

Abstract
Comparative studies of reproductive isolation (RI) by Coyne and Orr (1, 2) and others indicate a significant role for reinforcing natural selection in Drosophila speciation. The reinforcement hypothesis predicts increased prezygotic, but not postzygotic, RI between sympatric species pairs in response to maladaptive hybridization. We revisit this hypothesis and others using additional Drosophila, Lepidopteran, and toad (Bufo) data. In contrast to the predictions of reinforcement, we find increased premating and postzygotic RI between sympatric Drosophila species, including between recently diverged species pairs, as defined by Coyne and Orr (1) and others (i.e., DNei < 0.5). However, at slightly lower divergence thresholds increased postzygotic RI in sympatry is not statistically significant, while increased premating RI is, generally in agreement with the predictions of reinforcement. While premating data are unavailable, postzygotic RI is also increased between sympatric Lepidopteran and toad (Bufo) species. We find only modest support for “concordant asymmetries” in premating and postzygotic RI between sympatric Drosophila, described by others as uniquely supporting reinforcement. Finally, the proportion of geographic range overlap shared by species is positively associated with the magnitude of premating RI as predicted by reinforcement, but it is also positively associated with postzygotic RI, which cannot be explained by reinforcement. Taken together, our results demonstrate that comparisons of premating and postzygotic RI in sympatry depend greatly on divergence time, and suggest that fusion, extinction, and/or other mechanisms must combine with reinforcement to generate these patterns.
Significance Statement Understanding mechanisms underlying the origin of species remains a central goal of biology. By combining laboratory estimates of premating and postzygotic reproductive isolation (RI) with phylogenetic hypotheses, geographic range data, and genetic divergence estimates, the fundamental meta-analyses of Coyne and Orr provided compelling support that reinforcement contributes significantly to Drosophila speciation. Using additional Drosophila data, we demonstrate increased premating and postzygotic RI in sympatry. Postzygotic RI is also increased in sympatry between Lepidopteran and toad species. We also find only modest support for patterns previously described as uniquely supporting reinforcement. Together, our analyses demonstrate that comparisons of premating and postzygotic RI depend on divergence time, and suggest that fusion, extinction, and/or other mechanisms combine with reinforcement to produce these patterns.
Introduction
Speciation, the process in which one species splits into two, results from the accumulation of genetic differences and the evolution of reproductive isolation (RI) (3, 4). In broad terms, barriers to gene flow can be classified based on when they occur in the reproductive cycle, commonly in relation to the generation of a fertilized zygote (5, 6). These barriers fall into two general categories: prezygotic and postzygotic RI. Prezygotic RI includes all ecological, behavioral, mechanical, and gametic incompatibilities that occur before a zygote is formed (5), with most focus placed on premating traits (e.g., mate discrimination). Postzygotic barriers occur after a hybrid zygote is formed and include phenotypes as extreme as hybrid sterility and inviability (7, 8), but also more nuanced traits, such as hybrid behavioral defects (9–11) or delays in development (12–14). Coyne and Orr [(1, 2); hereafter “C&O”] completed the first in-depth meta-analyses of speciation that combined estimates of genetic divergence, premating RI, and hybrid viability and fertility in Drosophila with phylogenetic and geographic information. Their work demonstrated that both premating and postzygotic RI become stronger as the genetic distance between species increases, and that premating RI accumulates faster in sympatry.
Comparative analyses of speciation have greatly influenced our understanding of speciation (15). Analyses similar to those of C&O, and in some cases similar datasets, have revealed seminal aspects of how speciation proceeds. Studies of the rates of evolution of premating and postmating RI between Drosophila species pairs have enabled us to understand the role of sex chromosomes in speciation (16–18), and the potential drivers of speciation (19–22). Indeed, C&O sparked a cottage industry to study RI in different groups which has demonstrated that the monotonic increase in RI as divergence accrues is not specific to Drosophila, occurring in virtually all studied taxa (23, 24).
Arguably the most influential finding of C&O was that in contrast to postzygotic RI, mate discrimination is more pronounced between sympatric than between allopatric species pairs of the same age. These results were taken as evidence that natural selection favors the evolution of premating RI when allopatric species come into secondary contact. In this process, coined reinforcement, premating RI is strengthened as a byproduct of natural selection against hybridization (25–27). These analyses and others led to the conclusion that reinforcement is common during speciation (16, 28). As C&O (1, 2) noted, “any factor that reduces gene flow should inhibit fusion or extinction”. Observing increased prezygotic and postzygotic RI in sympatry, particularly for recently diverged species, would support a fusion and/or extinction hypothesis since only populations separated by strong mating discrimination may persist (1, 29, 30).
Additional premating and postzygotic RI estimates from Drosophila, in combination with estimates of postzygotic RI between Lepidopteran and between toad (Bufo) species, provide the opportunity to revisit and test the predictions of reinforcement and related hypotheses. First, and in contrast to C&O, we find increased premating and postzygotic RI between recently diverged sympatric relative to allopatric Drosophila pairs using their divergence threshold (DNei < 0.5); but at slightly lower thresholds increased postzygotic RI in sympatry is not statistically significant, while increased premating RI is, in agreement with the predictions of reinforcement. Second, while butterfly and toad data do not include estimates of premating RI, we also observe increased postzygotic RI in sympatry for these taxa, including for the most recently diverged toads. Third, our expanded Drosophila analyses find only modest evidence for stronger premating RI among the more costly reciprocal matings. Fourth, premating RI is positively associated with the proportion of geographic range overlap, but so is postzygotic RI, which cannot be explained by reinforcement. Our results suggest that fusion, extinction, and/or other mechanisms combine with reinforcement to generate the patterns we observe.
Results
Increased premating and postzygotic RI in sympatry
RI increases as species diverge, and several efforts have reported a negative correlation between parental species divergence and both the strength of mating discrimination (1, 2, 31) and the fitness of resulting hybrids (e.g., (32–34), reviewed in (15, 23, 24, 35)). We fit three types of models to study the evolution of RI between diverging species. For both premating and postzygotic RI in Drosophila, four-parameter logistic (henceforth abbreviated 4PL) models fit the data better than do linear or logistic models, for both allopatric and sympatric species pairs (Table S1). The 4PL model was also the best fit for postzygotic RI in toads and Lepidopterans, for both allopatric and sympatric species pairs. The 4PL model meets the biological expectation of a waiting time for mutations that generate either hybrid incompatibility (36–38) or behavioral isolation (31), followed by a rapid accumulation of RI that eventually asymptotes. Please note that even though Nei’s D, our metric of genetic differentiation between Drosophila parental species, is not a perfect predictor of the extent of differentiation, it represents a good proxy (Figure S1 in (9)).
Next, we re-assessed support for the hypothesis that reinforcement has driven the evolution of premating RI in sympatry for Drosophila. We leveraged the information provided by our model fits and compared the values of the lower asymptote, the upper asymptote, and the inflection point for sympatric and allopatric species pairs, for both premating and postzygotic RI (Figure 1A and B, respectively). Table S2 shows pairwise comparisons for the regression parameters for allopatric and sympatric species. To determine whether isolation evolved faster in sympatric species, we compared c, the inflection point of the 4PL model, and a proxy for how fast RI completes, for regressions using either sympatric or allopatric species. Consistent with previous studies (1, 2) premating RI between sympatric Drosophila species pairs reaches c earlier than allopatric species pairs (cSympatric-Drosophila-premating = 0.059; cAllopatric-Drosophila-premating = 0.318; Wilcoxon rank sum test with continuity correction: W = 978,080, P < 1 × 10-10; Figure 1C). The rate of increase once the inflection point is reached (Hill’s slope, or b) is higher for allopatric pairs which eventually leads to similar d (asymptote) values. In Figure S1, we present the other regression parameters: a, b, and d. Surprisingly, we observed that for postzygotic RI, c is lower and b is higher for sympatric than for allopatric pairs, indicating that postzygotic RI also increases faster as divergence accrues between sympatric species (cSympatric-Drosophila-postzygotic = 0.439; cAllopatric-Drosophila-postzygotic = 0.751; Wilcoxon rank sum test with continuity correction: W = 993,050, P < 1 × 10-10; Figure 1D). These results suggest that both premating and postzygotic RI increase faster between sympatric than between allopatric species pairs. We find similar results for both premating and postzygotic RI in Drosophila after using stringent subsamplings at the species-group level (Premating: cSympatric-Drosophila = 0.141; CAllopatric-Drosophila = 0.394, W = 16,917; P = 3.654 × 10-11; Postzygotic: cSympatric-Drosophila = 0.38; cAllopatric-Drosophila = 0.610, W = 8,342; P = 3.613 × 10-10; Figure S2).

Four-parameter logistic models for the evolution of premating RI (A) and postzygotic RI (B) between sympatric (blue) and allopatric (red) Drosophila species pairs. Semitransparent lines show 1,000 bootstrapped distributions. For both types of barriers, the inflection point (c) is at lower genetic distances for sympatric than for allopatric species pairs (premating: C; postzygotic: D). The vertical lines show the mean value of the bootstrap coefficients. Figure S1 presents the other three parameters for each of the two models.
Next, we fit a phylogenetically informed linear regression. The models included an effect for geographic origin and an interaction between origin and genetic distance. If RI evolves similarly in sympatry and allopatry, as expected for postzygotic RI under a scenario of pure-reinforcement driving the evolution of strengthened RI in sympatry, then effects of both origin and the interaction should be negligible. For premating RI, we found that sympatric species display generally stronger premating RI than do allopatric species (95% CI=[0.245, 0.464], P < 0.001), but the increase of premating RI is slower in sympatric species (95% CI = [−0.323, 0.058], P = 0.014; Table S3). For postzygotic RI, we found that sympatric and allopatric species have a similar level of postzygotic RI (95% CI = [−0.121, 0.198], P = 0.660), but that postzygotic RI increases faster with genetic distance in sympatry (95%CI = [0.138, 0.699], P = 0.002; Table S3). These results are consistent with previous reports of stronger RI in sympatry using a linear model of accumulation of premating and postzygotic RI (39). They are also consistent with the results from the uncorrected datasets, demonstrating that in Drosophila premating and postzygotic RI both evolve faster in sympatry.
While premating data are not available, we also studied these same parameters for postzygotic RI between Lepidopteran species and Bufo toads (Figure 2). The 4PL regression results for these taxa are similar to those in Drosophila (Figure 2A and B). The inflection point is lower for sympatric than for allopatric species pairs for both Lepidopterans (cSympatric-Lepidopterans = 0.686; cAllopatric-Lepidopterans = 0.834, W = 89,102, P < 1 × 10-10; Figure 2C) and BufO toads (cSympatric-Toads = 0.028; cAllopatric-Toads = 0.038, W = 130,280, P < 1 × 10-10; Figure 2D). For Lepidopterans, b is much higher for sympatric than for allopatric species pairs (bSympatric-Lepidopterans = 242.1179; bAllopatric-Lepidopterans = 7.321, W = 130,280, P < 1 × 10-10; Figure S3), which reflects that substantial postzygotic RI is rare between allopatric species pairs even in the most divergent crosses (i.e., Papilio xuthus × P. glaucus being an exception, Nei’s D = 1.161). Subsampling by genus produces a similar result. While c in sympatric species is similar to the complete dataset (cSympatric-Lepidopterans = 0.887), b is lower but positive (bSympatric-Lepidopterans = 5.801). Neither of these two parameters could be calculated for allopatric species because there was no increase in the magnitude of postzygotic RI over genetic distance for these species, and the 4PL regressions did not converge.
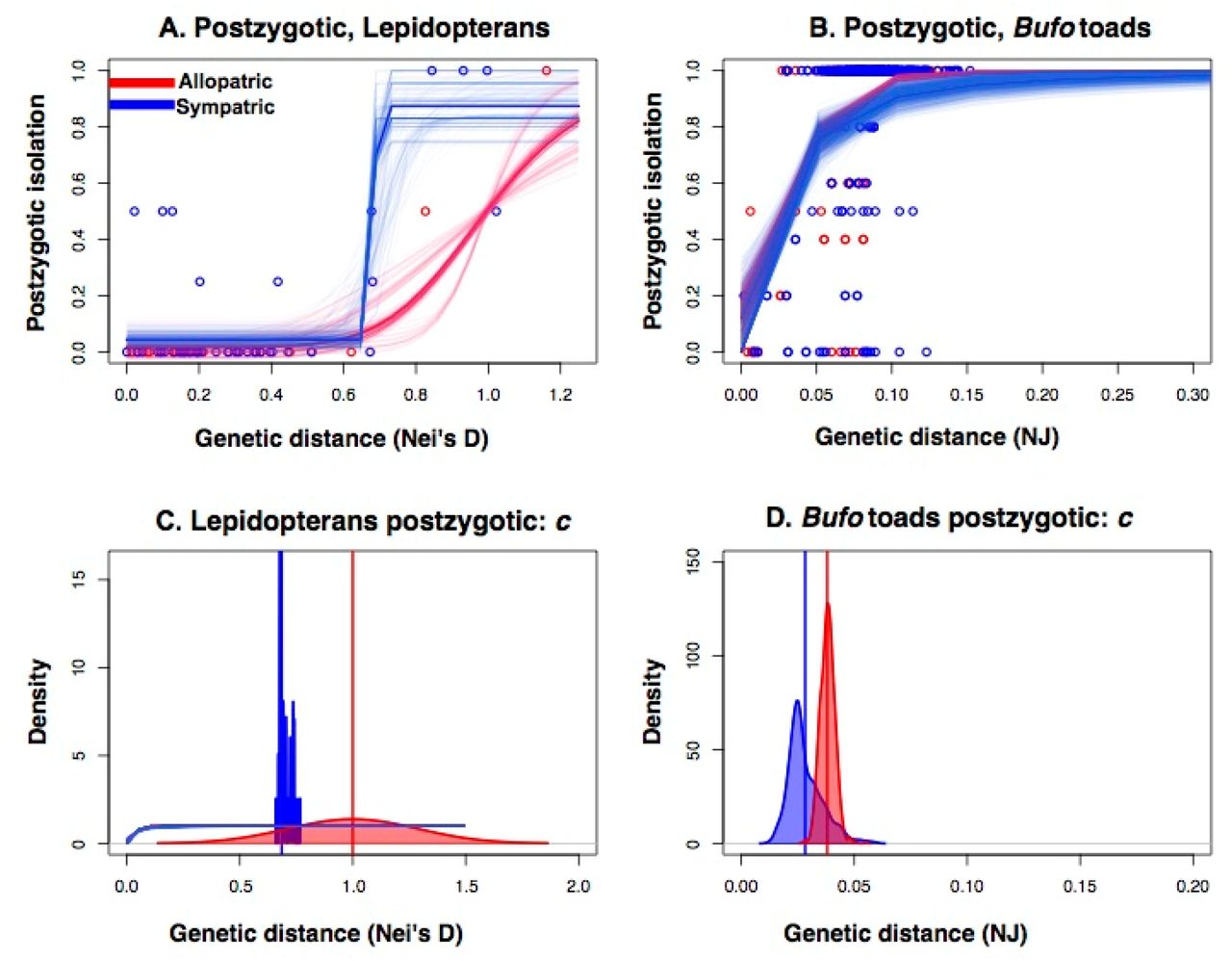
Four-parameter logistic models for the evolution of postzygotic RI between sympatric (blue) and allopatric (red) Lepidopteran (A) and Bufo (B) species pairs. Semitransparent lines show 1,000 bootstrapped distributions. For both barriers, the inflection point (c) is at lower genetic distances for sympatric than for allopatric species pairs (Lepidopterans: C; Bufo: D). The vertical lines show the mean value of the bootstrap coefficients. Figure S3 presents the other three parameters for each of the two models. Genetic distance is measured as Nei’s D for Lepidopterans and from a neighbor-joining tree for Bufo (39).
For Bufo, b is higher for allopatric than for sympatric species pairs (bSympatric-Toads = 1.734; bAllopatric-Toads = 2.899, W = 26,916, P < 1 × 10-10; Figure S3), which is consistent with the earlier inflection point for sympatric pairs and the similar asymptote of both cases (d = 1, Figure S3). A subsampling by subgenus was not possible for Bufo (See Methods).
We followed up the analyses in these two groups with a phylogenetically corrected linear regression. In the case of Lepidopterans, sympatric species have slightly higher postzygotic RI compared with allopatric species (95% CI = [−0.018, 0.257], P = 0.076), similar to premating RI in Drosophila. The increase of inviability with genetic distance is faster for sympatric species than for allopatric species (−95% CI = [0.674, −0.094], P = 0.014; Table S4). In Bufo, we find that postzygotic RI is generally lower in sympatric species (95% CI = [−0.272, −0.032], P = 0.006), but that RI increases faster with genetic distance between sympatric species pairs (95% CI = [0.486, 3.637], P = 0.012; Table S4). These regressions are more limited than 4PL regressions, as linear regressions do not differentiate between the inflection point (c) and the rate of increase with genetic distance (b). Nonetheless, these results are consistent with the 4PL results in that the rate of evolution of postzygotic RI is not equivalent in allopatric and sympatric species.
In sum, our results indicate that across the full range of divergence both premating and postzygotic RI are stronger between sympatric than allopatric species for three different animal taxa. Because analysis of RI across the full range of divergence may conflate conditions for co-occurrence of reproductively isolated lineages with the conditions that accelerate speciation, we next assessed RI for the most recently diverged species.
Comparisons of RI in sympatry depend on divergence time
We first restricted our dataset to recently diverged Drosophila pairs as defined by C&O (DNei < 0.5) to test if premating, but not postzygotic, RI is strengthened at earlier stages of divergence. Both premating (W = 479.5; P < 0.0001) and postzygotic (W = 902.5, P = 0.039) RI are stronger in sympatry using the C&O divergence threshold for young species. However, this result is highly contingent on the arbitrary choice of how to define recently diverged species, such that with a slightly lower threshold (DNei = 0.47) the strength of postzygotic RI in sympatry and allopatry does not differ (W = 881.5, P = 0.05; Figure S4). In contrast, postzygotic RI between toads is always significantly increased in sympatry relative to allopatry (Table S5). Fitting 4PL regressions to the most recently diverged Drosophila (DNei < 0.5) and Bufo pairs (NJ distance < 0.05; Figure S5) indicates that the inflection point of the regression occurs earlier for sympatric than for allopatric Drosophila (csympatric = 0.366; callopatric = 0.468; W = 812,777; P < 0.0001) and Bufo (csympatric = 0.029; callopatric > 0.5) pairs. Similar analyses were not possible for Lepidopterans because the 4PL regressions did not converge when the dataset included only species pairs with Nei’s D < 0.5.
In sum, our results indicate that comparisons of premating and postzygotic RI between sympatric Drosophila depend greatly on divergence time; and while comparisons that include only the most recently diverged Drosophila species support the predictions of reinforcement, elevated postzygotic RI between sympatric Drosophila and Bufo at relatively early stages of divergence implies other mechanisms also contribute to these patterns.
Proxies of the cost of hybridization are correlated with the strength of premating and postzygotic RI in Drosophila
We next tested three additional predictions of reinforcement as the primary driver of strengthened premating RI in sympatry. First, if premating RI evolves via reinforcement, then the selection pressure to evolve premating RI might be a function of the cost of hybridization (40, 41). It then follows that if the strength of RI differs between reciprocal crosses, the cross direction with stronger postzygotic RI, and thus a higher cost to hybridization, should also display stronger premating RI (42). A pattern of ‘concordant asymmetries’ (i.e., asymmetries in premating and postzygotic RI in the same direction) has been observed in selected sympatric Drosophila species (16 species pairs), but not in allopatric ones (20 species pairs), which is argued to uniquely support the prevalence of reinforcement ((42) but see (16)).
We extend this argument to the strength of asymmetries (i.e., cross directions with strong postzygotic RI should show relatively strong premating RI), testing for a positive correlation between the magnitude of premating and postzygotic RI asymmetries in Drosophila. Across all of the data, the strength of the asymmetry in postzygotic RI predicts the strength of the asymmetry in premating RI in sympatry (N = 73, F1,71 = 8.037, P = 0.006), but not in allopatry (N = 91, F1,89 = 0.002, P = 0.964), but the proportion of premating RI asymmetry explained by postzygotic RI asymmetry in sympatry is low (R2 = 0.089, Figure 3A). Figure S6 shows that the association between the strength of premating and postzygotic RI asymmetries is sensitive to inclusion parameters in the study. If only cases where the magnitude of postzygotic RI is asymmetric are included, the association between the magnitude of premating and postzygotic RI asymmetries is not significant (sympatry: F1,11= 1.381; P = 0.265; allopatry: F1,16 = 1.905, P = 0.186; dashed lines in Figures 3A). A partial correlation while controlling for genetic distance shows that the magnitude of premating and postzygotic RI asymmetries are only weakly correlated (ρ = 0.192, P = 0.050). Thus, even if concordant asymmetries are taken as incontrovertible evidence for reinforcement (see below), we interpret these results as supporting that other mechanisms also contribute to strengthened RI between sympatric Drosophila species.

{kind=link}
{kind=link}
{kind=link}
A. The strength of asymmetries in premating RI is weakly correlated with the strength of asymmetries in postzygotic RI in sympatry (solid blue), but not in allopatry (solid red). When completely symmetric pairs (i.e., those where the difference in the strength of RI between reciprocal crosses equals 0) are excluded, asymmetries in premating and postzygotic RI in sympatry are no longer significantly correlated (dashed blue line). Range overlap is correlated with the strength of both premating (B) and postzygotic RI (C).
Second, because the risk of hybridization is proportional to the extent of geographic range overlap, in sympatry reinforcement should produce a positive relationship between the strength of premating RI between species and the proportion of their range overlap. This relationship should not exist for postzygotic RI since there is no impetus for reinforcement to act on it (1). Consistent with the reinforcement hypothesis and findings of previous reports (43), we find that range overlap is positively associated with premating RI in Drosophila, even after controlling for genetic distance (Linear Model: F1,255: 50.809, P = 1.044 × 10-11; Figure 3B). However, the proportion of geographic range overlap is also positively associated with the magnitude of postzygotic RI in Drosophila (F1,105 = 14.098, P = 2.85 × 10-4; Figure 3C). Table S6 shows the results for the linear models. Notably, there is no relationship between geographic range overlap and the strength of premating or postzygotic RI in species whose ranges overlap more than 5%, suggesting the Drosophila result is driven by species pairs that are mostly allopatric (Table S7). This result suggests that in some instances overlap might drive the evolution of both premating and postzygotic RI, and is thus inconsistent with the predictions of reinforcement being the exclusive driver of premating RI in sympatry.
Finally, and as C&O (1) noted, if reinforcement is responsible for the strengthened RI in sympatry, then sympatric pairs should display a range of RI values that is higher and outside the range observed between allopatric pairs because natural selection will lead to RI after secondary contact. Conversely, if differential fusion and/or extinction is responsible for the pattern of stronger RI in sympatry (29, 30), the range of RI observed among sympatric pairs should be a subset of the RI observed among allopatric pairs because species pairs with weak RI will go extinct (1). The range of RI among sympatric pairs is similar to that of allopatric pairs (i.e., both are bounded by 0 and 1), including with the Coyne and Orr (1) threshold for young species (Nei’s D <0.5; Figure S7). (Please note that the range is not equivalent for Drosophila species pairs younger than Nei’s D < 0.25 ((42); see Discussion). Sympatric and allopatric Bufo pairs show a similar range of postzygotic RI regardless of divergence, but this is not the case for Lepidopterans (Figure S8). We interpret these results as indicating that multiple mechanisms likely contribute to increased premating RI in sympatry.
Discussion
Comparative studies are a powerful tool to study how RI accrues, divergence proceeds, and speciation occurs. Groundbreaking analyses by C&O and others broadly support the hypothesis that reinforcement contributes significantly to Drosophila speciation, but as C&O (1) noted “enhanced prezygotic RI could result from a process of fusion or extinction in sympatry” because “any factor that reduces gene flow should inhibit fusion or extinction” (pg. 376). Such a mechanism could produce higher premating RI in sympatry, in the absence of reinforcement (29). We discuss several lines of evidence that indicate multiple mechanisms likely contribute to increased RI in sympatry.
First, both premating and postzygotic RI are increased in sympatry for recently diverged Drosophila species as defined by Coyne and Orr (1), but at slightly lower divergence thresholds increased postzygotic RI in sympatry is no longer statistically significant. This pattern generally supports the predictions of reinforcement, while highlighting that comparisons of RI in sympatry depend greatly on divergence time. Postzygotic RI is also increased in sympatry between Lepidopteran and between the most recently diverged Bufo species. (Premating data are not available in either system.) Elevated postzygotic RI in sympatry for relatively recently diverged pairs in three divergent taxa implies multiple mechanisms combine to produce these patterns.
Second, the absence of very recently diverged and strongly isolated allopatric Drosophila species pairs has been argued as support for reinforcement and against fusion and extinction. If differential fusion and extinction contribute to elevated premating RI in sympatry, we should also observe strongly prezygotic isolated pairs in allopatry. In contrast to this prediction, Yukilevich ((42), Figure 1) reported a paucity of very recently diverged allopatric Drosophila pairs (D < 0.25) with strong premating RI (i.e., a “Templeton gap”). While this result supports the predictions of reinforcement, we argue it must depend on species definitions. For example, multiple instances in the literature report strong assortative mating between populations of the same species (i.e., D. lacertosa: Table 2 in (44); D. yakuba: Table 4 in (45); and 14 other species in (46)). Researchers are more likely to classify strongly isolated sympatric groups as different taxonomic species since their isolation has been “tested” in secondary contact. This creates an inherent bias when making these comparisons at the early stages of divergence, producing the illusion of a gap. This bias and the existence of strongly isolated allopatric populations within species that are weakly differentiated leads us to conclude that support for the Templeton gap in Drosophila is less compelling than previously argued. Notably, including these intraspecific observations lowers the inferred c value for allopatric species pairs, but not enough to change the conclusion that premating RI accumulates faster in sympatric species (Figure S9).
Third, asymmetries in postzygotic RI and the extent of geographic range overlap between species pairs have also been proposed as support for reinforcement of premating RI (42). Consistent with previous analyses, we find that in sympatry, sides of reciprocal Drosophila crosses that produce strong postzygotic RI also produce strong premating RI; however, asymmetry in postzygotic RI explains only 9% of the variance in premating RI asymmetries. While this pattern might be explained by the predictions of reinforcement, it might also be explained by differential fusion and/or extinction. In instances where premating and postzygotic RI act jointly to prevent fusion (47), one could also expect concordant asymmetries even if reinforcement is not driving the pattern of enhanced RI in sympatry. The same pattern of concordant asymmetries could also be explained by slight asymmetries in gene flow, which could produce asymmetrical accumulation of intrinsic postzygotic RI (16). Given that we also observe a strong effect of geographic range overlap on the evolution of premating RI and postzygotic RI, these results provide further evidence that no single mechanism explains increased RI in sympatry.
What then explains our observation of stronger postzygotic RI in sympatry for three divergent taxa? One possibility is that postzygotic RI also evolves through pervasive reinforcing selection. This would explain the faster evolution of premating and postzygotic RI in sympatry and the strong effect of geographic overlap in both barriers. Reinforcement of postmating RI has been demonstrated in Drosophila (48, 49) and Neurospora (50, 51), but in all cases the reinforced barriers act prior to zygote formation (i.e., postmating-prezygotic barriers). Even though hybrid inviability has been hypothesized to potentially evolve through selection against hybrids (52), reinforcement should only occur for hybrid inviability if aborting the hybrid embryos might represent an advantage to the parents. (No similar rationale has been proposed for hybrid sterility.) Since Drosophila, Lepidopterans, and toads all show external development, there is no impetus for selection to reinforce postzygotic RI.
A second possibility is that stronger postzygotic RI in sympatry results in part from fusion and/or extinction (29, 30, 53). In cases when postzygotic RI is increased in sympatry, and specifically in cases when both premating and postzygotic RI are stronger in sympatry, differential fusion is a more likely driver of the pattern of enhanced RI in sympatry than reinforcement (1). This idea has been disfavored because C&O did not observe elevated postzygotic RI for recently diverged sympatric Drosophila species, and because of the Templeton gap (see above). Our observation of increased postzygotic RI between relatively recently diverged Drosophila, butterflies, and between recently diverged toads implies a role for fusion, extinction, and/or other mechanisms.
A final, and under-discussed possibility, is that the pattern of stronger RI in sympatry is due to gene flow which might lead to a systematic underestimation of the age of sympatric species. This will disproportionately affect recently diverged sympatric species as the extent of gene flow seems to be negatively correlated with the divergence age between the parental species (54, 55). This possibility might affect our inference of stronger postzygotic RI in sympatry and specifically create the appearance of stronger RI at low genetic distances. We argue this is an unlikely possibility. In the case of Drosophila, the vast majority of introgression is purged rapidly (i.e., within the first ten generations) after admixture (56–58). The percentage of the genome that shows evidence of gene exchange among Drosophila species pairs ranges between 0.1% and 5% (59–61). Nonetheless, the effect of introgression on point measurements of differentiation deserves a systematic treatment.
Conclusions
Research from the last three decades has revealed dozens of cases of strengthened RI in sympatry resulting from reinforcement (reviewed in (26–28)). Experimental evolution has also demonstrated that RI can evolve rapidly if selection against hybridization is strong, lending additional support to the possibility of reinforcement in nature (48, 62–65). Using an expanded dataset, our Drosophila analyses generally agree with the main finding of C&O that premating, but not postzygotic, RI is increased between the most recently diverged sympatric species pairs. However, the dependence of this result on divergence time, our observation of increased postzygotic RI between sympatric butterflies and toads, and the other lines of evidence discussed above support that fusion, extinction, and/or other mechanisms must combine to produce these patterns.
Notably, strengthened RI does not seem to be a deterministic outcome of sympatry. Glycine and Silene plant pairs show similar levels of postmating-prezygotic RI in sympatry and allopatry (66). A concerted effort to compare the prevalence of strengthened RI in sympatry, and its potential causes across divergent taxa, is sorely needed. We expect that those analyses will reveal more support that no single mechanism underlies elevated RI in sympatry. While comparative studies of RI are useful for testing hypotheses across diverse groups of organisms, the results of our analyses suggest to us that we need a better understanding of the exact mechanisms underlying the evolution of RI in sympatry.
MATERIALS AND METHODS
Datasets
Our goal was to study whether the rate of evolution of postzygotic RI is similar in sympatric and allopatric animal species pairs. We gathered seven datasets that included metrics of RI (Table S8). Five of these datasets included estimates of genetic distance and the extent of geographic range overlap (i.e., whether species pairs are sympatric or allopatric). Two of these studies have fewer than five species pairs in each category, leaving only three datasets that include systematically gathered metrics of postzygotic RI in sympatric and allopatric species (Table S8). First, we used the data on over 630 Drosophila-species pairs from Yukilevich (42), the most extensive compilation of measurements of reproductive RI in Drosophila. This compendium included all of the data from C&O (1, 2), data on postzygotic RI across Drosophila from Bock (67), and new data collected by Yukilevich (42). In total, the dataset includes 288 interspecific hybridizations with estimates of genetic distance. Of these, 140 species pairs were classified as sympatric and 148 as allopatric. The second dataset was postzygotic RI in Lepidopterans (data from (68)). The dataset includes 212 interspecific hybridizations, 68 of which have measurements of genetic distance. Of these, 52 species pairs were classified as sympatric and 16 as allopatric. The final dataset, was postzygotic RI in toads (data from (39, 69)). This dataset includes 669 interspecific hybridizations, all of which have measurements of genetic distance (calculated from a neighbor-joining tree, (39)). Ninety-five species pairs were classified as sympatric and 574 as allopatric. Only the Drosophila dataset includes estimates of premating RI. Please note that the measurement of genetic distance between toad species (derived from a neighbor-joining tree) is not equivalent to that in Drosophila and Lepidopterans (Nei’s D), so we refrain from doing among comparison groups between insects and anurans.
Model fitting
We investigated the relationship between the strength of RI (either premating or postzygotic in Drosophila, or postzygotic in Lepidopterans and toads) and the genetic distance between the parental species. First, we compared the fit of three models to the accumulation of RI with genetic distance: linear, logistic increase, and a four-parameter logistic (i.e., dose-response) curve. We fit the linear model using the lm (library ‘stats’) function in R (70) and the other two models using the nlsLM (library ‘minpack.lm’, (71)) function in R. The logistic models followed the form:
 where Isolation is either a premating or postzygotic RI metric, a is when the genetic distance is zero, and b adjusts how quickly the probability changes with a single-unit change.
where Isolation is either a premating or postzygotic RI metric, a is when the genetic distance is zero, and b adjusts how quickly the probability changes with a single-unit change.
The four-parameter logistic models had the form:
 where a is the magnitude of isolation in a given cross at the minimum genetic distance (here where Nei’s D = 0), b is the rate of increase in RI at point c, the inflection point of the decay curve, and d is the maximum RI that can be obtained at high genetic distance. This model allows for an initial period where F1 hybrids do not suffer fitness consequences (34, 38) and includes that RI must asymptote at a value no higher than 1 (i.e., more genetic changes contributing to isolation become redundant as two lineages cannot be more isolated than completely isolated). Since nonlinear logistic regression has difficulties optimizing the values for each of the four constants in the equations, we tried 10 starting values per constant and found the model with the lowest Akaike Information Criterion (AIC, (72)) with the function ‘AIC’, (library ‘stats’; (70)).
where a is the magnitude of isolation in a given cross at the minimum genetic distance (here where Nei’s D = 0), b is the rate of increase in RI at point c, the inflection point of the decay curve, and d is the maximum RI that can be obtained at high genetic distance. This model allows for an initial period where F1 hybrids do not suffer fitness consequences (34, 38) and includes that RI must asymptote at a value no higher than 1 (i.e., more genetic changes contributing to isolation become redundant as two lineages cannot be more isolated than completely isolated). Since nonlinear logistic regression has difficulties optimizing the values for each of the four constants in the equations, we tried 10 starting values per constant and found the model with the lowest Akaike Information Criterion (AIC, (72)) with the function ‘AIC’, (library ‘stats’; (70)).
To determine which of the four models best describes the relationship between isolation and genetic divergence we used AICs (72). To find the confidence intervals of the logistic regressions we bootstrapped the datasets 1,000 times using the function nlsBoot (library(‘nlstools’, (73)). Finally, we compared the confidence intervals using Wilcoxon tests of the bootstrap values for each coefficient using the function wilcox.test (library ‘stats’, (70)).
Phylogenetic independence
Ideally, our estimates of RI would be phylogenetically independent from all others. However, our Drosophila dataset contains multiple species pairs with phylogenetic relationships that are not evolutionarily, and thus might not be statistically, independent. To account for this lack of independence, we used clade-level and species-level sampling schemes for Drosophila, and a species-level sampling scheme for Bufo frogs (9, 66). For the species group-level sampling scheme in Drosophila, we sampled a single cross from within each monophyletic clade of Drosophila (affinis, ananassae, athabasca, buzzatii, melanogaster, mesophragmatica, montium, mulleri, nasuta, obscura, planitibia, pseudobscura, repleta, takahashii, virilis, and willistoni; Suvorov et al. 2020) resulting in two datasets: one allopatric and one sympatric, each composed of about 16 species pairs. We subsampled the whole dataset 1,000 times and recalculated the value of c, the inflection point, for each iteration as described above (See Model fitting). We recorded the number of iterations that did not converge (i.e., the inferred c value was outside of bounds of the range of the function, in this case larger than the maximum Nei’s D value), but restricted comparisons between cSympatric and cAllopatric to regressions that converged. We followed a similar approach for Lepidopterans subsampling by genus (Anartia, Anthocharis, Callosamia, Choristoneura, Colias, Erebia, Heliconius, Helicoverpa, Heliothis, Hyalophora, Papilio, Phyciodes, Pieris, Pontia, and Ypomeneuta). We did not use this approach for Bufo toads because a large proportion of the hybridizations involved species from different Bufo sensu lato genera (Bufo sensu stricto, Sclerophrys, Schismaderma, Rhinella, Incillius, and Anaryxus).
We also formally corrected the data using the phylogenetic mixed model approach proposed by Castillo (39) for the three different taxa. We fitted a generalized linear mixed model using Markov Chain Monte Carlo to study the relationship between RI and genetic distance for sympatric and allopatric species. We used the function ginv (library MASS, (74)) to find the generalized inverse of the (1-genetic distance) matrix (39). We used the package MCMCglmm (75) and fitted a linear model, in which the magnitude of isolation (premating or postzygotic for Drosophila, and postzygotic for Lepidopterans and Bufo) was the response variable, genetic distance was a predictor variable, geographic overlap (i.e., whether a species pair was sympatric or allopatric) was a fixed effect, and the phylogenetic variance matrix was a random effect. The model also included an interaction between transformed genetic distance and geographic overlap. We ran two independent MCMC chains. To determine if the model converged in each of the two chains, we used the function gelman.diag (library coda, (76)). A chain was considered converged if all scale reduction factors for all variables (both fixed and random effects) were ≤1.1 for each of the two chains. We calculated the 95% confidence interval for the intercept and the slope using the function HPDinterval (library coda, (76)).
Asymmetries and geographic range
Asymmetries and overlap of geographic range are two proxies of the risk and cost of hybridization. For sympatric species pairs where postzygotic RI is stronger for one of the two reciprocal crosses, premating RI should also be asymmetric and stronger for the same cross direction if reinforcement has driven the evolution of premating RI. This correlation should not exist for allopatric species pairs (42). For the Drosophila dataset, we calculated the proportion of the variance in asymmetries in premating RI explained by asymmetries in postzygotic RI as the adjusted R2 from a linear regression. We fit linear models with the function lm (library ‘stats’, (70)) where the magnitude of the asymmetry in premating RI was the response, and the magnitude of the postzygotic RI was the only continuous effect (N = 167 species pairs). We also calculated the partial correlation coefficient between asymmetry in premating and postzygotic RI while controlling for genetic distance using the function pcor.test (ppcor library, (77)) for a dataset that included the magnitude of both asymmetries and genetic distance (N = 106 species pairs).
We used a linear model to study the effect of geographic range overlap on patterns of RI. Since we studied two types of RI barriers, we fitted two different linear models. The approach for the two models was identical and the model with the interaction followed the form:
 where Isolation refers to either a premating or postzygotic RI metric. To assess the significance of the interaction, we followed a maximum-likelihood model simplification approach (78, 79), in which we compared the model with the interaction to one without the interaction that followed the form:
where Isolation refers to either a premating or postzygotic RI metric. To assess the significance of the interaction, we followed a maximum-likelihood model simplification approach (78, 79), in which we compared the model with the interaction to one without the interaction that followed the form:

To compare the two linear models, we used the function lrtest (library ‘lmtest’, (80)).
Acknowledgements
We thank J. M. Coughlan and A. Dagilis for comments and J. M. Good for helpful discussions. Research reported in this publication was supported by the National Institute of General Medical Sciences of the National Institutes of Health (NIH) under Award Number R01GM121750 to DRM and R35GM124701 to BSC. The content is solely the responsibility of the authors and does not necessarily represent the official views of the NIH.
Footnotes
brandon.cooper{at}umontana.edu
REFERENCES
- 1.↵
- 2.↵
- 3.↵
- 4.↵
- 5.↵
- 6.↵
- 7.↵
- 8.↵
- 9.↵
- 10.
- 11.↵
- 12.↵
- 13.
- 14.↵
- 15.↵
- 16.↵
- 17.
- 18.↵
- 19.↵
- 20.
- 21.
- 22.↵
- 23.↵
- 24.↵
- 25.↵
- 26.↵
- 27.↵
- 28.↵
- 29.↵
- 30.↵
- 31.↵
- 32.↵
- 33.
- 34.↵
- 35.↵
- 36.↵
- 37.
- 38.↵
- 39.↵
- 40.↵
- 41.↵
- 42.↵
- 43.↵
- 44.↵
- 45.↵
- 46.↵
- 47.↵
- 48.↵
- 49.↵
- 50.↵
- 51.↵
- 52.↵
- 53.↵
- 54.↵
- 55.↵
- 56.↵
- 57.
- 58.↵
- 59.↵
- 60.
- 61.↵
- 62.↵
- 63.
- 64.
- 65.↵
- 66.↵
- 67.↵
- 68.↵
- 69.↵
- 70.↵
- 71.↵
- 72.↵
- 73.↵
- 74.↵
- 75.↵
- 76.↵
- 77.↵
- 78.↵
- 79.↵
- 80.↵




