

Abstract
Heart failure (HF) is one of the leading causes of death world-wide and is associated with cardiac metabolic perturbations. Human Type 2 Berardinelli-Seip Congenital Lipodystrophy (BSCL2) disease is caused by mutations in the BSCL2 gene. Global lipodystrophic Bscl2−/− mice exhibit hypertrophic cardiomyopathy. Whether BSCL2 plays a direct role in regulating cardiac substrate metabolism and/or contractile function remains unknown. Here we show that mice with cardiac-specific deletion of Bscl2 (Bscl2cKO) developed dilated HF. Myocardial BSCL2 deletion led to elevated ATGL expression and FA oxidation (FAO) along with reduced cardiac lipid contents. Cardiac dysfunction in Bscl2cKO mice was independent of mitochondrial dysfunction and oxidative stress, but associated with decreased metabolic reserve and ATP levels. Importantly, heart failure in Bscl2cKO mice could be partially reversed by pharmacological inhibition of FAO, or prevented by high fat diet (HFD) feeding. Lipidomic analysis further identified markedly reduced glycerolipids, glycerophospholipids, NEFA and acylcarnitines in Bscl2cKO hearts, which were partially normalized by FAO inhibition or HFD. Our study reveals a new form of HF with excessive lipid catabolism, and identifies a crucial cardiomyocyte-specific role of BSCL2 in controlling cardiac lipid catabolism, energy state and contractile function. It also provides novel insights into metabolically treating energy-starved HF using FAO inhibitor or HFD.
Introduction
Heart failure (HF) is one of the leading causes of morbidity and mortality worldwide1. In healthy individuals, the heart exhibits striking metabolic flexibility, being capable of utilizing carbohydrate, lipid, amino acids and/or ketone bodies to meet energetic demands, cellular constituent turnover, and metabolic signaling; these processes are critical for maintenance of mechanical work. Oxidation of fatty acids (FAs) predominates, accounting for 60-70% of myocardial oxygen consumption2, 3. In patients with hypertrophied and failing hearts, derangements of substrate utilization include an increased reliance on glycolysis concomitant with an overall reduced oxidative metabolism4. The severely failing heart usually demonstrates a lower concentration of ATP, supporting the concept that energy starvation contributes significantly to the pathogenesis of HF5. Indeed, there is a striking correlation between cardiac energetic status and survival in HF patients6. Thus, targeting metabolic processes in the heart may represent a promising way to develop new therapeutic approaches for HF.
Cardiac lipid metabolism is precisely controlled to maintain a balance between FA uptake, triglyceride (TG) synthesis, TG hydrolysis, and FA oxidation (FAO)7. Imbalances in these processes are commonly seen in obese and diabetic patients (as well as animal models), which are associated with cardiac steatosis and contractile dysfunction7. Recent preclinical and clinical evidence argues for an important role of adipose triglyceride lipase (ATGL)-mediated cardiac lipolysis in promoting mitochondrial FAO and ATP production thus contractile function8. Constitutive Atgl−/− mice develop severe cardiac steatosis and HF, associated with a high mortality9. Conversely, cardiomyocyte-specific overexpression of ATGL maintains normal cardiac function in lean mice, and reduces cardiac TG content and improves cardiac function during diabetes and obesity10, 11. The precise mechanisms linking cardiac lipid accumulation with contractile dysfunction remain obscure.
Berardinelli-Seip Congenital Lipodystrophy 2 (BSCL2, a.k.a. SEIPIN) is a highly conserved endoplasmic reticulum (ER) protein expressed in most tissues, with the highest levels in testis, neuronal and adipose tissue12. Global Bscl2-deficient (Bscl2−/−) mice recapitulate human BSCL2 disease, exhibiting congenital lipodystrophy and severe insulin resistance13–15. Various molecular functions of BSCL2 have been proposed, ranging from regulating lipid droplet (LD) biogenesis16, 17 to mitochondrial metabolism18, 19. Human and Drosophila BSCL2 assembles as an undecamer or dodecamer respectively and play crucial roles in lipid transfer and/or LD formation20, 21. BSCL2 has been shown to interact with 1-acylglycerol-3-phosphate O-acyltransferase 2 (AGPAT2)22, LIPIN123, glycerol-3-phosphate acyltransferase 3 (GPAT3)24 and Promethin25, 26. We and others demonstrate that BSCL2 plays a key role in regulating cyclic AMP/protein kinase A (cAMP/PKA) mediated TG lipolysis essential for adipocyte differentiation and maintenance13, 15. Recently, we reported that Bscl2−/− mice develop hypertrophic cardiomyopathy associated with reduced cardiac steatosis27. Despite the relatively low expression of BSCL2 in murine hearts28, whether BSCL2 plays a cell-autonomous role in modulating cardiac lipid metabolism and function has not been fully addressed.
The present study highlights BSCL2 as a key player governing cardiac lipid metabolism. Cardiomyocyte-specific deletion of BSCL2 enhanced ATGL expression and FAO, resulting in markedly reduced cardiac lipid reserve, associated with compromised ATP production and contractile dysfunction. Inhibition of FAO or supplying FAs by high fat diet (HFD) feeding partially alleviated cardiac energetic stress and augmented contractility. These findings improve our understanding of how perturbations in lipid utilization/storage contribute towards HF development.
Results
Myocardial deletion of BSCL2 induces dilated heart failure
Previously we demonstrated lipodystrophic Bscl2−/− mice develop cardiac dysfunction 27. In order to interrogate the specific role of cardiac BSCL2, we generated a mouse model with a cardiomyocyte-specific deletion of BSCL2 (Bscl2cKO) via Myh6-Cre. Gene expression analysis confirmed an approximate 75% reduction of Bscl2 specifically in cardiac muscle but not in liver and skeletal muscle of Bscl2cKO mice (Supplemental Figure S1A). Current available antibodies were not sensitive enough to detect endogenous BSCL2 protein in murine heart tissue, preventing us from confirming the cardiac deletion of BSCL2 at the protein level.
We next determined the impact of cardiomyocyte-specific deletion of Bscl2 on cardiac function, in comparison to two distinct control groups [Cre-; Bscl2f/f (designated as Ctrl) and Cre+; Bscl2w/w]. No changes were observed in total body weights at either 3 months or 6 months of age between experimental groups (Supplemental Table S1). At 3 months old, there were also no significant differences in ventricle mass in proportion to body weight (Figure 1A) or tibia length (Supplemental Figure S1B). By echocardiography, 3-month-old mice did not display noticeable changes in left ventricular post wall thickness at end systole (LVPWs) (Figure 1B), left ventricular anterior thickness at end systole (LVAWs) (Figure 1C), left ventricular internal diameters at end systole (LVIDs) (Figure 1D), ejection fraction (Figure 1E) and fractional shortening (Figure 1F) between the three experimental groups. When mice aged to 6 months old, ratios of ventricle mass to body weight (Figure 1A) and tibia length (Supplemental Figure S1B) were still comparable in all groups, suggesting no gross cardiac hypertrophy. In agreement with the previous report29, 6-month-old Cre+, Bscl2w/w mice demonstrated no differences in LVPWs, LVAWs, or LVIDs (Figure 1B-D), but exhibited a minor reduction in ejection fraction and fractional shortening (Figure 1E-F) compared with Ctrl mice. In contrast, 6-month-old Bscl2cKO mice exhibited decreased wall thickness and increased dilation at both systoles and diastoles (Figure 1B-D and Supplemental Table S1), along with marked reductions in contractility compared with Ctrl and Cre+, Bscl2w/w mice (Figure 1E-G). A 4 chamber analysis confirmed mild dilatation in 6-month-old Bscl2cKO mice (Figure 1H). Histological analysis of ventricular cross-sectional area showed no evidence of abnormal cardiomyocyte morphology compared with both control groups (Supplemental Figure S1C). In addition, despite similar induction of atrium natriuretic peptide (Nppa) in hearts of Cre+; Bscl2w/w and Bscl2cKO mice when compared with Ctrl mice, we observed a greater induction of brain natriuretic peptide (Nppb) and growth differentiation factor 15 (Gdf15), biomarkers for stressed myocardium, in 6-month-old Bscl2cKO hearts versus Ctrl hearts (Figure 1I). The expression of adult Myh6 gene expression in Bscl2cKO hearts was further downregulated relative to Ctrl hearts (Figure 1I). Moreover, the BSCL2 deletion-induced cardiac dysfunction was not accompanied by excessive myocardial fibrosis (assessed by trichrome staining of collagen deposition) in hearts of Bscl2cKO mice (Supplemental Figure S1D). Together, these data suggest that cardiomyocyte-specific BSCL2 deletion leads to dilated heart failure independent of the long-term expression of transgene Myh6-Cre.
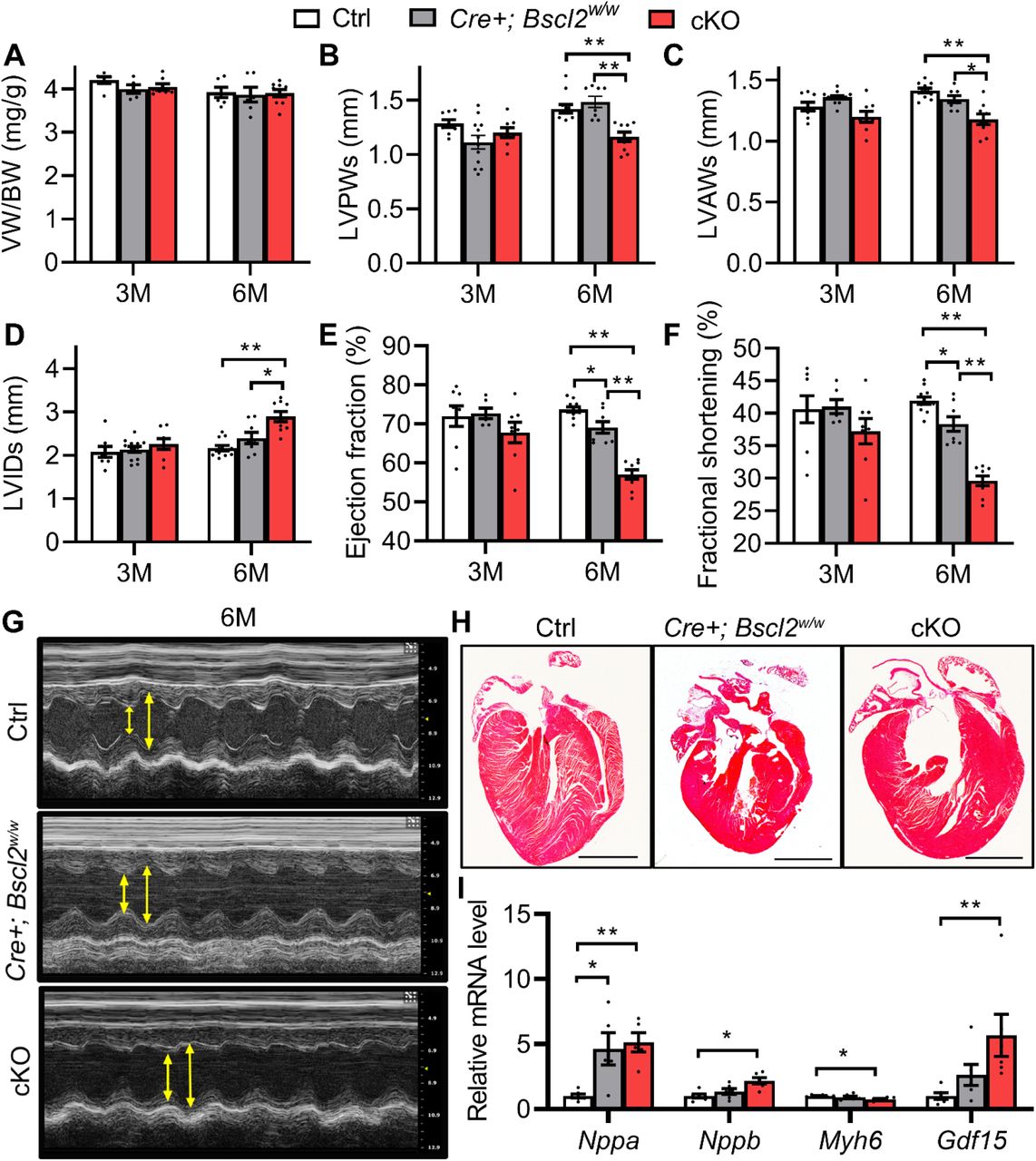
(A) Ventricle weight (VW) normalized to body weight (BW) in 3-month-old (3M) and 6-month-old (6M) male Cre-; Bscl2f/f (Ctrl), Cre+; Bscl2w/w, and Cre+; Bscl2f/f (cKO) mice. 3M old: Ctrl, n=6; Cre+; Bscl2w/w, n=5; cKO, n=6. 6M old: Ctrl, n=6; Cre+; Bscl2w/w, n=6; cKO, n=9. (B) left ventricle post wall thickness at end systole (LVPWs, mm); (C) left ventricle anterior wall thickness at end systole (LVAWs, mm); (D) left ventricle internal diameter at end systole (LVIDs, mm); (E) ejection fraction (%) and (F) fractional shortening (%) in 3M and 6M old male mice. 3M old: Ctrl, n=8; Cre+; Bscl2w/w, n=8; cKO, n=12. 6M old: Ctrl, n=11; Cre+; Bscl2w/w, n=8; cKO, n=9. (G) Representative echocardiography and (H)4 chamber view of 6-month-old mice. Scale bar: 2 mm. (I) RT-PCR analysis of cardiac stress genes in ventricles of 6-month-old male mice. n=6 per group. *: P< 0.05; **: P< 0.005. Two-way ANOVA followed by Tukey’s post-hoc tests.
Myocardial-specific deletion of BSCL2 causes elevated TG turnover and FAO preceding functional decline
Although we have previously demonstrated reduced cardiac TG content, increased ATGL and FAO in lipodystrophic Bscl2−/− hearts27, it remains to be determined whether this was due to a direct loss of BSCL2 from cardiomyocytes. Indeed, we found ventricular TG content in 3-month-old Bscl2cKO mice was reduced by 57% as compared to Ctrl mice (Figure 2A). Bscl2cKO hearts displayed increased (≈2.5-fold) ATGL, but not HSL, protein expression compared with that of Ctrl hearts (Figure 2B-C). Such changes were not attributed to alterations in the transcript level of Pnpla2 (Supplemental Figure S2A), supporting its post-transcriptional regulation. Isolated adult Bscl2cKO cardiomyocytes displayed similar upregulation of ATGL, but not HSL, compared with Ctrl cells (Figure 2D-E), further supporting a cell-autonomous effect of BSCL2 deletion on ATGL expression. Ctrl and Bscl2cKO cardiomyocytes responded similarly to the stimulation of isoproterenol in terms of PKA-mediated phospholamban (PL) phosphorylation (Supplemental Figure S2B). Analysis of whole heart lysates also revealed comparable basal PKA-mediated substrate phosphorylation between Ctrl and Bscl2cKO mice (Supplemental Figure S2C). These data suggest that cardiac-specific deletion of BSCL2 did not affect cAMP/PKA signaling in hearts, different from what we previously observed in BSCL2-deleted adipose tissue30–32.
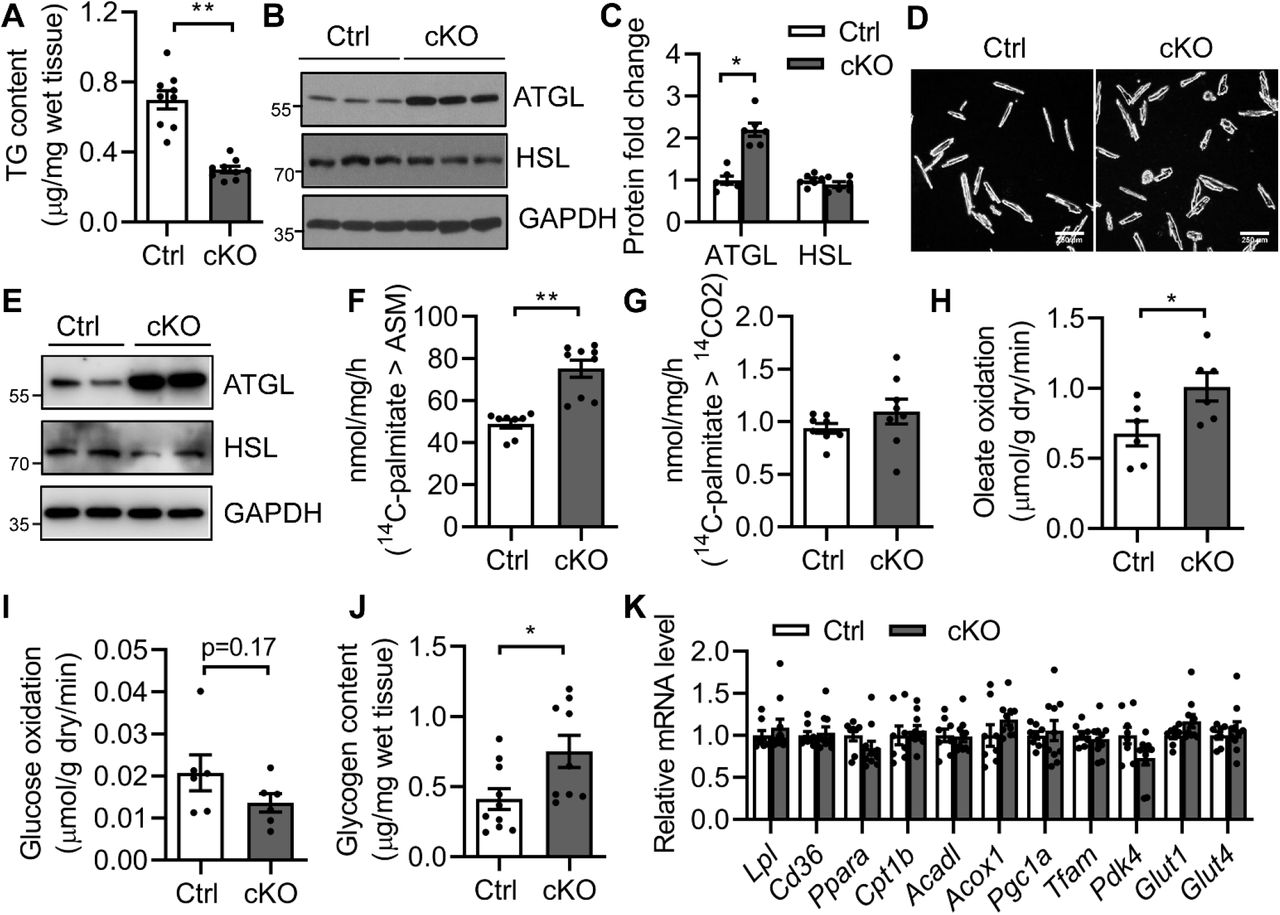
(A) Ventricular TG content as normalized to wet tissue weight. n=9 per group. (B-C) Representative Western blotting and quantification of lipolytic proteins in heart homogenates. n=6 per group. (D-E) Isolated adult cardiomyocytes and representative Western blotting in duplicates from two independent experiments. (F-G) Acid soluble metabolites (ASM) and CO2 production after incubating heart crude mitochondrial fraction with 14C-palmitate. Ctrl, n=8; cKO, n=9. (H) Oleate oxidation rate; (I) glucose oxidation rate in ex vivo perfused working hearts. n=6 per group. (J) Glycogen content. Ctrl, n=10; cKO, n=9. (K) RT-PCR analysis of genes involved in fatty acid metabolism, mitochondrial biogenesis and glucose metabolism. Ctrl, n=8; cKO, n=10. All experiments used 3-month-old male Cre-; Bscl2f/f (Ctrl) and Bscl2cKO (cKO) mice. *: P< 0.05; **: P<0.005, unpaired t tests (parametric).
We next investigated FAO in Bscl2cKO hearts. When incubating heart homogenates with 14C-palmitate, 3-month-old Bscl2cKO mice demonstrated elevated release of 14C-labeled acid soluble metabolites (ASM) (Figure 2F) despite a lack of change in 14CO2 production (Figure 2G). To systemically assess cardiac substrate metabolism preceding cardiac functional decline, hearts from 3-month-old male Ctrl and Bscl2cKO mice were subject to ex vivo perfusions in working mode using radiolabeled substrates. We found no differences in heart rates between two genotypes (Supplemental Figure S2D). The Bscl2cKO hearts demonstrated a 33% increase in oleate oxidation (Figure 2H) concomitant with a tendency of lower glucose oxidation (Figure 2I). The cardiac oxygen consumption, cardiac efficiency as well as cardiac power (Supplemental Figure S2E-G, respectively) were all comparable between 3-month-old Ctrl and Bscl2cKO hearts, suggesting maintained cardiac function at this age. Interestingly, the baseline glycogen content in 3-month-old Bscl2cKO hearts was increased by 83% (Figure 2J), potentially resulting from a tendency toward lower glucose oxidation rate in Bscl2cKO hearts. In spite of enhanced FAO and glycogen accumulation, Bscl2cKO hearts demonstrated no changes in the expression of genes involved in FA uptake (Lpl, Cd36), mitochondrial and peroxisomal β-oxidation (Pparα, Cpt1β, Acadl, Acox1), mitochondrial biogenesis (Pgc1α, Tfam), and glucose metabolism (Pdk4, Glut1, Glut4) (Figure 2L). Lack of changes in mitochondrial biogenesis was further confirmed by the similar ratios of mitochondrial DNA (mtDNA)-encoded mt-Rnr2 to nuclear DNA (nDNA)-encoded Hk2 intron 9 (Supplemental Figure S2H) and protein levels of each of the electron transport chain (ETC) complexes between heart lysates of 3-month-old Ctrl and Bscl2cKO mice (Supplemental Figure S2I). Together, these data clearly suggest that cardiomyocyte-specific BSCL2 deficiency results in higher rates of cardiac TG turnover and FAO independent of transcriptional changes of mitochondrial and extra-mitochondrial metabolic genes.
Chronic derangements in myocardial FAO leads to massive lipid remodeling and reduced endogenous substrates in Bscl2cKO hearts
To identify mechanisms underlying the progressive development of HF, we performed untargeted lipidomic analyses of ventricles from 6-month-old Ctrl and Bscl2cKO mice. Total normalized lipid ion abundances identified in Bscl2cKO hearts were reduced by about 45% (Figure 3A). Heatmap analysis revealed massive reductions in the absolute levels of five broadly classified lipid classes defined by the Lipid MAPS consortium; i.e. glycerophospholipids, fatty acyls [mainly nonesterified fatty acids (NEFA)], sphingolipids, sterol lipids and glycerolipids in Bscl2cKO mice (Figure 3B and Supplemental Excel 1). When comparing the % distributions of these five lipid classes, the proportions of glycerophospholipids, sphingolipids and sterol lipids were significantly higher in Bscl2cKO hearts as compared to Ctrl hearts (Figure 3C). The proportions of NEFA were relatively comparable between two genotypes, while the proportion of glycerolipids was markedly reduced by 58% in Bscl2cKO hearts (Figure 3C). Analysis of the absolute levels of glycerolipids revealed 78%, 50% and 75% reductions in TG, diacylglycerol (DAG) and monoacylglycerol (MAG), respectively in Bscl2cKO hearts (Figure 3D). The absolute levels of NEFA and total acylcarnitines (ACs) were significantly lower in Bscl2cKO hearts (Figure 3E). Specifically, the abundances of the major long-chain ACs (AC16:0, AC16:1, AC18:0, AC18:1, AC18:2) were all reduced by approximately 70% (Figure 3F). These data suggest myocardial BSCL2 deletion results in a dramatic remodeling of lipid composition highlighted by reduced levels of energy providing substrates indicative of impairment of cardiac metabolic reserve.

(A) The total lipid ion abundance normalized to tissue weight. (B) Heatmap of lipid species including glycerophospholipids [i.e. phosphatidylcholine (PC), phosphatidylethanolamine (PE), phosphatidylserine (PS), phosphatidylinositol (PI), phosphatidylglycerol (PG), cardiolipin (CL), phosphatidic acid (PA), and acylcarnitines (AC)], nonesterified free fatty acids (NEFA), sphingolipids, sterols and glycerolipids based on Z-scores calculated from the summed ion abundances normalized to tissue weight. (C) Pie chart representing the proportional distribution of summed ion abundances of glycerolipid, glycerophospholipid, sphingolipid, NEFA, and sterol lipid classes. (D-F) Comparison of the total normalized ion abundances for (D) glycerolipids including TG, DAG and MAG, (E) NEFA and ACs, and (F) specific AC and hydroxyl acylcarnitines (OHAC) species. Global lipidomic analysis of ventricles by shotgun mass spectrometry was performed in nonfasting 6-month-old male Cre-; Bscl2f/f (Ctrl) and Bscl2cKO (cKO) mice. n=4 per group with each pooled from 3 animals. *: P< 0.05; **: P< 0.005, unpaired t tests (nonparametric).
To exclude the effect of the chronic expression of Cre transgene on massive lipid remodeling in Bscl2cKO mice, we included both Ctrl and Cre+; Bscl2w/w mice as controls to compare their cardiac TG contents and protein expression at 6 months of age. As expected, no differences were observed between Ctrl and Cre+; Bscl2w/w hearts, and only Bscl2cKO hearts displayed reduced TG (Supplemental Figure S3A) and increased ATGL expression (Supplemental Figure S3B). In addition, we observed similar expression of PPARα and its target proteins (CD36, CPT1β) between three genotypes (Supplemental Figure S3B). Collectively, these data emphasize a BSCL2-specific regulation of lipid remodeling in Bscl2cKO hearts independent of transcriptional activation of PPARα.
Cardiomyopathy in 6-month-old Bscl2cKO mice is associated with energy deficiency independent of intrinsic mitochondrial dysfunction and oxidative stress
We next examined whether the massive lipid remodeling exerts maladaptive effects on mitochondrial function, leading to the development of HF in 6-month-old Bscl2cKO mice. Transmission electron microscopy images revealed a complete lack of LDs in 6-month-old Bscl2cKO hearts, in support of cardiac TG reduction (Figure 4A). Sarcomere arrangement, mitochondrial morphology and sizes, as well as mitochondria distribution were generally preserved in both Ctrl and Bscl2cKO hearts (Figure 4A). We performed respirational analysis of isolated mitochondria from hearts of Ctrl and Bscl2cKO mice in the presence of exogenous substrates (succinate), and identified similar basal and maximal oxygen consumption rates (Figure 4B). There were also no differences in coupled (ADP-driven) and uncoupled respiration between two genotypes, suggesting lack of obvious mitochondrial dysfunction (Figure 4C). In line with that, BSCL2 deletion did not affect the levels of marker proteins for mitochondrial ETC complexes expressed per μg total mitochondrial protein (Figure 4D). The absence of mitochondrial dysfunction and biogenesis was also confirmed in 6-month-old Bscl2cKO hearts as evidenced by similar expression of ETC complex proteins, PGC1α and mitochondrial stress marker Prohibitin compared with both Ctrl and Cre+; Bscl2w/w hearts (Supplemental Figure S4A). Despite an increased FAO in 3-month-old Bscl2cKO hearts, oxidation of 2’, 7’-dichlorofluorescein diacetate (DCFDA) was not augmented in heart extracts of 6-month-old Bscl2cKO mice (Supplemental Figure S4B). The level of lipid peroxides malondialdehyde (MDA) was even reduced in Bscl2cKO hearts compared with Ctrl hearts (Supplemental Figure S4C). Consistent with lack of oxidative stress, there were no differences in the expression of SOD1, SOD2 and Catalase in Bscl2cKO hearts relative to both Ctrl and Cre+; Bscl2w/w hearts (Supplemental Figure S4B). Together, these data suggest mitochondrial dysfunction and oxidative stress are unlikely to play a role in maladaptive cardiac remodeling and progression of HF in Bscl2cKO mice.
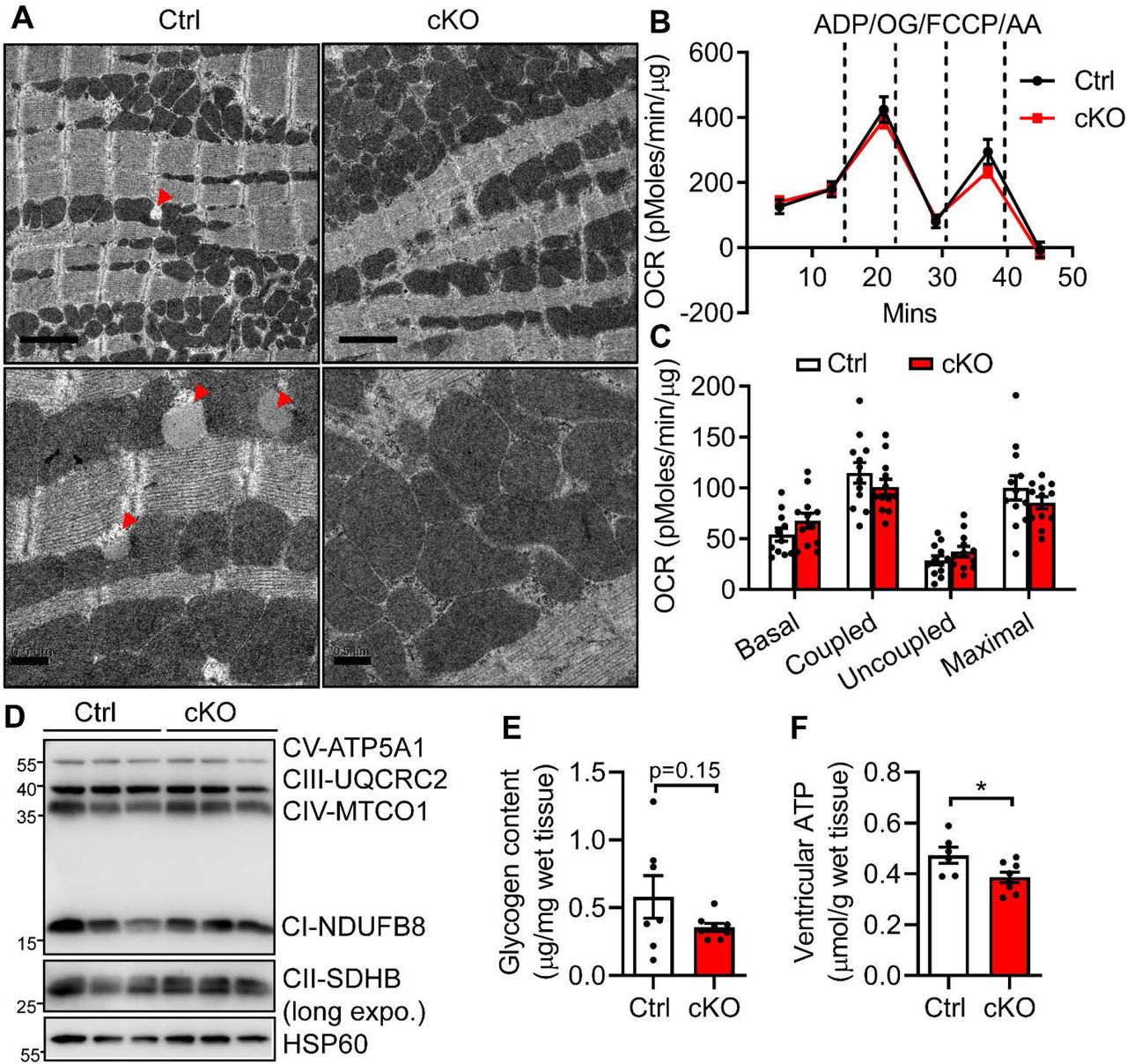
(A) Representative transmission electron micrographs of ventricles from 6-month-old Cre-; Bscl2f/f (Ctrl) and Bscl2cKO (cKO) mice. Red arrowheads: lipid droplets (LDs). Upper panels: scale bar = 2 μm; lower panels: scale bar = 0.5 μm. (B-C) Succinate/rotenone driven mitochondrial oxygen consumption rates (OCR) were measured by Seahorse XF24 analyzer with sequential addition of ADP, oligomycin (OG), FCCP and antimycin A (AA). Basal, coupled, uncoupled and maximal mitochondrial respiration were shown in (C). n=12 per group. (D) Representative Western blotting in isolated mitochondria from ventricles of 6-month-old Ctrl and Bscl2cKO mice with mitochondrial HSP60 as controls. n=3 per group. Three independent experiments. (E) Ventricular glycogen contents, n=7 per group, and (F) ventricular ATP contents in nonfasting 6-month-old male Ctrl and Bscl2cKO mice. Ctrl, n=6; cKO, n=8. *: P< 0.05, unpaired t tests (parametric).
Interestingly, in contrast to 3-month-old Bscl2cKO mice (Figure 2J), we identified a tendency toward lower glycogen content in 6-month-old Bscl2cKO hearts (Figure 4E). There were no differences in the plasma concentrations of glucose and lipid substrates (TG, cholesterol, NEFA and glycerol) between 6 month-old Ctrl and Bscl2cKO mice (Supplemental Table S2). Despite the availability of extracellular substrates, the ventricular ATP content was significantly lower in hearts of 6-month-old Bscl2cKO mice (Figure 4F), in line with its reduced metabolic reserve (i.e. lipid and glycogen stores). Collectively, our data suggest chronic higher FAO caused by BSCL2 deletion in hearts leads to cardiac nutritional deprivation which is likely responsible for the cardiac energetic and contractile failure in 6-month-old Bscl2cKO mice.
Inhibition of FAO reprograms cardiac lipidome and partially rescues cardiac dysfunction in Bscl2cKO mice
Since cardiac dysfunction in Bscl2cKO mice is associated with higher FAO, we treated mice with TMZ, a specific 3-Ketoacyl-CoA thiolase (3-KAT) inhibitor that inhibits FAO33, for up to 6 weeks starting at 6-month-old when Bscl2cKO mice already developed cardiac dysfunction. TMZ did not alter body weight (Supplemental Figure S5A) or liver weight (Supplemental Figure S5B), but tended to increase white fat mass in both genotypes (Supplemental Figure S5C). TMZ elevated circulating cholesterol levels in both Ctrl and Bscl2cKO mice, but there were no differences in plasma glucose, NEFA, glycerol or TG levels in the experimental groups (Supplemental Table S3). Ctrl and Bscl2cKO mice also demonstrated similar ventricle to body weight (Supplemental Figure S5D) and tibia length (Figure 5A) ratios after TMZ treatment. TMZ caused no changes in cardiac dilation and contractility of Ctrl mice, but was able to significantly improve wall thickness and cardiac contractility without notably normalizing dilatation of Bscl2cKO hearts (Figure 5B-D and Supplemental Table S3). Attenuation of cardiac dysfunction by TMZ was also evident in female Bscl2cKO mice (Supplemental Figure S5E-H).

6-month-old male Cre-; Bscl2f/f (Ctrl) and Bscl2cKO (cKO) mice received daily i.p. injection with PBS or trimetazidine (TMZ) at 15 mg/kg body weights (BW) for a total of 6 weeks. (A) Ratio of ventricle weight (VW) to tibia length (TL). Ctrl, n=5; cKO, n=9. (B) Left ventricle post wall thickness at end systole (LVPWs, mm); (C) left ventricular internal diameter in end systole (LVIDs); and (D) fractional shortening. Ctrl, n=6; cKO, n=9. (E) Heatmap of major lipid species based on Z-score calculated from the summed ion abundances normalized to milligram of wet tissue; (F-G) comparison of the total normalized ion abundances for (F) glycerolipids and NEFA as well as (G) ACs. n=3 per PBS-treated group. n=4 per TMZ-treated group. Each sample was pooled from 3 animals. (H) Ventricular glycogen. PBS-Ctrl, n=5; PBS-cKO, n=9. n=9 per TMZ-treated group. (I) Ventricular ATP contents normalized to gram of wet tissue. PBS-Ctrl, n=5; PBS-cKO, n=8. TMZ-Ctrl, n=5, TMZ-cKO, n=9. (J-K) Representative Western blotting and quantification of total cell extracts from ventricles of 6-month-old PBS and TMZ-treated Ctrl and Bscl2cKO mice. n=6 per group. Data were normalized to GAPDH with PBS-treated Ctrl set to 1. Two independent experiments. (L) RT-PCR analysis of cardiac stress genes in ventricles of 6-month-old PBS and TMZ-treated male mice. n=6 per group. *: P< 0.05; **: P< 0.005. Two-way ANOVA followed by Tukey’s post-hoc tests.
We performed another independent set of untargeted lipidomics to examine the effect of TMZ on cardiac lipidome. As indicated in Figure 5E, PBS-treated Bscl2cKO hearts recapitulated all changes in cardiac lipid contents as we previously observed in 6-month-old Bscl2cKO hearts (Figure 3). TMZ unexpectedly reduced the absolute levels of cardiac lipid contents in all categories except ACs in Ctrl hearts. In contrast, TMZ dramatically enhanced the accumulation of these lipid classes in Bscl2cKO hearts relative to PBS-treated Bscl2cKO hearts. Especially, the absolute levels of phospholipids, sphingolipids, and sterols were even greater in TMZ-treated Bscl2cKO than PBS-treated Ctrl hearts (Figure 5E and Supplemental Excel 2). Specifically, TMZ was able to markedly increase TG, NEFAs and ACs in Bscl2cKO hearts (Figure 5F-G). Moreover, TMZ slightly lowered glycogen in Ctrl hearts. However, it caused more glycogen accumulation in Bscl2cKO hearts (Figure 5H). Despite a significant upregulation of metabolic reserve, there was only a very minimal but non-significant improvement of ventricular ATP content in TMZ-treated Bscl2cKO hearts (Figure 5I). As expected, TMZ itself exerted no effect on ATGL upregulation in Bscl2cKO hearts, since it acts on FAO downstream of ATGL (Figure 5J). However, the expression of BDH1, a crucial enzyme responsible for cardiac ketolysis, was significantly higher in Bscl2cKO hearts after TMZ treatment (Figure 5J). There was also a trend for TMZ to attenuate the gene expression of cardiac stress markers in Bscl2cKO hearts (Figure 5K). Together, our data suggest inhibiting FAO in Bscl2cKO hearts is able to partially restore cardiac function, potentially through modifying cardiac metabolism.
HFD significantly improves cardiac energy substrates and prevents heart failure in Bscl2cKO mice
To gain further insight into the importance of metabolic reserve in energy-starved Bscl2cKO hearts, we also fed 3-month-old Ctrl and Bscl2cKO male mice a normal chow diet (NCD) or a 60% high fat diet (HFD) for a period of 3 months. By 6 months old, Ctrl and Bscl2cKO mice exhibited similar obese phenotype with comparable weight gain after HFD feeding (Supplemental Table S2). They exhibited similarly higher levels of plasma glucose, cholesterol, NEFA and glycerol relative to their NCD-fed counterparts, and there were no differences in serum TG concentrations in all four groups (Supplemental Table S2). HFD slightly increased the ventricle weight to tibia length ratios in both genotypes, indicating mild but comparable cardiac hypertrophy (Figure 6A). We found no significant changes in wall thickness, dilation and contractility in Ctrl mice after 3 months of HFD feeding (Figure 6B-D and Supplemental Table S2). However, HFD was able to increase LVPWs and LVAWs, and improve cardiac contractility of Bscl2cKO mice to the similar levels of HFD-fed Ctrl mice despite exerting no effect on dilation (Figure 6B-D and Supplemental Table S2).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
3-month-old Bscl2f/f (Ctrl) and Bscl2cKO (cKO) mice were fed with normal chow diet (NCD) or high fat diet (HFD, 60% fat calories) for a total of 3 months. (A) ratio of ventricle weight (VW) to tibial length (TL). Ctrl, n=5; cKO, n=9. (B) Left ventricle post wall thickness at end systole (LVPWs, mm); (C) left ventricular internal diameter in end systole (LVIDs); and (D) fractional shortening. NCD-Ctrl, n=9; NCD-cKO, n=12; HFD-Ctrl, n=12; HFD-cKO, n=16. (E) Heatmap of major lipid species based on Z-score calculated from summed ion abundances normalized to milligram of wet tissue; (F-G) comparison of the total normalized ion abundances for (F) glycerolipids and NEFA as well as (G) ACs. n=3 per NCD group. n=4 per HFD group. Each sample was pooled from 3 animals. (H) Myocardial ATP content as normalized to gram of wet tissue. NCD-Ctrl, n=8; NCD-cKO, n=8; HFD-Ctrl, n=7; HFD-cKO, n=11. (I) Representative Western blotting of total cell extracts from ventricles of Ctrl and Bscl2cKO mice. n=3 per group. Two independent experiments. *: P< 0.05; **: P< 0.005. Two-way ANOVA followed by Tukey’s post-hoc tests.
We also determined cardiac lipidome of HFD-fed hearts with the above PBS-treated hearts as NCD controls. Interestingly, in Ctrl hearts, we found HFD reduced the levels of phospholipids (i.e. PC, PS, PI, PG, CL and PA) and sphingolipids, but preferentially increased the levels of sterols and glycerolipids when comparing with NCD (Figure 6E). The amounts of almost all lipid classes were increased in Bscl2cKO hearts after HFD feeding, albeit still lower than HFD-fed Ctrl hearts (Figure 6E and Supplemental Excel 2). Specifically, TG levels were greatly upregulated in both Ctrl and Bscl2cKO hearts after HFD feeding; whereas the levels of NEFA and ACs in Bscl2cKO hearts were increased to a lesser extent by HFD when compared with NCD (Figure 6F-G). Nevertheless, while Bscl2cKO hearts contained less ATP under NCD, the ventricular ATP contents in HFD-fed Bscl2cKO mice was almost restored to the levels of HFD-fed Ctrl hearts, suggesting improved cardiac energetics (Figure 6H). Immunoblot analysis revealed similar upregulation of ATGL in Bscl2cKO relative to Ctrl hearts regardless of diets (Figure 6I). Collectively, HFD could restore cardiac function by improving cardiac energetics via increasing energy supply in Bscl2cKO mice.
Discussion
In this study, we show that genetic deletion of BSCL2 in cardiomyocytes leads to dramatic cardiac lipid remodeling and dilated HF in mice. Mechanistically, cardiac BSCL2 ablation causes ATGL overexpression, excessive FAO and overt cardiac lipid remodeling. Increased lipid catabolism ultimately exhausts intramyocellular lipid and glycogen reserve and is likely responsible for the energetic and contractile failure in Bscl2cKO mice. Importantly, inhibiting FAO by promoting substrate switch or HFD feeding via increasing lipid supply alleviate cardiac dysfunction in Bscl2cKO mice. Our results thus identify a novel and indispensable role of BSCL2 in regulating a preferential oxidation of FAs from endogenous cardiac TG lipolysis which governs cardiac energetics and function.
BSCL2 deletion enhances cAMP/PKA triggered ATGL-mediated lipolysis and FAO in adipose tissue30–32. Similar to global Bscl2−/− hearts27, Bscl2cKO hearts exhibited higher ATGL expression associated with reduced TG contents and accelerated FAO, highlighting the cell-autonomous role of cardiac BSCL2 in lipid catabolism despite its low expression in hearts. Interestingly, Bscl2cKO mice developed more severe cardiomyopathy than global Bscl2−/− mice, suggesting differential pathological mechanisms. Lipodystrophic Bscl2−/− mice undergo mild hypertrophic cardiomyopathy, which can be attenuated by partially restoring fat mass and/or improving whole-body and cardiac insulin resistance27, 34. This suggests insulin resistance largely accounts for the pathophysiology of metabolic cardiomyopathy in lipodystrophic Bscl2−/− mice, resembling diabetic hearts. On the other hand, Bscl2cKO mice developed energy deficit-induced dilated HF which was independent of profound remodeling of structural changes (e.g. hypertrophy and fibrosis) (Figure 1) and insulin resistance (data not shown). We speculate that hyperphagia in lipodystrophic Bscl2−/− mice may actually provide more circulating lipid metabolites which protect BSCL2-deleted heart from cardiac substrate exhaustion and HF as we observed in Bscl2cKO mice. Notably, a previous report identified no differences in cardiac function of 55-60 week-old mice with cardiac deletion of BSCL2 driven by the same Myh6-Cre35. This discrepancy could be due to differences in construct design or background strain.
Comparisons of cardiac lipidomes between Bscl2−/− and Bscl2cKO mice clearly demarcate the autonomous and non-autonomous effects of BSCL2 deletion on lipid remodeling. Bscl2cKO mice exhibited an almost 50% reduction in total lipid contents concomitant with downregulation of a wide array of lipid species from glycerolipids to phospholipids to ACs (Figure 3). On the contrary, Bscl2−/− hearts only demonstrated reduced glycerolipid contents with a tendency toward lower levels of glycerophospholipids, NEFA and ACs 27. Such discrepancy could again be attributed to the excess circulating FA levels in hyperphagic Bscl2−/− mice. Notably, the levels of cardiac PA, the important glycerolipid intermediates and phospholipid precursors, were consistently reduced in both Bscl2−/− and Bscl2cKO hearts, suggesting a potential specific role of BSCL2 in regulating PA metabolism. Different from our results, PA levels were shown to be enriched in BSCL2-deleted murine adipose tissue36 and yeasts37, 38. Whether this can be ascribed to differences in tissue depots or species is not known. Additionally, BSCl2-deleted Drosophila S2 cells also displayed reduced phospholipids which potentially account for the formation of giant lipid droplets39. More work is needed to understand whether BSCL2 plays a direct role in mediating phospholipid metabolism.
PGC-1α and PPARα are key activators of TG dynamics and content in the heart40, 41. Especially, ATGL-mediated fat catabolism has been directly linked to cardiac PGC-1 and PPARα expression as well as FAO rates8. While Bscl2cKO hearts clearly exhibited excessive FAO in ex vivo perfused working hearts preceding impaired cardiac function, none of PGC-1α and PPARα and their downstream target genes were altered (Figure 2). Interestingly, lean mice with myocardial ATGL overexpression10, 11 or acetyl CoA carboxylase 2 (ACC2) deletion42 maintain normal cardiac energetics and performance despite higher TG turnover or FAO rates. Thus, it needs to be recognized that other changes in cellular metabolism independent of ATGL-mediated intramyocardial lipid catabolism may exist to contribute to the cardiac energy deficit and progressive deterioration of cardiac function in Bscl2cKO mice at baseline, which warrants further investigation. Regarding the posttranscriptional regulation of ATGL, ATGL is known to be ubiquitinated [43 and data not shown], and we have previously demonstrated enhanced ATGL stability in Bscl2−/− cardiomyocytes and mouse embryonic fibroblasts (MEFs)27. However, we were not able to pull down endogenous ATGL using current available antibodies, which prevents us from examining ATGL ubiquitination in BSCL2-deleted hearts. Therefore, the molecular events for the posttranscriptional regulation of ATGL in the absence of BSCL2 remain to be identified.
Reliance on FAO in obesity and/or diabetes is correlated with lower cardiac efficiency, impaired mitochondrial respiratory function and increased ROS production44. Alteration of the cardiac lipidome may also mediate functional impairment through dampening mitochondrial function45. However, none of these abnormalities occurred in our Bscl2cKO mice despite massive reduction of cardiac lipid contents. In fact, BSCL2-deleted mitochondria maintain efficient oxidative phosphorylation in the presence of exogenous substrates (Figure 4). This implies that the ATP deficit in Bscl2cKO hearts is mainly due to insufficient endogenous substrate in vivo independent of the intrinsic defects in mitochondrial function. Higher cardiac FAO is normally associated with increased exogenous lipid import as observed in diabetic hearts or hearts with cardiac-restricted overexpression of PPARα46. However, our Bscl2cKO mice displayed similar expression of lipid uptake genes and comparable circulating lipid metabolites, suggesting no defect in lipid transport. This may ultimately result in an imbalance of lipid consumption and supply within Bscl2cKO myocardium leading to downregulation of vast amounts of lipid substrates (Figure 3). In addition, AC levels tightly reflect the FAO rates, and AC profiling has been used to identify FAO dysregulation47. Previous reports suggest muscle AC levels correlate negatively with FAO in the postabsorptive state48. Notably, Bscl2cKO hearts demonstrated no alterations in CPT1 expression, suggesting intact carnitine shuttle (Figure 3K). The almost unanimous reduction of long-chain ACs in Bscl2cKO hearts highlights the presence of increased FAO which may eventually deplete mitochondrial ACs thus reducing substrates entering TCA cycles and causing energy deficit. Thus, our Bscl2cKO mice constitute as the first animal model that demonstrates excessive myocardial lipid catabolism associated with progressive deterioration of metabolic reserve and HF in the absence of elevated lipid uptake.
TMZ has been reported to improve cardiac function in experimental models of ischemia/reperfusion injury49–51 and ischemic HF patients52–54. It predominantly acts by shifting energy production from NEFA to glucose oxidation in the heart50. TMZ exerts cardioprotective role in our Bscl2cKO mice. Yet, the underlying mechanisms may not be simply explained by alleviation of energy deficit, as TMZ failed to significantly improve intracellular ATP levels in Bscl2cKO hearts. Although our focus was primarily on TMZ effect on cardiac function, we surprisingly uncovered that TMZ remodels cardiac lipidome by downregulating the lipid contents of almost all lipid classes in normal mouse hearts. We speculate inhibiting FAO may also put a brake on lipid transport and/or synthesis in the heart. Conversely, TMZ promotes drastic lipid and glycogen accumulation in metabolic-stressed Bscl2cKO hearts. The prominence of glycogen in TMZ-treated Bscl2cKO hearts may reflect enhanced glucose utilization as glycogen content has also been shown to commensurate with augmented carbohydrate metabolism such as in GLUT1-overexpressing hearts55. Notably, none of these changes can be associated with differences in the protein expression of lipid and glucose metabolic genes (data not shown). Interestingly, aside from glucose utilization, TMZ may also trigger enhanced cardiac ketolysis in Bscl2cKO hearts which demonstrated higher BDH1 expression. This finding concurs with a recent report on TMZ’s induction of cardiac β-hydroxybutyrate flux to attenuate isopropanol-induced rat heart failure56. Whether TMZ indeed induces substrate switch to both glucose and ketone bodies in Bscl2cKO hearts needs to be further dissected through ex vivo perfused working hearts. In addition, we cannot exclude the nonmetabolic effects of TMZ in preventing left ventricle remodeling independent of its inhibitory activity on FAO57, 58. Nevertheless, our study underscores that TMZ has potential in ameliorating cardiac function and slows HF progression in a non-ischemic model of HF.
HFD alone has been shown to be cardioprotective especially in alleviating energy-compromised HF59. In the present study, we find that decreased cardiac function in Bscl2cKO mice can be attenuated by HFD feeding. Lipidomics study further confirmed improved cardiac metabolic substrates mainly in the form of TG in HFD-fed Bscl2cKO mice. This was in line with the notion that HFD feeding provides more coronary circulation of substrates to match up the rate of enhanced lipid utilization thus attenuating the myocardial ATP deficit in the energy-deprived failing Bscl2cKO hearts. In spite of an improvement in cardiac contractility, we were not able to observe a significant reduction of cardiac stress markers in HFD-treated Bscl2cKO mice (not shown). Nonetheless, results from our HFD feeding studies support the cardio-protective effect of HFD on the energy-deprived HF.
In conclusion, our study highlights an important link between BSCL2 and myocardial energy metabolism and function and advances our understanding of the relationship between TG dynamics, FAO rates and the pathogenesis of HF. It may also provide insights into the therapeutic approaches in the treatment of cardiac disorders related to dysregulated metabolism in general.
Materials and Methods
Mice
Cre+; Bscl2f/f mice (designated as Bscl2cKO) mice were generated by breeding Bscl2f/f mice13 with transgenic mice expressing Cre recombinase under the cardiac-specific alpha myosin-heavy chain 6 (Myh6) promoter (JAX Cat#: 011038)60. The Myh6-Cre mice (designated as Cre+; Bscl2w/w) were included as controls when assessing cardiac functions. Cre-; Bscl2f/f mice (designated as Ctrl) were used as controls for the majority of the studies. To inhibit mitochondrial β-oxidation, 6-month-old Ctrl and Bscl2cKO mice were i.p. injected with PBS or trimetazidine (TMZ) at 15 mg/kg/day for 6 weeks, a dose that does not induce whole-body insulin resistance33. Ctrl and Bscl2cKO mice were fed a 60% HFD (Research Diets; D12492) starting at 3 months of age for a duration of 3 months. Most experiments were performed in ad libitum male mice and repeated in female mice. All mice were housed in the central animal facility with room temperature controlled at 21°C, and an artificial 12 h:12 h light: dark cycle (lights on at 06:00 am). Mice were directly sacrificed by cervical dislocation and hearts were rapidly excised. In ex vivo perfused working heart experiments, hearts were rapidly excised (<30 s) following anesthesia via intraperitoneal ketamine/xylazine (80/10 mg/kg) injection. All procedures involving animals and tissues conform to the NIH Guide for the Care and Use of Laboratory Animals and were approved by the Institutional Animal Care and Use Committee at the Augusta University.
Echocardiography
Echocardiographic analyses were performed on mice under anesthesia which was first induced with 5% isoflurane in 100% O2 for 1 min in a vaporizer then maintained during spontaneous breathing of 1.25% isoflurane in 100% O2 at a flow rate of 1 l/min via a small nose cone. Two-dimensional guided M-mode echoes were obtained from short-and long-axis views at the levels of the largest left ventricular diameter using a a VisualSonics Vevo 2100 echocardiography machine equipped with a 30 MHz probe (VisualSonics). All measurements and calculations were done in triplicates.
Tissue TG analyses
For tissue TG enzymatic analyses, lipids were extracted from tissue homogenates and dissolved in chloroform. The concentrations of TG were measured using a triglyceride assay kit (Infinity™ triglycerides kit, Thermo Fisher Scientific) and normalized to tissue weights as previously described13.
Histology and transmission electron microscopy (TEM)
Whole hearts were fixed, embedded and cut along the coronal plane to visualize the four-chamber view. The apex of left ventricle (LV) tissue was fixed and stained for electron microscopical imaging in a JEM 1230 transmission electron microscope (JEOL USA Inc., Peabody, MA) at 110 kV with an UltraScan 4000 CCD camera & First Light Digital Camera Controller (Gatan Inc., Pleasanton, CA) as previously described27.
Lipidomic analysis by high resolution/accurate mass spectrometry and tandem mass spectrometry
Total lipids from frozen ventricles were extracted and resuspended in isopropanol:methanol:chloroform (4:2:1 v:v:v) containing 20 mM ammonium formate followed by untargeted lipidomic analysis. Relative quantification of abundances between samples was performed by normalizing target lipid ion peak areas to the PC (14:0/14:0) internal standard followed by normalization to tissue weights as previously described27.
FAO assays
FAO reaction assays with LV homogenates were prepared and carried out as detailed previously 61. Briefly, ≈ 25 mg pieces of freshly isolated ventricular tissues were placed in STE buffer (250 mM Sucrose, 10 mM Tris, pH=7.5 and 1 mM EDTA) and homogenized using a glass dounce homogenizer (20 loose and 20 tight strokes). 20 μL tissue homogenates were incubated with 380 μL reaction buffer containing 1 μCi/mL [1–14C] palmitic acid substrate for fatty acid oxidation. The released [14C]CO2 was captured by hydroamine soaked filter paper and measured by scintillation counting, while acid soluble metabolites (ASM) were analyzed by centrifugation and counting of 14C radioactivity in the supernatant. Data were normalized to the total protein contents for LV homogenates.
Isolation and culture of adult cardiomyocytes
The isolation of adult mouse cardiomyocytes was carried out based on established procedures 62. The cardiomyocytes were suspended in plating media and plated onto laminin (5 μg/mL) precoated tissue culture plates. 1 h after plating, myocytes were changed to the culture media in the absence of 2,3-butanedione monoxime (BDM), ITS (Insulin/transferrin/selenium supplement) and lipid and kept in culture for 4 h, before exposure to isoproterenol (1 μM) for 20 mins.
Substrate metabolism in isolated working mouse hearts
Myocardial substrate utilization and contractile function were measured ex vivo in hearts isolated from 14 to 16 week-old male Ctrl and Bscl2cKO mice. All hearts were prepared and perfused in the working mode (non-recirculating manner) for 30 minutes with a preload of 12.5 mmHg and an afterload of 50 mmHg as previously described63, 64. Standard Krebs–Henseleit buffer was supplemented with 8 mM glucose, 0.4 mM oleate conjugated to 3%BSA (fraction V, FA-free; dialyzed), 10 μU/ml insulin (basal/fasting concentration), 0.05 mM L-carnitine, and 0.13 mM glycerol. Metabolic fluxes were assessed through the use of distinct radiolabeled tracers: 1) [U-14C]-glucose (0.12 mCi/L from MP Biomedicals; glucose oxidation); and 2) [9, 10-3H]-oleate (0.067 mCi/L from Sigma-Aldrich; β-oxidation). Measures of cardiac metabolism (e.g., oleate and glucose oxidation, and oxygen consumption) and function (e.g., cardiac power) were determined. At the end of the perfusion period, hearts were snap-frozen in liquid nitrogen and stored at −80°C prior to analysis. Data were presented as steady state values (i.e., the mean of the last two time points during a distinct perfusion condition for each individual heart).
Mitochondrial isolation and measurement of mitochondrial respiration
Fresh ventricles were isolated and minced for mitochondrial isolation and measurement of mitochondrial respiration by XF24 Extracellular Flux Analyzer (Seahorse Bioscience) as previously described32. See Supplemental Materials and Methods for details.
Tissue ATP measurement
ATP content was determined by using ATP Bioluminescent Assay Kit (FL-AA; Sigma-Aldrich, Saint Louis, MO, USA) according to the manufacturer’s procedure. Briefly, frozen heart tissues were homogenized in cold 10% trichloroacetic acid buffer, centrifuged at 5000 × g for 10 min at 4°C followed by neutralization with 50 mM Tris-acetate (pH 7.8). The ATP content was then determined by a multi-mode microplate reader with luminescence luminometer (FLUOstar Omega; BMG Labtech). Data were normalized to tissue weight.
RNA isolation and real-time quantitative PCR
Total RNA was extracted with Trizol Reagent (Thermo Fisher) and reverse-transcribed using MLV-V reverse transcriptase using random primers (Invitrogen). Real-time quantitative RT-PCR was performed on the Strategene MX3005 system. Data were normalized to 2 housekeeping genes (Actb and 36B4) based on Genorm algorithm (medgen.ugent.be/genorm/) and expressed as fold changes. All tissue gene expression studies were performed in nonfasted mice. RT-PCR Primers were listed in Supplemental Table S4.
Immunoblotting
Tissues were lysed in lysis buffer containing 25 mM Tris-HCl (pH 7.4), 150 mM NaCl, 2 mM EDTA, 1% Triton X-100 and 10% glycerol with freshly added protease and phosphatase inhibitor cocktail (Sigma). The protein concentration was determined by Bradford protein assay (Bio-Rad). Western blotting and quantification were performed as previously described27. Specific antibodies were listed in Supplemental materials.
Statistical analysis
Quantitative data were presented as means ± SEM. Animal experiments were performed with at least three independent cohorts. Statistical comparisons were made by using unpaired t test, one-way ANOVA followed by Dunnett’s multiple comparisons test, two-way ANOVA followed by Tukey’s post-hoc tests, or multiple t tests after correction using the Holm-Sidak method using the built-in statistics of GraphPad Prism 9 software. A P value of less than 0.05 was considered statistically significant.
Additional methodological details are included in the Supplemental Materials.
Competing Interests
None declared.
Author contributions
H. Zhou and W. Chen conceived the project and designed the research and wrote the manuscript. H. Zhou performed most physiological, biochemical and molecular studies; Ji Li and M.E. Young performed and supervised ex vivo perfused working hearts. Todd A. Lydic performed lipidomic analysis and assisted with data interpretation. Jie Li and H. Su performed and supervised adult cardiomyocyte isolation. H. Su, Ji Li and M.E. Young helped editing the manuscript.
Acknowledgement
We thank the Electron Microscopy and Histology Core at Augusta University for technical assistance and electron microscope imaging. This work was supported by National Heart, Lung and Blood Institute [1R01HL132182-01 to W.C.], National Institute of General Medical Sciences [R01GM124108 to J.L.], National Institute on Aging [R01AG049835 to J.L], and the American Heart Association Career Development Award (18CDA34080244 to H.Z.).
References




