

Abstract
The B.1.617 variant emerged in the Indian state of Maharashtra in late 2020 and has spread throughout India and to at least 40 countries. There have been fears that two key mutations seen in the receptor binding domain L452R and E484Q would have additive effects on evasion of neutralising antibodies. Here we delineate the phylogenetics of B.1.617 and spike mutation frequencies, in the context of others bearing L452R. The defining mutations in B.1.617.1 spike are L452R and E484Q in the RBD that interacts with ACE2 and is the target of neutralising antibodies. All B.1.617 viruses have the P681R mutation in the polybasic cleavage site region in spike. We report that B.1.617.1 spike bearing L452R, E484Q and P681R mediates entry into cells with slightly reduced efficiency compared to Wuhan-1. This spike confers modestly reduced sensitivity to BNT162b2 mRNA vaccine-elicited antibodies that is similar in magnitude to the loss of sensitivity conferred by L452R or E484Q alone. Furthermore we show that the P681R mutation significantly augments syncytium formation upon the B.1.617.1 spike protein, potentially contributing to increased pathogenesis observed in hamsters and infection growth rates observed in humans.
Introduction
Global control of the SARS-CoV-2 pandemic has yet to be realised despite availability of highly effective vaccines1,2. Emergence of new variants with multiple mutations is likely the result of chronic infections within individuals who are immune compromised3. New variant emergence and transmission has coincided with rollout of vaccines, potentially threatening their success in controlling the pandemic4,5.
India experienced a wave of infections in mid 2020 and was controlled by a nationwide lockdown. Since easing of restrictions, India has seen expansion in cases of COVID-19 since March 2021. The B.1.1.7 variant has been growing in the north of the country and is known to be more transmissible than previous viruses bearing the D614G spike mutation6. The B.1.617 variant emerged in the state of Maharashtra in late 2020/early 20217 and has spread throughout India and to at least 40 countries. It was labelled initially as a double mutant since two of the mutations L452R and E484Q were matched to an in-house screening database for mutations leading to probable evasion of antibodies and/or being linked to increased transmissibility.
The defining mutations in spike are L452R and E484Q in the critical receptor binding domain that interacts with ACE28. L452R was observed in the California variant and is associated with increase in viral load and around 20% increased transmissibility 9. It was also associated with increased ACE2 binding, increased infectivity10 and 3-6 fold loss of neutralisation sensitivity to vaccine elicited sera in experiments with pseudotyped virus (PV) particles10,11. Little is known about E484Q, though E484K is a defining feature of two VOCs, B.1.351 and P.112. E484K is known to confer around 10 fold loss of sensitivity to neutralising antibodies in vaccine and convalescent sera13,14. In contrast to L452R, E484Q has not been observed in transmissible variants.
Here we demonstrate three lineages of B.1.617, all bearing the L452R mutation. We report key differences in amino acids between sub-lineages and focus on B.1.617.1 bearing three key mutations: L452R, E484Q and P681R. We report modestly reduced sensitivity of the ancestral B.1.617.1 spike protein to BNT162b2 mRNA vaccine-elicited antibodies that is similar in magnitude to the loss of sensitivity conferred by L452R or E484Q alone. Furthermore we show that the P681R mutation significantly augments syncytium formation upon the B.1.617.1 spike protein, potentially contributing to increased pathogenesis observed in hamsters15 and high growth rates observed in humans.
Results
Three B.1.617 sub-lineages are characterised by L452R and P681R in spike
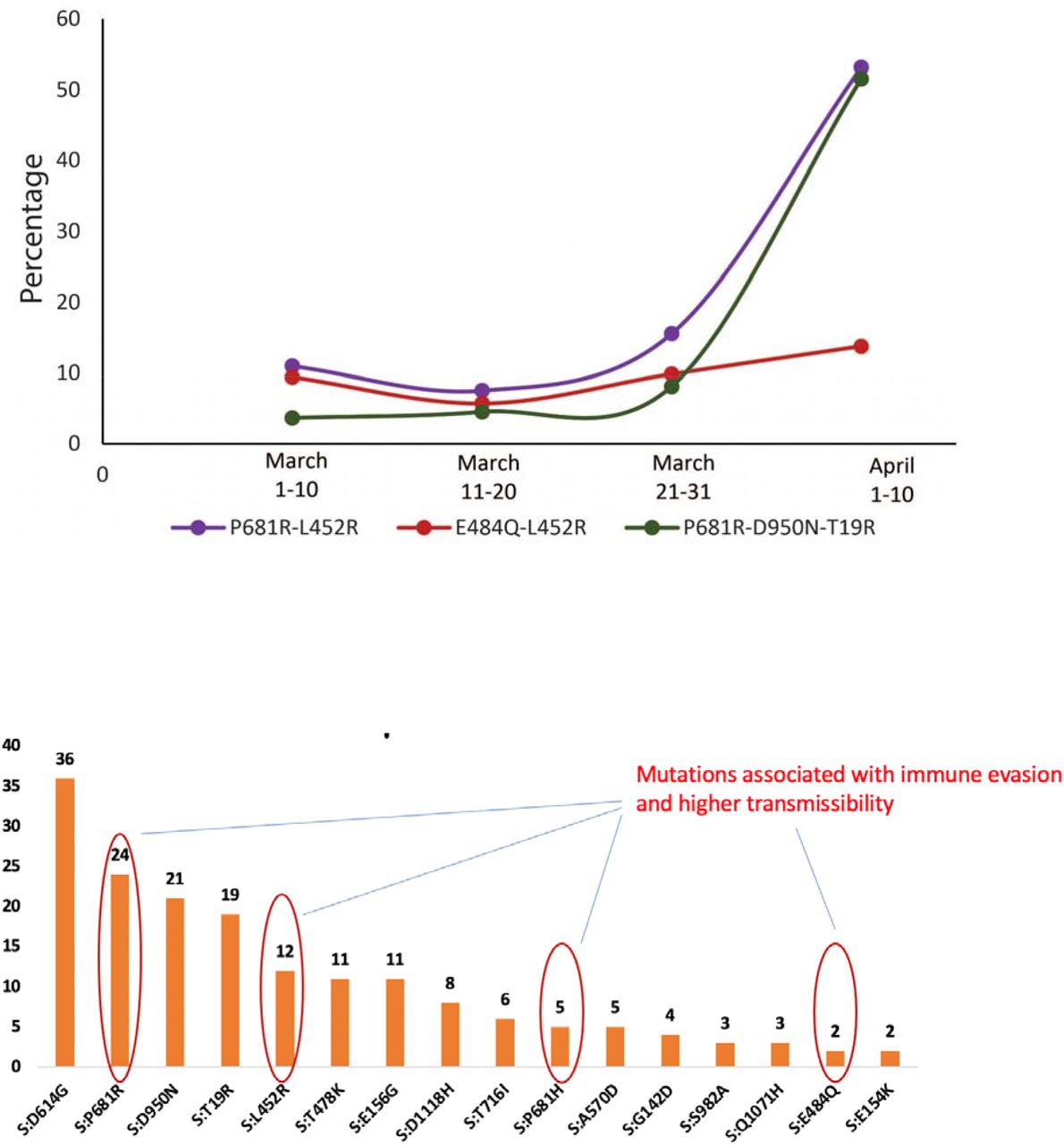
We downloaded whole genome SARS-CoV-2 sequences containing L452R from GISAID, subsampled and inferred a maximum likelihood phylogenetic tree (Figure 1A). We annotated the sequences based on the accompanying mutations and observed that B.1.617.1 has three key spike mutations L452R, E484Q and P681R, whereas B.1.617.2 is characterised by L452R, T478K and P681R. This likely signifies loss of E484Q in B.1.617.2 given that B.1.617.3 also bears E484Q in 90% of sequences (Figure 1). The number of sequenced isolates of B.1.617.1 and B.1.617.2 has been steadily increasing both globally (Figure 1B) and in India (Figure 1C), though with the caveat of very low sequencing of prevalent cases. A high proportion of global sequences is from the UK where B.1.617.2 appears to be dominating imported cases and local transmissions. There is already significant diversity in B.1.617.1 as demonstrated in Figure 1D.

A. Maximum-likelihood phylogeny of lineages bearing L452R in spike. All sequences with the L452R mutation were downloaded from https://gisaid.org and manually aligned to reference strain MN908947.3 with mafft. Sequences were deduplicated and a random subset of 400 global sequences, and 100 USA sequences were then selected with seqtk. All sequence lineages were assigned using pangolin v2.4. Major lineages are indicated as straight lines adjacent to the heatmap, alongside mutations of current interest. The phylogeny was inferred with IQTREE2 v2.1.3. B, C. The cumulative number of B. global and C. Indian B.1.617 sequences between December 2020 - April 2021 are indicated D. Table of relevant mutations in B.1.617 at prevalence of >50%. All data collected on 4th May 2021. E. Cases and designated lineages of a localised hospital-based outbreak amongst fully vaccinated staff in Delhi, India.
Vaccine breakthrough in a Delhi health facility is dominated by B1.617
Vaccination of health care workers in Delhi was started in early 2021, with the ChdOx-1 vaccine. Surveillance has suggested B.1.1.7 dominance in the Delhi area during early 2021, with growth of B.1.617 since March 2021 (personal communication). During the wave of infections during March and April an outbreak of SARS-CoV-2 was confirmed in 33 vaccinated staff members at a single tertiary centre (age 27-77 years). Sequencing16 revealed the majority were B.1.617.2 with a range of other B lineage viruses including B.1.1.7 (Figure 1E). Importantly no severe cases were documented in this event.
B.1.617.1 Spike confers partial evasion of BNTl62b2 vaccine elicited antibodies
Spike mutations L452R and E484Q are in the receptor binding domain that not only binds ACE28, but is a target for neutralising antibodies17–19 (Figure 2). We tested the neutralisation sensitivity of B.1.617.1 Spike bearing L452R, E484Q and P681R using a previously reported pseudotyped virus (PV) system with HIV-1 particles bearing SARS-CoV-2 spike (Figure 3). PV testing against neutralising antibodies has been shown to be highly correlated with live virus systems20. We tested nine stored sera from Pfizer BNT162b2 vaccinees against a range of spike mutation bearing PV. As expected E484K conferred a ten-fold reduction in neutralisation by vaccine sera, and E484Q had a slightly milder yet significant impact. When E484Q and L452R were combined, there was a statistically significant loss of sensitivity as compared to wild type, but the fold change was similar to that observed with each mutation individually with no evidence for an additive effect.

A. Surface representation of the spike protein in closed formation (PDB: 6ZGE) in a vertical view with the location of L452 and E484 mutations highlighted as red and green sphere, respectively. Each monomer in the homotrimer is coloured accordingly B. Surface representation of the same spike protein in closed confirmation in a “top-down” view along the trimer axis. The residues associated with RBD substitutions L452R, and E484Q are highlighted in red and green spheres respectively, on a single monomer. C. Ribbon representation of a single monomer of the same Spike, with residues L452 and E484 highlighted as spheres coloured by element. D. Surface representation of the spike protein in open formation with neutralising antibody H4 (pink spheres, PDB: 7L58, Rapp et al, 2021) bound to one monomer of the spike protein. Residues L452 and E484 are indicated with red and green sphere, respectively. Note that E484 is partially occluded by the bound monoclonal antibody E. Ribbon representation of the interaction between the neutralising antibody H4 and the RBD of a spike monomer. Residue E484 has direct interaction with the antibody, suggesting that mutations at this site may be involved in immune escape.

A-B. Serial dilutions of sera used and approx. 250,000 RLU of virus used for each mutant at each dilution. B. indicates linked serum samples Geometric Mean Titre (GMT) shown with 95% CI. ** p<0.01.
P681R confers increased syncytium formation capability on B.1.617.1 spike
Spike is known to mediate cell entry via interaction with ACE2 and TMPRSS221 and is a major determinant of viral infectivity. Given that that B.1.617.1 does not appear to be highly immune evasive, we hypothesised that it may be have higher infectivity. We tested single round viral entry using the PV system, infecting target 293T cells over-expressing ACE2 and TMPRSS2, as well as Calu-3 lung cells expressing endogenous levels of ACE2 and TMPRSS2 (Supplementary figure 1). We observed similar entry efficiency across mutants, all of which appeared lower than the Wuhan-1 D614G wild type. (Figure 4).

PV were generated in 293T cells filtered and then used to infect target cells. Luciferase was measured 48 hours after infection. Infectivity data were corrected for genome copy input using SG-PERT. Mean is plotted with error bars representing SEM.

PV were generated in 293T cells filtered and then used to infect target cells. Luciferase was measured 48 hours after infection. Input virus inoculum was corrected for genome copy input using SG-PERT. Mean is plotted with error bars representing SEM. Data are representative of two experiments.
SARS-CoV-2 infection in clinically relevant cells is TMPRSS2 dependent and requires fusion at the plasma membrane, potentially to avoid restriction factors in endosomes22. The plasma membrane route of entry, and indeed transmissibility in animal models, is critically dependent on the polybasic cleavage site (PBCS) between S1 and S222,23. Mutations at P681 in the PBCS have been observed in multiple SARS-CoV-2 lineages, most notably in the B.1.1.7 variant that likely emerged in the UK.
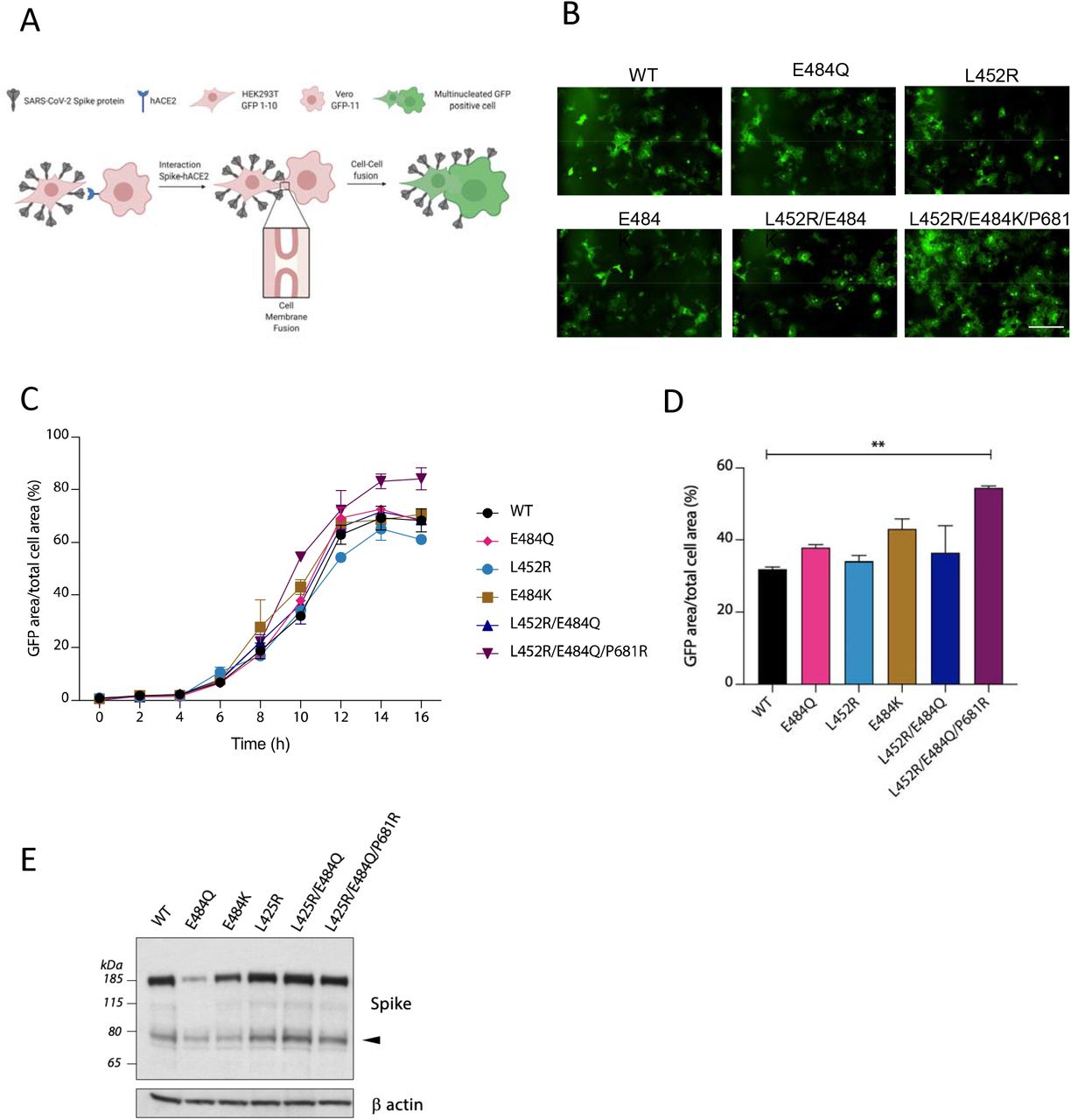
We previously showed that B.1.1.7 spike, bearing P681H, had significantly higher fusogenic potential than a D614G Wuhan-1 virus24. We therefore tested a series of mutations in B.1.617 spike using a split GFP system. We transfected spike bearing plasmids into donor cells and co-cultured them with acceptor cells (Figure 5). We found no significant differences for the single mutants tested L452R, E484Q/K, or indeed the double mutant L452R+E484Q. However, the triple mutant with P681R demonstrated higher fusion activity and syncytium formation, mediated specifically by P681R (Figure 5, supplementary figure 2).


A. Quantification of cell-cell fusion kinetics showing percentage of green area to total cell area over time. Mean is plotted with error bars representing SEM B. Quantification of cell-cell fusion of the indicated Spike mutants at 10 hours post transfection. Mean is plotted with error bars representing SEM. **p<0.005 Unpaired Student t test P681R combinations are seen increasingly more frequently during the ongoing outbreak in Delhi. Loss of E484Q has not reduced fitness of B.1.617 variants. The most rapidly expanding lineage is B.1.617.2 that lacks E484Q B) In sequenced post-vaccination breakthroughs P681R was the most frequently observed mutation after D614G.

A. Schematic of cell-cell fusion assay. B Reconstructed images at 10 hours of GFP positive syncytia formation. Scale bars represent 400 mm. C. Quantification of cell-cell fusion kinetics showing percentage of green area to total cell area over time. Mean is plotted with error bars representing SEM D. Quantification of cell-cell fusion of the indicated Spike mutants at 10 hours post transfection. D. Mean is plotted with error bars representing SEM. **p<0.005 Unpaired Student t test. E. Western blotting of donor cell lysates using an antibody against S2.
Discussion
Here we have shown that B.1.617 spike bearing L452R, E484Q and P681R has modest ability to avoid neutralising antibodies elicited by BNT162b2 vaccination. The fold reduction for the two RBD mutations L452R and E484Q was no greater than the individual mutations alone, arguing against use of the term “double mutant”. The loss of neutralisation of B.1.617 has likely contributed to an epidemic wave in India where background infection to the Wuhan-1 D614G in 2020 was between 20-50%25. Despite in vitro data showing only small loss of neutralisation against B.1.617 with the Covaxin vaccine26, here we also show vaccine breakthrough in health care workers at a single tertiary hospital who were fully vaccinated with ChAdOx-1 vaccine (adenovirus vectored). These infections were predominantly B.1.617, with a mix of other lineages bearing D614G in spike. The dominance of B.1.617 could be explained by prevalence of this lineage in community infection or simply reflect transmission between HCWs. The data nonetheless raise the possibility of a transmission advantage of B.1.617 in vaccinated individuals.
We measured spike mediated entry into target cell lines exogenously or endogenously expressing ACE2 and TMPRSS2 receptors. The E484K, L452R and P681R mutant had reduced entry efficiency relative to wild type. A recent report using a spike B.1.617 with a larger set of mutations found variable entry efficiency relative to wild type across cell types27.
Virus infectivity and fusogenicity mediated by the PBCS is a key determinant of pathogenicity and transmissibility22,28 and there are indications that giant cells/syncitia formation are associated with fatal disease29. We find that P681R is associated with enhanced capacity to induce cell-cell fusion and syncitia formation, and that P681R alone confers this ability on the B.1.617.1 spike with RBD mutations L452R and E484Q.
It is unclear whether B.1.617 variants will prove more transmissible than B.1.1.7, also circulating in India and now globally dominant. In the absence of published data on transmissibility of B.1.617 we predict that this variant will have a transmission advantage relative to Wuhan-1 with D614G in individuals with pre-existing immunity from vaccines/natural infection as well as in settings where there is low vaccine coverage and low prior exposure. Lower protection against B.1.351, the variant with least sensitivity to neutralising antibodies, has been demonstrated for at least three vaccines2,30–32. However, progression to severe disease and death was low in all studies. Therefore, at population scale, extensive vaccination will likely protect against moderate to severe disease and will reduce transmission of B.1.617 given the in vitro neutralisation data we and others have presented.
Methods
Phylogenetic Analysis
All sequences excluding low-quality sequences (>5% N regions) with the L452R mutation were downloaded from https://gisaid.org33 on the 4th May 2021 and manually aligned to reference strain MN908947.3 with mafft v4.475 34 using the --keeplength --addfragments option. Sequences were de-duplicated using bbtools dedupe.sh. Due to the high proportion of USA-centric sequences containing L452R, all USA sequences were extracted and saved to a separate fasta file. A random subset of 400 global sequences (excluding USA), and 100 USA sequences were then selected with seqtk and concatenated. Sequence lineages were assigned to all sequences with pangolin v2.4 (https://github.com/cov-lineages/pangolin) and pangolearn (04/05/2021).
Phylogenies were then inferred using maximum-likelihood in IQTREE v2.1.3 35 using a GTR+R6 model and the -fast option. Mutations of interest were determined using a local instance of nextclade-cli v0.14.2 (https://github.com/nextstrain/nextclade). The inferred phylogeny was annotated in R v4.04 using ggtree v2.2.436 and rooted on the SARS-CoV-2 reference sequence, and nodes arranged in descending order. Major lineages were annotated on the phylogeny, as well as a heatmap indicating which mutations of interest were carried by each viral sequence.
Structural Analyses
The PyMOL Molecular Graphics System v.2.4.0 (https://github.com/schrodinger/pymol-open-source/releases) was used to map the location of the two RDB mutants L452R and E484Q onto two previously published SARS-CoV-2 spike glycoprotein structures. Th two structures included a closed-conformation spike protein - PDB: 6ZGE37 and a spike protein in open conformation, bound to nAb H4 38.
Serum samples and ethical approval
Ethical approval for use of serum samples. Controls with COVID-19 were enrolled to the NIHR BioResource Centre Cambridge under ethics review board (17/EE/0025).
Cells
HEK 293T CRL-3216, Vero CCL-81 were purchased from ATCC and maintained in Dulbecco”s Modified Eagle Medium (DMEM) supplemented with 10% fetal calf serum (FCS), 100 U/ml penicillin, and 100mg/ml streptomycin. All cells were regularly tested and are mycoplasma free.
Pseudotype virus preparation
Plasmids encoding the spike protein of SARS-CoV-2 D614 with a C terminal 19 amino acid deletion with D614G, were used as a template to produce variants lacking amino acids at position H69 and V70, as well as mutations N439K and Y453F. Mutations were introduced using Quickchange Lightning Site-Directed Mutagenesis kit (Agilent) following the manufacturer”s instructions. B. 1.1.7 S expressing plasmid preparation was described previously, but in brief was generated by step wise mutagenesis. Viral vectors were prepared by transfection of 293T cells by using Fugene HD transfection reagent (Promega). 293T cells were transfected with a mixture of 11ul of Fugene HD, 1μg of pCDNAΔ19 spike-HA, 1ug of p8.91 HIV-1 gag-pol expression vector and 1.5μg of pCSFLW (expressing the firefly luciferase reporter gene with the HIV-1 packaging signal). Viral supernatant was collected at 48 and 72h after transfection, filtered through 0.45um filter and stored at −80°C as previously described. Infectivity was measured by luciferase detection in target 293T cells transfected with TMPRSS2 and ACE2.
Standardisation of virus input by SYBR Green-based product-enhanced PCR assay (SG-PERT)
The reverse transcriptase activity of virus preparations was determined by qPCR using a SYBR Green-based product-enhanced PCR assay (SG-PERT) as previously described39. Briefly, 10-fold dilutions of virus supernatant were lysed in a 1:1 ratio in a 2x lysis solution (made up of 40% glycerol v/v 0.25% Trition X-100 v/v 100mM KCl, RNase inhibitor 0.8 U/ml, TrisHCL 100mM, buffered to pH7.4) for 10 minutes at room temperature.
12μl of each sample lysate was added to thirteen 13μl of a SYBR Green master mix (containing 0.5μM of MS2-RNA Fwd and Rev primers, 3.5pmol/ml of MS2-RNA, and 0.125U/μl of Ribolock RNAse inhibitor and cycled in a QuantStudio. Relative amounts of reverse transcriptase activity were determined as the rate of transcription of bacteriophage MS2 RNA, with absolute RT activity calculated by comparing the relative amounts of RT to an RT standard of known activity.
Plasmids for split GFP system to measure cell-cell fusion
pQCXIP□BSR□GFP11 and pQCXIP□GFP1□10 were from Yutaka Hata40 Addgene plasmid #68716; http://n2t.net/addgene:68716; RRID:Addgene_68716 and Addgene plasmid #68715; http://n2t.net/addgene:68715; RRID:Addgene_68715)
Generation of GFP1□ 10 or GFP11 lentiviral particles
Lentiviral particles were generated by co-transfection of 293T cells with pQCXIP□BSR□GFP11 or pQCXIP□GFP1□10 as previously described 41. Supernatant containing virus particles was harvested after 48 and 72 hours, 0.45 μm filtered, and used to infect 293T or Vero cells to generate stable cell lines. 293T and Vero cells were transduced to stably express GFP1 10 or GFP11 respectively and were selected with 2 μg/ml puromycin.
Cell-cell fusion assay
Cell-cell fusion assay was carried out as previously described41,42 but using a Split-GFP system. Briefly, 293T-GFP1-10 and Vero-GFP11 cells were seeded at 80% confluence in a 24 multiwell plate the day before. 293T cells were co-transfected with 1.5 μg of spike expression plasmids in pCDNA3 using Fugene 6 and following the manufacturer”s instructions (Promega). 293T-GFP1-10 cells were then detached 5 hours post transfection, mixed together with the Vero-GFP11 cells, and plated in a 12 multiwell plate. Cell-cell fusion was measured using an Incucyte and determined as the proportion of green area to total phase area. Data were then analysed using Incucyte software analysis. Graphs were generated using Prism 8 software. Furin inhibitor CMK (Calbiochem) was added at transfection.
Western blotting
Cells were lysed and supernatants collected 18 hours post transfection. Purified virions were prepared by harvesting supernatants and passing through a 0.45 μm filter. Clarified supernatants were then loaded onto a thin layer of 8.4% optiprep density gradient medium (Sigma-Aldrich) and placed in a TLA55 rotor (Beckman Coulter) for ultracentrifugation for 2 hours at 20,000 rpm. The pellet was then resuspended for western blotting. Cells were lysed with cell lysis buffer (Cell signalling), treated with Benzonase Nuclease (70664 Millipore) and boiled for 5 min. Samples were then run on 4%-12% Bis Tris gels and transferred onto nitrocellulose or PVDF membranes using an iBlot or semidry (Life Technologies and Biorad, respectively).
Membranes were blocked for 1 hour in 5% non-fat milk in PBS + 0.1% Tween-20 (PBST) at room temperature with agitation, incubated in primary antibody (anti-SARS-CoV-2 Spike, which detects the S2 subunit of SARS-CoV-2 S (Invitrogen, PA1-41165), anti-GAPDH (proteintech) or anti-p24 (NIBSC)) diluted in 5% non-fat milk in PBST for 2 hours at 4°C with agitation, washed four times in PBST for 5 minutes at room temperature with agitation and incubated in secondary antibodies anti-rabbit HRP (1:10000, Invitrogen 31462), anti-bactin HRP (1:5000; sc-47778) diluted in 5% non-fat milk in PBST for 1 hour with agitation at room temperature. Membranes were washed four times in PBST for 5 minutes at room temperature and imaged directly using a ChemiDoc MP imaging system (Bio-Rad).
Serum pseudotype neutralisation assay
Spike pseudotype assays have been shown to have similar characteristics as neutralisation testing using fully infectious wild type SARS-CoV-220. Virus neutralisation assays were performed on 293T cell transiently transfected with ACE2 and TMPRSS2 using SARS-CoV-2 spike pseudotyped virus expressing luciferase43. Pseudotyped virus was incubated with serial dilution of heat inactivated human serum samples or convalescent plasma in duplicate for 1h at 37°C. Virus and cell only controls were also included. Then, freshly trypsinized 293T ACE2/TMPRSS2 expressing cells were added to each well. Following 48h incubation in a 5% CO2 environment at 37°C, the luminescence was measured using Steady-Glo Luciferase assay system (Promega).
COG-UK CONSORTIUM
Funding acquisition, Leadership and supervision, Metadata curation, Project administration, Samples and logistics, Sequencing and analysis, Software and analysis tools, and Visualisation:
Samuel C Robson13.
Funding acquisition, Leadership and supervision, Metadata curation, Project administration, Samples and logistics, Sequencing and analysis, and Software and analysis tools:
Nicholas J Loman41 and Thomas R Connor10, 69.
Leadership and supervision, Metadata curation, Project administration, Samples and logistics, Sequencing and analysis, Software and analysis tools, and Visualisation:
Tanya Golubchik5.
Funding acquisition, Metadata curation, Samples and logistics, Sequencing and analysis, Software and analysis tools, and Visualisation:
Rocio T Martinez Nunez42.
Funding acquisition, Leadership and supervision, Metadata curation, Project administration, and Samples and logistics:
Catherine Ludden88.
Funding acquisition, Leadership and supervision, Metadata curation, Samples and logistics, and Sequencing and analysis:
Sally Corden69.
Funding acquisition, Leadership and supervision, Project administration, Samples and logistics, and Sequencing and analysis:
Ian Johnston99 and David Bonsall5.
Funding acquisition, Leadership and supervision, Sequencing and analysis, Software and analysis tools, and Visualisation:
Colin P Smith87 and Ali R Awan28.
Funding acquisition, Samples and logistics, Sequencing and analysis, Software and analysis tools, and Visualisation:
Giselda Bucca87.
Leadership and supervision, Metadata curation, Project administration, Samples and logistics, and Sequencing and analysis:
M. Estee Torok22, 101.
Leadership and supervision, Metadata curation, Project administration, Samples and logistics, and Visualisation:
Kordo Saeed81, 110 and Jacqui A Prieto83, 109.
Leadership and supervision, Metadata curation, Project administration, Sequencing and analysis, and Software and analysis tools:
David K Jackson99.
Metadata curation, Project administration, Samples and logistics, Sequencing and analysis, and Software and analysis tools:
William L Hamilton22.
Metadata curation, Project administration, Samples and logistics, Sequencing and analysis, and Visualisation:
Luke B Snell11.
Funding acquisition, Leadership and supervision, Metadata curation, and Samples and logistics:
Catherine Moore69.
Funding acquisition, Leadership and supervision, Project administration, and Samples and logistics:
Ewan M Harrison99, 88.
Leadership and supervision, Metadata curation, Project administration, and Samples and logistics:
Sonia Goncalves99 and Leigh M Jackson91.
Leadership and supervision, Metadata curation, Samples and logistics, and Sequencing and analysis:
Ian G Goodfellow24, Derek J Fairley3, 72, Matthew W Loose18 and Joanne Watkins69.
Leadership and supervision, Metadata curation, Samples and logistics, and Software and analysis tools:
Rich Livett99.
Leadership and supervision, Metadata curation, Samples and logistics, and Visualisation:
Samuel Moses25, 106.
Leadership and supervision, Metadata curation, Sequencing and analysis, and Software and analysis tools:
Roberto Amato99, Sam Nicholls41 and Matthew Bull69.
Leadership and supervision, Project administration, Samples and logistics, and
Sequencing and analysis:
Darren L Smith37, 58, 105.
Leadership and supervision, Sequencing and analysis, Software and analysis tools, and Visualisation:
Jeff Barrett99, David M Aanensen14, 114.
Metadata curation, Project administration, Samples and logistics, and Sequencing and analysis:
Martin D Curran65, Surendra Parmar65, Dinesh Aggarwal95, 99, 64 and James G Shepherd48.
Metadata curation, Project administration, Sequencing and analysis, and Software and analysis tools:
Matthew D Parker93.
Metadata curation, Samples and logistics, Sequencing and analysis, and Visualisation:
Sharon Glaysher61.
Metadata curation, Sequencing and analysis, Software and analysis tools, and Visualisation:
Matthew Bashton37, 58, Anthony P Underwood14, 114, Nicole Pacchiarini69 and Katie F Loveson77.
Project administration, Sequencing and analysis, Software and analysis tools, and Visualisation:
Alessandro M Carabelli88.
Funding acquisition, Leadership and supervision, and Metadata curation:
Kate E Templeton53, 90.
Funding acquisition, Leadership and supervision, and Project administration:
Cordelia F Langford99, John Sillitoe99, Thushan I de Silva93 and Dennis Wang93.
Funding acquisition, Leadership and supervision, and Sequencing and analysis:
Dominic Kwiatkowski99, 107, Andrew Rambaut90, Justin O’Grady70, 89 and Simon Cottrell69.
Leadership and supervision, Metadata curation, and Sequencing and analysis:
Matthew T.G. Holden68 and Emma C Thomson48.
Leadership and supervision, Project administration, and Samples and logistics:
Husam Osman64, 36, Monique Andersson59, Anoop J Chauhan61 and Mohammed O Hassan-Ibrahim6.
Leadership and supervision, Project administration, and Sequencing and analysis:
Mara Lawniczak99.
Leadership and supervision, Samples and logistics, and Sequencing and analysis:
Ravi Kumar Gupta88, 113, Alex Alderton99, Meera Chand66, Chrystala Constantinidou94, Meera Unnikrishnan94, Alistair C Darby92, Julian A Hiscox92 and Steve Paterson92.
Leadership and supervision, Sequencing and analysis, and Software and analysis tools:
Inigo Martincorena99, David L Robertson48, Erik M Volz39, Andrew J Page70 and Oliver G Pybus23.
Leadership and supervision, Sequencing and analysis, and Visualisation:
Andrew R Bassett99.
Metadata curation, Project administration, and Samples and logistics:
Cristina V Ariani99, Michael H Spencer Chapman99, 88, Kathy K Li48, Rajiv N Shah48, Natasha G Jesudason48 and Yusri Taha50.
Metadata curation, Project administration, and Sequencing and analysis:
Martin P McHugh53 and Rebecca Dewar53.
Metadata curation, Samples and logistics, and Sequencing and analysis:
Aminu S Jahun24, Claire McMurray41, Sarojini Pandey84, James P McKenna3, Andrew Nelson58, 105, Gregory R Young37, 58, Clare M McCann58, 105 and Scott Elliott61.
Metadata curation, Samples and logistics, and Visualisation:
Hannah Lowe25.
Metadata curation, Sequencing and analysis, and Software and analysis tools:
Ben Temperton91, Sunando Roy82, Anna Price10, Sara Rey69 and Matthew Wyles93.
Metadata curation, Sequencing and analysis, and Visualisation:
Stefan Rooke90 and Sharif Shaaban68.
Project administration, Samples and logistics, Sequencing and analysis:
Mariateresa de Cesare98.
Project administration, Samples and logistics, and Software and analysis tools:
Laura Letchford99.
Project administration, Samples and logistics, and Visualisation:
Siona Silveira81, Emanuela Pelosi81 and Eleri Wilson-Davies81.
Samples and logistics, Sequencing and analysis, and Software and analysis tools:
Myra Hosmillo24.
Sequencing and analysis, Software and analysis tools, and Visualisation:
Áine O’Toole90, Andrew R Hesketh87, Richard Stark94, Louis du Plessis23, Chris Ruis88, Helen Adams4 and Yann Bourgeois76.
Funding acquisition, and Leadership and supervision:
Stephen L Michell91, Dimitris Grammatopoulos84, 112, Jonathan Edgeworth12, Judith Breuer30, 82, John A Todd98 and Christophe Fraser5.
Funding acquisition, and Project administration:
David Buck98 and Michaela John9.
Leadership and supervision, and Metadata curation:
Gemma L Kay70.
Leadership and supervision, and Project administration:
Steve Palmer99, Sharon J Peacock88, 64 and David Heyburn69.
Leadership and supervision, and Samples and logistics:
Danni Weldon99, Esther Robinson64, 36, Alan McNally41, 86, Peter Muir64, Ian B Vipond64, John BoYes29, Venkat Sivaprakasam46, Tranprit Salluja75, Samir Dervisevic54 and Emma J Meader54.
Leadership and supervision, and Sequencing and analysis:
Naomi R Park99, Karen Oliver99, Aaron R Jeffries91, Sascha Ott94, Ana da Silva Filipe48, David A Simpson72 and Chris Williams69.
Leadership and supervision, and Visualisation:
Jane AH Masoli73, 91.
Metadata curation, and Samples and logistics:
Bridget A Knight73, 91, Christopher R Jones73, 91, Cherian Koshy1, Amy Ash1, Anna Casey71, Andrew Bosworth64, 36, Liz Ratcliffe71, Li Xu-McCrae36, Hannah M Pymont64, Stephanie Hutchings64, Lisa Berry84, Katie Jones84, Fenella Halstead46, Thomas Davis21, Christopher Holmes16, Miren Iturriza-Gomara92, Anita O Lucaci92, Paul Anthony Randell38, 104, Alison Cox38, 104, Pinglawathee Madona38, 104, Kathryn Ann Harris30, Julianne Rose Brown30, Tabitha W Mahungu74, Dianne Irish-Tavares74, Tanzina Haque74, Jennifer Hart74, Eric Witele74, Melisa Louise Fenton75, Steven Liggett79, Clive Graham56, Emma Swindells57, Jennifer Collins50, Gary Eltringham50, Sharon Campbell17, Patrick C McClure97, Gemma Clark15, Tim J Sloan60, Carl Jones15 and Jessica Lynch2 111.
Metadata curation, and Sequencing and analysis:
Ben Warne8, Steven Leonard99, Jillian Durham99, Thomas Williams90, Sam T Haldenby92, Nathaniel Storey30, Nabil-Fareed Alikhan70, Nadine Holmes18, Christopher Moore18, Matthew Carlile18, Malorie Perry69, Noel Craine69, Ronan A Lyons80, Angela H Beckett13, Salman Goudarzi77, Christopher Fearn77, Kate Cook77, Hannah Dent and Hannah Paul77.
Metadata curation, and Software and analysis tools:
Robert Davies99.
Project administration, and Samples and logistics:
Beth Blane88, Sophia T Girgis88, Mathew A Beale99, Katherine L Bellis99, 88, Matthew J Dorman99, Eleanor Drury99, Leanne Kane99, Sally Kay99, Samantha McGuigan99, Rachel Nelson99, Liam Prestwood99, Shavanthi Rajatileka99, Rahul Batra12, Rachel J Williams82, Mark Kristiansen82, Angie Green98, Anita Justice59, Adhyana I.K Mahanama81, 102 and Buddhini Samaraweera81, 102.
Project administration, and Sequencing and analysis:
Nazreen F Hadjirin88 and Joshua Quick41.
Project administration, and Software and analysis tools:
Radoslaw Poplawski41.
Samples and logistics, and Sequencing and analysis:
Leanne M Kermack88, Nicola Reynolds7, Grant Hall24, Yasmin Chaudhry24, Malte L Pinckert24, Iliana Georgana24, Robin J Moll99, Alicia Thornton66, Richard Myers66, Joanne Stockton41, Charlotte A Williams82, Wen C Yew58, Alexander J Trotter70, Amy Trebes98, George MacIntyre-Cockett98, Alec Birchley69, Alexander Adams69, Amy Plimmer69, Bree Gatica-Wilcox69, Caoimhe McKerr69, Ember Hilvers69, Hannah Jones69, Hibo Asad69, Jason Coombes69, Johnathan M Evans69, Laia Fina69, Lauren Gilbert69, Lee Graham69, Michelle Cronin69, Sara Kumziene-SummerhaYes69, Sarah Taylor69, Sophie Jones69, Danielle C Groves93, Peijun Zhang93, Marta Gallis93 and Stavroula F Louka93.
Samples and logistics, and Software and analysis tools:
Igor Starinskij48.
Sequencing and analysis, and Software and analysis tools:
Chris J Illingworth47, Chris Jackson47, Marina Gourtovaia99, Gerry Tonkin-Hill99, Kevin Lewis99, Jaime M Tovar-Corona99, Keith James99, Laura Baxter94, Mohammad T. Alam94, Richard J Orton48, Joseph Hughes48, Sreenu Vattipally48, Manon Ragonnet-Cronin39, Fabricia F. Nascimento39, David Jorgensen39, Olivia Boyd39, Lily Geidelberg39, Alex E Zarebski23, Jayna Raghwani23, Moritz UG Kraemer23, Joel Southgate10, 69, Benjamin B Lindsey93 and Timothy M Freeman93.
Software and analysis tools, and Visualisation:
Jon-Paul Keatley99, Joshua B Singer48, Leonardo de Oliveira Martins70, Corin A Yeats14, Khalil Abudahab14, 114, Ben EW Taylor14, 114 and Mirko Menegazzo14.
Leadership and supervision:
John Danesh99, Wendy Hogsden46, Sahar Eldirdiri21, Anita Kenyon21, Jenifer Mason43, Trevor I Robinson43, Alison Holmes38, 103, James Price38, 103, John A Hartley82, Tanya Curran3, Alison E Mather70, Giri Shankar69, Rachel Jones69, Robin Howe69 and Sian Morgan9.
Metadata curation:
Elizabeth Wastenge53, Michael R Chapman34, 88, 99, Siddharth Mookerjee38, 103, Rachael Stanley54, Wendy Smith15, Timothy Peto59, David Eyre59, Derrick Crook59, Gabrielle Vernet33, Christine Kitchen10, Huw Gulliver10, Ian Merrick10, Martyn Guest10, Robert Munn10, Declan T Bradley63, 72 and Tim Wyatt63.
Project administration:
Charlotte Beaver99, Luke Foulser99, Sophie Palmer88, Carol M Churcher88, Ellena Brooks88, Kim S Smith88, Katerina Galai88, Georgina M McManus88, Frances Bolt38, 103, Francesc Coll19, Lizzie Meadows70, Stephen W Attwood23, Alisha Davies69, Elen De Lacy69, Fatima Downing69, Sue Edwards69, Garry P Scarlett76, Sarah Jeremiah83 and Nikki Smith93.
Samples and logistics:
Danielle Leek88, Sushmita Sridhar88, 99, Sally Forrest88, Claire Cormie88, Harmeet K Gill88, Joana Dias88, Ellen E Higginson88, Mailis Maes88, Jamie Young88, Michelle Wantoch7, Sanger Covid Team (www.sanger.ac.uk/covid-team)99, Dorota Jamrozy99, Stephanie Lo99, Minal Patel99, Verity Hill90, Claire M Bewshea91, Sian Ellard73, 91, Cressida Auckland73, Ian Harrison66, Chloe Bishop66, Vicki Chalker66, Alex Richter85, Andrew Beggs85, Angus Best86, Benita Percival86, Jeremy Mirza86, Oliver Megram86, Megan Mayhew86, Liam Crawford86, Fiona Ashcroft86, Emma Moles-Garcia86, Nicola Cumley86, Richard Hopes64, Patawee Asamaphan48, Marc O Niebel48, Rory N Gunson100, Amanda Bradley52, Alasdair Maclean52, Guy Mollett52, Rachel Blacow52, Paul Bird16, Thomas Helmer16, Karlie Fallon16, Julian Tang16, Antony D Hale49, Louissa R Macfarlane-Smith49, Katherine L Harper49, Holli Carden49, Nicholas W Machin45, 64, Kathryn A Jackson92, Shazaad S Y Ahmad45, 64, Ryan P George45, Lance Turtle92, Elaine O’Toole43, Joanne Watts43, Cassie Breen43, Angela Cowell43, Adela Alcolea-Medina32, 96, Themoula Charalampous12, 42, Amita Patel11, Lisa J Levett35, Judith Heaney35, Aileen Rowan39, Graham P Taylor39, Divya Shah30, Laura Atkinson30, Jack CD Lee30, Adam P Westhorpe82, Riaz Jannoo82, Helen L Lowe82, Angeliki Karamani82, Leah Ensell82, Wendy Chatterton35, Monika Pusok35, Ashok Dadrah75, Amanda Symmonds75, Graciela Sluga44, Zoltan Molnar72, Paul Baker79, Stephen Bonner79, Sarah Essex79, Edward Barton56, Debra Padgett56, Garren Scott56, Jane Greenaway57, Brendan AI Payne50, Shirelle Burton-Fanning50, Sheila Waugh50, Veena Raviprakash17, Nicola Sheriff17, Victoria Blakey17, Lesley-Anne Williams17, Jonathan Moore27, Susanne Stonehouse27, Louise Smith55, Rose K Davidson89, Luke Bedford26, Lindsay Coupland54, Victoria Wright18, Joseph G Chappell97, Theocharis Tsoleridis97, Jonathan Ball97, Manjinder Khakh15, Vicki M Fleming15, Michelle M Lister15, Hannah C Howson-Wells15, Louise Berry15, Tim Boswell15, Amelia Joseph15, Iona Willingham15, Nichola Duckworth60, Sarah Walsh60, Emma Wise2 111, Nathan Moore2 111, Matilde Mori2 108, 111, Nick Cortes2 111, Stephen Kidd2 111, Rebecca Williams33, Laura Gifford69, Kelly Bicknell61, Sarah Wyllie61, Allyson Lloyd61, Robert Impey61, Cassandra S Malone6, Benjamin J Cogger6, Nick Levene62, Lynn Monaghan62, Alexander J Keeley93, David G Partridge78, 93, Mohammad Raza78, 93, Cariad Evans78, 93 and Kate Johnson78, 93.
Sequencing and analysis:
Emma Betteridge99, Ben W Farr99, Scott Goodwin99, Michael A Quail99, Carol Scott99, Lesley Shirley99, Scott AJ Thurston99, Diana Rajan99, Iraad F Bronner99, Louise Aigrain99, Nicholas M Redshaw99, Stefanie V Lensing99, Shane McCarthy99, Alex Makunin99, Carlos E Balcazar90, Michael D Gallagher90, Kathleen A Williamson90, Thomas D Stanton90, Michelle L Michelsen91, Joanna Warwick-Dugdale91, Robin Manley91, Audrey Farbos91, James W Harrison91, Christine M Sambles91, David J Studholme91, Angie Lackenby66, Tamyo Mbisa66, Steven Platt66, Shahjahan Miah66, David Bibby66, Carmen Manso66, Jonathan Hubb66, Gavin Dabrera66, Mary Ramsay66, Daniel Bradshaw66, Ulf Schaefer66, Natalie Groves66, Eileen Gallagher66, David Lee66, David Williams66, Nicholas Ellaby66, Hassan Hartman66, Nikos Manesis66, Vineet Patel66, Juan Ledesma67, Katherine A Twohig67, Elias Allara64, 88, Clare Pearson64, 88, Jeffrey K. J. Cheng94, Hannah E. Bridgewater94, Lucy R. Frost94, Grace Taylor-Joyce94, Paul E Brown94, Lily Tong48, Alice Broos48, Daniel Mair48, Jenna Nichols48, Stephen N Carmichael48, Katherine L Smollett40, Kyriaki Nomikou48, Elihu Aranday-Cortes48, Natasha Johnson48, Seema Nickbakhsh48, 68, Edith E Vamos92, Margaret Hughes92, Lucille Rainbow92, Richard Eccles92, Charlotte Nelson92, Mark Whitehead92, Richard Gregory92, Matthew Gemmell92, Claudia Wierzbicki92, Hermione J Webster92, Chloe L Fisher28, Adrian W Signell20, Gilberto Betancor20, Harry D Wilson20, Gaia Nebbia12, Flavia Flaviani31, Alberto C Cerda96, Tammy V Merrill96, Rebekah E Wilson96, Marius Cotic82, Nadua Bayzid82, Thomas Thompson72, Erwan Acheson72, Steven Rushton51, Sarah O’Brien51, David J Baker70, Steven Rudder70, Alp Aydin70, Fei Sang18, Johnny Debebe18, Sarah Francois23, Tetyana I Vasylyeva23, Marina Escalera Zamudio23, Bernardo Gutierrez23, Angela Marchbank10, Joshua Maksimovic9, Karla Spellman9, Kathryn McCluggage9, Mari Morgan69, Robert Beer9, Safiah Afifi9, Trudy Workman10, William Fuller10, Catherine Bresner10, Adrienn Angyal93, Luke R Green93, Paul J Parsons93, Rachel M Tucker93, Rebecca Brown93 and Max Whiteley93.
Software and analysis tools:
James Bonfield99, Christoph Puethe99, Andrew Whitwham99, Jennifier Liddle99, Will Rowe41, Igor Siveroni39, Thanh Le-Viet70 and Amy Gaskin69.
Visualisation:
Rob Johnson39.
1 Barking, Havering and Redbridge University Hospitals NHS Trust, 2 Basingstoke Hospital, 3 Belfast Health & Social Care Trust, 4 Betsi Cadwaladr University Health Board, 5 Big Data Institute, Nuffield Department of Medicine, University of Oxford, 6 Brighton and Sussex University Hospitals NHS Trust, 7 Cambridge Stem Cell Institute, University of Cambridge, 8 Cambridge University Hospitals NHS Foundation Trust, 9 Cardiff and Vale University Health Board, 10 Cardiff University, 11 Centre for Clinical Infection & Diagnostics Research, St. Thomas’ Hospital and Kings College London, 12 Centre for Clinical Infection and Diagnostics Research, Department of Infectious Diseases, Guy’s and St Thomas’ NHS Foundation Trust, 13 Centre for Enzyme Innovation, University of Portsmouth (PORT), 14 Centre for Genomic Pathogen Surveillance, University of Oxford, 15 Clinical Microbiology Department, Queens Medical Centre, 16 Clinical Microbiology, University Hospitals of Leicester NHS Trust, 17 County Durham and Darlington NHS Foundation Trust, 18 Deep Seq, School of Life Sciences, Queens Medical Centre, University of Nottingham, 19 Department of Infection Biology, Faculty of Infectious & Tropical Diseases, London School of Hygiene & Tropical Medicine, 20 Department of Infectious Diseases, King’s College London, 21 Department of Microbiology, Kettering General Hospital, 22 Departments of Infectious Diseases and Microbiology, Cambridge University Hospitals NHS Foundation Trust; Cambridge, UK, 23 Department of Zoology, University of Oxford, 24 Division of Virology, Department of Pathology, University of Cambridge, 25 East Kent Hospitals University NHS Foundation Trust, 26 East Suffolk and North Essex NHS Foundation Trust, 27 Gateshead Health NHS Foundation Trust, 28 Genomics Innovation Unit, Guy’s and St. Thomas’ NHS Foundation Trust, 29 Gloucestershire Hospitals NHS Foundation Trust, 30 Great Ormond Street Hospital for Children NHS Foundation Trust, 31 Guy’s and St. Thomas’ BRC, 32 Guy’s and St. Thomas’ Hospitals, 33 Hampshire Hospitals NHS Foundation Trust, 34 Health Data Research UK Cambridge, 35 Health Services Laboratories, 36 Heartlands Hospital, Birmingham, 37 Hub for Biotechnology in the Built Environment, Northumbria University, 38 Imperial College Hospitals NHS Trust, 39 Imperial College London, 40 Institute of Biodiversity, Animal Health & Comparative Medicine, 41 Institute of Microbiology and Infection, University of Birmingham, 42 King’s College London, 43 Liverpool Clinical Laboratories, 44 Maidstone and Tunbridge Wells NHS Trust, 45 Manchester University NHS Foundation Trust, 46 Microbiology Department, Wye Valley NHS Trust, Hereford, 47 MRC Biostatistics Unit, University of Cambridge, 48 MRC-University of Glasgow Centre for Virus Research, 49 National Infection Service, PHE and Leeds Teaching Hospitals Trust, 50 Newcastle Hospitals NHS Foundation Trust, 51 Newcastle University, 52 NHS Greater Glasgow and Clyde, 53 NHS Lothian, 54 Norfolk and Norwich University Hospital, 55 Norfolk County Council, 56 North Cumbria Integrated Care NHS Foundation Trust, 57 North Tees and Hartlepool NHS Foundation Trust, 58 Northumbria University, 59 Oxford University Hospitals NHS Foundation Trust, 60 PathLinks, Northern Lincolnshire & Goole NHS Foundation Trust, 61 Portsmouth Hospitals University NHS Trust, 62 Princess Alexandra Hospital Microbiology Dept., 63 Public Health Agency, 64 Public Health England, 65 Public Health England, Clinical Microbiology and Public Health Laboratory, Cambridge, UK, 66 Public Health England, Colindale, 67 Public Health England, Colindale, 68 Public Health Scotland, 69 Public Health Wales NHS Trust, 70 Quadram Institute Bioscience, 71 Queen Elizabeth Hospital, 72 Queen’s University Belfast, 73 Royal Devon and Exeter NHS Foundation Trust, 74 Royal Free NHS Trust, 75 Sandwell and West Birmingham NHS Trust, 76 School of Biological Sciences, University of Portsmouth (PORT), 77 School of Pharmacy and Biomedical Sciences, University of Portsmouth (PORT), 78 Sheffield Teaching Hospitals, 79 South Tees Hospitals NHS Foundation Trust, 80 Swansea University, 81 University Hospitals Southampton NHS Foundation Trust, 82 University College London, 83 University Hospital Southampton NHS Foundation Trust, 84 University Hospitals Coventry and Warwickshire, 85 University of Birmingham, 86 University of Birmingham Turnkey Laboratory, 87 University of Brighton, 88 University of Cambridge, 89 University of East Anglia, 90 University of Edinburgh, 91 University of Exeter, 92 University of Liverpool, 93 University of Sheffield, 94 University of Warwick, 95 University of Cambridge, 96 Viapath, Guy’s and St Thomas’ NHS Foundation Trust, and King’s College Hospital NHS Foundation Trust, 97 Virology, School of Life Sciences, Queens Medical Centre, University of Nottingham, 98 Wellcome Centre for Human Genetics, Nuffield Department of Medicine, University of Oxford, 99 Wellcome Sanger Institute, 100 West of Scotland Specialist Virology Centre, NHS Greater Glasgow and Clyde, 101 Department of Medicine, University of Cambridge, 102 Ministry of Health, Sri Lanka, 103 NIHR Health Protection Research Unit in HCAI and AMR, Imperial College London, 104 North West London Pathology, 105 NU-OMICS, Northumbria University, 106 University of Kent, 107 University of Oxford, 108 University of Southampton, 109 University of Southampton School of Health Sciences, 110 University of Southampton School of Medicine, 111 University of Surrey, 112 Warwick Medical School and Institute of Precision Diagnostics, Pathology, UHCW NHS Trust, 113 Wellcome Africa Health Research Institute Durban and 114 Wellcome Genome Campus.
Acknowledgments
RKG is supported by a Wellcome Trust Senior Fellowship in Clinical Science (WT108082AIA). COG-UK is supported by funding from the Medical Research Council (MRC) part of UK Research & Innovation (UKRI), the National Institute of Health Research (NIHR) and Genome Research Limited, operating as the Wellcome Sanger Institute. This study was supported by the Cambridge NIHRB Biomedical Research Centre. SAK is supported by the Bill and Melinda Gates Foundation via PANGEA grant: OPP1175094. We would like to thank Paul Lehner for Calu-3 cells. We thank Wendy Barclay and Thomas Peacock for helpful discussions and the Geno2pheno UK consortium. This study was also partly funded by Rosetrees Trust.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}