

Abstract
Control of voluntary limb movement is predominantly attributed to the contralateral motor cortex. Nevertheless, increasing evidence suggests the involvement of ipsilateral cortical networks in this process. Ipsilateral control particularly emerges in motor tasks requiring bilateral coordination, which is an essential characteristic of locomotion. Here, we combined a unilateral thoracic spinal cord injury (SCI) with a cortical neuroprosthetic intervention that uncovered a functional role of the ipsilateral cortex in rat movement. In all rats, after SCI, ipsilesional cortex excitation promoted a bilateral synergy, whereby the elevation of the contralateral foot was complemented by ipsilateral hindlimb extension. In two animals, we found that stimulation of a medial cortical sub-region modulated ipsilateral hindlimb flexion. Ipsilateral cortical stimulation delivered after SCI alleviated multiple locomotor and postural deficits. These results establish a causal link between cortical activation and a remarkably fine and proportional ipsilateral control of hindlimb movement, a potential target for future neuroprosthetic technology.
Introduction
Cortical commands primarily regulate contralateral limb movements. This lateralization bias is reflected (1) anatomically, by a majority of crossed cortico-spinal tract (CST) projections (Hicks & D’Amato, 1975), (2) electrophysiologically, by a predominance of contralateral muscle recruitments by cortical stimulation (Kwan et al., 1978), (3) functionally, by contralateral deficits induced by cortical lesions (Passingham et al., 1983). However, lateralization of cortical control is incomplete, yet there is limited evidence on the functional significance of cortical ipsilateral regulation of movement (Montgomery et al., 2013). A minority of direct cortico-spinal projections are uncrossed (Vahlsing & Feringa, 1980). Ipsilateral impairments have been reported after unilateral cortical injury or transient interference (e.g., via transcranial magnetic stimulation) accompanied with increased cortical activity from the opposite hemisphere (Blasi et al., 2002; Chen, Gerloff, et al., 1997; Johansen-Berg et al., 2002; Jones et al., 1989; Kim et al., 2003; Marque et al., 1997; Yarosh et al., 2004). Nevertheless, the function of the ipsilateral motor cortex is unclear and its role in the recovery of motor control after injury remains controversial (Caramia et al., 2000; Chen, Cohen, et al., 1997; Dancause et al., 2006; Hallett, 2001; Hummel & Cohen, 2006; Jankowska & Edgley, 2006; Serrien et al., 2004; Stoeckel & Binkofski, 2010; Turton et al., 1996). It has been reported that the ipsilateral motor cortex regulates tasks involving bilateral movements (Ames & Churchland, 2019; Donchin et al., 1998). Imaging studies have shown that lower extremities movements and walking, which require efficient bilateral coordination, are associated with bilateral activity in primary sensorimotor cortices and supplementary motor areas (Miyai et al., 2001). Yet cortical dynamics underlying locomotion have been primarily studied in relation to contralateral kinematics (Bonizzato et al., 2018; Brown & Martinez, 2021; DiGiovanna et al., 2016; Yin et al., 2014). The relationship between cortical commands and locomotion has received attention in the last decades (Amboni et al., 2013). Recent studies have shown that not only the cortex proactively controls high-level and goal-oriented motor planning but it is also involved during stereotyped locomotion (Artoni et al., 2017). Nevertheless, demonstrations of functional hindlimb controllability by cortical networks are still lacking, especially with respect to ipsilateral cortical contribution.
In order to address this knowledge gap, we designed a behavioral neuromodulation framework to assess the gait-phase-specific effects of intracortical neurostimulation on ipsilateral hindlimb kinematics during locomotion. We evaluated the regulation of hindlimb trajectory and posture both in intact rats and after a unilateral hemisection SCI. This side-specific lesion preserves the majority of crossed projections from the ipsilateral cortex while maximizing the loss of direct efferences from the contralateral cortex. As early as one week after injury, different modalities of ipsilateral cortical neuroprosthetic stimulation immediately alleviated SCI-induced deficits, including lack of hindlimb support, weak hindlimb extension and flexion, and dragging.
Finally, by longitudinally acquiring chronic motor maps in awake rats, we sought to provide a parallel description of the time course of ipsilateral cortico-spinal transmission and spontaneous recovery of locomotor function after SCI.
Our functional causal approach to ipsilateral movement directly challenges the classical view whereby ipsilateral motor cortex control of movement is epiphenomenal and functionally limited. Our work shows that the cortex has direct functional control of hindlimb motor synergies and that its action can reverse SCI locomotor deficits.
Results
We designed cortical neurostimulation that unveiled the contribution of the motor cortex in controlling ipsilateral hindlimb movements. Our goal was to characterize whether the stimulation manipulated the ongoing locomotor output in intact conditions and alleviated motor deficits after hemiparesis induced by lateralized SCI. Phase-coherent intracortical stimulation was synchronized with gait phases such as the contralateral foot lift. The expected timing of the gait event was predicted by real-time processing of muscle activity (Bonizzato & Martinez, 2021). We induced selective unilateral hindlimb deficits with a thoracic lateral hemisection of the spinal cord (Brown & Martinez, 2019b). In this SCI model, the hindlimb on the side of the spinal lesion is transitory paralyzed due to the loss of main supraspinal inputs, but the ipsilesional cortex retains most of its crossed connections to the sublesional spinal circuits.
For the sake of clarity, throughout the manuscript, the terms “ipsilateral” and “ipsilesional” will only be used to indicate the left implanted motor cortex and left leg, which are located on the same side of the spinal hemisection. Conversely “contralateral” and “contralesional” will solely refer to the right cortex and right leg. In brief, left = ipsi-; right = contra-.
Phase-coherent stimulation in intact rats
Online detection of muscle activation was used to predict gait events and consequently trigger stimulation through a 32-channel intracortical array implanted into the left motor cortex. Stimulation effects on locomotor behavior were evaluated in n=6 intact rats (Fig. 1A). Gait modulation was timing-dependent: maximal effects were obtained for stimuli delivered during the preparation and early execution of the right swing and consisted of an increase in right hindlimb flexion (Fig. 1B). When the stimulation was delivered between the left contact and the right mid-swing (in ‘phase-coherence’ with foot lift (Bonizzato & Martinez, 2021)), right step height was increased (+129±17%, p=8E-5) and gait pattern modifications resulted in a right swing dominance (+25±2%, p=5E-4) and a left stance dominance (+9±1%, p=6E-4, Fig. 1B).
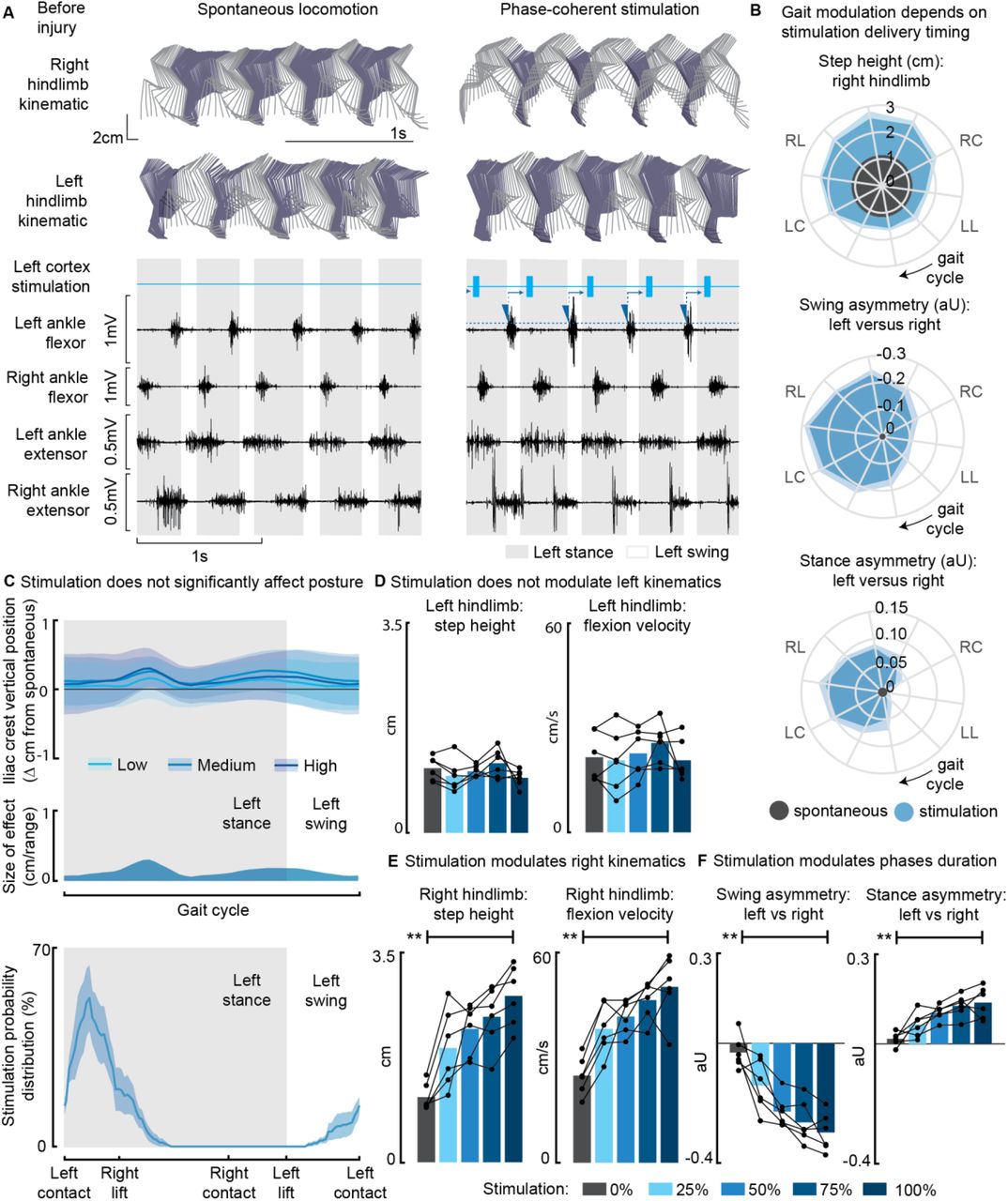
(A) Stick diagrams and electromyographic (EMG) activity during spontaneous locomotion and phase-coherent stimulation. The stimulation was triggered by left ankle flexor activation and was delivered during the late right stance (early left stance). (B) Polar plots showing contralateral step height in cm and gait phase asymmetries in arbitrary units (aU) for stimulation delivered with different timings along the whole gait cycle. Positive asymmetry index values refer to left-side dominance. For ease of reading, the radial axis of the swing symmetry plot has been inverted (outer values are negative). For the three polar plots, the most effective kinematic neuromodulation corresponds to the largest radius. The gait cycle progresses clockwise, and the gait event references are reported as LC: left contact, RL: right lift, RC: right contact, and LL: left lift. (C) Analysis of the effects of cortical stimulation on the posture of rats (top) and experimental stimulation distribution (bottom). Posture is shown as the height of the left iliac crest during the gait cycle, which was not modulated by increasing cortical stimulation amplitude. (D) Characterization of ipsilateral kinematics. Left step height and flexion speed were not affected by increasing cortical stimulation amplitudes. (E) Modulation of contralateral kinematics. Right step height and flexion speed were linearly increased with greater stimulation amplitudes. (F) Modulation of bilateral gait phase duration. The absolute values of swing and stance asymmetry indexes were linearly increased with greater stimulation amplitudes. Positive asymmetry index values refer to left-side dominance. The data are represented as the mean ± SEM. ** p < 0.01.
While characterizing the effects of modulating phase-coherent stimulation amplitudes, we found no effects on posture (Fig. 1C) nor ipsilateral kinematics (Fig. 1D) for all of the intact rats. Nevertheless, stimulation modulated the walking duty cycle and contralateral swing kinematics. Right step height (+157±13%, p=2E-5), flexion velocity (+107±21%, p=2E-4), swing (30±3%, p=2E-5) and stance (14±2%, p=4E-4) asymmetry indexes increased linearly when increasing stimulation amplitudes (R2=[79±5, 78±6, 80±4, 73±9]%, Fig. 1E-F).
Phase-coherent stimulation of SCI rats
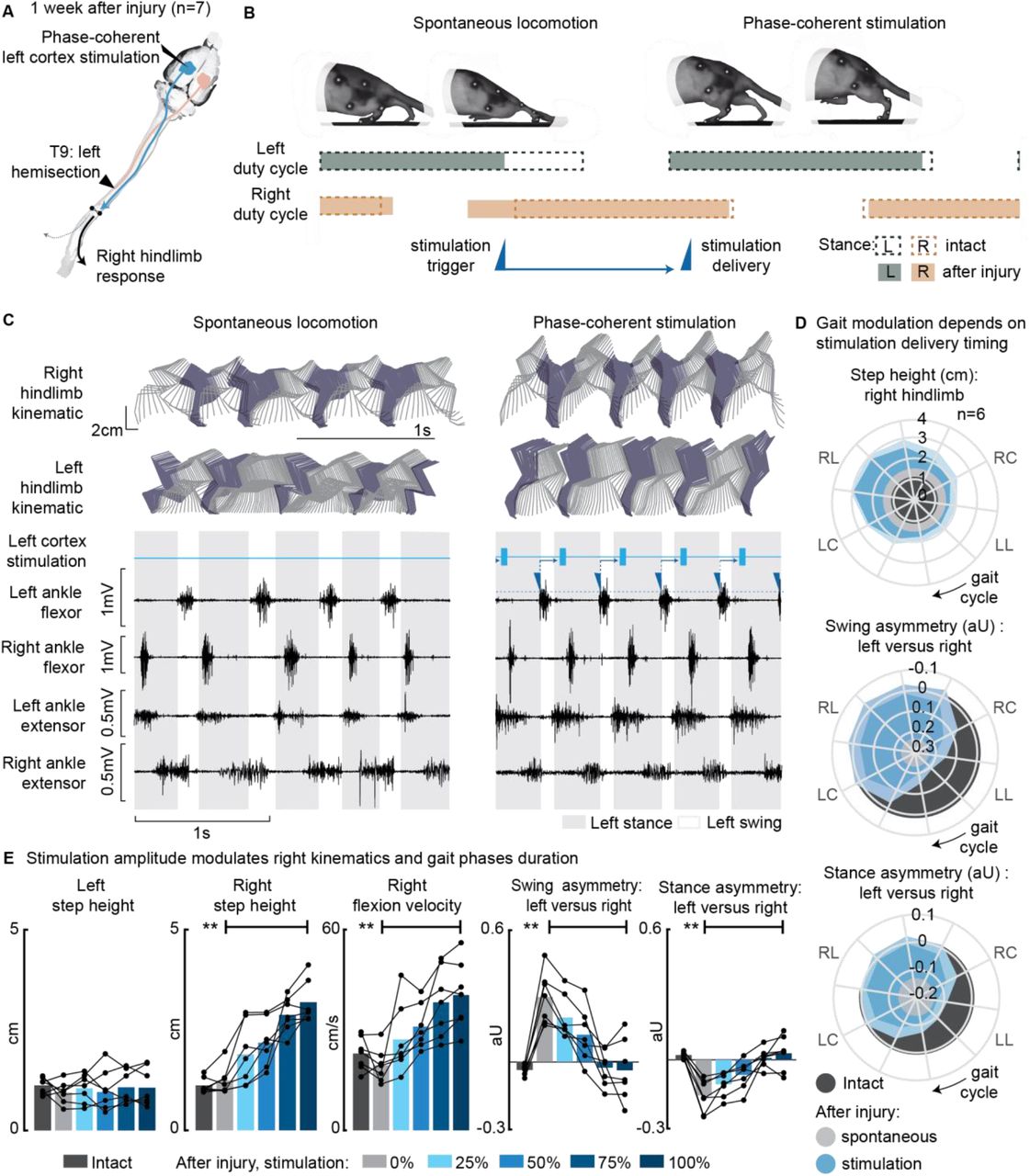
After a left spinal hemisection, rats exhibited left hindlimb motor deficits, i.e. on the same side as the lesion. Approximately 1 week after injury (5 to 10 days depending on the injury severity), as soon as the animal had recovered alternated plantar hindlimb stepping, locomotor behavior was evaluated in n=7 rats. The lack of left hindlimb support as well as weaker left flexion and extension induced asymmetries in the gait pattern (left swing dominance 29±4% p=2E-4, right stance dominance 16±3% p=1E-4, Fig. 2E). Phase-coherent stimulation of the left motor cortex (see scheme in Fig. 2A) proportionally enhanced contralateral step height. This effect was behaviorally expressed as a bilateral synergy, composed of a contralateral hindlimb flexion and an ipsilateral extension. Consequently, ipsilateral weight-bearing was intensified and prolonged, reversing the motor deficits and promoting the recovery of the balanced gait phase distribution between the left and right hindlimbs (Fig. 2B-C, Video 1). The maximal effects were obtained for stimuli delivered during the preparation and early execution of the right swing (as seen for the intact condition). These strongest effects included an increase in contralateral step height (+85±18%, p=0.008) as well as a counterbalance between swing (p=0.003) and stance durations (p=0.006) that reversed the asymmetry deficit up to 116±11% and 115±9% respectively, compared to intact walking (Fig. 2D). When delivered during the late right or early left stance, stimulation amplitude linearly modulated right step height (+172±30%, p=3E-6) and flexion velocity (+115±22%, p=4E-4). In addition, the swing (deficit reversed up to 123±10%, p=4E-4) and stance (deficit reversed up to 125±10%, p=5E-4) asymmetry indexes proportionally decreased (linear fits R2=[85±4, 86±4, 80±7, 86±5]%, Fig. 2E).

(A) A schematic representation of the injury and neurostimulation model showing the thoracic left hemisection (T9) and left (ipsilesional) motor cortex stimulation. (B) A schematic representation of spontaneous locomotion and phase-coherent stimulation effects on postural changes, gait phase duration and alternation as well as stimulation trigger and delivery timings. The stimulation, triggered in correspondence with the left lift and delivered just before the right lift, resulted in a stronger right swing and a synchronous stronger left stance. (C) Stick diagrams and EMG activity during spontaneous locomotion and phase-coherent stimulation. The stimulation was triggered by left ankle flexor activation and was delivered during the late right stance (early left stance). (D) Polar plots showing contralateral step height (cm) and gait phase asymmetry variations (aU) for stimulation delivered at different timings along the whole gait cycle. Positive asymmetry index values refer to left-side dominance. For ease of reading, the radial axis of the swing symmetry plot has been inverted (outer values are negative). For the three polar plots, the condition of strongest neuromodulation corresponded to the largest radius. Gait phase symmetry, highly affected during spontaneous locomotion, was recovered for stimulation delivered after the left contact and before the right contact. The gait cycle progresses clockwise, and the gait event references are reported as LC: left contact, RL: right lift, RC: right contact, and LL: left lift. (E) The contralateral kinematics and gait phase durations were linearly modulated with increasing stimulation amplitudes. Positive asymmetry index values refer to left-side dominance. Phase-coherent stimulation generated an increase in the step height and flexion speed of the right hindlimb and mediated the recovery of the physiological symmetry between the left and right swing and stance phases. The data are represented as the mean ± SEM. ** p < 0.01. See Fig. S1 for additional details about the rats’ hemisection profiles.
We then assessed the effects of phase-coherent stimulation on muscle activity (Fig. 3A). The ipsilesional ankle extensor was remarkably affected during spontaneous locomotion after the injury (−72±4% burst duration p=0.007, −92±2% total activation p=0.007, with respect to intact conditions, Fig. 3B). Phase-coherent stimulation reinstated the contribution from this muscle, increasing burst duration (90±18% of the deficit, p=0.004, Fig. 3B) and total activation (56±13% of the deficit, p=0.004, Fig. 3B) linearly with the injected stimulus amplitude (R2=[84±7, 84±10]%). After injury, rats displayed low posture caused by the loss of weight acceptance on their left hindlimb. Postural deficits depended on the SCI severity (Fig. 4A). We found that during the recovery process, postural compensation occurred, culminating in an exceedingly elevated posture in chronic rats (Fig. 4B). Phase-coherent stimulation, delivered in the early left stance, immediately alleviated postural deficits 1 week after injury and the iliac crest height proportionally increased with greater stimulation amplitudes (p=0.03, R2=76±9%, Fig. 4C).

(A) EMG envelopes during spontaneous locomotion before and after injury as well as phase-coherent stimulation after injury. Activities of the left and right ankle flexor (tibialis anterior) and left and right ankle extensor (medial gastrocnemius). The gait event references are reported as LC: left contact, RC: right contact. (B) Left medial gastrocnemius activity was modulated by the stimulation. The burst duration and the level of muscle activation were linearly increased with greater stimulation amplitudes. The data are represented as the mean ± SEM. ** p < 0.01. See Fig. S1 for additional details about the rats’ hemisection profiles.

Posture is shown as the height of the left iliac crest during locomotion with respect to the spontaneous condition before injury. The data are represented as the mean ± SEM. (A) Postural deficits depend on injury severity. Rats with severe SCI exhibit a weaker posture 1 week after injury. (B) Variation over 1 month of spontaneous recovery. Posture is raised and overcompensated. (C) Effect of phase-coherent stimulation 1 week after injury. Posture is increasingly raised with greater stimulation amplitudes.
Awake motor maps
In n=12 awake rats, we collected ipsilesional cortical maps, measuring cortical transmission to both hindlimbs for 8 weeks following SCI (Fig. 5A). The lesion initially decreased corticospinal transmission on both sides: 5 days after injury the ipsilateral (left cortex to left hindlimb) and contralateral (left cortex to right hindlimb) transmissions were decreased by –90±7% and –53±13% (p=[2 E-4, 3 E-4], Fig. 5B), respectively. The map size substantially increased 2 weeks after injury (+238±28% p=1E-4, Fig. 5B). Over time, the representation of bilateral hindlimb movements significantly increased compared to the intact condition (+124±30%, p=0.005, Fig. 5B).
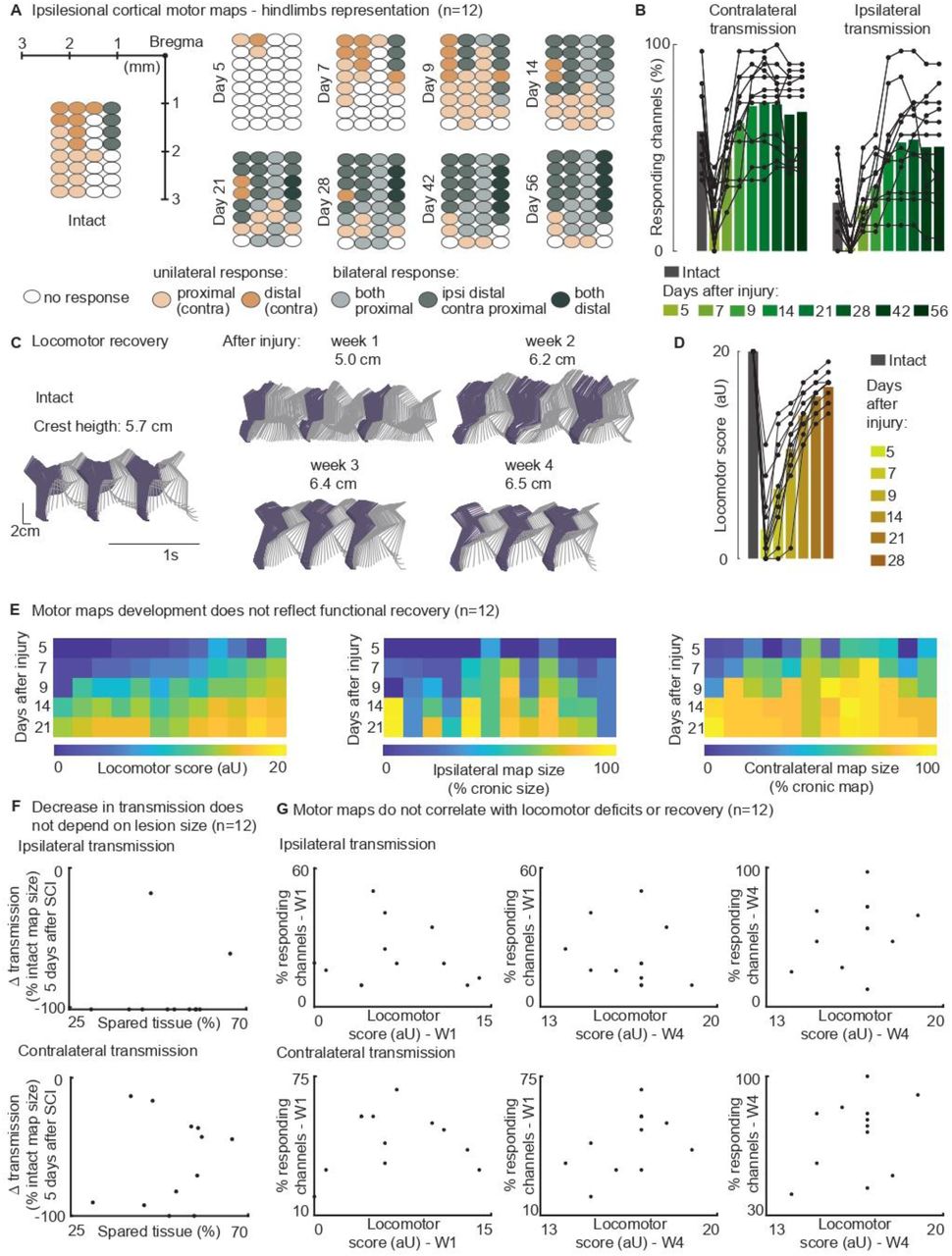
(A) Awake cortical motor map representation before injury and up to 2 months after injury. The responding channels were identified by the rule of majority voting (a site response in at least n=6 rats out of 12). Bilateral representation of hindlimb movements increases over time compared to the intact condition. (B) Quantification of responding channels from the intact condition and up to 2 months after injury. (C) Stick diagrams from treadmill locomotion and iliac crest height before injury and during the first 4 weeks after injury. (D) Quantification of locomotor score from the intact condition and up to 1 month after injury. (E) Cortical transmission and locomotor performance. An increase in map size did not correlate with motor recovery. (F) Lack of correlation between map size and lesion size 5 days after injury. An ipsilateral and contralateral decrease in transmission does not parallel the spared tissue at the lesion epicenter. (G) Lack of correlation between map size and locomotor score. Time points are reported as W1: week 1 and W4: week 4. Ipsilateral and contralateral transmission does not parallel the motor deficit, predict long-term performance nor reflect recovery. Bars: mean of individual data points.
The upregulation of cortical transmission and postural changes during spontaneous locomotion were mostly expressed within the same timeframe, specifically 1 to 2 weeks after SCI (Fig.5A-D). Between weeks 1 and 2, 91±22% of the overall postural correction (Fig. 4B, 5C) and 71±2% of the overall motor score recovery occurred (Fig. 5D). 83±7% of the ipsilateral map size post-lesional increment took place within the same time interval (Fig. 5B). Nevertheless, we found that motor map development did not correlate with functional motor recovery across subjects (Fig. 5E-G).
Ipsilateral neuromodulation of hindlimb flexion
Cortical control of hindlimb movements in behaving rats has been primarily associated with contralateral limb flexion and elevation (Bonizzato & Martinez, 2021; Bonizzato et al., 2018; DiGiovanna et al., 2016; Rigosa et al., 2015). Our results presented thus far show that for all tested animals, extensor synergies in the ipsilesional hindlimb emerged within the framework of a contralateral hindlimb flexion. Nevertheless, in n=2 rats, an additional distinct motor response was represented in a medial area of the motor cortex. In these rats, that were tested 2 weeks after SCI, stimulation of specific array sites preferentially evoked ipsilateral flexor responses (Fig. 6, rat#1: 3 channels with 271±36% ipsilateral dominance, rest of responding channels 43±4%, p=0.006, rat#2: 6 channels 452±87%, all others 18±4%, p=5E-4, Fig. 6D). The site with the highest ipsilateral dominance (rat#1: 327±109%, rat#2: 692±84%) was chosen to test the modulation of ipsilateral swing trajectories (Fig. 6B). Stimuli delivered during the late left stance resulted in kinematic modulation: step height (+133±18,+99±23%, p=[1E-4,0.001]) and flexion velocity (+46±19,+101±19%, p=[0.01,1E-4]) increased linearly (rat#1 R2=[90, 91]%, rat#2 R2=[95, 86]%) with greater stimulation amplitudes (Fig. 6E, Video 2). As a result, dragging was immediately alleviated (−46±6, −100%, p=[1E-6,7E-4]). This result was unique for ipsi-dominant cortical sites; no other tested electrode produced ipsilateral flexion facilitation (see Fig. 2E). The sites produced a similar functional effect as contralesional cortical stimulation (Bonizzato & Martinez, 2021).

(A) A schematic representation of the injury and neurostimulation model. After lateral hemisection, ipsilesional motor cortex stimulation evoked ipsilateral responses. (B) Stick diagrams and EMG activity during spontaneous locomotion and phase-coherent stimulation. The stimulation was triggered by right ankle flexor activation and was delivered during the late left stance. (C) Samples of single train stimulation of specific channels that preferentially evoked ipsilateral muscle activation in two different animals. (D) Ipsilateral dominance of EMG responses. Awake cortical motor maps were obtained as a ratio between left and right tibialis anterior activation. Channels that presented a left preference were located in the most medial region of the map. (E) Phase-coherent stimulation modulated ipsilateral kinematics and reduced the foot drop deficit. Left step height, flexion speed, and dragging alleviation linearly increased with greater stimulation amplitudes. Two subjects are presented independently, n=10 steps per condition. The data are represented as the mean ± SEM. *, ** p < 0.05 and < 0.01, respectively.
Cortical neuromodulation of hindlimb alternated rhythms
We next asked whether long-train intracortical stimulation in awake resting rats can evoke complex multi-modal motor responses (Graziano et al., 2002) and whether the effects on hindlimb movement are bilateral. The stimulation was 250ms in duration, approximately the time scale of locomotor movement preparation and initiation (Bonizzato et al., 2021). In n=6 intact rats, we found that long-train stimulation of one motor cortex evoked locomotor-like rhythms (Fig. 7A-B, Video 3), expressed by bilateral alternated whole-leg movements. We then determined that one week after unilateral hemisection SCI, long-train stimulation of the ipsilesional cortex failed to evoke bilateral rhythms in half of the tested rats. These rats exhibited a more severe injury phenotype, which was also confirmed by lower ladder crossing performance. Bilateral alternated locomotor-like rhythms returned by week 2 or 3 in all rats (Fig. 7C). The remaining three less severe rats displayed bilateral alternated hindlimb rhythms when receiving ipsilesional cortical stimulation as early as one week after injury (Fig. 7D). Classically, stimulation studies of cortical control and recovery of movement are often performed under ketamine sedation (Brown & Martinez, 2018; Nudo et al., 1996). To visualize the well-characterized absence of rhythmic hindlimb activity after ketamine sedation, n=1 intact rat was also tested and recorded before and after ketamine injection, confirming suppression of rhythmic hindlimb responses (Fig. 7E).

(A) Schematic representation of the locomotor-like rhythmic movements evoked by long-train (250ms) cortical stimulation. Evoked rhythms are characterized by alternated hindlimb movements. (B) In all n=6 tested intact rats, stimulation of the left cortex generated bilateral alternated hindlimb rhythms. After SCI, rats are sorted by injury severity, using their ladder score at week 1 for ranking. One week after injury, long-train cortical stimulation failed to evoke bilateral alternated rhythms in half of the cohort. In two of these rats, contralateral rhythms were still present and bilateral alternated rhythms were recovered by week 2. In the most severe rat, contralateral-only rhythms were evoked on week 2 and bilateral alternated rhythms on week 3. For the remaining half of the cohort, long-train cortical stimulation recruited bilateral alternated rhythms at all tested time points. (C) Stimulus-synchronized ankle flexor EMG traces from n=1 rat with a moderate-severe injury, showing loss (week 1) and following recovery (week 2-3) of ipsilateral evoked hindlimb rhythms. (D) Stimulus-synchronized EMG trace from n=1 rat with mild injury, showing that bilateral alternated evoked rhythms are preserved at week 1. (E) Stimulus-synchronized EMG trace from n=1 intact rat before and after ketamine sedation, showing transient loss of bilateral alternated rhythms.
Discussion
A cortical neuroprosthesis facilitates the control of ipsilateral hindlimb extension
In this study, we demonstrated that after a lateralized SCI, the ipsilesional cortex (with most of its crossed efferences preserved) exhibited an increased contribution in controlling bilateral hindlimb movements. An ipsilateral cortical neuroprosthesis alleviated SCI-induced locomotor and postural deficits across different levels of injury severity (Fig. S1, Table 1). The lateralized lesion model and phase-coherent cortical stimulation revealed functional ipsilateral motor control. The evoked movement was characterized by flexion of the contralateral hindlimb with a synchronous extension of the ipsilateral hindlimb. Thus, the ipsilesional motor cortex can activate bilateral lumbar synergistic networks through descending connections spared from the injury. We suggest that the acute expression of this bilateral synergy (1 week after injury) is compatible with adaptive or compensatory upregulation of pre-existing functional networks after SCI. Rapid onset of postural compensation is also displayed behaviorally by rats during the same timeframe (Fig. 4B). Although this outcome may reflect the participation of several supralesional networks, lateralized injuries capitalize the role of the ipsilesional cortex in voluntary postural and weight-bearing adjustments. We propose that this phenomenon reflects a need to preserve the functional role of the motor cortex in modulating contralateral step height during locomotion. In the absence of appropriate support from the opposite hindlimb due to the injury, the ability to elevate the foot would be compromised. Therefore, cortex-driven descending pathways may increase the excitatory transmission to ipsilesional extensor networks, thus recovering appropriate hindlimb support and retrieving precise functional control of contralateral step height.
Rats marked with * did not receive left motor cortex implantation. They were included in the study for establishing spontaneous changes in posture over time (Fig. 4A-B).
Ipsilesional motor map progression after SCI did not correlate with spontaneous recovery
After a unilateral cortical injury, plastic changes are observed in the opposite hemisphere (Axelson et al., 2013; Rehme et al., 2012; Shimizu et al., 2002; Strens et al., 2003; Witte et al., 2000). Laterally unbalanced SCI induces asymmetric activity changes in the contralesional and ipsilesional cortex, which may participate in functional recovery or compensation mechanisms (Bonizzato & Martinez, 2021; Brown & Martinez, 2018; Brown & Martinez, 2019a, 2021). We tracked the progression of motor representation of both hindlimbs in the ipsilesional motor cortex and found that in all animals, cortico-spinal transmission significantly decreased after injury (Fig. 5B), independently from the subject-specific size of the injury (Fig. 5F). This finding is consistent with a major loss of connectivity, including damage to the uncrossed ventral CST (Weidner et al., 2001) and ipsilateral cortico-reticulo-spinal transmission (Bonizzato & Martinez, 2021). The loss of excitability was quickly recovered within 2 weeks (Fig. 5B), with a return of cortico-spinal transmission consistent with the upregulation of the descending pathways spared from the injury. However, the subject-specific evolution of cortical motor maps in this timeframe did not correlate with the behaviorally expressed motor performance (Fig. 5G). Conversely, we previously showed that contralesional cortical map changes tightly correlated with locomotor recovery (Bonizzato & Martinez, 2021). Comparison of these two results suggested that recovery of hindlimb movement after SCI may be more tightly connected to changes in the contralateral cortical motor representation rather than the ipsilateral cortical motor representation, even in fully lateralized thoracic injuries, which disproportionally affect the crossed projections from the contralateral cortex. Nevertheless, since acute cortical inactivation 3 weeks after SCI immediately reinstated bilateral hindlimb deficits, we previously concluded that the ipsilesional cortex also supports locomotor recovery in this SCI model (Brown & Martinez, 2018). These combined results suggested that, although not a precise predictor of motor performance, the return of bilateral cortico-spinal transmission from the ipsilesional cortex after SCI is an important excitatory drive that supports bilateral skilled hindlimb movement.
A cortical neuroprosthesis facilitates the control of ipsilateral hindlimb flexion
We demonstrated a distinctive case of ipsilateral flexion modulation, which requires specific consideration. In two rats, phase-coherent stimulation of a cortical sub-region predominantly modulated the ipsilateral hindlimb flexion. The localization of the specific channels to the most medial region of the array (Fig. 6D), hence closest to the interhemispheric fissure, may suggest the involvement of transcallosal interactions in mediating transmission of the cortical command generated in the ipsilateral cortex (Brus-Ramer et al., 2009). However, the hemisection (particularly severe in one of the two animals, see Table S1) should exclude a contribution from the interrupted left CST. Further experiments are required to understand the mechanism(s) underlying this unconventional instance of cortical control of movement. A compelling research question arising from these results is whether similar findings can be found in the primate motor cortex.
A cortical neuroprosthesis unveiled ipsilateral functional control of movement
Multiple hypotheses have been proposed to explain ipsilateral motor cortical activity during movement (Ames & Churchland, 2019), including the following: (1) an abstract limb-independent representation of movement (Porro et al., 2000), (2) an efference copy of signals generated by the contralateral cortex (Ganguly et al., 2009), (3) uncrossed descending connectivity (Weidner et al., 2001), (4) bilateral termination of crossed descending connectivity (Rosenzweig et al., 2009) and (5) distribution of motor cortical computations across the two hemispheres (Ames & Churchland, 2019; Li et al., 2016). Our results contribute to this debate and establish specific causal links in brain-behavior interactions (Silvanto & Pascual-Leone, 2012). We employed cortical stimulation that was delivered during endogenous execution of movements and provided evidence of cortical-mediated control of functional, complex, and diverse ipsilateral movements in the rat. Our results do not support the hypothesis that ipsilateral motor cortex participation in movement is solely epiphenomenal, such as a purely abstract representation or a lone efference copy. Furthermore, lateralized injury completely abolished all uncrossed descending connectivity in a subset of animals, whereas the described effect persisted. Thus, our results are consistent with the integration of the latter two proposed explanations described above: crossed cortical descending projections (either direct or indirect through brainstem relays (Asboth et al., 2018)) display a high degree of bilaterality and complexity in both rats (Bonizzato & Martinez, 2021) and primates (Rosenzweig et al., 2009), a feature which underlies an equally complex bilateral distribution of motor cortical computations (Aizawa et al., 1990). This hypothesis is further supported by the finding that ipsilesional cortical inactivation in the rat immediately reinstates leg control deficits 3 weeks after hemisection (Brown & Martinez, 2018).
Long-train cortical stimulation recruits spinal locomotor circuits
The short duration of the stimulus train used during phase-coherent stimulation experiments can limit the display of complete, coordinated movements which may be evoked and modulated by cortical networks when activated for a time scale of hundreds of milliseconds (Graziano et al., 2002), such as what happens endogenously when movement is initiated (Bonizzato et al., 2021). We used long-train cortical stimulation in resting animals to unfold a complex generation of locomotor-like rhythms. The evoked movements were highly coordinated bilaterally across the entire hindlimb system. We interpret the apparent discrepancy between the selective outcomes of short duration stimulation, which recruits a predictable unilateral flexion or extension pattern, with the alternate recruitment of both flexion and extension within each leg (Video 3) produced by long stimulation, in light of the different excitability of spinal circuits within the dissimilar contexts of walking or being elevated with no contact to the ground. Afferent inputs are critical for spinal locomotion, nevertheless here we show that unilateral cortical drive can recruit the spinal locomotor circuits evoking alternated “air-stepping” in awake rats in absence of cutaneous interaction with a ground surface. We also show that thoracic hemisection can initially restrict the effects of cortical excitation to the unilateral generation of spinal rhythms, suggesting that cortical projections jointly recruit independent (i.e., side-specific (Grillner & Wallen, 1985)) rhythm-generating spinal units. Recovery of bilateral alternated rhythms within 2-3 weeks after hemisection suggests changes within the spinal circuitry below the lesion, possibly mediated by the persistent interaction between commissural interneurons and efferences responsible for cortico-spinal transmission (Gossard et al., 2015; Martinez et al., 2011). The role of supraspinal drive on spinal locomotor circuits has been previously discussed with respect to “fictive” locomotion (decerebrate) preparations. In the cat, pyramidal stimulation was found to reset the locomotor rhythm by initiating bursts of activity in either extensor (Leblond et al., 2001) or flexor muscles (Orlovsky, 1972), but repetitive burst stimulation was required to temporize cycles into a structured rhythm, which falls short to the rhythms-evoking capacity we demonstrated through long-train cortical stimulation in awake rats.
Ipsilateral cortical control of movement
In summary, our results show that short phasic cortical stimulation is mediated by a gating system allowing kinematic expression of selective (flexion/extension) cortical commands to both hindlimbs in the correct phase of the locomotor cycle; long cortical stimulation triggers spinal locomotor circuits, transforming unilateral cortical neuromodulation into bilateral alternated output. Hence, the rodent motor cortex displays a complex executive relationship with spinal networks which mediate cortical initiation and modulation of ongoing movement, overall allowing a remarkable bilateral efferent transmission, which coherently integrates and regulates spinal states. Movement is generated by distributed cortical, subcortical, brainstem, and spinal networks, each of which is strongly functionally interconnected with contralateral counterparts. Multiple cortical networks involved in the generation of movement have been shown to become activated in a limb-independent mechanism. In the dorsal stream of visuomotor processing, the posterior parietal cortex contributes to grasping (Kermadi, 2000) or locomotor movements such as obstacle avoidance (Andujar et al., 2009). During the execution of these tasks, neurons responding to both left and right limb movement predominate. Premotor cortical areas also harbor neurons that become activated during ipsilateral movement (Cisek et al., 2003; Kermadi, 2000; Michaels & Scherberger, 2018). Our results indicated that this bilaterality is not extinguished in the cortical line of sensorimotor integration. Instead, it is at least selectively preserved in the functional network properties of the ultimate cortical actuator of movement, the primary motor cortex.
Cortical neuroprostheses
These results, beyond providing new evidence of complex ipsilateral control of movement in mammals, have potential translational implications for the future development of neuroprosthetic solutions. Since cortical stimulation immediately alleviated motor deficits in rats, it may also promote more efficient movement execution in individuals with lateralized SCI or hemiparesis due to cortical or subcortical stroke. Improved motor performance may lead to a broad range of potential benefits, including better and more sustained access to activity-based training. A limitation of this potential treatment is the invasive nature of the intracortical interface utilized in the rats. Less invasive solutions exist including transcranial magnetic stimulation, which requires further targeted research since (1) it has not yet been tested as a ‘priming’ agent for movement in the subacute phases of neurotrauma (Smith & Stinear, 2016) and (2) it is usually intended as an inhibitory agent for the non-lesioned cortex (Nowak et al., 2009), in line with the interhemispheric inhibition stroke model. A clear trade-off between invasiveness and efficacy of neurostimulation techniques needs to be established to determine which set of neurostimulation methods holds the potential to improve the generation of cortical motor commands in individuals with neurotrauma.
Materials and Methods
Experimental model and subject details
Animals
All procedures followed the guidelines of the Canadian Council of Animal Care and were approved by the Comité de déontologie de l’expérimentation sur les animaux (CDEA, animal ethics committee) at Université de Montréal. A total of n=16 (see Table S1) female Long-Evans rats (Charles River Laboratories, line 006, 270-350 g) were used in this study. Additional rats (n=25) were added to analyze spontaneous postural changes after injury (Fig. 4A-B). After a period of acclimatization and handling habituation, rats were trained to walk on a motorized treadmill with positive reinforcement (food). The rats were housed in a group (n=3) before surgery and were housed individually after implantation. Blinding did not apply since kinematic analysis was automatically performed by DeepLabCut. The output was curated to avoid detection mistakes, and corrections involved less than 0.5% of the conditioned points.
Study design
The number of animals used in this study was determined based on power analysis. The objective of this study was to maximize the extension/stance phase and promote the weight support recovery of the affected hindlimb after unilateral SCI. At the beginning of the study, we ran a pilot experiment with two animals and found that ipsilesional phase-coherent intracortical stimulation increased the contralateral stance phase duration by > 80% of the intra-subject variability. Under this condition, power analysis estimated a 97% probability of achieving significant results (α=0.05) with n=5 subjects and a 99% probability with n=6 subjects (one-sided, paired t-test). We characterized n=6 intact animals. For SCI subjects, we increased the sample size to n=7 to allow sufficient power for electromyographic (EMG) investigation. Starting with recordings from n=7 rats and excluding poor quality signals, we obtained an EMG analysis with n=5 animals for each muscle.
Method details
Surgical procedures
All surgical procedures were performed under isoflurane general anesthesia. Lidocaine (2%) was delivered at the incision sites. Analgesic (buprenorphine) and antibiotic (Baytril) were administrated for 3-4 days after surgery. In the first surgery, we implanted the EMG electrodes and the intracortical array. Differential EMG wires were inserted into the left and right tibialis anterior and medial gastrocnemius muscles. Common ground wires were subcutaneously placed around the torso. After a craniotomy and removal of the dura matter from the left motor cortex hindlimb area, a Tucker-Davis Technologies 32-channel array (8 rows, 4 columns, 1.125 × 1.75 mm) was inserted into the cortical layer V (1.5 mm depth) with the top-right site of the array positioned at coordinates [1.1 mm posterior, 1.1.mm lateral] from bregma. The EMG connector and intracortical array were then embedded in dental acrylic and fixated on the head with four screws. During the second surgery, rats received SCI. A partial T9 laminectomy was performed and 2% lidocaine was used to lower spinal reflexes, followed by left spinal cord hemisection(Brown & Martinez, 2019b). In rats that were unable to micturate, their bladders were manually expressed for a few days after injury, until they spontaneously regained control of micturition.
Behavioral assessments
The following three tasks were used to assess the motor performance of the rats: (1) ladder crossing, (2) open-field, and (3) treadmill. The ladder test was used to evaluate skilled locomotion. In this test, rats were recorded (100 frames/s) while crossing a horizontal ladder (130 cm long) with regularly spaced rungs (3 mm rungs spaced by 2 cm). In each session, trials with only consecutive steps were analyzed and five trials per rat were averaged. Each trial consisted of approximately 10 steps. The scoring system was based on the foot fault score (Metz & Whishaw, 2002). At 7 days after lesion induction, the performance was used as a reference to classify the severity of the animal’s injury. The injuries were classified based on the number of partial or correct paw placements on the rungs over the total number of steps (referred to as paw placements), Thus, the injuries were classified as: (1) mild (left hindlimb > 20% paw placement), (2) moderate (left hindlimb < 20% paw placement and right hindlimb > 75% paw placement) and (3) severe (bilateral deficit, right hindlimb < 75% paw placement). The open-field test was utilized to evaluate spontaneous overground locomotion. In this test, rats were recorded (30 frames/s) during spontaneous locomotion within a circular Plexiglas arena (96 cm diameter, 40 cm wall height) with an anti-skid floor. The locomotor score was assigned using the Martinez scale (Brown & Martinez, 2019b; Martinez et al., 2009). The treadmill task was used to evaluate stimulation effects. In this task, 10 consecutive steps were considered for each trial. The treadmill speed was 23 cm/s and kinematics were captured (119.2 Hz) with the use of six reflective markers identifying the iliac crest, trochanter, knee, fifth metatarsal and fourth toe tip. The posture of the rats was evaluated from the height of the iliac crest during the gait cycle and was compared to intact rats. The ladder and open-field scoring as well as kinematic analysis were performed offline. The kinematics were tracked with DeepLabCut (Mathis et al., 2018) and manually curated to avoid misdetections.
Awake motor maps
To evaluate motor evoked responses, the 32 channels of the cortical array were tested in awake animals. A 40 ms pulse train (330 Hz, biphasic, 200 μs/phase) was delivered to each site and hindlimb responses were assessed visually while the animal was resting and held with trunk support. Starting with stimulation amplitudes of 100 μA, the response type (proximal or distal) was assessed and the minimum amplitude evoking a visible twitch was then identified as the threshold value. A joint motor map from n=12 subjects was defined by majority voting (Fig. 5). In the case of n=2 rats, in which specific channels preferentially evoked ipsilateral motor responses, EMG signals were recorded during 10 additional rounds of testing for all channels. After normalizing each muscle activity to spontaneous locomotion, we quantified the ipsilateral dominance of muscle activation as the ratio of the left and right tibialis anterior evoked responses.
Phase-coherent cortical stimulation
The phase-coherent neurostimulation strategy was previously presented in detail (Bonizzato & Martinez, 2021). The EMG activity was processed online during treadmill locomotion and a trigger event was detected when the signal crossed a manually selected activation threshold. A biphasic 40 ms train at 330 Hz was then delivered with a specific delay. Among the 32 sites of the cortical array, the stimulation channel was chosen which evoked the strongest right hindlimb flexion (or left hindlimb flexion in the case of ipsilateral modulation) during motor maps assessment. For amplitude characterization, the left flexor was always used as a synchronization signal and the delay was fixed and corresponded to 140-190 ms depending on the rat gait pattern. In this protocol, the stimulation was delivered in the late right stance or corresponding early left stance. The amplitude values were linearly spaced within a functional range defined from a minimum visible effect (40-100 µA before injury, 25-70 µA after injury) to a maximum comfortable value for the animal (125-300 µA before injury, 70-200 µA after injury). For timing characterization, the synchronization was alternatively based on the right flexor and the left flexor or right extensor activity. The amplitude was fixed and equal to a medium value of the functional range. The delay varied among trials in order for the stimulation delivery to complementarily cover the whole gait cycle (0-200 ms for the flexors, 80-280 ms for the extensor, in steps of 40 ms). In specific cases of ipsilateral kinematics modulation, the trigger was detected from the right flexor signal, the delay was fixed (160 ms, corresponding to late left stance), and the amplitudes varied within the functional range (lower bound 50 and 100 µA, upper bound 200 and 175 µA). In each characterization, trials were randomly permuted whenever possible.
Long-train cortical stimulation
For each tested rat, the cortical channel which evoked the strongest hindlimb responses was preliminarily selected by visual investigation. Awake resting rats were manually supported at the torso and forelimbs, while the hindlimbs were left relaxed with no support. Hindlimb responses to long-train stimuli (250ms, 330 Hz, biphasic, cathodic first pulses, 200 μs/phase) were captured on the camera (120 Hz). On three rats, EMG data from both ankle flexor muscles (Tibialis Anterior) was also collected synchronously (6 kHz). Stimulus amplitude was set at 100 µA for all long-train experiments. Experiments were repeated in the intact state and weekly for three weeks after SCI. One intact rat received one single dose of ketamine (120mg/kg, IP) to confirm the absence of alternated evoked rhythms in this condition. The rat was tested 10 minutes after injection, while moderately sedated (corneal and paw withdrawal reflexes were preserved, but no overt spontaneous movement). In this case, only a stimulus amplitude of 150 µA was delivered.
Current spread
Current propagation of intracortical microstimulation is regulated by a well-characterized physical law that takes into consideration the stimulation current and the propagation medium. The relationship between the distance r of the effective current spread from the electrode tip in mm and the current intensity I in µA is expressed by an equation of the form I=kr2 (Stoney et al., 1968), where the constant k for the cortical medium is equal to 1292 µA/mm2. We verified that at no point during the experiments presented in this study we had a propagation further than half-millimeter distance given that we never exceeded 300 µA threshold for the stimulation current.
Histology
At the end of the experiments, the rats were euthanized with pentobarbital administration (Euthanyl, 100 mg/kg, intraperitoneal). Transcardiac perfusion was performed with a 0.2% phosphate-buffered saline (PBS) solution followed by a 4% paraformaldehyde (PFA) solution (pH 7.4). The spinal cords were extracted and first stored in a 4% PFA solution and then in a 20% sucrose solution. To evaluate the lesions, spinal sections around the T9 segment were cut in 40 µm slices and tissue damage was assessed under a microscope. A reconstruction of the lesion profiles at the epicenter level was traced and the extent of healthy and damaged tissue was quantified.
Quantification and statistical analyses
All results are shown as the mean value ± the standard error of the mean (SEM). The statistical analyses were performed with the Student’s t-test for normally distributed populations or the Wilcoxon rank-sum test for other populations. All tests involved paired population samples. Samples with p < 0.05 were considered statistically significant.
Funding
This work was supported by the Craig H. Neilsen Foundation and the Natural Sciences and Engineering Research Council of Canada. M.M. was supported by a salary award from Fonds de Recherche Québec-Santé (FRQ-S). M.B. was supported by fellowships from the FRQ-S, the Institut de valorisation des données (IVADO), the TransMedTech Institute, and a departmental postdoctoral fellowship in memory of Tomás A. Reader. E.M. was supported by a fellowship from the TransMedTech Institute.
Author contributions
M.B. and M.M. conceived the research; E.M., M.B. and M.M. designed the experiments; M.B. developed the overall system integration; E.M. and M.B. performed the surgeries and collected the experimental data; E.M., M.B., M.M. analyzed the data; E.M. and M.B. drafted the manuscript; E.M., M.B., M.M., edited, revised manuscript and approved its final version.
Competing interests
M.B. and M.M. submitted an international patent application (PCT/CA2020/051047) covering a device allowing performing coherent cortical stimulation during locomotion. They are founders of a company developing a stimulation-based therapy to restore movement after SCI.
Supplemental figures and video

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
(A) Hemisection profiles at the epicenter level. (B) Classification of the injury severity. Injury severity groups were defined according to skilled locomotion performance during ladder crossing 7 days after injury. The injuries were classified as mild: left hindlimb > 20% paw placement, moderate: left hindlimb < 20% paw placement and right hindlimb > 75% paw placement and severe: right hindlimb < 75% paw placement (bilateral deficits).
Video 1. Cortical neuroprosthesis mediated control of ipsilateral hindlimb extension. Treadmill walking during spontaneous locomotion and with phase-coherent stimulation 1 week after injury. The stimulation was delivered during the late right or early left stance and resulted in a flexion of the right hindlimb and a synchronous extension of the left hindlimb.
Video 2. Cortical neuroprosthesis mediated control of contralateral hindlimb flexion. Treadmill walking during spontaneous locomotion and with phase-coherent stimulation 2 weeks after injury. The stimulation was delivered during the late left or early right stance and resulted in a flexion of the left hindlimb.
Video 3. Long-train cortical stimulation recruits spinal locomotor circuits. Hindlimb responses to long-train intracortical stimulation in the intact state, with and without ketamine sedation, and at two time points (1 and 2 weeks) after SCI. The stimulation was delivered in resting rats manually supported at torso and forelimbs, with the hindlimbs left free to move.
Acknowledgments
The authors would like to thank Roxanne Drainville for her participation in animal handling, Émilie Délage and Victorine Artot for participating in data analysis; Andrew Brown and Mohamad Sawan for the fruitful discussion on this work’s material and methods; Philippe Drapeau and Marc Bourdeau for technical assistance; Marjolaine Homier, Stéphane Ménard, Raphaël Santamaria and the staff at the Division des Animaleries for supporting our animal care.
References




