

Abstract
Distinguishing between genetic, environmental, and genotype-by-environment effects is central to understanding geographic variation in phenotypic clines. Two of the best-documented phenotypic clines are Bergmann’s rule and Allen’s rule, which describe larger body sizes and shortened extremities in colder climates, respectively. Although numerous studies have found inter- and intraspecific evidence for both ecogeographic patterns, we still have little understanding about whether these patterns are driven by genetics, environment, or both. Here, we measured the genetic and environmental contributions to Bergmann’s rule and Allen’s rule across introduced populations of house mice (Mus musculus domesticus) in the Americas. First, we documented clines for body mass, tail length, and ear length in natural populations, and found that these conform to both Bergmann’s rule and Allen’s rule. We then raised descendants of wild-caught mice in the lab and showed that these differences persisted in a common environment, indicating that they have a genetic basis. Finally, using a full-sib design, we reared mice under warm and cold conditions. We found very little plasticity associated with body size, suggesting that Bergmann’s rule has been shaped by strong directional selection in house mice. However, extremities showed considerable plasticity, as both tails and ears grew shorter in cold environments. These results indicate that adaptive phenotypic plasticity as well as genetic changes underlie major patterns of clinal variation in house mice and likely facilitated their rapid expansion into new environments across the Americas.
Introduction
Clines in phentoypes have historically been attributed to natural selection and reflect adaptation to local environments (Huxley 1939; Endler 1977). Two of the most well described phenotypic clines are Allen’s rule and Bergmann’s rule. Allen’s rule is the observation that extremities, such as limb length and tail length, are shorter in colder climates compared to warmer regions, resulting in latitudinal clines (Allen 1877). Bergmann’s rule is the observation that body sizes are larger in colder climates, resulting in latitudinal clines in body size (Bergmann 1847). Shortened extremities and larger body sizes minimize heat loss by reducing surface area to volume ratios and are viewed as thermoregulatory adaptations (Mayr 1956). Numerous studies have documented Bergmann’s rule and Allen’s rule within and across species of birds (Johnston and Selander 1964; James 1970; Laiolo and Rolando 2001; Romano et al. 2020) and mammals (Brown and Lee 1969; Griffing 1974; Yom-Tov and Nix 1986; Fooden and Albrecht 1999), including humans (Ruff 1994, 2002; Foster and Collard 2013; Betti et al. 2015). Moreover, various meta-analyses have supported the generality of these rules (Ashton et al. 2000; Ashton 2002; Freckleton et al. 2003; Meiri and Dayan 2003; Blackburn and Hawkins 2004; Millien et al. 2006; Nudds and Oswald 2007; Olson et al. 2009; Symonds and Tattersall 2010). On the other hand, several meta-analyses have questioned the ubiquity of these patterns arguing that statistical support is weak (Geist 1987; Gohli and Voje 2016; Riemer et al. 2018) or that phenotypic differences are more likely to be driven by resource abundance rather than by considerations of temperature (McNab 1971; Geist 1987; Alhajeri and Steppan 2016). The contradicting results found across the literature are unsurprising given the variation within and among datasets, such as choice of taxonomic groups, environmental variables, and inconsistencies in measurements. Moreover, virtually all studies to date are based on observations of individuals sampled in natural populations in which factors such as age, reproductive condition, social status, pathogen and parasite loads, and overall health are not easily controlled. Thus, we still have very little understanding of the mechanisms underlying Allen’s rule and Bergmann’s rule.
Missing from many of these discussions are careful analyses determining which traits are genetically encoded, environmentally influenced, or both. Most traits associated with Bergmann’s rule and Allen’s rule are complex, meaning they are both polygenic and strongly influenced by the environment (Falconer and Mackay 1996; Lynch et al. 1998). Disentangling genetics from environmental effects in natural populations is difficult when using phenotypic data collected from wild-caught animals. Genetic contributions to trait values may be masked by environmental effects and genotype-by-environment interactions (Conover and Schultz 1995; Alho et al. 2011). Phenotypic plasticity may also generate clinal patterns, giving a false impression of adaptive clines (James 1983). In fact, many temporal changes in body size are driven by the environment and not genetic adaptation in birds (Teplitsky et al. 2008; Husby et al. 2011) and mammals (Ozgul et al. 2009, 2010). Furthermore, we have little understanding of how populations conforming to these ecogeographic rules vary in the degree and direction of plasticity they exhibit in response to environmental stimuli. Variation in plasticity and gentoype-by-environment interactions may faciliate adaptation and divergence in polygenic traits (Via and Lande 1985; Gillespie and Turelli 1989; Gomulkiewicz and Kirkpatrick 1992; West-Eberhard 2003). However, controlling for environmental effects and measuring the contributions of phenotypic plasticity is difficult, as transplant experiments and common garden experiments are infeasible for many taxa. These limitations have impeded our ability to make substantial progress on understanding the evolutionary and ecological mechanisms underlying Bergmann’s rule and Allen’s rule.
House mice (Mus musculus domesticus) provide a tractable system for disentangling the genetic and environmental contributions to complex traits. House mice have recently expanded their range from Western Europe to the Americas, where they can be found from the tip of South America to Alaska. Across this broad latitudinal range, mice are exposed to various environmental gradients, from cold, temperate environments to warm, tropical and arid environments (Phifer-Rixey and Nachman 2015). Despite residing in these novel environments for only ~500 generations (Phifer-Rixey et al. 2018), there is evidence for clinal variation across latitudes. Specifically, mice in eastern North America follow Bergmann’s rule (Lynch 1992), with larger mice in more northern populations. These body size differences persist in a commmon environment over several generations, indicating that they have a genetic basis (Lynch 1992; Phifer-Rixey et al. 2018). Selection on house mice over ten generations in the laboratory recapitulates these clinal patterns: mice bred at lower temperatures become larger and undergo genetic divergence in body size (Barnett and Dickson 1984). Furthermore, previous work has revealed an environmental influence on tail length when exposed to cold temperatures. Specficially, house mice reared in a cold environment grew significantly shorter tails than mice reared at warm temperatures, consistent with Allen’s rule (Sumner 1909, 1915; Barnett 1965). However, these earlier studies investigated only a single population of mice or used classical inbred laboratory strains of mice, making it difficult to place the results in an explicit evolutionary framework. We still have little understanding of the phenotypic variation of house mice across their entire latitudinal distribution, and even less understanding of the contributions of genetics and environment to these complex traits.
Here, we use a combination of approaches to tease apart genetics from plasticity in Bergmann’s rule and Allen’s rule in house mice from North and South America. First, we determined if house mice conform to both Bergmann’s rule and Allen’s rule across their entire introduced range by analyzing phenotypic data from wild-caught individuals. Second, we collected temperate and tropical populations of house mice from the ends of their latitudinal distribution, brought them back to the lab, and established wild-derived colonies. We analyzed phenotypic differences between populations and across generations in a common lab environment and identified a genetic basis to both Allen’s rule and Bergmann’s rule. Third, to measure the influence of environment on body size and extremity length, we performed a second common garden experiment by rearing both populations of mice using a full-sib design in a cold and warm environment and measured the effects on body size and extremity length. Measuring developmental plasticity within and between populations allowed us to assess the influence of temperature on complex traits and to understand the evolutionary mechanisms underlying these clinal patterns. Specifically, we found that unlike body size, tail and ear length are highly plastic and this plastic response goes in the same direction as the evolved response, highlighting an example of adaptive phenotypic plasticity.
Materials and Methods
Phenotypic data from wild-caught mice
To determine if house mice conform to Allen’s rule and Bergmann’s rule, we tested for associations between body mass, tail length, ear length, and latitude in wild house mice collected across North and South America. We dowloaded specimen data of all house mouse records from VertNet (Constable et al. 2010) on October 13, 2020, using the search query: vntype:specimen, genus:Mus. We obtained 62,139 museum records and retained records that included Mus musculus specimens collected in North or South America (excluding islands). We omitted individuals listed as pregnant, juvenile, subadult, or immature, and included individuals listed as adult, mature, or with no age class specified. We also manually coded females and males as ‘adult’ if they fulfilled any of the following criteria: females - presence of placental scars, parous, or lactating; males - presence of seminal vesicles, testes descended (TD), or testes scrotal (TS). Tail lengths shorter than 20mm and longer than 120mm (n = 8), and ear lengths greater than 30mm (n = 1) were considered extreme outliers (greater than 3.5 standard deviations from the mean) and were removed from downstream analyses. Sample information for the final VertNet dataset (n = 3,018) is provided in Data S1.
Laboratory-reared mice - common garden experiment 1
For the first common garden experiment, we collected live animals from two locations that represent the ends of a latitudinal transect: Manaus, Amazonas, Brazil (MAN), located near the equator at 3oS latitude, and Saratoga Springs, New York, USA (SAR), located at 43oN latitude. Details of this common garden experiment are given in (Phifer-Rixey et al. 2018) and (Suzuki et al. 2020). Briefly, live mice from both Brazil and New York were brought back to the lab at the University of California, Berkeley. Within each population, unrelated pairs of wild-caught mice were mated to produce first generation (N1) lab-reared mice, and these inbred lines have subsequently been maintained through sib-sib matings each generation for over 10 generations. Wild-caught mice and their descendants were housed in a standard laboratory environment at 21°C with a 12-hr dark and 12-hr light cycle. Water and commercial rodent chow (Teklad Global, 18% protein, 6% fat) were provided ad libitium. Standard museum measurements (total length, tail length, hind foot length, ear length, and body mass) were taken for all wild-caught, N1, and N2 mice from each population (see Data S2). Tail lengths less than 50mm (n = 2) and ear lengths less than 8mm (n = 1) were considered outliers (greater than three standard deviations away from the mean) and were removed from downstream analyses.
Developmental phenotypic plasticity - common garden experiment 2
For the second common garden experiment, we used two wild-derived inbred lines each from Brazil (MANA, MANB) and New York (SARA, SARB). Each line has been inbred for more than 10 generations, and thus mice within a line are nearly identical genetically. Equal numbers of males and females were produced for each within-line comparison (n = 80; see Data S3 and S4). Full-sibs were born at room temperature (21°C) and singly-housed at weaning (~21 days old). After a brief acclimation period, we randomaly assigned 3.5-week-old mice into size-matched groups based on sex-specific body mass, and then housed mice at either 5°C or remained at 21°C for the duration of the experiment (~50 days total). We measured initial body mass and tail length and recorded subsequent body mass and tail lengths once a week for each mouse. At the end of the experiment, we euthanized mice at 75 ± 3 days of age, and recorded final body mass and tail length, in addition to standard museum measurements. Two final ear lengths were not included in downstream analyses due to ear damage. We deposited skulls and skeletons of all mice in the Museum of Vertebrate Zoology, University of California, Berkeley (catalog numbers are given in Data S4). All experimental procedures were in accordance with the UC Berkeley Institutional Animal Care and Use Committee (AUP-2017-08-10248).
Data Analysis
All data analyses and visualizations were completed in R (v. 4.0.3). Within R, we used the tidyverse (v. 1.3.0) (Wickham et al. 2019), performance (v. 0.7.1) (Lüdecke et al. 2021), cowplot (v. 1.1.1), here (v. 1.0.1), and rmarkdown (v. 2.7) (Allaire et al. 2021) packages, along with R base library. Relative tail length and relative ear length were calculated by dividing tail or ear length by body mass for each individual. We also performed all analyses using tail length residuals and ear length residuals (by regressing length from body mass across individuals) and obtained similar results.
We tested for clinal patterns of body mass and extremity length across latitude in wild-caught house mice using Spearman correlations. For common garden experiment one, we fitted a linear model using lm{stats} to predict body mass, tail length, and ear length with sex, population, and generation (formula: (trait) ~ Sex * Population * Generation). The significance of interactions was evaluated using summary{base} and analysis of variance (ANOVA) based on type III (partial) sums of squares, implemented in the CAR library (v. 3.0.10) (Fox and Weisberg 2019). For common garden experiment two, we fitted a linear mixed model (estimated using restricted maximum likelihood) using lmer{lme4} (v. 1.1.26) (Bates et al. 2015) to predict body mass, tail length, and ear length with sex, population and environment (formula: (trait) ~ Sex * Population * Environment). The model included line as a random effect (formula: ~1 | Line). Results were evaluated using summary{lmerTest} (v. 3.1.3) (Kuznetsova et al. 2017) and Anova{car}. We peformed post hoc comparisons on significant two-way interactions using Tukey’s HSD tests or Mann-Whitney U tests. The code to perform analyses for this study are available as a git-based version control repository on GitHub (https://github.com/malballinger/Ballinger_allenbergmann_AmNat_2021). The analysis can be reproduced using a GNU Make-based workflow with built-in bash tools (v. 3.2.57(1)-release) and R (v. 4.0.3).
Results
Evidence for Bergmann’s rule and Allen’s rule in wild house mice
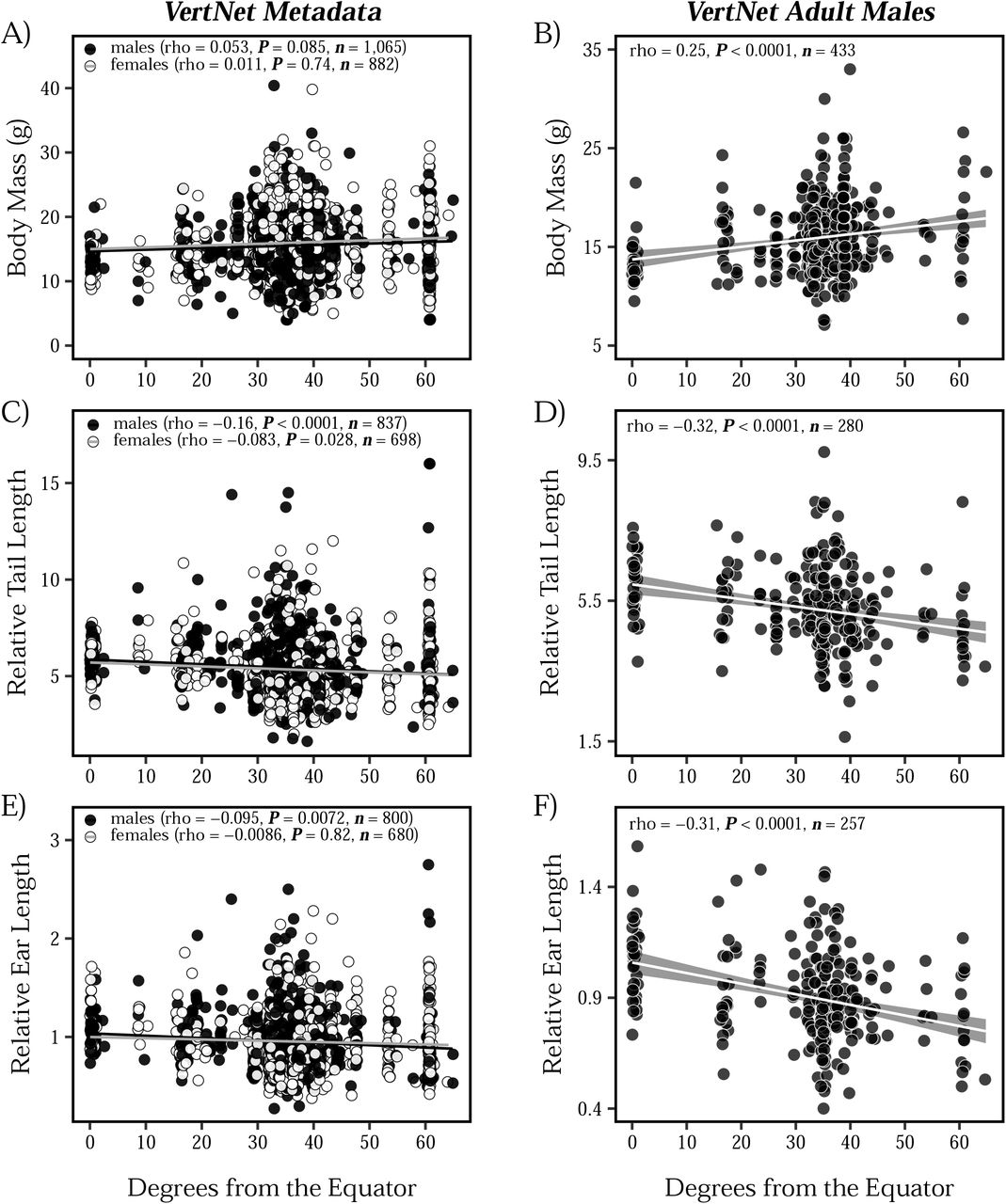
We assessed the relationship between tail length, ear length, body mass, and latitude in mice collected across North and South America to determine if populations of house mice conform to Allen’s rule and Bergmann’s rule. Using a large dataset downloaded from VertNet (n = 3,018; Data S1), we find little evidence for Bergmann’s rule, as body mass showed a non-significant, positive correlation with latitude across both males and females (Figure 1A). In contrast, we found stronger evidence for Allen’s rule in American house mice, with both tail length (Figure 1C) and ear length (Figure 1E) showing a significant, negative correlation with latitude. These patterns of extremity length largely hold true across both sexes (Figure 1C, 1E). In each of these comparisons, however, latitude explained only a very small amount of the phenotypic variation.
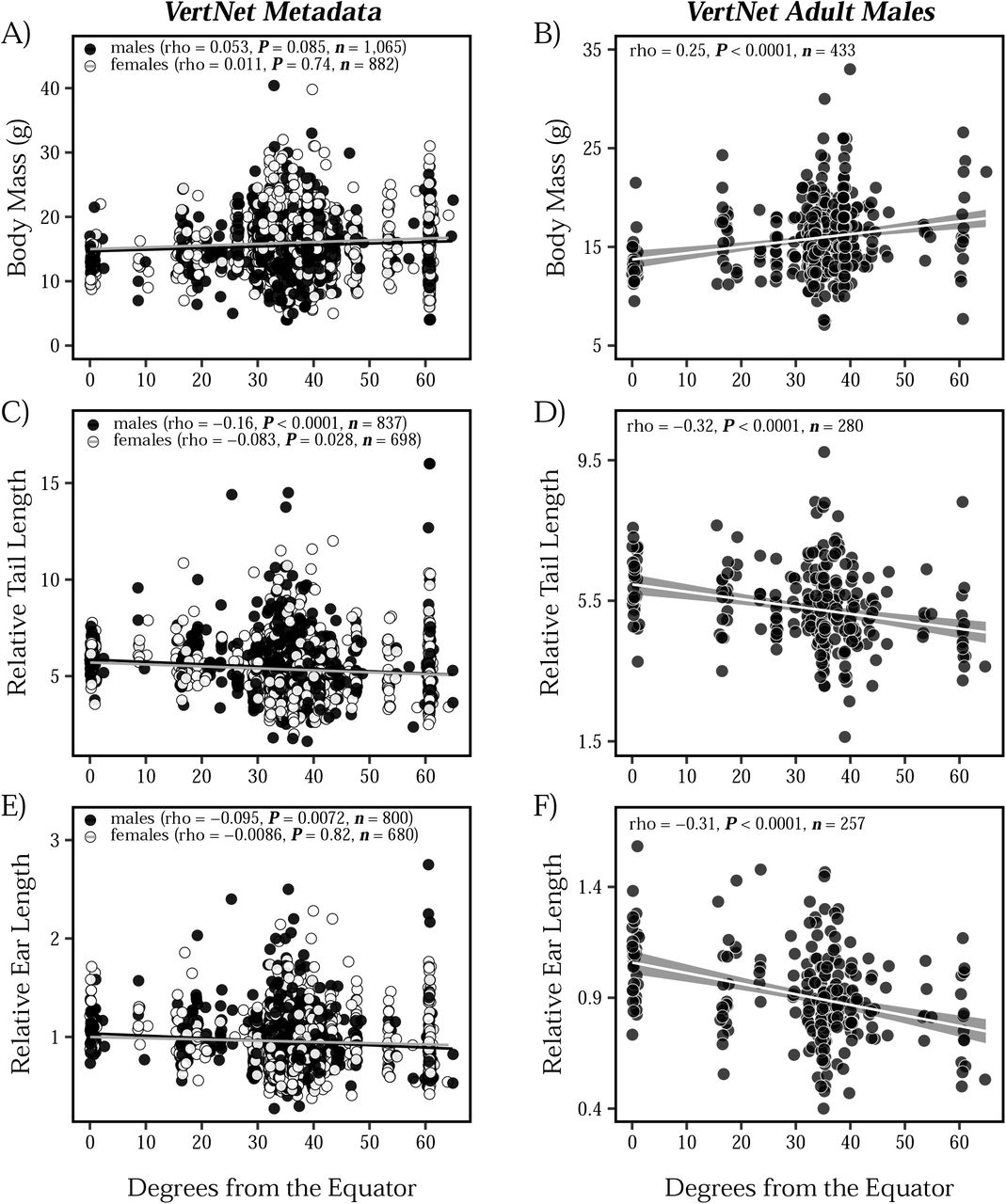
Associations between body mass (A-B), tail length (C-D), ear length (E-F), and absolute latitude across wild-caught North and South American house mice. Tail length and ear length are plotted relative to body mass for each individual. Individuals are represented as individual points, with males depicted in black and females depicted in white. Results from Spearman correlations are presented in each plot, along with sample sizes. For clarity, standard error shading is ommitted from linear regression lines associated with the VertNet Metadata (panels A, C, and E).
The lack of evidence for Bergmann’s rule in Figure 1A may be due to the influence of uncontrolled factors (e.g., age, diet, health), environmental effects, or phenotypic plasticity. Although we minimized the inclusion of records of non-adult specimens by removing pregnant females, juveniles, and subadults, we still see large variation across all three traits, likely due to various factors that were not recorded. To reduce this variation, we filtered the VertNet dataset to only include adult males (Figure 1B, 1D, 1F; n = 445). We focused on males since females show substantial variation associated with reproductive condition. In this more controlled set of adult males, we see strong evidence for both Bergmann’s rule (Figure 1B) and Allen’s rule (Figure 1D, 1F). The comparison between the larger dataset (Figures 1A, 1C, 1E) and the more curated dataset (Figure 1B, 1D, 1F) highlights how uncontrolled variation in collated museum metadata may obscure broad ecogeographic patterns.
Differences in body mass and extremity length persist in a common environment
The phenotypic clines observed across wild house mice could represent genetic differences, phenotypic plasticity, or both. To disentangle genetics from plasticity, we collected live mice from near the equator (Manaus, Amazonas, Brazil) and from 43oN latitude (Saratoga Springs, New York, USA) and brought them into a common laboratory environment. Population-specific differences in body mass in wild-caught mice (N0) persisted across the first two generations of laboratory-reared mice (N1 and N2; Figure 2A). Specifically, mice from New York were larger than mice from Brazil (Kruskal–Wallis, F1,439=282.54, P<0.001) (Figure 2A; Figure S1), and these differences persisted across generations (Mann-Whitney U, P<0.05). Sex-specific differences in body mass were also seen across generations, with males larger than females (Kruskal–Wallis, F1,439=50.79, P<0.001) (Figure 2A; Figure S1), and the direction and magnitude of sexual dimorphism was the same among New York and Brazil house mice. The maintenance of body mass differences in a common environment and across generations suggests a strong genetic basis in house mice.

Differences in body mass (A), tail length (B), and ear length (C) between New York mice (blue) and Brazil mice (gold) across generations. Tail length and ear length are plotted relative to body mass for each individual. Population-level data are depicted as boxplots overlayed on density plots, with boxplot vertical lines denoting 1.5x the inerquartile range. Individuals are represented as individual points. Results from linear models are presented in the upper right corner of each panel (*P<0.05; Figure S1). Sample sizes: (A) n = 441; (B) n = 432; (C) n = 434.
Similarly, population-specific differences in extremity length (i.e., relative tail length and relative ear length) in wild-caught mice (N0) persisted across the first two generations of laboratory-reared mice (N1 and N2; Figure 2B-C). Specifically, mice from New York had shorter tails (ANOVA, F1,420=321.40, P<0.001) (Figure 2B; Figure S1) and shorter ears (ANOVA, F1,422=193.08, P<0.001) (Figure 2C; Figure S1) compared to mice from Brazil, and these differences persisted across generations (Tukey’s HSD, P<0.05). The maintenance of tail length and ear length differences in a common environment and across generations suggests a strong genetic basis in house mice.
Extremity length, but not body size, is greatly influenced by temperature
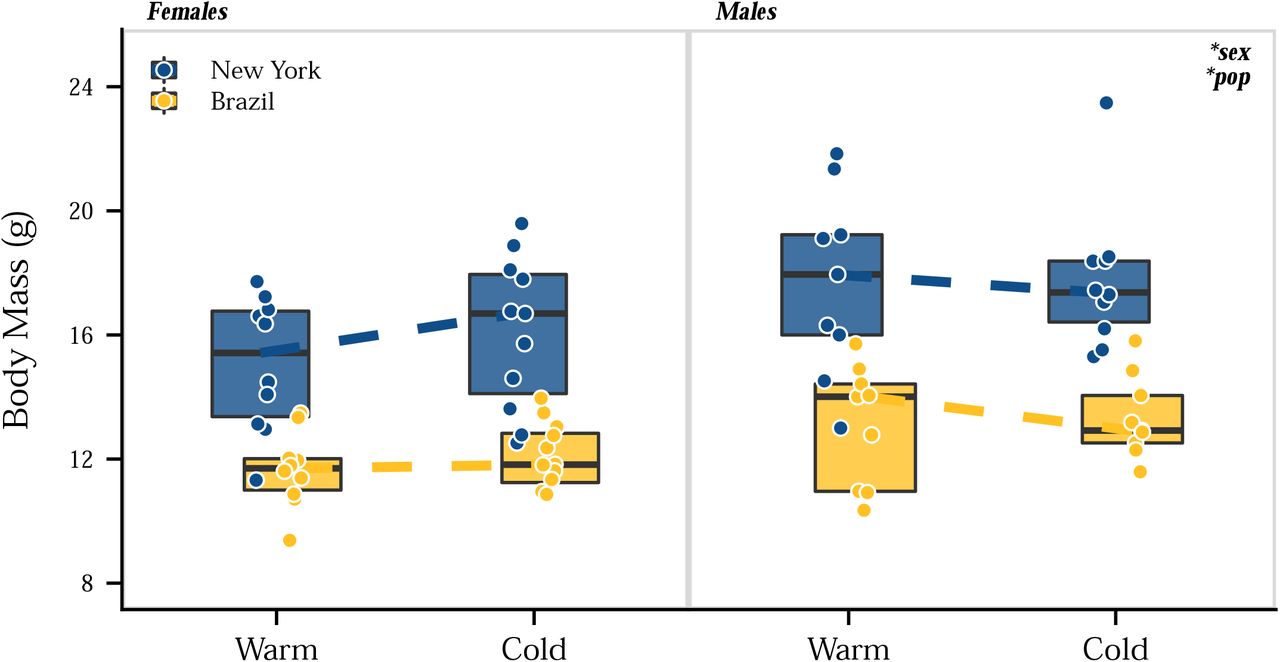
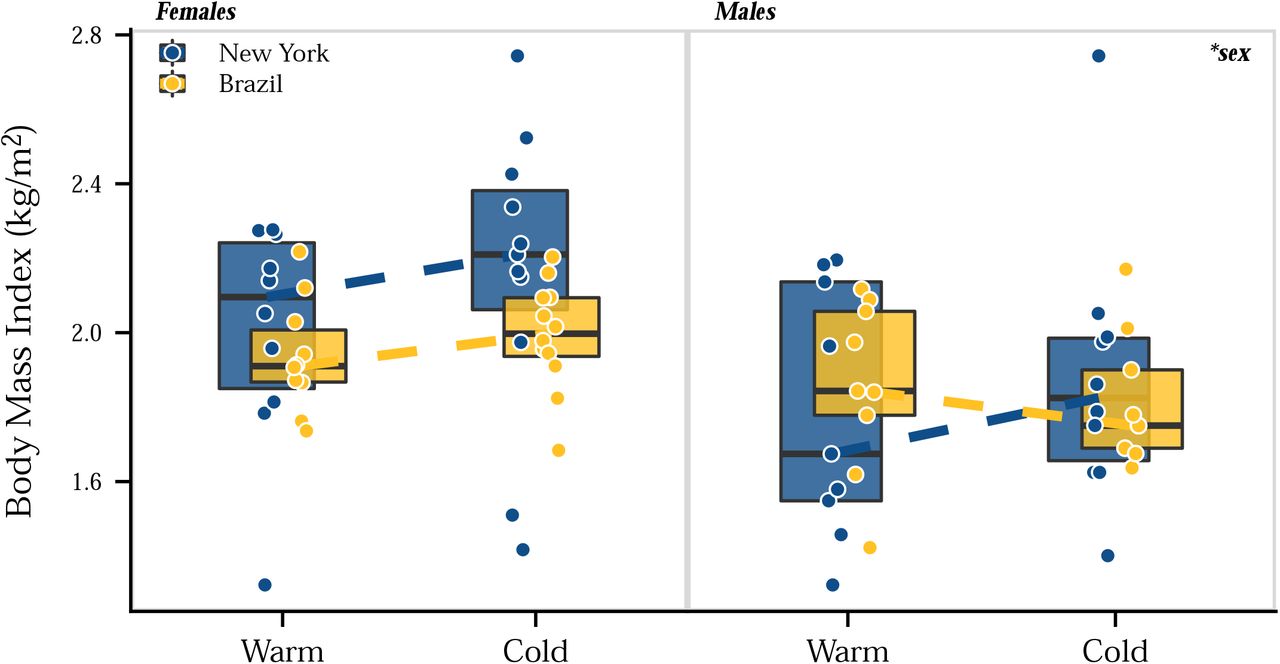
The results presented above identified phenotypic divergence in body mass and tail and ear length in house mice, with New York mice having shorter tails and ears and larger body sizes than mice from Brazil, consistent with Allen’s rule and Bergmann’s rule, respectively. To determine the influence of phenotypic plasticity on these traits, we performed a second common garden experiment by rearing laboratory-born mice from both populations in a cold and warm environment. We used temperature as the environmental variable because temperature is highly correlated with latitude (Millien et al. 2006) and phenotypic variation in wild house mice across North and South America is explained most by temperature-related variables (Suzuki et al. 2020). Evolved differences in body mass were evident at weaning (Figure 3), with New York mice larger than Brazil mice. These body mass differences between populations persisted across developmental stages from 3 to 11 weeks. In all cases, males were larger than females. Full-sibs (of the same sex) reared at different temperatures showed no differences in body mass (Figure 3). At the end of the experiment, body size differences recapitulated patterns seen across generations, with New York mice larger than Brazil mice (ANOVA, χ2=4, P<0.05) and males larger than females (ANOVA, χ2=42.15, P<0.001) (Figure 4, Figure S2). The lack of plasticity in body mass was not a result of differences in fat accumulation, as body mass index (BMI) did not differ between populations (ANOVA, χ2=0.50, P>0.05) or environments (ANOVA, χ2=1.28, P>0.05) (Figure S2; Figure S3). These results suggest that phenotypic plasticity does not play a significant role in body size evolution of house mice.

Body mass growth trajectories across environments in New York (blue) and Brazil (gold) house mice. Cold-reared mice are denoted as dotted lines and warm-reared mice are denoted as solid lines. Individuals are plotted as semi-transparent points (n = 80), with population means depicted as smoothed regression fits, with standard error shading. The same individuals depicted here are also depicted in Figure 4.
Individuals are represented as individual points (n = 80), and boxplots indicate the 25th, median, and 75th quartiles. Results from linear mixed models are presented in the upper right corner (*P<0.05; Figure S2). The same individuals depicted here are also depicted in Figure 3.
In contrast to body mass, tail length was greatly influenced by developmental temperature, with the first few weeks post-weaning having the greatest influence on absolute tail length (Figure 5). Specifically, mice reared in a cold environment grew shorter tails than mice reared in a warm environment (ANOVA, χ2=25.59, P<0.001) (Figure 5; Figure 6A; Figure S2). The magnitude of this effect was striking in Brazil mice, corresponding to 5.40 mm, on average, or 7% of the total tail length. Despite developmental temperature playing a signficant role in tail length, evolved differences in tail length were evident at the end of the experiment, with Brazil mice growing longer tails than New York mice in both environments (ANOVA, χ2=6.54, P<0.05) (Figure 5; Figure 6A; Figure S2). Thus, tail length exhibits both evolved divergence and phenotypic plasticity. Similarly, cold-reared mice grew shorter ears than warm-reared mice (ANOVA, χ2=20.07, P<0.05) (Figure 6B; Figure S2), and the magnitude of the plastic response corresponded to 5% of the total ear length in both populations. These temperature-growth responses of the extremities were not a simple consequence of body size differences, as body mass did not differ between treatments (Figure 4). Overall, unlike body size, extremity length showed significant plasticity in response to temperature, with mice growing shorter extremities in a cold environment, consistent with patterns of Allen’s rule.

Absolute tail length growth trajectories across environments in New York (blue) and Brazil (gold) house mice. Cold-reared mice are denoted as dotted lines and warm-reared mice are denoted as solid lines. Individuals are plotted as semi-transparent points (n = 80), with population means depicted as smoothed regression fits, with standard error shading. The same individuals depicted here are also depicted in Figure 6.

Tail length (A) and ear length (B) are plotted relative to body mass for each individual. Individuals are represented as individual points (tail length: n = 80; ear length: n = 78), and boxplots indicate the 25th, median, and 75th quartiles. Both sexes were combined for simplicity. Results from linear mixed models are presented in the upper right corner (*P<0.05; Figure S2). The same individuals depicted here are also depicted in Figure 5.
Adaptive phenotypic plasticity in extremity length
Differences between warm- and cold-reared mice revealed a strong plastic response to temperature in extremity length. Because plasticity is considered adaptive when a phenotype is altered in the same direction as natural selection (Ghalambor et al. 2007), we next asked whether phenotypic plasticity of Brazil mice goes in the same or opposite direction as the evolved response of New York mice. For absolute tail length, the plastic response in Brazil house mice nearly recapitulates the evolved tail length of New York mice (Figure 5, Figure 6A), highlighting an example of adaptive phenotypic plasticity. Interestingly, the plastic response of tail length for New York mice raised in the cold was attenuated in comparison to the plastic response of tail length for Brazil mice (Figure 5, Figure 6A), suggesting that New York house mice may be closer to the phenotypic optimum or that there is a developmental constraint on tail length. Lastly, plasticity in ear length of Brazil house mice went in the same direction as the evolved response of New York house mice (Figure 6B), further illustrating adaptive phenotypic plasticity. The overall degree and direction of plasticity in extremities mirror patterns associated with Allen’s rule. In fact, the plastic response of both tail length and ear length in Brazil mice explains roughly 40% of the mean phenotypic differences we observe in wild mice (i.e., between N0 Brazil mice and N0 New York mice).
Discussion
Previous studies have provided conflicting assessments of the generality of Bergmann’s and Allen’s rules and have rarely combined field and laboratory studies to identify the contribution of genetic and non-genetic effects to phenotypic variation. Here, we focused on one species that has recently expanded its range across many degrees of latitude, and we studied phenotypic variation in the wild and in the lab. Moreover, by rearing genetically identical mice at different temperatures we were able to assess the contribution of phenotypic plasticity to patterns seen in nature.
First, we found that wild house mice across North and South America conform to Bergmann’s rule and Allen’s rule, as house mice are larger in size with shortened extremities farther from the equator. Second, persistent differences in body mass and tail and ear length in a common environment across more than 10 generations indicated a genetic basis to Bergmann’s rule and Allen’s rule, presumably reflecting thermoregulatory adaptations. Finally, we measured the contributions of phenotypic plasticity to these traits and found that tail and ear length are highly plastic in response to cold temperature, while body size is not. The plastic response in extremity length to cold temperatures appears adaptive, matching the direction and magnitude of the evolved response in temperate house mice. Adaptive plasticity associated with Allen’s rule, in conjunction with strong selection for body size, likely promoted the rapid expansion of house mice into new environments across the Americas.
Genetic contributions to ecogeographic rules
Parallel phenotypic clines across multiple transects provide strong evidence for natural selection (Endler 1977). Body mass in house mice increases as latitude increases (Figure 1A-B), resembling patterns seen in eastern North America (Lynch 1992; Phifer-Rixey et al. 2018), South America (Suzuki et al. 2020), and Australia (Tomlinson and Withers 2009). These clinal patterns are consistent with Bergmann’s rule. Notably, this pattern is much clearer when looking at a dataset that includes only adult males (Figure 1B) compared to a dataset with all animals (Figure 1A). This difference may help to explain the discrepancies between previous meta-analyses based on museum collections (e.g., Ashton et al. 2000; Meiri and Dayan 2003; Riemer et al. 2018). Moreover, the substantial trait variation in Figure 1 among individuals, even when sampled at the same latitude, provides strong motivation for studying these traits in a common laboratory environment.
Phenotypic measurements of lab-reared mice from temperate and tropical populations revealed a genetically determined difference in body mass, with mice from colder climates significantly larger than mice from tropical environments (Figure 2A). This pattern was seen in the first two generations (Figure 2A) as well as in mice that had been in the lab for over 10 generations (Figure 3, Figure 4). These results agree with previous studies that also found a genetic basis for body size differences in house mice from eastern North America (Lynch 1992; Phifer-Rixey et al. 2018), western North America (Ferris et al. 2021), and South America (Suzuki et al. 2020), and suggest that there has been strong directional selection for body size in house mice. Selection over short time scales leading to latitudinal clines for body size has also been shown for other non-native species, such as in the genus Drosophila. Specifically, body size clines in introduced species of Drosophila have been repeated across continents, in common garden experiments, and through experimental evolution studies (Cavicchi et al. 1985; Coyne and Beecham 1987; Partridge et al. 1994; James et al. 1995; Land et al. 1999; Huey et al. 2000; Gilchrist et al. 2001, 2004). Latitudinal clines for body size have also arisen rapidly in introduced populations of house sparrows (Johnston and Selander 1964, 1971) and starlings (Cardilini et al. 2016). Together, these results suggest that introduced species may undergo strong selection while colonizing new environments, allowing patterns conforming to Bergmann’s rule to become quickly established.
Phenotypic measurements of wild mice revealed clines for relative tail length and relative ear length, with shorter extremities seen in mice collected farther from the equator, consistent with Allen’s rule (Figure 1). As with body size, a clearer picture of clinal variation emerged when only considering adult males (Figure 1D and Figure 1F) compared to all animals (Figure 1C and Figure 1E). Measurements of lab mice showed that these differences persist and thus have a genetic basis (Figure 2, Figure 5, Figure 6). Shorter tails in northern populations of house mice likely confer selection for heat conservation and adaptation to the cold, as tail length shows a positive correlation with tempeature of the coldest month across rodents (Alhajeri et al. 2020). Similar trends and correlations have also been found for limb length and bill length in birds (Nudds and Oswald 2007; Symonds and Tattersall 2010; Danner and Greenberg 2015; Friedman et al. 2017). In addition to thermoregulatory advantages, alternative mechanisms for Allen’s rule have been postulated to explain why longer tails are found in the tropics, such as enhanced climbing ability with increased arboreality (Alroy 2019; Mincer and Russo 2020). Because house mice are commensal with humans, it is unlikely that longer tails confer a climbing advantage in the tropics.
Contributions of phenotypic plasticity to ecogeographic rules
Body size in house mice shows very little plasticity in response to cold temperature (Figures 3 and 5), reaffirming that there has likely been strong directional selection for body size in house mice. Lack of plasticity associated with Bergmann’s rule is consistent with previous studies in laboratory mice (Sumner 1909, 1915; Ashoub 1958; Serrat et al. 2008; Serrat 2013) and, in addition to selection, may be due to a number of physiological factors. First, the environmental influence of temperature may need to occur preweaning or prenatal to elicit a plastic response (e.g., Weaver and Ingram 1969; Burness et al. 2013; Andrew et al. 2017). Second, traits that provide flexible and immediate heat conservation, such as increased fur insulation (e.g., Sumner 1915; Weaver and Ingram 1969), may provide an initial phenotypic response to cold temperature and thereby mitigate selection on body size. Lastly, exposure to high temperatures instead of low temperatures may elicit a plastic response in body size, as seen previously in some endotherms (Ashoub 1958; Gordon 2012; Burness et al. 2013; Andrew et al. 2017).
Unlike Bergmann’s rule, Allen’s rule can be generated via developmental phenotypic plasticity, as extremity length is highly senstive to ambient temperature in both mammals and birds (Serrat 2014; Tattersall et al. 2017). Our results in wild house mice agree with previous studies in mammals, with mice growing shorter tails and ears in a cold environment (Figure 6) (Ogle and Mills 1933; Harland 1960; Chevillard et al. 1963; Weaver and Ingram 1969). In laboratory mice, temperature directly affects the growth of cartilage in both tails and ears, influencing extremity length (Serrat et al. 2008). Furthermore, the widespread patterns of tail length plasticity in response to cold are also recapitulated at the skeletal level, with both the length and number of caudal vertebrae decreasing in response to cold temperatures in mice (Barnett 1965; Noel and Wright 1970; Thorington Jr 1970; Al-Hilli and Wright 1983). Although we did not measure skeletal differences between New York and Brazil mice, it seems likely that the tail length plasticity we observed is a result of plasticity in both number and length of individual caudal vertebrae. Moreover, ear length shows the greatest plasticity in both populations, with both New York and Brazil mice growing shorter ears in the cold. The pronounced plastic response of ears compared to tails may indicate that smaller appendages consisting entirely of cartilage are less developmentally canalized. Less constraint associated with extremities may also underly the highly plastic nature of Allen’s rule compared to Bergmann’s rule. This is illustrated by tail length and ear length plasticity accounting for roughly 40% of the observed differences among wild New York and Brazil house mice.
Adaptive phenotypic plasticity and Allen’s rule
Phenotypic plasticity is adaptive when it aligns with the direction of selection, moving traits closer to the local phenotypic optima (Baldwin 1896; West-Eberhard 2003; Ghalambor et al. 2007). We found evidence for adaptive phenotypic plasticity underlying Allen’s rule, as plasticity produced shorter ears and tails in cold environments. We also observed an attenuated plastic response for tail length in New York house mice compared to Brazil house mice, suggesting that New York mice are closer to the phenotypic optimum and are better adapted to colder environments. Overall, plasticity in house mouse extremities mirrors general evolutionary patterns of shorter extremity lengths in colder climates and may play an important role in generating Allen’s rule.
There are two ways by which adaptive phenotypic plasticity can facilitate the colonization of new environments. Adaptive plasticity can incompletely move the trait value closer to the phenotypic optimum, with directional selection refining the trait value, leading to subsequent genetic changes (Price et al. 2003; Ghalambor et al. 2007). Alternatively, adaptive plasticity can slow or impede evolution by moving individuals completely to the phenotypic optimum, shielding genetic variation from natural selection (Price et al. 2003; Ghalambor et al. 2007). We find evidence for the first scenario for both tail and ear length in house mice. Specifically, both genetic and plastic contributions generate shorter tails and ears in colder environments. Despite the plastic response of extremity length in Brazil mice nearly recapitulating the magnitude of the evolved response of New York mice, we see clear evidence of genetic differences in tail length and ear length between New York and Brazil house mice. This suggests that phenotypic plasticity moves extremity length close to the local optimum but does not shield it from subsequent selection. Overall, adaptive phenotypic plasticity with Allen’s rule, in addition to strong, directional selection underlying Bergmann’s rule, likely facilitated the rapid expansion of house mice into new environments across the Americas.
Supplemental Figures

Points and ranges represent model estimates and 95% credibility estimates for the linear model, with color indicating the phenotypic trait (purple: body mass; black: tail length; green: ear length). Solid lines that do not cross the dotted, vertical line are significant.

Points and ranges represent model estimates and 95% credibility estimates for the linear mixed model, with color indicating the phenotypic trait (purple: body mass; black: tail length; green: ear length; gray: BMI). Solid lines that do not cross the dotted, vertical line are significant.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Individuals are represented as individual points (n = 80). The same individuals depicted here are also depicted in Figure 4.
Acknowledgements
We thank Michael Sheehan and Felipe Martins for collecting wild mice, and we thank Kathleen Ferris, Gabriela Heyer, Dana Lin, Felipe Martins, Megan Phifer-Rixey, Michael Sheehan, and Taichi Suzki for help with mouse husbandry. M.A.B was supported by a National Science Foundation Graduate Research Fellowship (DGE 1106400), Junea W. Kelly Museum of Vertebrate Zoology Graduate Fellowship, and a UC Berkeley Philomathia Graduate Fellowship. This work was supported by graduate student research funds from the Museum of Vertebrate Zoology and Department of Integrative Biology to M.A.B and an NIH grant to M.W.N. (R01GM127468).
References




