizing a State-1-like conformation of the membrane Env precursor, we treated cells expressing wild-type HIV-1 Env with BMS-806. BMS-806 treatment decreased both gp160 cleavage and the addition of complex glycans, implying that gp160 conformational flexibility contributes to the efficiency of these processes. Selective pressure to maintain flexibility in the precursor of functional Env allows the uncleaved Env to sample asymmetric conformations that potentially skew host antibody responses toward pNAbs.
IMPORTANCE The envelope glycoprotein (Env) trimers on the surface of human immunodeficiency virus (HIV-1) mediate the entry of the virus into host cells and serve as targets for neutralizing antibodies. The functional Env trimer is produced by cleavage of the gp160 precursor in the infected cell. We found that the HIV-1 Env precursor is highly plastic, allowing it to assume different asymmetric shapes. This conformational plasticity is potentially important for Env cleavage and proper modification by sugars. Having a flexible, asymmetric Env precursor that can misdirect host antibody responses without compromising virus infectivity would be an advantage to a persistent virus like HIV-1.
INTRODUCTION
Human immunodeficiency virus (HIV-1), the etiologic agent of acquired immunodeficiency syndrome (AIDS), utilizes a metastable envelope glycoprotein (Env) trimer to engage host receptors and enter target cells (1). The functional Env trimer consists of three gp120 exterior subunits and three gp41 transmembrane subunits (1–3). During virus entry, gp120 engages the receptors, CD4 and CCR5/CXCR4, and gp41 fuses the viral and cell membranes (4–16). Env is the only virus-specific protein on the viral surface and is targeted by host antibodies (17–20).
In infected cells, the HIV-1 Env trimer is synthesized in the rough endoplasmic reticulum (ER), where signal peptide cleavage, folding, trimerization and the addition of high-mannose glycans take place (21–24). The resulting gp160 Env precursor is transported to the Golgi apparatus, where some of the glycans are modified to complex types and proteolytic cleavage by host furin-like proteases produces the gp120 and gp41 subunits (25–41). The proteolytically processed, mature Env trimers are transported to the cell surface and incorporated into virions.
On the membrane of primary HIV-1, Env exists in a pre-triggered, “closed” conformation (State 1) that resists the binding of commonly elicited antibodies (42–47). Binding to the receptor, CD4, on the target cell releases the restraints that maintain Env in State 1, allowing transitions through a default intermediate conformation (State 2) to the pre-hairpin intermediate (State 3) (42,48,49). In the more “open” State-3 Env conformation, a trimeric coiled coil composed of the gp41 heptad repeat (HR1) region is formed and exposed, as is the gp120 binding site for the second receptor, either CCR5 or CXCR4 (50–58). Binding to these chemokine receptors is thought to promote the insertion of the hydrophobic gp41 fusion peptide into the target cell membrane and the formation of a highly stable six-helix bundle, which mediates viral-cell membrane fusion (14-16,59-62).
The ability of HIV-1 to establish persistent infections in humans requires an Env trimer that minimally elicits neutralizing antibodies and resists the binding of antibodies generated during the course of natural infection. In addition to a heavy glycan shield and surface variability, the conformational flexibility and plasticity of Env may help HIV-1 avoid the host antibody response (45,47,63–66). Flexible Envs could present epitopes that are not exposed on the State-1 Env trimer, misdirecting host antibodies away from the functional virus spike. The vast majority of antibodies elicited by Env during natural HIV-1 infection are unable to bind the functional State-1 Env trimer, and instead recognize downstream conformations (States 2, 2A and 3) (67–71). These antibodies cannot access their epitopes once the virus has bound CD4 and therefore do not neutralize efficiently (70). Uncleaved Envs that assume State-2/3 conformations are abundant on the surface of HIV-1-infected cells, in some cases reaching the cell surface by bypassing the Golgi (72). Poorly neutralizing antibodies (pNAbs) with State-2/3 specificity typically recognize these uncleaved Envs more efficiently than cleaved Env (73–79). Crosslinking the uncleaved cell-surface Env exerted effects on Env antigenicity similar to those resulting from gp120-gp41 cleavage, suggesting that the uncleaved Env might be more flexible than mature Env (80). Indeed, recent single-molecule fluorescence resonance energy transfer (smFRET) analysis confirmed that, in contrast to the dominant State-1 conformation of the wild-type Env, an Env mutant unable to be proteolytically processed due to an alteration of the cleavage site occupies States 2 and 3 more frequently than State 1 (81). Thus, the abundant, cell-surface-accessible and conformationally heterogeneous uncleaved Env could misdirect host immune responses away from the elicitation of broadly neutralizing antibodies, which generally recognize the State-1 Env conformation (42,45,46,48,81). Broadly neutralizing antibodies (bNAbs) typically appear after several years of HIV-1 infection and only in a minority of HIV-1-infected individuals (83–91).
Here, we investigate the conformation of the uncleaved HIV-1 Env trimer, both on the cell surface and purified from membranes. Cryo-electron microscopy (cryo-EM) reconstructions reveal that purified uncleaved Envs preferentially assume asymmetric trimer conformations, exposing epitopes for pNAbs. We identified a gp41 region in which structural changes are coupled to the asymmetric opening of the Env trimer. We tested the effect of a State-1-stabilizing gp120 ligand, BMS-378806 (herein called BMS-806) on the cleavage and glycosylation of the wild-type Env. Our findings indicate the importance of conformational plasticity of the uncleaved HIV-1 Env trimer for efficient proteolytic maturation, complex glycan addition and evasion of host antibody responses.
RESULTS
Analysis of the conformation of uncleaved HIV-1 Env on cell surfaces
Cleavage of the HIV-1 Env precursor affects its antigenicity (73–79). The recognition of the uncleaved and mature HIV-1JR-FL Envs on the surface of transfected HOS cells exhibited distinct patterns for State 1-preferring bNAbs versus State 2/3-preferring pNAbs (Fig. 1A). Whereas the uncleaved Env was bound by antibodies capable of recognizing all three states, the mature Env was bound only by the potently neutralizing antibodies with State-1 preferences. The uncleaved Env apparently samples multiple conformations, but the mature Env assumes a conformation that precludes the binding of pNAbs.
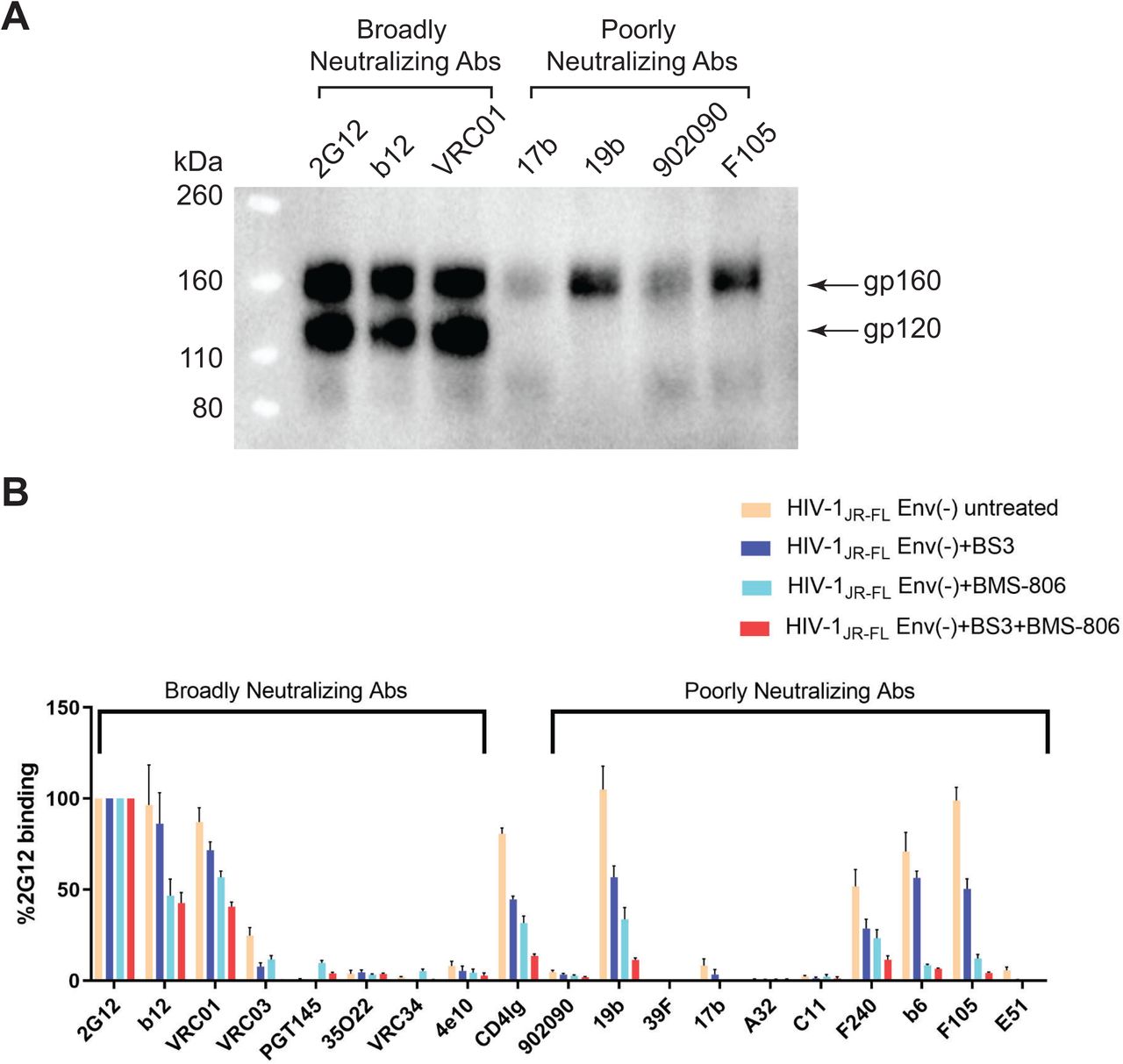
(A) HOS cells transiently expressing the wild-type HIV-1JR-FL Env, a fraction of which is cleaved in these cells, were incubated with the indicated broadly neutralizing antibodies or poorly neutralizing antibodies. After washing and lysis of the cells, the antibody-Env complexes were purified with Protein A-Sepharose beads and analyzed by Western blotting with a rabbit anti-gp120 polyclonal serum. (B) The effect of crosslinking with BS3 and/or BMS-806 treatment on antibody binding to HIV-1JR-FL Env(-) on the surface of CHO cells was evaluated by cell-based ELISA. BMS-806 (10 µM) was added to the CHO cells at the time of induction of Env(-) expression with doxycycline. All values were normalized against 2G12 antibody binding and were derived from at least three independent experiments. Note that the HIV-1JR-FL Env(-) glycoprotein is not recognized by the PGT145 V2 quaternary antibody, which serves as a negative control.
The HIV-1 entry inhibitor, BMS-806, hinders transitions from State 1 and modestly increases the occupancy of State 1 by the mature, wild-type HIV-1 Env (see Table 1) (42,53,54,79,81). BMS-806 treatment or glutaraldehyde crosslinking has been shown to shift the antigenic profile of uncleaved HIV-1 Env closer to that of the cleaved Env (79, 80). Incubating virions containing uncleaved Env with BMS-806 significantly enriched the low-FRET State-1 conformation, resulting in a conformational profile closer to that of the unliganded mature HIV-1 Env (Table 1) (81). We tested the effects of BMS-806 and the lysine-specific crosslinker, bis (sulfosuccinimidyl) suberate (BS3), on the antigenic profile of cleavage-defective HIV-1JR-FL Env(-) expressed on the surface of CHO cells (Fig. 1B). Treatment with BMS-806 and BS3 additively decreased Env(-) recognition by pNAbs (19b, b6, F105 and F240) and CD4-Ig, which preferentially bind Env conformations other than State 1 (45,48,54,78,81). In comparison, for the bNAbs 2G12, b12 and VRC01, the BMS-806/BS3-treated Env(-) was recognized at more than 40% the level observed for the untreated Env(-). These results are consistent with previous studies suggesting that BMS-806 can decrease the State-2/3 occupancy of uncleaved HIV-1 Envs anchored in the viral or cell membranes (Table 1) (79, 81).
Purification and characterization of Env(-) trimers
To investigate further the range of conformations sampled by the uncleaved HIV-1 Env, we purified full-length HIV-1JR-FL Env(-) trimers from the membranes of inducibly-expressing CHO cells (Fig. 2A and B). The CHO cells were incubated with BMS-806 during Env(-) synthesis in an attempt to shift occupancy from States 2/3 to State 1. BMS-806 treatment of the Env(-)-expressing cells reduced the synthesis of sialidase-sensitive and Endoglycosidase H-resistant glycoforms that are relatively enriched in complex carbohydrates (Fig. 2C). Glycosylation analysis revealed that BMS-806 treatment led to decreased complex sugar addition to the glycans modifying gp120 asparagine residues 88, 156, 160, 241, 362 and 463 (Fig. 2D and E). The effects of BMS-806 on Env(-) conformation apparently influence the conversion of particular high-mannose glycans to complex carbohydrates in the Golgi.

(A) Purified HIV-1JR-FL Env(-) without and with crosslinking by BS3 was run on a NUPAGE 4-12% BT gel stained by Coomassie Blue. (B) Purified HIV-1JR-FL Env(-) crosslinked by BS3 was run on a NativePAGE 4-16%BT gel and subjected to Western blotting with an HRP-conjugated anti-HIV-1 gp120 antibody. (C-E) To evaluate the effect of BMS-806 on the glycosylation of the synthesized Env(-) glycoprotein, BMS-806 (10 µM) was added to the CHO cells at the time of doxycycline induction. (C) The effect of BMS-806 on HIV-1JR-FL Env(-) glycosylation was evaluated by Western blotting after digestion with glycosidases (sialidase, Peptide-N-glycosidase F (PNGase F), and Endoglycosidase H (Endo H)). The purified HIV-1JR-FL Env(-) glycoproteins were digested with the indicated glycosidase, run on a NUPAGE 4-12% BT gel, and subjected to Western blotting with an HRP-conjugated anti-HIV-1 gp120 antibody. The results shown are representative of those obtained in three independent experiments. Note that BMS-806 treatment decreases Env(-) heterogeneity by reducing the levels of sialidase-sensitive and Endo H-resistant glycoforms. (D,E) The bar graphs show the glycan profiles at each glycosylation site of HIV-1JR-FL Env(-) purified from untreated CHO cells (D) or CHO cells treated with 10 µM BMS-806 (E), as determined by mass spectrometry. The glycan composition (in percent) was broadly characterized as high-mannose (red bars) or processed (complex + hybrid) glycans (blue bars).
To purify the Env(-) trimer complexes, membranes from BMS-806-treated CHO cells were incubated with saturating concentrations of BMS-806, crosslinked with BS3, and solubilized in Cymal-5. The detergent in the Env(-) glycoprotein solution was exchanged to a mixture of 4.5 mg/ml amphipol A8-35 and 0.005% Cymal-6 prior to cryo-plunging the samples in preparation for eventual cryo-electron microscopy (cryo-EM) imaging. Parallel smFRET studies estimated that only 26% of detergent-solubilized Env(-) was in a low-FRET conformation consistent with State 1 (Fig. 3A). The majority (74%) of the solubilized Env(-) glycoproteins assumed high- and intermediate-FRET conformations consistent with States 2 and 3, respectively. Thus, compared with BMS-806-treated Env(-) on virions, the Env(-) glycoproteins solubilized and purified from CHO cells exhibit less State 1 and more State 2/3 conformations (Table 1).

(A) HIV-1JR-FL Env(-) with V1 and V4 labeling tags was purified from 293T cell membranes using a protocol identical to that used for preparation of Env(-) for cryo-EM imaging. The purified Env(-) was labeled and analyzed by smFRET. FRET trajectories were compiled into a population FRET histogram and fit to the Gaussian distributions associated with each conformational state, according to a hidden Markov model (42). The percentage of the population that occupies each state as well as the number of molecules analyzed (N) is shown. The error bars represent the standard deviation from three independent data sets. (B) Membranes from BMS-806-treated CHO cells expressing HIV-1JR-FL Env(-) were crosslinked with BS3 and then solubilized in Cymal-5 detergent. The lysate was successively incubated with the 19b anti-gp120 (V3) antibody and Protein-A Sepharose beads. The Env(-) glycoproteins precipitated by the 19b antibody or by the negative-control Protein A-Sepharose beads during the indicated rounds of immunoprecipitation were analyzed by SDS-PAGE and Western blotting (upper left panel). The Env(-) glycoproteins in the initial cell membrane lysate (Input) and those glycoproteins remaining after four rounds of 19b counterselection were precipitated with Ni-NTA beads or the indicated antibodies; the precipitated Env(-) glycoproteins were analyzed by SDS-PAGE and Western blotting (upper right panel). The total amounts of Env(-) glycoprotein in the input and after 19b counterselection, normalized to the input Env(-) glycoprotein amount precipitated by the Ni-NTA beads, are shown in the bar graph (lower panel). Means and standard deviations derived from two independent experiments are shown.
The increased exposure of the gp120 V3 loop is a sensitive indicator of HIV-1 Envs that have undergone transitions from a State-1 conformation (48,54,92–94). We tested the ability of the 19b anti-V3 antibody, which does not neutralize most primary HIV-1 strains, to precipitate the BMS-806-treated, BS3-crosslinked Env(-) trimers solubilized in Cymal-5 detergent (Fig. 3B). After successive precipitations with the 19b antibody, approximately 85% of the Env(-) glycoprotein was removed from the CHO cell lysate. Therefore, even in the presence of BMS-806 and after BS3 crosslinking, most of the Env(-) trimers solubilized in Cymal-5 detergent apparently sample non-State-1 conformations. Together with the above cell-based ELISA and smFRET results, these experiments suggest that detergent solubilization destabilizes the uncleaved Env, even after BMS-806 and BS3 treatment. Therefore, the cell membrane and lipid-protein interactions may be important for the stabilization of the Env(-) State-1 conformation.
Env(-) structure determination by cryo-electron microscopy (cryo-EM)
The BMS-806-treated, BS3-crosslinked HIV-1JR-FL Env(-) trimers, purified in Cymal-5 and exchanged into amphipol A8-35 and Cymal-6, were analyzed by cryo-EM. We collected cryo-EM data from both a 200-kV FEI Tecnai Arctica microscope without an energy filter and a 300-kV FEI Titan Krios microscope with a Gatan BioQuantum energy filter, in video frames of a super-resolution counting mode with the Gatan K2 Summit direct electron detector (Fig. 4A-F). While both 200-kV and 300-kV cryo-EM datasets gave rise to consistent results, the final reconstructions at near-atomic resolution were achieved using the 300-kV cryo-EM dataset; the 300-kV dataset incorporates single-particle data collected at a high tilt angle of the sample stage to alleviate the effect of the strong orientation preference of the Env(-) particles. By contrast, the 200-kV cryo-EM dataset, which lacks tilted data, fell short of achieving a comparable level of resolution and suffered from the orientation preference of the particle images. However, despite the modest level of resolution (5.5-8 Å), extensive 3D classification of the 200-kV dataset, as detailed in a bioRxiv preprint (95), indicated the existence of multiple Env(-) conformations, some of which are consistent with the higher-resolution reconstructions obtained from the 300-kV dataset. This paper focuses on interpreting two higher-resolution maps of the uncleaved Env(-) trimer derived from the 300-kV dataset.
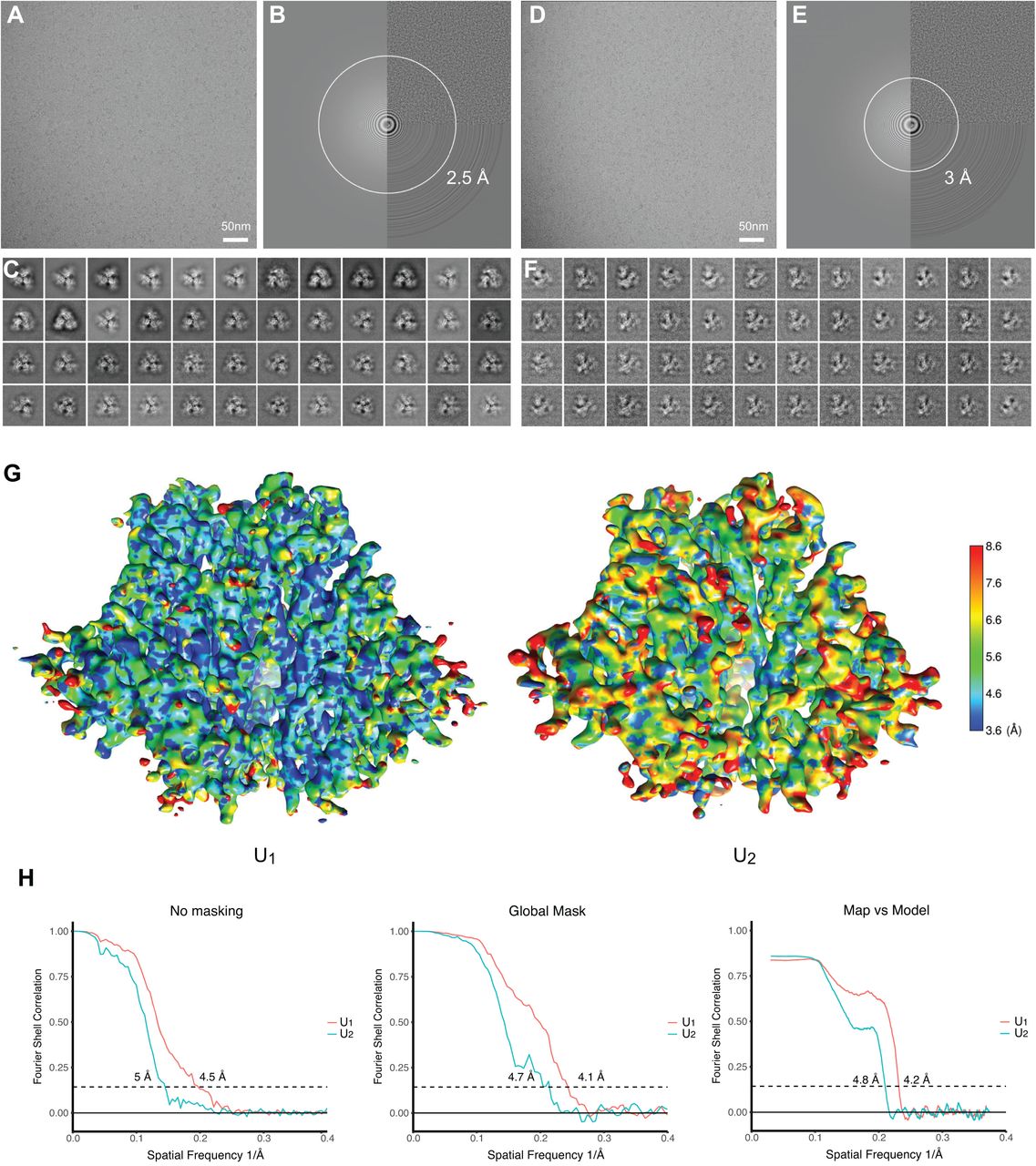
(A) A typical cryo-EM micrograph of Env(-) trimers taken with a Gatan K2 direct electron detector at 0 degrees of tilt. (B) Fourier transform of the image in A . (C) Unsupervised 2D class averages for non-tilt particles. (D) A typical cryo-EM micrograph of Env(-) trimers taken with a Gatan K2 direct electron detector at 45 degrees of tilt. (E) Fourier transform of the image in D. (F) Unsupervised 2D class averages for tilted particles. (G) The local resolution measurement of the State-U1 and State-U2 maps, as measured by ResMap (143). The maps are colored according to the local resolution, indicated by the color gradient (units in Angstroms). Side views of the Env(-) maps are shown, with gp120 at the bottom of the figure and gp41 at the top. (H) The gold-standard FSC plots of the State-U1 and State-U2 cryo-EM maps.
Analysis of the 300-kV data resulted in two major 3D classes, herein designated State U1 and State U2, respectively comprising 37% and 17% of the imaged particles, after removal of junk particles. The State-U1 and State-U2 maps were refined to 4.1 and 4.7 Å, respectively, without imposing any symmetry during refinement and reconstruction (Fig. 4G and H). The map quality allowed atomic modelling and refinement with accuracy to the level of the Cα backbone trace. By contrast, imposing C3 symmetry during refinement and reconstruction resulted in lower resolution and poorer structural features in the refined density maps of both State-U1 and State-U2, suggesting that both conformations indeed lack rigorous three-fold symmetry. Other 3D classes derived from the HIV-1JR-FL dataset were not able to be refined to comparable levels of resolution, and thus are not further analyzed and discussed herein. Curiously, no major 3D classes with rigorous three-fold symmetry were found when extensive 3D classification was conducted using the maximum-likelihood method without imposing C3 symmetry (96). This likely reflects the intrinsic conformational plasticity of the Env(-) glycoprotein, although we do not rule out the contribution of preparation-dependent variables, such as asymmetric crosslinking between adjacent protomers.
Key structural features of the asymmetric uncleaved HIV-1 Env trimers
The U1 and U2 Env(-) trimers share an overall topology with existing structures of soluble and membrane HIV-1 Env trimers (97–108) (Fig. 5A). A central feature of all these structures is a 3-helix bundle (3-HBC) formed by the C-terminal portion of the gp41 HR1 region (HR1C); the gp120 subunits project outward from this central helical coiled coil. These common features allowed us to use existing symmetric and asymmetric HIV-1 Env trimer structures as references to build structural models of states U1 and U2. All three individual protomers in the U1 and U2 trimers exhibit similar folds, with Cα RMSD values of ∼2 Å (Fig. 5B and C). While both the U1 and U2 conformations of Env(-) are asymmetric, they exhibit different degrees of such asymmetry in terms of the relative rotation of the individual protomers with respect to the trimer axis. The protomers are differentially translated and rotated with respect to each other in unique ways in the U1 and U2 trimers (Fig. 5D), generating ∼3-4 Å movement overall in the gp120 outer domain (OD). When one of the protomers is used to align both conformations, the other two protomers of U2 are notably rotated by 2.8 and 4 degrees relative to the corresponding protomers of U1 (Fig. 5A). This creates the smallest and largest openings between two adjacent protomers in U2, the more asymmetric of the two Env(-) conformations. Alignment of all three protomer structures in each conformation indicates that the asymmetric conformations are facilitated by local structural rearrangements of residues 546-568 at the inter-protomer interface. This gp41 segment (HR1N) is immediately N-terminal to the central 3-HBC and exhibits the greatest local structural variation among the promoters. Notably, the overall structural variation of gp41 among the U1 and U2 protomers is greater than that of the gp120 core structure, presumably because gp41 contributes more interactions to the inter-protomer interface. Consistently, the gp120 trimer association domain (TAD), which includes the V1/V2 and V3 regions, exhibits greater conformational variation in U2 than in U1, leading to an overall greater extent of asymmetry in U2 (Fig. 5B and C). There is similarly greater gp41 structural variation among the protomers in U2 than in U1.

(A) Protomer 2 of the State-U1 and State-U2 models are superposed, showing that protomer 1 and protomer 3 are rotated 4.0° and 2.8°, respectively. (B) Three protomers of the State-U1 model are superposed. (C) Three protomers of the State-U2 model are superposed. (D) With protomer 2 of the State-U1 and State-U2 models superposed, the Cα distances between the same residues on the U1 and U2 structures are measured for four residues (from (i) to (iv): T90, D230, S481 and N392). In the side views of Env(-) shown in B-D, gp120 is at the bottom of the figure and gp41 at the top.
Comparison with structures of cleaved HIV-1 Env trimers
We compared the U1 and U2 HIV-1JR-FL Env(-) structures to those of mature (cleaved) HIV-1 Env trimers. The structure of the unliganded HIV-1BG505 sgp140 SOSIP.664 glycoprotein (PDB: 4ZMJ) provides an example of a stabilized soluble Env trimer with C3 symmetry (104). Structures of cytoplasmic tail-deleted, detergent-solubilized HIV-1JR-FL and HIV-1AMC011 EnvΔCT trimers have been solved in complex with Fab fragments of the PGT151 neutralizing antibody (PDB: 5FUU and 6OLP, respectively) (105, 108). Binding of the PGT151 Fabs introduces asymmetry into the Env trimer, limiting the binding stoichiometry to two Fabs per trimer.
The folds of the U1 and U2 Env(-) protomers resemble those of the sgp140 SOSIP.664 and PGT151-bound EnvΔCT protomers (Fig. 6). The largest structural difference is localized in HR1N residues 534-570 leading to the central 3-HBC of gp41. When the U1 and sgp140 SOSIP.664 trimer structures are aligned using one of the protomers, the other two protomers of U1 exhibit rotations in opposite directions relative to the symmetric sgp140 SOSIP.664 trimer structure, causing a prominent narrowing of the opening angle between these two protomers in the U1 trimer structure (Fig. 6A). By contrast, when the U1 structure is aligned to the PGT151-bound EnvΔCT trimer using the protomer free of the antibody, both the other two protomers exhibit rotations in the same direction; this results in two smaller opening angles and one notably larger opening angle in comparison with those seen in the symmetric sgp140 trimer (Fig. 6B). In addition to relative rotation, the gp120 components of the U1 protomers also exhibit outward movement in both comparisons (Fig. 6A and B), giving rise to a slightly wider trimer footprint (Fig. 7A). Some local divergence in the gp120 V1/V2 region and gp41 α8 helix between HIV-1JR-FL Env(-) and HIV-1BG505 sgp140 SOSIP.664 likely results from strain-dependent differences in primary sequence. Consistent with this explanation, the protomer structures of the Env(-) and EnvΔCT trimers, both derived from the HIV-1JR-FL strain, align well in these regions. As is the case for all current HIV-1 Env trimer structures, the gp41 membrane-proximal external region (MPER) and transmembrane region are disordered in the U1 and U2 maps.

(A) Left: Protomer 1 of the State-U1 trimer is superposed on the unliganded HIV-1BG505 sgp140 SOSIP.664 trimer (PDB ID 4ZMJ) (104), demonstrating how the other two protomers in State-U1 are rotated towards each other. Right: Side views of the superposed protomers, with red parts representing the major areas of difference between the two protomers. (B) Left: Protomer 1 of the State-U1 trimer is superposed on the HIV-1JR-FL EnvΔCT trimer complexed with PGT151 Fabs (PDB ID 5FUU) (105), indicating that binding of the PGT151 antibodies introduces asymmetry into the Env trimer that differs from that of U1. Right: Side views of the superposed protomers, with red parts representing the major areas of difference between the two protomers. In the side views of the Env protomers shown in the right-hand panels of A and B, gp120 is at the bottom of the figure and gp41 at the top.

(A) The inter-protomer distances (in Å) between selected atoms of gp120 and gp41 are shown for different Env structures: the smaller, inner triangle depicts distances measured between gp41 residues W628 and I635; the larger, outer triangle depicts distances measured between gp120 residues A336 and Q352. The U1 and U2 structures are compared with those of the unliganded sgp140 SOSIP.664 trimer (PDB ID 4ZMJ) (104) and the PGT151-bound HIV-1JR-FL and HIV-1AMC011 EnvΔCT trimers (PDB IDs 5FUU and 6OLP, respectively) (105,108). For 5FUU and 6OLP, the sides of the Env trimer that are bound by the PGT151 Fabs are marked. (B) The three gp120 subunits of four Env trimer atomic structures were superposed with the gp120 subunits of the State-U1 Env(-) trimer. Each protomer was aligned separately. After gp120 alignment, the relative positions of the gp41 HR1C helixes are jointly shown here. In each case, the U1 HR1C helices are colored cyan. With gp120 aligned, the gp41 in State U1 is displaced compared with the other structures. Upper row: top views of 3-helix bundles; Bottom row: side views of 3-helix bundles. 5FYK is the structure of an HIV-1JR-FL sgp140 SOSIP.664 trimer complexed with several neutralizing antibody Fabs (65).
We next compared the topology of the Env(-) trimers to that of cleaved Env trimers. The inter-protomer distances between arbitrarily chosen atoms on the outer surface of gp120 and gp41 provide a measure of trimer geometry (Fig. 7A). Of the trimers that we compared, the symmetric HIV-1BG505 sgp140 SOSIP.664 trimer is the most tightly packed, with the respective gp120 and gp41 sides 77.3 and 39.3 Å in length. The two sides of the EnvΔCT trimers bound to the PGT151 antibody Fabs are similar in length (gp120: 75.4, 77.1/gp41: 37.4, 37.4 Å and gp120: 75.5, 76.0/gp41: 37.5, 37.8 Å in the HIV-1JR-FL and HIV-1AMC011 EnvΔCT trimers, respectively); these Fab-bound sides are shorter than the “opened” unliganded side (gp120: 83.6/gp41: 46.2 Å and gp120: 84.8/gp41: 46.6 Å in the HIV-1JR-FL and HIV-1AMC011 EnvΔCT trimers, respectively). The asymmetry of the U1 Env(-) trimer is qualitatively similar to that of the U2 trimer; the asymmetry of the Env(-) trimers is distinguished by three sides of different lengths and therefore differs from the asymmetry in the EnvΔCT trimers induced by the PGT151 antibody. Notably, the average lengths of the gp120/gp41 sides of the Env(-) trimers are longer than those of the unliganded sgp140 SOSIP.664 or PGT151-bound EnvΔCT trimers, indicating that the uncleaved Env(-) trimers are packed less tightly than the cleaved Env trimers.
To evaluate the basis for the increased “openness” of the uncleaved Env(-) trimers, we compared the structures of the gp41 3-HBC coiled coil and HR1N region in the Env(-) and cleaved Env trimers. Changes in the packing or orientation of the 3-HBC coiled coil could potentially influence trimer topology. Although it appears that the crossing angles between two adjacent helices in the gp41 3-HBC coiled coil are very similar in the U1 and U2 trimers, these 3-HBC helices exhibit differential packing and asymmetric features in U1 and U2 that are amplified into a greater degree of overall trimeric asymmetry. Compared to the PGT151-bound cleaved Env structures (PDB IDs 5FUU and 6OLP), the U1 conformation has clearly larger crossing angles and thus a greater 3-HBC coiled-coil footprint (Fig. 7B). By contrast, the crossing angles in U1 are nearly identical to those of the sgp140 SOSIP.664 trimers, but the U1 3-HBC helices exhibit marked translation in opposite directions that breaks the trimer symmetry seen in the crystal structures of the sgp140 SOSIP.664 trimers (PDB IDs 5FYK and 4ZMJ). Being able to sustain structural rearrangements involving both of the orthogonal degrees of freedom demonstrates that the Env trimer metastability and lability is potentially rooted in the conformational plasticity and flexibility of the central 3-HBC structure.
Despite a high degree of primary sequence conservation among HIV-1 strains, the gp41 HR1N region (residues 541-570) exhibits significant conformational polymorphism among current HIV-1 Env trimer structures. In the pre-triggered (State-1) Env conformation, the gp41 HR1N region has been implicated in the non-covalent association with gp120; in the pre-hairpin intermediate (State 3), the HR1N region forms part of the extended HR1 helical coiled coil (14-16,109-111). Therefore, HR1N may transition from an as-yet-unknown State-1 structure to a helical coiled coil (State 3) as Env “opens” upon binding CD4. The HR1N region is relatively disordered in most sgp140 SOSIP.664 structures, probably as a result of the I559P change used to stabilize these soluble Env trimers (112–115). Even in asymmetric structures of sgp140 SOSIP.664 trimers bound to soluble CD4 and the E51 CD4-induced antibody (116), HR1N disorder precludes analysis. We therefore limited our comparison to asymmetric Env trimers for which HR1N structural information is available. Comparison of the HR1N conformation in the asymmetric Env trimers suggested that the helicity of the HR1N region is related to the degree of “openness” of the corresponding protomer (Fig. 8). Lower helicity of the HR1N region leads to a somewhat collapsed conformation that is correlated with a smaller inter-protomer opening angle. This is consistent with the notion that a non-helical, loop-like and more collapsed HR1N, which is located in the crevice formed by the protomer arms, would not have sufficient structural strength to sustain a wider opening angle. These observations support the proposition that the HR1N conformation is allosterically coupled with asymmetric features of the 3-HBC and the overall asymmetry of the entire trimer.

(A) Sequences of the gp41 HR1N region from three U1 protomers are shown, with residues in α-helices highlighted in red. (B) The relationship between HR1N helicity and the opening angle of three asymmetric HIV-1 Env trimers (U1 and two PGT151-Fab-bound EnvΔCT trimers (PDB IDs 5FUU and 6OLP)) is shown. The x-axis represents the opening angle for each of three sides, measured using the “angle_between_domains” command in Pymol (142). The y-axis represents the number of residues in an α-helical conformation for the HR1N region of that side. (C) The HR1N and HR1C regions from the three indicated atomic models are superposed. (D) The HR1N regions from the three protomers in State U1 are shown .
Env(-) glycosylation
Most of the peptide-proximal density associated with N-linked glycosylation is preserved in the U1 map and was modeled (Fig. 9). Most distal glycan residues are not well resolved, reflecting their dynamic nature and heterogeneity. As has been previously shown, the high-mannose glycans are clustered in a patch on the surface of the gp120 outer domain (39,40,65,117). No glycan-associated density on Asn 297 is detectable, and the glycan signal on Asn 448 is weak. The signals associated with the complex glycans on gp41 residues Asn 611 and Asn 637 are buried in noise. The most membrane-proximal gp41 glycan on Asn 616 is largely modified by high-mannose glycans.

Glycans on State-U1 trimers are colored according to the following scheme: glycans that exhibit significant decreases in the addition of processed glycans as a result of BMS-806 treatment are colored purple; high-mannose glycans are colored yellow; and the remaining mixed or processed glycans that are not affected by BMS-806 binding are colored green.
BMS-806 treatment of Env(-)-expressing cells led to a reduction in the modification of glycans on Asn 88, 156, 160, 241, 362 and 463. Asn 88 and 241 are located at the gp120-gp41 interface, and Asn 156 and 160 at the trimer apex (Fig. 9). Previous studies have suggested that BMS-806 can strengthen inter-subunit and inter-protomer interactions in the Env trimer, increasing the binding of neutralizing antibodies that recognize the gp120-gp41 interface and trimer apex (79). Strengthening these interactions may render the carbohydrates in these regions less available for modification to complex carbohydrates. Consistent with this, two other BMS-806-sensitive glycans (on Asn 362 and Asn 463) reside on the perimeter of the gp120 outer domain that, in a more closed trimer, might be sterically limited by inter-protomer effects.
BMS-806 binding site
The binding site of BMS-806 in sgp140 SOSIP.664 complexes (PDB: 5U7M) has been previously characterized (118). In the Env(-) maps, density corresponding to the location of BMS-806 in the sgp140 SOSIP.664 complexes is evident. In the Env(-) complexes, BMS-806 is located in the gp120 Phe 43 cavity and the adjacent water-filled channel, sandwiched between Trp 427 and Trp 112. Although the level of resolution does not allow unambiguous definition of the binding mode, the position and orientation of BMS-806 is consistent with that in the sgp140 SOSIP.664 complexes (118) (Fig. 10). In the U1 Env(-)-BMS-806 structure, as in the unliganded and BMS-806-bound sgp140 SOSIP.664 structures (104, 118), Layer 1 of the gp120 inner domain appears to be stabilized by the insertion of Trp 69 into the back end of the Phe 43 cavity, where it interacts orthogonally with Trp 112. During the course of Env binding to CD4, Layer 1 is thought to undergo rearrangement to decrease the off-rate of CD4 (119); fixation of Layer 1 by BMS-806 could help to inhibit Env conformational transitions to the CD4-bound State 3.

The BMS-806 binding sites within three protomers of the State-U1 structure (cyan) are compared with those in the BMS-806-bound sgp140 SOSIP.664 trimer (PDB 5U70) (118).
Effect of BMS-806 on processing of wild-type HIV-1 Env
BMS-806 and its analogues block transitions from the pre-triggered Env conformation; thus, addition of these compounds to cleaved and uncleaved Envs on virions enriches State 1 (Table 1) (42,53,54,79,81). The studies shown in Figure 2D and E suggest that limiting the conformational flexibility of the cleavage-defective Env(-) by exposing Env(-)-expressing cells to BMS-806 can influence the processing of carbohydrate structures. To evaluate more thoroughly how Env conformation influences its processing, we used A549-Gag/Env cells, which produce virus-like particles (VLPs) containing Env (72). The wild-type HIV-1AD8 Env in the A549-Gag/Env cells is proteolytically processed and the VLPs contain mostly cleaved Env, as is the case for authentic HIV-1 virions (72). Therefore, the use of A549-Gag/Env cells allowed us to evaluate the effects of BMS-806 on the cleavage and glycosylation of wild-type HIV-1 Env in cells and on VLPs.
We incubated A549-Gag/Env cells with BMS-806 and studied Env in cell lysates and VLPs. BMS-806 treatment during Env expression resulted in a decrease in the efficiency of Env cleavage (Fig. 11A). The uncleaved Env produced in the presence of BMS-806 was efficiently incorporated into VLPs (Fig. 11B). This contrasts with the relative exclusion of uncleaved Env from VLPs produced in the absence of BMS-806 (Fig. 11B) (72). In the untreated cells, some of the glycans on the uncleaved Env are Endoglycosidase Hf-resistant and therefore are complex carbohydrates (Fig. 11A). The Endoglycosidase Hf-resistant fraction of the uncleaved Env migrated faster on SDS-polyacrylamide gels following BMS-806 treatment, indicating that fewer complex sugars are added to Env produced in A549-Gag/Env cells treated with BMS-806 (Fig. 11A). Nonetheless, in the BMS-806-treated cells, the uncleaved Env that is modified by complex glycans (and therefore has passed through the Golgi) is incorporated into VLPs (Fig. 11B). These results suggest that the BMS-806-induced reduction in the conformational flexibility of the Env precursor decreases the efficiency of gp160 cleavage and addition of some complex glycans, without significantly affecting Env transport through the Golgi or incorporation into VLPs.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
A549-Gag/Env cells were treated with BMS-806 (10 µM) or mock treated during doxycycline induction of Gag/Env expression. Lysates were prepared from cells (A) and supernatants containing virus-like particles (VLPs) (B), and were treated with Peptide-N-glycosidase F (PNGase F) or Endoglycosidase Hf (Endo Hf), or mock treated (no Rx). The Envs were run on reducing SDS-polyacrylamide gels and analyzed by Western blotting. The deglycosylated gp160, gp120 and gp41 proteins (dgp160, dgp120 and dgp41, respectively) are indicated by arrows (red – PNGase F-treated sample; green – Endo Hf-treated sample). Data in this figure are representative of those obtained in two independent experiments.
DISCUSSION
The uncleaved HIV-1 Env serves as a precursor to the cleaved functional Env and, by eliciting poorly neutralizing antibodies, as a potential decoy to the host immune system. Antibody or ligand binding and smFRET analyses indicate that the Env precursor can sample multiple conformations that resemble States 1, 2 and 3 of the mature viral Env spike (73–81). The conformational plasticity of the Env precursor contrasts with the behavior of the mature Env, which in the absence of ligands largely resides in State 1 (42, 81). Therefore, proteolytic cleavage stabilizes State-1 Env, which is highly resistant to neutralization by antibodies recognizing other Env conformations. Although proteolytic maturation also primes the membrane-fusing potential of other Class I viral membrane fusion proteins, the effects of cleavage on HIV-1 Env conformational plasticity are unusual. For example, crystal structures comparing the influenza virus precursor, HA0, with the cleaved HA1/HA2 trimer showed differences only in the immediate vicinity of the cleavage site (120). Uncleaved HIV-1 Envs can be transported from the endoplasmic reticulum to the cell surface by bypassing the Golgi or, when trafficking through the classical secretory pathway, by escaping furin cleavage in the Golgi (72). Both subsets of uncleaved Envs on the surface of expressing cells can be recognized by pNAbs and therefore represent a potentially abundant source of Env conformations other than State 1 (72, 79). The resulting diversion of host antibody responses away from State-1 Env, the major target for neutralizing antibodies, would have considerable advantages for a persistent virus like HIV-1.
BMS-806 can enrich State 1 in the uncleaved membrane-anchored Env (79, 81) and BS3 crosslinking could hypothetically help to stabilize this conformation. Nonetheless, once Env(-) glycoproteins were solubilized in detergent, these treatments did not prevent Env(-) from assuming non-State-1 conformations. The loss of membrane interactions (122), the effects of detergents or other manipulations during purification may have contributed to diminished State-1 occupancy in this case.
Our structural and biophysical analyses indicate that the cleaved Env conformation seen in the sgp140 SOSIP.664 trimers is also sampled by the uncleaved Env, but notably, in an asymmetric fashion. Thus, although the asymmetry of the U1 and U2 uncleaved Env trimers alters the quaternary relationships among the Env protomers, the fold of the individual Env(-) protomers resembles those of sgp140 SOSIP.664 and PGT151-bound EnvΔCT trimers. Analysis by smFRET has suggested that these Envs are predominantly in a State-2-like conformation (121). By analogy, we deduce that U1 and U2 represent State-2-like conformations. State 2 has been suggested to represent a default intermediate conformation favored by Envs that experience a destabilization of State 1 (48,49,54,82,121). CD4 binding to the wild-type HIV-1 Env trimer sequentially induces State-2 and State-3 conformations in the bound protomer, whereas the other, ligand-free protomers in the Env trimer assume State-2 conformations (49). Although PGT151 is a broadly neutralizing antibody and can presumably interact with State-1 Envs, it induces asymmetry in the Env trimer, causing the Env protomers to assume State-2-like conformations (121). Thus, breaking symmetry in the HIV-1 Env trimer often results in the adoption of a State-2 conformation, consistent with the proposed default nature of this intermediate.
Asymmetry of both uncleaved and cleaved Env trimers appears to be related to the structural plasticity and flexibility of the gp41 HR1N region, which is directly situated in the inter-protomer interface and is allosterically coupled with the quaternary Env conformation. On the one hand, the HR1N structure can directly affect the packing of the central 3-HBC coiled coils; on the other hand, the HR1N rigidity can allosterically regulate the inter-protomer opening angle. Mutagenesis studies have suggested that in the pre-triggered (State-1) Env conformation, the HR1N region contributes to the non-covalent association of gp120 with gp41 (109-111). We observed a relationship between the inter-protomer opening angle of asymmetric Env trimers and HR1N helicity. As initial CD4 binding to the Env trimer occurs asymmetrically, with State-2 conformations assumed by the unbound protomers (49), the HR1N regions presumably transition from as-yet-unknown State-1 conformations to predominantly helical conformations. Subsequent assembly of three HR1N helices into the extended gp41 coiled coil [(HR1N+C)3] projects the fusion peptide toward the target membrane.
The symmetry of the mature, pre-triggered (State-1) HIV-1 Env trimer likely contributes to its ability to evade pNAbs. Supporting this assertion is the previous observation that the fraction of cell-surface Env recognized by bNAbs crosslinked into trimers, whereas the cell-surface Env that was recognized by pNAbs crosslinked into dimers and monomers, possible reflecting trimer asymmetry (72). The asymmetry observed for the uncleaved Env(-) U1 and U2 trimers potentially allows pNAbs to access their epitopes with minimal steric hindrance. Indeed, pNAbs directed against the gp120 V3 region or CD4-binding site can be docked into the open face of the U1 Env trimer with only minimal readjustment of surrounding structures to remove steric clashes (data not shown). Maintaining C3 symmetry may be one prerequisite for preserving an antibody-resistant State-1 Env conformation. Our study implicates the conformationally labile gp41 HR1N segment in maintaining trimer symmetry, and the high-resolution structure of this functionally important region in a State-1-compatible Env conformation is a future goal.
The intrinsic conformational heterogeneity of the uncleaved HIV-1 Env trimer and the low occupancy of certain conformational states present significant challenges to their structural characterization. Previous studies of detergent-solubilized uncleaved HIV-1 Envs with truncated cytoplasmic tails were performed without extensive 3D classification and with C3 symmetry imposed, resulting in lower-resolution structures (123, 124). Our current study takes advantage of subsequent advances in 3D classification in cryo-EM technology and data processing to identify two major classes of Env(-) trimers, both asymmetric. Cryo-EM and smFRET analyses support the existence of other conformations in the Env(-) preparation, but high-resolution reconstruction of these conformers was unsuccessful (95). Current 3D hierarchical classification methods are prone to ignore or completely miss lowly populated conformational states or experience difficulties in precisely classifying these low-population conformations, which then leads to insufficient resolution for structure determination and functional interpretation (125). A more complete characterization of the multiple conformations assumed by the uncleaved HIV-1 Env may require approaches better able to deal with a high degree of structural heterogeneity than maximum-likelihood-based 3D classification (125,126).
BMS-806 inhibits HIV-1 entry, blocking CD4-induced transitions of the mature Env from a pre-triggered (State-1) conformation to downstream states (42,53,54,79,81). On the cell or viral membrane, uncleaved Env can respond to treatment with BMS-806 by increasing the occupancy of State 1 (79, 81). Consequently, BMS-806 decreases recognition of uncleaved Env by pNAbs, whereas recognition by most bNAbs is maintained or increased (55, 79). We found that BMS-806 also exerts a significant effect on Env during its maturation. BMS-806 treatment of cells expressing wild-type HIV-1 Env resulted in decreases in both gp160 cleavage and modification by complex carbohydrate structures; transport through the Golgi and incorporation into VLPs were not apparently blocked by BMS-806. These observations imply that gp160 conformational flexibility contributes to the efficiency with which the Env precursor is acted upon by furin and glycosylation enzymes. The requirement that functional Env is cleaved (25, 127) therefore provides selective pressure to maintain flexibility in the HIV-1 Env precursor. The resulting conformational heterogeneity of the Env precursor represents a potential advantage for a persistent virus like HIV-1 by skewing host antibody responses away from State 1. For immunization strategies employing membrane-anchored HIV-1 Env or during natural HIV-1 infection, treatment with BMS-806 analogues could potentially increase the presentation of the State-1 Env conformation to the immune system. BMS-806 analogues (79) could also assist future investigation of State-1-like conformations of uncleaved and cleaved HIV-1 Env trimers.
MATERIALS AND METHODS
Protein expression and purification
For expression of the uncleaved full-length membrane-anchored HIV-1JR-FL Env(-) glycoprotein, the env cDNA was codon-optimized and was cloned into an HIV-1-based lentiviral vector. These Env sequences contain a heterologous signal sequence from CD5 in place of that of wild-type HIV-1 Env. The proteolytic cleavage site between gp120 and gp41 was altered, substituting two serine residues for Arg 508 and Arg 511. In the HIV-1JR-FL Env(-) glycoprotein, the amino acid sequence LVPRGS-(His)6 was added to the C-terminus of the cytoplasmic tail. For Env(-) expression, the env coding sequences were cloned immediately downstream of the tetracycline (Tet)-responsive element (TRE). Our expression strategy further incorporated an internal ribosomal entry site (IRES) and a contiguous puromycin (puro) T2A enhanced green fluorescent protein (EGFP) open reading frame downstream of env (TRE-env-IRES-puro.T2A.EGFP). Uncleaved membrane-anchored Env(-) was produced by exogenous expression in CHO cells. Briefly, the HIV-1-based lentiviral vector encoding HIV-1JR-FL Env(-) was packaged, pseudotyped with the vesicular stomatitis virus (VSV) G protein, and used to transduce CHO cells (Invitrogen) constitutively expressing the reverse Tet transactivator (rtTA). High-producer clonal cell lines were derived using a FACSAria cell sorter (BD Biosciences) to isolate individual cells expressing high levels of EGFP. The integrity of the recombinant env sequence in the clonal lines was confirmed by sequence analysis of PCR amplicons. Clonal cultures were adapted for growth in a serum-free suspension culture medium (CDM4CHO; Thermo Fisher).
For the exogenous production of the Env(-) glycoprotein, cells were expanded in a suspension culture using a roller bottle system (Thermo) and were treated with 1 μg/ml of doxycycline and 10 μM BMS-378806 (herein referred to as BMS-806) (Selleckchem) after reaching a density of >4 × 106 cells per ml. After 18 to 24 h of culture with doxycycline and BMS-806, the cells were harvested by centrifugation. During the remainder of the purification procedure, 10 μM BMS-806 was added to all buffers. The cell pellets were homogenized in a homogenization buffer (250 mM sucrose, 10 mM Tris-HCl [pH 7.4], and a cocktail of protease inhibitors [Roche Complete EDTA-free tablets]). Membranes were then extracted from the homogenates by ultracentrifugation. The extracted crude membrane pellet was collected, resuspended in 1×PBS to a final concentration of 5 mg of wet membrane per ml of 1×PBS and crosslinked with 5 mM BS3 (Proteochem), followed by solubilization with a solubilization buffer containing 100 mM (NH4)2SO4, 20 mM Tris-HCl (pH 8.0), 300 mM NaCl, 20 mM imidazole, 1% (wt/vol) Cymal-5 (Anatrace), and a cocktail of protease inhibitors (Roche Complete EDTA-free tablets). The suspension was ultracentrifuged for 30 min at 100,000 × g and 4°C. The supernatant was collected and was mixed with a small volume of preequilibrated Ni-nitrilotriacetic acid (NTA) beads (Qiagen) for 2 h on a rocking platform at 4°C. The mixture was then injected into a small column and washed with a buffer containing 20 mM Tris-HCl (pH 8.0), 100 mM (NH4)2SO4, 1 M NaCl, 30 mM imidazole, and 0.5% Cymal-5. The beads were resuspended in a buffer containing 20 mM Tris-HCl (pH 8.0), 100 mM (NH4)2SO4, 250 mM NaCl, 4.5 mg/ml Amphipol A8-35 (Anatrace), 0.006% DMNG (Anatrace) and a cocktail of protease inhibitors (Roche Complete EDTA-free tablets), and incubated for 2 hours on a rocking platform. The mixture was applied to a column and the buffer was allowed to flow through. The beads were then resuspended in a buffer containing 20 mM Tris-HCl (pH 8.0), 100 mM (NH4)2SO4, 250 mM NaCl, 4.5 mg/ml Amphipol A8-35 (Anatrace) and a cocktail of protease inhibitors (Roche Complete EDTA-free tablets), and incubated for 2 hours on a rocking platform. The mixture was added to a column and the buffer allowed to flow through, followed by washing with 10 bed volumes of a buffer containing 20 mM Tris-HCl (pH 8.0), 100 mM (NH4)2SO4, and 250 mM NaCl. Proteins were eluted from the bead-filled column with a buffer containing 20 mM Tris-HCl (pH 8.0), 100 mM (NH4)2SO4, 250 mM NaCl, and 250 mM imidazole. The buffer of the eluted Env(-) glycoprotein solution was exchanged with imaging buffer containing 20 mM Tris-HCl (pH 8.0), 100 mM (NH4)2SO4, and 250 mM NaCl with a Centrifugal Filter (Millipore), and was concentrated. Before cryo-plunging, Cymal-6 (Anatrace) was added to the Env(-) glycoprotein solution at a final concentration of 0.005%.
Expression of wild-type HIV-1 Env and virus-like particles (VLPs)
Human A549 lung epithelial cells (ATCC) inducibly expressing Env and an HIV-1 Gag-mCherry fusion protein under the control of a tetracycline-regulated promoter were established as described (72). Briefly, A549-rtTA cells constitutively expressing the reverse tet transactivator were transduced with an HIV-1-based lentivirus vector expressing Rev and Env from HIV-1AD8, a primary HIV-1 strain (128). These A549-Env cells were transduced with a lentivirus vector expressing the HIV-1 Gag precursor fused with mCherry (72). The doxycycline-regulated expression of the Gag-mCherry fusion protein resulted in the release of Env-containing VLPs into the medium. Herein, we designate these cells A549-Gag/Env. The A549-Gag/Env cells were grown in DMEM/F12 supplemented with 10% FBS, L-glutamine and penicillin-streptomycin.
Antibodies
Antibodies against HIV-1 Env were kindly supplied by Dr. Dennis Burton (Scripps), Drs. Peter Kwong and John Mascola (Vaccine Research Center, NIH), Dr. Barton Haynes (Duke), Dr. Hermann Katinger (Polymun), Dr. James Robinson (Tulane) and Dr. Marshall Posner (Mount Sinai Medical Center). In some cases, anti-Env antibodies were obtained through the NIH AIDS Reagent Program. Antibodies for Western blotting include goat anti-gp120 polyclonal antibody (ThermoFisher) and the 4E10 human anti-gp41 antibody (Polymun). An HRP-conjugated goat anti-human IgG (Santa Cruz) and an HRP-conjugated goat anti-rabbit antibody (Santa Cruz) were used as secondary antibodies for Western blotting.
Single-molecule FRET: sample preparation, data acquisition and analysis
Analysis of the conformational dynamics of HIV-1 Env was performed after enzymatic labeling of the V1 and V4 regions of gp120 on the purified (His)6-tagged HIV-1JR-FL Env(-) glycoprotein with Cy3 and Cy5 fluorophores, respectively, as previously described (42). A transfection ratio of 20:1 of non-tagged: V1/V4-tagged HIV-1JR-FL Env(-) was used to ensure that only one protomer within a trimer carries enzymatic tags for site-specific labeling. The HIV-1JR-FL Env(-) glycoprotein was purified from transiently expressing 293T cells that had been treated with BMS-806 and crosslinked with BS3, as described above. The purified HIV-1JR-FL Env(-) glycoprotein in buffer (20 mM Tris-HCl (pH 8.0), 10 mM MgCl2, 10 mM CaCl2, 100 mM (NH4)2SO4, 250 mM NaCl, 0.005% Cymal-6, 10 µM BMS-806) was labeled with Cy3B(3S)-cadaverine (0.5 µM), transglutaminase (0.65 µM; Sigma Aldrich), LD650-CoA (0.5 µM) (Lumidyne Technologies), and AcpS (5 µM) at room temperature overnight. After labeling, Env(-) trimers were purified using ZebaTM spin desalting columns (ThermoFisher) to remove free dyes. Finally, prior to imaging, fluorescence-labeled HIV-1JR-FL Env(-) carrying the (His)6 epitope tag was incubated with biotin-conjugated anti-(His)6 tag antibody (HIS.H8, Invitrogen) at 4° for two hours.
All smFRET data were acquired on a home-built total internal reflection fluorescence (TIRF) microscope, as previously described (42, 129). Fluorescently labeled HIV-1JR-FL Env(-) trimers were immobilized on passivated streptavidin-coated quartz microscopy slides and washed with pre-imaging buffer specifically made for this experiment. The pre-imaging buffer consisted of 20 mM Tris HCl (pH 8.0), 100 mM (NH4)2SO4, 250 mM NaCl, 0.005% Cymal-6, and 10 µM BMS-806. For smFRET analysis, a cocktail of triplet-state quenchers and 2 mM protocatechuic acid (PCA) with 8 nM protocatechuic 3,4-dioxygenase (PCD) were added to the above pre-imaging buffer to remove molecular oxygen. Cy3 and Cy5 fluorescence was detected with a 60x water-immersion objective (Nikon), split by a diachronic mirror (Chroma), and imaged on two synchronized ORCA-Flash4.0v2 sCMOS cameras (Hamamatsu) at 40 frames/seconds for 80 seconds.
smFRET data analysis was performed on the customized Matlab (Mathworks) program SPARTAN (129). Fluorescence intensity trajectories were extracted from recorded movies, and FRET efficiency (FRET) was calculated based on FRET= IA/(ϒID+IA), where ID and IA are the fluorescence intensities of donor (D) and acceptor (A), respectively, and ϒ is the correlation coefficient, which incorporates the difference in quantum yields of donor and acceptor and detection efficiencies of the donor and acceptor channels. FRET trajectories were further compiled into a FRET histogram, which provides information about the distribution of Env(-) molecules among the conformational states. The state distributions in the FRET histogram were then fitted to the sum of three Gaussian distributions (based on previously identified FRET trajectories) (42,81,121) in Matlab, and the occupancy of each state was further obtained from the area under each Gaussian distribution.
Immunoprecipitation of cell-surface Env
One day prior to transfection, HOS cells were seeded in 6-well plates (6 x 105 cells/well). The cells were transfected the next day with 0.4 µg of the pSVIIIenv plasmid expressing the wild-type HIV-1JR-FL Env and 0.05 µg of a Tat-expressing plasmid. Two days later, the cells were washed twice with blocking buffer (1×PBS with 5% FBS) and then incubated for 1 hour at 4°C with 6 µg/µl anti-gp120 monoclonal antibody. Cells were then washed four times with blocking buffer, four times with washing buffer (140 mM NaCl, 1.8 mM CaCl2, 1 mM MgCl2 and 20 mM Tris, pH 7.5), and lysed in NP-40 buffer (0.5 % NP-40, 0.5 M NaCl and 10 mM Tris, pH 7.5) for 5 min at 4°C with gentle agitation. Lysates were cleared by centrifugation at 15,000 x g for 30 min at 4°C. Antibody-bound Env was precipitated using Protein A-Sepharose beads and analyzed by SDS-PAGE and Western blotting with a horseradish peroxidase (HRP)-conjugated rabbit anti-gp120 polyclonal serum.
Cell-based enzyme-linked immunosorbent assay (ELISA)
CHO cells expressing HIV-1JR-FL Env(-) were induced with 1 µg/ml doxycycline with or without 10 µM BMS-806. Fifteen to twenty-four hours later, the cells were washed twice with washing buffer #1 (20 mM Hepes, pH 7.5, 1.8 mM CaCl2, 1 mM MgCl2, 140 mM NaCl), and crosslinked with 5 mM BS3 or incubated in buffer without crosslinker. Forty-five minutes later, the cells were quenched with quench buffer (50 mM Tris, pH 8.0, 1.8 mM CaCl2, 1 mM MgCl2, 140 mM NaCl). The cells were blocked with a blocking buffer (35 mg/ml BSA, 10 mg/ml non-fat dry milk, 1.8 mM CaCl2, 1 mM MgCl2, 25 mM Tris, pH 7.5 and 140 mM NaCl) and incubated with the indicated primary antibody in blocking buffer for 30 min at 37°C. Cells were then washed three times with blocking buffer and three times with washing buffer #2 (140 mM NaCl, 1.8 mM CaCl2, 1 mM MgCl2 and 20 mM Tris, pH 7.5) and re-blocked with the blocking buffer. A horseradish peroxidase (HRP)-conjugated antibody specific for the Fc region of human IgG was then incubated with the samples for 45 min at room temperature. Cells were washed three times with blocking buffer and three times with washing buffer #2. HRP enzyme activity was determined after addition of 35 µl per well of a 1:1 mix of Western Lightning oxidizing and luminal reagents (Perkin Elmer Life Sciences) supplemented with 150 mM NaCl. Light emission was measured with a Mithras LB940 luminometer (Berthold Technologies).
Analysis of Env(-) glycoforms in BMS-806-treated cells
CHO cells expressing HIV-1JR-FL Env(-) were treated with 1 µM BMS-806 or an equivalent volume of the carrier, DMSO. After 18-24 h of culture, the cells were harvested and lysed in homogenization buffer (see above) and treated with different glycosidases following the manufacturer’s instructions. The lysates were analyzed by Western blotting with a horseradish peroxidase (HRP)-conjugated anti-HIV-1 gp120 antibody, as described above.
Analysis of Env glycopeptides
The sample preparation and mass spectrometric analysis of Env(-) glycopeptides has been described previously (39, 40), and no changes were made to the procedure for the current analysis. Briefly, the Env(-) glycoprotein was denatured with urea, reduced with TCEP, alkylated with iodoacetamide, and quenched with dithiothreitol. The protein was then buffer exchanged, digested with trypsin alone or with a combination of trypsin and chymotrypsin, generating glycopeptides.
The glycopeptides were analyzed by LC-MS on an LTQ-Orbitrap Velos Pro (Thermo Scientific) mass spectrometer equipped with ETD (electron transfer dissociation) that was coupled to an Acquity Ultra Performance Liquid Chromatography (UPLC) system (Waters). About 35 micromoles of digest was separated by reverse phase HPLC using a multistep gradient, on a C18 PepMap™ 300 column. The mass spectrometric analysis was performed using data-dependent scanning, alternating a high-resolution scan (30,000 at m/z 400), followed by ETD and collision-induced dissociation (CID) data of the five most intense ions. The glycopeptides were identified in the raw data files using a combination of freely available glycopeptide analysis software and expert identification, as described previously (39).
Analysis of A549-Gag/Env cells and VLPs treated with BMS-806
To analyze the effect of BMS-806 on the processing of the wild-type HIV-1AD8 Env, 150-mm dishes of 30-40% confluent A549-Gag/Env cells were seeded and, on the following day, treated with 2 µg/ml doxycycline. At the same time, 10 µM BMS-806 was added. Approximately 72 hours after induction, cell lysates and medium were harvested. To prepare VLPs, the culture medium was cleared by low-speed centrifugation (500 x g for 15 minutes at 4°C) and 0.45-µm filtration. VLPs were pelleted by centrifugation at 100,000 x g for one hour at 4°C. The resuspended VLP preparation was clarified by low-speed centrifugation.
Env solubilized from cell lysates and VLPs was denatured by boiling in denaturing buffer (New England Biolabs) for 10 minutes. Samples were mock-treated or treated with PNGaseF or Endo Hf (New England Biolabs) for 1.5 hours according to the manufacturer’s protocol. The treated samples were then analyzed by reducing SDS-PAGE and Western blotting.
Cryo-EM sample preparation
A 3-μl drop of 0.3 mg/ml Env(-) protein solution was applied to a glow-discharged C-flat grid (R1/1 and R1.2/1.3, 400 Mesh, Protochips, CA, USA), blotted for 2 sec, then plunged into liquid ethane and flash-frozen using an FEI Vitrobot Mark IV.
Cryo-EM data collection
Cryo-EM grids were first visually screened on a Tecnai Arctica transmission electron microscope (FEI) operating at 200 kV. Qualified grids were then imaged in a 200-kV FEI Tecnai Arctica microscope, equipped with an Autoloader, at a nominal magnification of 210,000 times, and in a 300-kV Titan Krios electron microscope (FEI) equipped with a Gatan BioQuantum energy filter, at a nominal magnification of 105,000 times, operating at 300 kV. Coma-free alignment and astigmatism were manually optimized prior to data collection. Cryo-EM data from the 200-kV Arctica microscope were collected semi-automatically by Leginon version 3.1 (130, 131) on the Gatan K2 Summit direct electron detector camera (Gatan Inc., CA, USA) in a super-resolution counting mode, with a dose rate of 8 electrons/pixel/second and an accumulated dose of 50 electrons/Å2 over 38 frames per movie. The calibrated physical pixel size and the super-resolution pixel size were 1.52 Å and 0.76 Å, respectively. The defocus for data collection was set in the range of -1.0 to -3.0 µm. A total of 12,440 movies were collected on the 200-kV Arctica microscope without tilting the stage, from which 10,299 movies were selected for further data analysis after screening and inspection of data quality.
Cryo-EM data from the 300-kV Krios microscope, including both zero-tilted and 45°-tilted images, were collected on the K2 Summit direct electron detector (Gatan) at a pixel size of 0.685 Å in a super-resolution counting mode, with an accumulated dose of ∼53 electrons/Å2 across 40 frames per movie. With defocus ranging from -1.0 to -2.7 μm, a total of 10,929 movies were acquired across three sessions.
Zero-tilted and 45°-tilted images were collected by a semi-automatic process set up in Serial EM (132), which is compatible with customized scripts. For the collection of zero-tilted movies, the process normally involved the following steps: Square selection and focusing, hole selection, serial local focusing and data acquisition. In the final step, precise adjustment of the defocus was conducted each time before recording movies for a new group of holes. However, for the collection of tilted movies, precise adjustment of the defocus was performed for all holes in the first place, followed by an extra coordinate transformation for the x-axis and y-axis. Tilted movies were then recorded serially with the new defocus and coordinates.
Cryo-EM data processing and analysis
The raw movie frames of each dataset were first corrected for their gain reference and each movie was used to generate a micrograph that was corrected for sample movement and drift with the MotionCor2 program (133) at a super-resolution pixel size (0.76 Å for the 200-kV dataset, 0.685 Å for the 300-kV dataset). These drift-corrected micrographs were used for the determination of the actual defocus of each micrograph with the CTFFind4 (134) and Gctf (135) programs. Icy or damaged micrographs were removed through manual per-image screening.
For the 200-kV dataset, using DeepEM, a deep learning-based particle extraction program that we developed (136), 1,436,424 particles of Env(-) were automatically selected in a template-free fashion. All 2D and 3D classifications were done at a pixel size of 1.52 Å. After the first round of reference-free 2D classification, bad particles were rejected upon inspection of class-average quality, which left 1,366,095 particles. The initial model, low-pass filtered to 60 Å, was used as the input reference to conduct unsupervised 3D classification into 5 classes with C3 symmetry, using an angular sampling of 7.5° and a regularization parameter T of 4. Iterative 3D classification in RELION (137) and ROME (138) resulted in a 3D class of 121,979 particles that reached a resolution of 5.5 Å (gold-standard FSC at 0.143 cutoff) after refinement, with imposition of C3 symmetry. More details of this preliminary, intermediate analysis were described in an online bioRxiv preprint (95).
For the zero-tilt 300-kV dataset, micrographs without dose-weighting were used by Gctf (135) to estimate the global CTF parameters; for the 45°-tilt dataset, particles were first picked by a program based on a VGG deep neural network improved from the DeepEM algorithm design (136). The coordinates were then applied for local CTF estimation in Gctf (135). We found that for most of 45°-tilted micrographs, limiting the resolution range used for CTF determination in Fourier space improved the accuracy of the resulting CTF parameters. This was realized by including the variables “local_resL” and “local_resH” in the Gctf (135) command. Automatic picking followed by manual examination yielded 1,941,541 particles of the HIV-1JR-FL Env(-) trimers, with 785,844 zero-tilted and 1,155,697 tilted particles.
All 2D and 3D classifications of the particles from the 300-kV datasets were conducted with dose-weighted micrographs generated by MotionCor2 (133). Particles were stacked at 2.74 Å/pixel using a box size of 84*84 for initial sorting. Two rounds of reference-free 2D classification were performed in RELION 3.0 (137), followed by one round in ROME (138), which combines maximum likelihood-based image alignment and statistical manifold learning-based classification. Bad particles were rejected upon inspection of the class average’s quality after each round of 2D classification, leaving 572,205 particles for 3D refinement. The initial model was generated in RELION 3.0 (137) using particles from diversely oriented 2D classes, and was low-pass filtered to 60 Å.
3D classification and refinement of the 300-kV datatset were performed in RELION 3.0 (137), as summarized in Table 3. In the first round of unsupervised 3D classification, the Healpix order was enhanced from 2 to 3 at the 20th iteration. To prevent tilted particles from being separated as a sole 3D class, the resolution limit to restrict the probability calculation was set at 15 Å in the preceding 20 iterations and 10 Å in the posterior iterations. The 2nd round of 3D classification retained the same parameters except that K (the number of classes) was changed to 6. The 3rd round of 3D classification was performed by local searching (σ=4, meaning that the standard deviation of the Euler angles equals 4 times the Healpix order) to discard amorphous particles. Particles with the correct size and detailed secondary structures were selected and binned two-fold into 1.37 Å/pixel for further refinement. The selected 278,582 particles were first aligned together by auto-refinement, and then were classified into 12 classes within a soft, global mask without alignment. Particles from 5 classes with complete domain constitution were sorted out and used for per-particle CTF refinement in RELION 3.0 (137). Imposed with updated CTF correction, the sorted stacks were classified with local searching into two major classes.
As observed in Chimera (139), the distribution of particles concentrated in the top-view orientation for both maps, leading to anisotropy of the final resolution. Therefore, we retrieved the tilt-view particles excluded by previous rounds of 3D classification, and combined them with particles from the two classes. This was accomplished by several rounds of screening satisfying classes from the results of deep 2D classification in ROME (138). The new particle dataset, containing 171,342 zero-tilted particles and 157,607 45°-tilted particles, was used for one round of 3D classification under global searching with Healpix order 2. Particles from 3 of the 4 classes were identified as HIV-1JR-FL Env(-) trimers with improved isotropic resolution; these 284,664 particles were combined for the next round of 3D classification. Another round of 3D classification using the same parameters except for K=3 was performed to exclude particles with poor quality. The principal class consisting of 92% of this round’s particles was reserved.
For elaborate 3D classification, we adopted a hierarchical enhancement of Healpix order in the next 9 rounds (Table 3): Sorted particles from the previous round of 3D classification were used for auto-refinement followed by classification into four classes with local searching under a Healpix order of 4. In every round, this process produced a major class consistent with the structure of the conventional Env trimer and consisting of more than 80% of the input particles, while the other classes appeared in incomplete form. Therefore, this major class of particles was used for auto-refinement and was chosen as input for next round of 3D classification. This classification-selection-refinement-classification process was iterated four times, using different K (class number) values and the same Healpix order 4, until the result demonstrated more than one principal class. C1 symmetry was imposed throughout all these unsupervised 3D classifications. In the last two rounds, we enhanced the Healpix order to 5 to perform local-searching 3D classification again, and finally obtained five classes. Four of these classes, consisting of 96% of the input particles, exhibited different degrees of asymmetry. By carefully comparing their features, two classes with similar topology were designated State-U1 while the other two classes were designated State-U2, containing 123,372 and 55,571 particles respectively. The last round of auto-refinement for the U1 and U2 datasets was done in RELION 3.0 (137), applied with a soft-edged global mask when it fell into local searching. According to the in-plane shift and Euler angles of each particle from the final refinement, we reconstructed the two half-maps of each state at a super-resolution counting mode with a pixel size of 0.685 Å. The overall masked resolutions for the reconstructed maps of State-U1 and State-U2 were 4.1 Å and 4.7 Å respectively, measured by the gold-standard FSC at 0.143-cutoff.
Atomic model building and refinement
The symmetric structure of the HIV-1BG505 sgp140 SOSIP.664 trimer with three BMS-806 molecules bound (PDB: 6MTJ) (118) and the asymmetric structure of the HIV-1JR-FL EnvΔCT glycoprotein bound to PGT151 Fabs (PDB: 5FUU) (105) were used as reference models to build a U1 structure. The template structures were docked in Coot (140), and then main-chain and side-chain fitting was improved manually to generate the starting coordinate file. The fitting of the U1 model was further improved by real_space_refinement with secondary structure restraints in Phenix (141). Glycans of U1 were manually refined in Coot (140) with “Glycan” model, using 5FUU as a reference. The U1 model was used as a whole to perform rigid-body fitting into the U2 density. Structural comparison was conducted in Pymol (142) and Chimera (139). All figures of the structures were produced in Pymol (142).
Accession numbers
The cryo-EM reconstructions of states U1 and U2 reported in this paper have been deposited in the Electron Microscopy Data Bank under accession numbers EMD-XXXX and EMD-XXXX, respectively. The models of U1 and U2 have been deposited in the Protein Data Bank under ID codes XXXX and XXXX. The cryo-EM raw data, including the motion-corrected micrographs and the particle stacks of U1 and U2 used for final refinement, have been deposited into the Electron Microscopy Pilot Image Archive (www.ebi.ad.uk/emdb/ampiar) under accession no. EMPIAR-10163.
Author contributions
J.S. and Y.M. conceived this study. H.Ding and J.C.K. prepared the Env(-)-expressing CHO cells and the A549-Gag/Env cells. S.Z. and R.T.S. analyzed Env(-) antigenicity and established a purification scheme for the Env(-) protein. S.Z. and W.L.W. screened the samples for optimization of cryo-EM imaging. W.L.W. conducted cryo-electron microscopy, collected all data and preprocessed the data. K.W. and S.C. performed data analysis and refined the maps. K.W., S.Z., S.C. and Y.M. built the structural models. E.P.G., S.Z. and H.Desaire analyzed the Env(-) glycans. M.L. and S.Z. conducted smFRET experiments. H.T.N. studied the effect of BMS-806 on the processing of wild-type Env. Y.M. and J.S. wrote the manuscript. All authors contributed to data analysis and manuscript preparation.
Acknowledgments
This work was funded in part by NIH grants AI125093 (H. Desaire), AI93256, AI100645, AI125093, AI145547, AI127767, AI150471/GM56550 and AI124982 (J.S.), by an Intel academic grant (Y.M.), by grants from the Natural Science Foundation of Beijing Municipality grant No. Z180016/Z18J008 and the National Natural Science Foundation of China grant No. 11774012 (Y.M.), and by a gift from the late William F. McCarty-Cooper. M.L. was supported by a grant (109998-67-RKVA) from the American Foundation for AIDS Research (amfAR). The research was also supported by the Basic Research Core of the University of Alabama, Birmingham Center for AIDS Research (NIH grant AI027767). The cryo-EM experiments were performed in part at the Center for Nanoscale Systems at Harvard University, a member of the National Nanotechnology Coordinated Infrastructure Network (NNCI), which is supported by the National Science Foundation under NSF award no. 1541959. The cryo-EM facility was funded through the NIH grant AI100645, Center for HIV/AIDS Vaccine Immunology and Immunogen Design (CHAVI-ID). The data processing was performed in part in the Sullivan cluster, which is supported by a gift from Mr. and Mrs. Daniel J. Sullivan, Jr.
Footnotes
Authorship order was determined by mutual agreement.
REFERENCES
- 1.↵
- 2.
- 3.↵
- 4.↵
- 5.
- 6.
- 7.
- 8.
- 9.
- 10.
- 11.
- 12.
- 13.
- 14.↵
- 15.
- 16.↵
- 17.↵
- 18.
- 19.
- 20.↵
- 21.↵
- 22.
- 23.
- 24.↵
- 25.↵
- 26.
- 27.
- 28.
- 29.
- 30.
- 31.
- 32.
- 33.
- 34.
- 35.
- 36.
- 37.
- 38.
- 39.↵
- 40.↵
- 41.↵
- 42.↵
- 43.
- 44.
- 45.↵
- 46.↵
- 47.↵
- 48.↵
- 49.↵
- 50.↵
- 51.
- 52.
- 53.↵
- 54.↵
- 55.↵
- 56.
- 57.
- 58.↵
- 59.↵
- 60.
- 61.
- 62.↵
- 63.↵
- 64.
- 65.↵
- 66.↵
- 67.↵
- 68.
- 69.
- 70.↵
- 71.↵
- 72.↵
- 73.↵
- 74.
- 75.
- 76.
- 77.
- 78.↵
- 79.↵
- 80.↵
- 81.↵
- 82.↵
- 83.↵
- 84.
- 85.
- 86.
- 87.
- 88.
- 89.
- 90.
- 91.↵
- 92.↵
- 93.
- 94.↵
- 95.↵
- 96.↵
- 97.↵
- 98.
- 99.
- 100.
- 101.
- 102.
- 103.
- 104.↵
- 105.↵
- 106.
- 107.
- 108.↵
- 109.↵
- 110.
- 111.↵
- 112.↵
- 113.
- 114.
- 115.↵
- 116.↵
- 117.↵
- 118.↵
- 119.↵
- 120.↵
- 121.↵
- 122.↵
- 123.↵
- 124.↵
- 125.↵
- 126.↵
- 127.↵
- 128.↵
- 129.↵
- 130.↵
- 131.↵
- 132.↵
- 133.↵
- 134.↵
- 135.↵
- 136.↵
- 137.↵
- 138.↵
- 139.↵
- 140.↵
- 141.↵
- 142.↵
- 143.




