

Abstract
Disease tolerance describes a host’s ability to maintain health independently of the ability to clear microbe loads. However, we currently know little about the mechanisms that underlie disease tolerance or how known mechanisms of tissue damage signalling and repair may contribute to variation in tolerance. The Jak/Stat pathway plays a pivotal role in Drosophila humoral innate immunity, signalling tissue damage and triggering cellular renewal, making it a potential mechanism underlying the disease tolerance phenotype. Here, we show that disrupting the Jak/Stat pathway in Drosophila melanogaster alters disease tolerance during Pseudomonas entomophila systemic infection. Overall, flies with disrupted Jak/Stat show variation in survival that is not explained by variation in pathogen loads. For instance, mutations disrupting the function of ROS-producing dual oxidase (duox) or the negative regulator of Jak/Stat, Socs36E render males less tolerant to systemic bacterial infection but not females. We also investigated whether the negative regulator of Jak/Stat, G9a -which has previously been associated with tolerance of viral infections – is also implicated in tolerance of bacterial infection. While female flies lacking G9a showed higher mortality and reduced bacterial clearance, disease tolerance did not differ between G9a mutants and the wildtype. This suggests that G9a does not affect tolerance during systemic bacterial infection as it appears to do with viral infection. Overall, our findings highlight that Jak/Stat signalling mediates disease tolerance during systemic bacterial infection and that this response differs between males and females. Our work therefore suggests that differences in Jak/Stat mediated disease tolerance may be a potential source of sexually dimorphic response to infection in Drosophila.
Introduction
When organisms acquire infection, they face two major challenges in order to return to a healthy state. The first challenge is to identify and clear the source of the infection. While crucial, pathogen clearance alone will not result in a healthy host, because after pathogen elimination, what is left is the tissue damage caused by pathogen growth and as a side-effect of immunopathology. The second challenge to return to healthy state is therefore to minimise damage even before it occurs, and to repair and regenerate tissue after it has been damaged during infection (Martins et al. 2019; McCarville and Ayres 2018; Oliveira, Bahia, and Vale 2020; Soares, Teixeira, and Moita 2017; Vale, Fenton, and Brown 2014).
Individuals capable of dealing with the first challenge are typically labelled ‘resistant’, and exhibit low microbe loads because their immune clearance mechanisms are very effective (Boon et al. 2009; Ganz and Ebert 2010; Lazzaro, Sackton, and Clark 2006; Wang, Lu, and Leger 2017). The mechanisms underlying host resistance have been well characterized empirically and often involve detecting pathogen-derived molecular patterns such as peptidoglycans and triggering signalling cascades that result in the expression of antimicrobial peptides (AMPs) which directly kill the replicating pathogen (Kleino and Silverman 2014; Henna Myllymäki, Valanne, and Rämet 2014; H. Myllymäki and Rämet 2014; Palmer, Varghese, and Van Rij 2018; Valanne, Wang, and Rämet 2011). These responses have been described in great detail in the fruit fly Drosophila, and include the immune deficiency (Imd), Toll & Jak/Stat pathways (Bang 2019; Ferrandon et al. 2007; Hoffmann 2003; Hultmark 2003; Lemaitre and Hoffmann 2007).
Individuals with very effective mechanisms of damage limitation and repair, however, may not necessarily have a strong capacity for pathogen clearance, but will be successful in preventing or repairing tissue damage (Martins et al. 2019; Medzhitov, Schneider, and Soares 2012; Schneider and Ayres 2008; Soares, Teixeira, and Moita 2017; Soares, Gozzelino, and Weis 2014). Effective mechanisms of damage signalling and repair may explain why some individuals are tolerant of infection, and are able to experience relatively high health even if their pathogen loads remain high or are not completely cleared (Martins et al. 2019; Soares, Gozzelino, and Weis 2014). Compared to well-described pathogen clearance mechanisms, we are only beginning to unravel the mechanistic basis of disease tolerance. Likely candidate mechanisms include those that regulate inflammation to reduce immunopathology (Adelman et al. 2013; Cornet et al. 2014; Sears et al. 2011); detoxification of host or pathogen derived metabolites (Ferreira et al. 2011; Soares, Teixeira, and Moita 2017; Vale, Fenton, and Brown 2014); or tissue protection and regeneration (Jamieson et al. 2013; Soares, Teixeira, and Moita 2017; Soares, Gozzelino, and Weis 2014). However, the few disease tolerance candidate genes arising from genome-wide association or transcriptomic studies - such as ghd (grainyhead), dsb (debris buster), crebA (cyclic response element binding protein) and, dfoxo (forkhead box, sub-group O) - do not appear to be associated with these functions, or with classical immune pathways (Dionne et al. 2006; Howick and Lazzaro 2014; Lissner and Schneider 2018; Troha et al. 2018). This may indicate that disease tolerance is a complex phenotype and that dissecting its mechanistic basis may be more successful through a more focused examination of the effect of specific candidate signalling pathways.
Here we take advantage of the detailed knowledge of Drosophila infection and immunity to investigate whether damage signalling plays a role in disease tolerance during systemic bacterial infection. We focus on the Janus kinase/signal transducer and activator of transcription (Jak/Stat) pathway which is known to have important roles in signalling damage and tissue renewal. The Jak/Stat pathway was originally shown to control multiple biological processes including development, homeostasis, metabolism, and immunity in insects and mammals (Brivanlou and Darnell 2002; Harrison 2012; H. Myllymäki and Rämet 2014; Stark and Darnell 2012; Zhou and Agaisse 2012). In response to different types of stresses such as mechanical injury, oxidative stress, and infection the Jak/Stat pathway is activated by cytokine-like ligands of the unpaired family namely upd-1, upd-2 and upd-3 (Agaisse et al. 2003; Chakrabarti et al. 2016; Dostert et al. 2005; Ekengren et al. 2001; Ekengren and Hultmark 2001; Gilbert et al. 2005; Harrison et al. 1998). Upd-3 is produced during damage caused by reactive oxygen species (ROS), which in turn are produced by dual oxidase (duox) (Klebanoff 1974; Babior 1995; Lee and Kim 2014) (see Fig. 1). The extracellular binding of upd-3 to Domeless (dome), leads to the phosphorylation of Hopscotch (hop). This then leads to the phosphorylation of Stat92E, and its translocation to the nucleus (H. Myllymäki and Rämet 2014). In the nucleus, in addition to the production of factors that are necessary for repairing cellar damage, Stat92E also induces the expression of Socs36E, a negative regulator of Hopscotch (Kiu and Nicholson 2012). Recent work has also highlighted the role of the histone H3 lysine 9 methyltransferase (also called G9a) in negatively regulating the expression of the Jak/Stat pathway during infection (Merkling et al. 2015).

A simplified version of Drosophila Jak/Stat pathway known to have important roles in signalling damage and tissue renewal. In response to infection, the Jak/Stat pathway is activated by cytokine-like ligands of the unpaired family namely upd-3, produced  during damage caused by reactive oxygen species (ROS), which in turn are produced by dual oxidase (duox). The extracellular binding of upd-3 to Domeless (dome), leads to the phosphorylation of Hopscotch (hop). This then leads to the phosphorylation of Stat92E, and its translocation ↘ to the nucleus. In the nucleus, Stat92E induces the expression of Socs36E, a negative regulator
during damage caused by reactive oxygen species (ROS), which in turn are produced by dual oxidase (duox). The extracellular binding of upd-3 to Domeless (dome), leads to the phosphorylation of Hopscotch (hop). This then leads to the phosphorylation of Stat92E, and its translocation ↘ to the nucleus. In the nucleus, Stat92E induces the expression of Socs36E, a negative regulator  of Hopscotch. Recent work has also highlighted the role of the histone H3 lysine 9 methyltransferase (also called G9a) in negatively regulating
of Hopscotch. Recent work has also highlighted the role of the histone H3 lysine 9 methyltransferase (also called G9a) in negatively regulating  the expression of the Jak/Stat pathway during infection.
the expression of the Jak/Stat pathway during infection.
Focusing on its role in immunity, there is substantial evidence that Jak/Stat signalling plays a key role in wound healing, gut immunity, and downstream AMP production (Chakrabarti et al. 2016; Kemp et al. 2013; Lamiable and Imler 2014; Tafesh-Edwards and Eleftherianos 2020). For instance, during enteric bacterial infection in Drosophila, the Jak/Stat pathway contributes to intestinal immunity by regulating intestinal stem cell (ISC) proliferation and epithelial cell renewal via epidermal growth factor (EGFR) signalling (Buchon et al. 2010; Chakrabarti et al. 2016; Ohlstein and Spradling 2006). The absence of epithelial renewal leads to a loss of structural integrity and increased susceptibility to bacterial infections (Buchon et al. 2009). In cellular immunity, Jak/Stat signalling is central to the production, differentiation and maintenance of blood cells in insects (Banerjee et al. 2019; Meister and Lagueux 2003).
Jak/Stat is also important in humoral immunity to viral infection (Dostert et al. 2005), where a loss of regulation of Jak/Stat by the epigenetic negative regulators G9a results in reduced tolerance of Drosophila C virus infections due to increased immunopathology (Merkling et al. 2015). This specific result led us to question whether the effects of G9a-mediated Jak/Stat regulation on tolerance were specific to viral infection, or if the regulation of Jak/Stat also affects disease tolerance during bacterial infection. Here we investigated the tolerance response of Drosophila during septic infection with the bacterial pathogen P. entomophila, using deletion mutant flies lacking various components of Jak/Stat signalling and regulation. Further motivated by the widespread observation of sexually dimorphic immunity reviewed in (Belmonte et al. 2020; Klein and Flanagan 2016) and particularly that the effects of G9a on tolerance of DCV infection are more pronounced in female flies (Gupta and Vale 2017; Merkling et al. 2015), we also focused on assessing how Jak/Stat affects males and females separately.
Materials and methods
Fly strains and maintenance
we used several D. melanogaster strains with loss-of-function mutations for the Jak/Stat pathway. All the fly strains were on the same genetic background as that of yw (189) including Domeless (Bloomington stock# 14726), Hopscotch (15097), Socs36E (16744), Duox (16468). We also used G9a mutant flies (i.e., G9a-/-) and control (G9a+/+), also known as G9aDD2 see (Merkling et al. 2015) for details about G9a mutants. Previous work has shown that yw flies have lower basal levels of nitric oxide and are more susceptible to infection compared to other lab wild types such as w1118, Oregon R, Canton-S (Eleftherianos et al. 2003). However, we found that males and females of both yw and another wildtype of outcrossed flies showed comparable survival following P. entomophila systemic infection and exhibited similar levels of internal bacterial loads when measured around 24 hours following infection (see supplementary Fig. S1A & B, Table S1 for survival; Fig. S1C, for bacterial load). We maintained all the fly lines in a 12ml plastic vials on a standard cornmeal diet or Lewis’s medium see (Siva-Jothy et al. 2018), at 25°C (±2°C). We used 3-5 day old adult flies for all our experiments (see below). First, we housed 2 males and 5 females for egg laying (48 hours) onto a vial containing fresh food. We then removed the adults and the vials containing the eggs were kept in 25°C incubator for 14 days, or until pupation. We placed the newly eclosed individuals (males and females separately) in fresh food vials until the experimental day (around 3 days).
Bacterial culture preparation
We used P. entomophila cultured overnight in Luria broth (LB) at 37°C under constant agitation i.e., 120 revolutions per minute (rpm). P. entomophila is a gram-negative bacterium naturally found in soil and aquatic environments, known to be highly pathogenetic for D. melanogaster (Dieppois et al. 2015; Vodovar et al. 2005)). Upon reaching ∼0.75 OD600 we pelleted the culture using a 4°C cold centrifuge during 5 minutes at 5000rpm and then removed the supernatant. We resuspended the bacteria in 1X phosphate buffer saline (PBS) and prepared the final infection dose of OD600 of 0.05 for all our infection assays.
Systemic infection assay
We used a split-vial experimental design (see Fig. 2), wherein after infection each vial containing about 25 flies (of each treatment, sex and fly line combination) were divided into 2 vials for measuring (A). survival following infection (∼15 flies/combination) and (B). internal bacterial load (see Fig. 2II). With this split-vial design we were able to use replicate-matched data for both survival and bacterial load in order to estimate disease tolerance for each fly line (i.e., fly survival with respect to internal bacterial load). We infected 3-5-day old male and female adult flies using a 0.14mm insect minutein needles bent at 90° angle to avoid damaging the internal tissues by dipping in P. entomophila bacterial solution. For mock controls we substituted bacterial solution with sterile 1X PBS. After stabbed the flies in the sternopleural region of the thorax (Khalil et al. 2015). We then placed males and females separately onto fresh food vials and incubated at 25°C. We scored the flies (both infected and control) every 2-3 hours for the first 48 hours following infection, then 2-3 times each day for the next 6 days (150 hours).

(I) Design of experiments to assay (1) survival (n= ∼15 vials of around 25 flies/vial/infection treatment/sex/fly line) and (2) internal bacterial load following systemic bacterial infection with Pseudomonas entomophila to test the role of the Jak/Stat pathway (n= ∼15 vials of around 25 flies/vial/infection treatment/sex/fly line) on (3) disease tolerance. Each point in disease tolerance (3) panel represents replicate-matched data [n= ∼15 vials/infection treatment/sex/fly line – with each vial containing around 25 flies] from survival (1) and bacterial load (2). (II) A split-vial experimental design, wherein after infection each vial containing about 25 flies of each treatment, sex and fly line combination were divided into 2 vials for measuring (A). survival following infection (∼15 flies/combination) and (B). internal bacterial load (∼10 flies/combination).
Measuring bacterial load
To quantify internal bacterial load after P. entomophila infection first, we thoroughly washed each fly with 70% ethanol for ∼30 sec to surface sterilize and then rinsed twice with autoclaved distilled water. We plated the second wash on LB agar plates to confirm whether the surface bacteria were successfully removed after sterilization. We then transferred individual fly onto 1.5ml micro centrifuge tubes and homogenized using a motorized pestle for approximately 30-60 seconds in 100μl LB broth (n=30 homogenates/sex/infection treatment/ fly line). We performed serial dilution of each fly homogenate up to 10−6 fold and added 4μL aliquot on a LB agar plate. We incubated the plate overnight for ∼18h at 30°C and counted the resultant bacterial colonies manually (Siva-Jothy et al. 2018). We note that mock-infected control fly homogenates did not produce any colonies on LB agar plates.
Statistics
Survival: We analysed the survival data with a Cox mixed effects model using the R package ‘coxme’ (Therneau 2015) for different treatment groups (P. entomophila systemic infection and mock controls) across males and females. We specified the model as: survival ∼ fly line * treatment * sex * (1|vials/block), with ‘fly line’, ‘treatment’ and ‘sex’ and their interactions as fixed effects, and ‘vials’ nested within a ‘block’ as a random effect for wild type yw and Jak/Stat mutant fly lines.
Bacterial load: We found that the bacterial load data were not normally distributed (tested with Shapiro–Wilks’s test for normality). Hence, we first log transformed the data and then confirmed that the transformed residuals were still not normally distributed. Hence, we used a non-parametric one-way ANOVA Kruskal-Wallis test to test the effects of each fly lines (wild type yw and Jak/Stat mutants) on males and females following bacterial infection.
Measuring disease tolerance: Finally, to understand how the Jak/Stat signalling contributes to the phenotype disease we analysed tolerance to bacterial infection by linear reaction norm (i.e., measuring the linear relationship between fly survival against bacterial load) (Raberg, Sim, and Read 2007; Ayres and Schneider 2012; Louie et al. 2016; Oliveira, Bahia, and Vale 2020). We analysed using ANCOVA for each of the fly lines (wild type yw and Jak/Stat mutants) by fitting ‘fly line’ and ‘sex’ as categorical fixed effects and ‘bacterial load’ as a continuous covariate and their interactions as fixed effects. Because we were interested in identifying how the Jak/Stat mutants differ from wildtype flies during disease tolerance - we compared the estimates of the slope using pairwise comparison (f-test; w1118 vs. different Jak/Stat mutants) to find out what extent these Jak/Stat mutants significantly differed from yw wild type in tolerating bacterial infections.
Results & discussion
1. Following systemic bacterial infection, loss of different components of Jak/Stat pathway result in variable survival outcomes
Overall, we found that disruption of Jak/Stat signalling affected fly survival during bacterial P. entomophila infections (Fig. 3A and B, Table 1 & S2). Both male and female flies lacking duox (ROS producing dual oxidase) were more susceptible to P. entomophila infections compared to the wildtype (yw) (Fig. 3A and B, Table 1 & S2). However, other mutants showed improved survival relative to wild types. These included male and female flies lacking the transmembrane receptor domeless, males lacking the negative regulator Soc36E and both sexes lacking hopscotch (see hazard ratio in Fig. 3B, Table 1 & S2).
Summary of estimated hazard ratio from the cox proportional model. A greater hazard ratio (>1) indicates that Jak/Stat mutant flies are more susceptible to P. entomophila infection than yw wildtype flies while lower ratio (<1) indicates that Jak/Stat mutants have better survival than wildtype.

(A) Survival curves for wildtype yw flies and flies lacking Jak/Stat pathway components for females and males exposed to systemic P. entomophila of infection dose OD600=0.05 (n=20-30 flies/vial (∼15 vials)/fly line/treatment/sex/infection dose). [‘*’ indicates that the Jak/Stat mutants are significantly different from yw flies]. (B) Estimated hazard ratios calculated from the survival curves for males and female flies (yw & with flies lacking components of Jak/Stat). A greater hazard ratio (>1) indicates higher susceptibility of Jak/Stat mutants than wild type while (<1) indicates Jak/Stat mutants have better survival than wild type yw flies to systemic bacterial infection (p=<0.05). (C) Bacterial load (mean ln10) measured around 24 hours following infection. [significantly different fly lines are connected with different alphabets using Tukey’s HSD as a post hoc analysis of pairwise comparisons].
2. Both wildtype (yw) and Jak/Stat mutant flies exhibit similar bacterial loads
Next, we tested whether the variation we observed between mutants in mortality could be explained by differences in their bacterial load. Given that most mortality occurred just after 24 hours for most of our fly genotypes (Fig. 3A) we quantified bacterial load at 24 hours following infection. Both wild type and Jak/Stat mutants exhibited similar levels of bacterial load around 24 hours following infection with P. entomophila (Fig. 3C, Table S3). Therefore, despite no substantial difference in microbe loads at 24 hours post infection, Jak/Stat mutants showed variable survival. This would fit the functional definition of disease tolerance as for the same bacterial load some lines appear to be more tolerant (survive longer, such as domeless) while others are less tolerant (e.g., duox).
3. During systemic bacterial infection, loss of Jak-Stat signalling leads to sex differences in disease tolerance phenotypes
While the results above are indicative of variable tolerance depending on the Jak/Stat mutation, we carried out a formal analysis of disease tolerance using the slope of the linear reaction norm between fly survival and microbe load, where each data point is the matched survival / CFU data for one replicate vial (see methods for description of split-vial design and Fig. 2). Here, the differences in tolerance between Jak/Stat mutant and yw fly lines are indicated by a significant interaction between the bacterial load and the fly line for survival, which reflects the overall rate at which fly health (survival) changes with bacterial load (tolerance) between fly lines. Overall, we found that the Jak/Stat mutants showed differences in disease tolerance phenotypes compared to wild type yw in both males and females (Fig. 4, Table 2). Females lacking duox or Socs36E showed higher tolerance than males to P. entomophila (Fig. 4B and 4A, Table 3). Given the role of duox in producing ROS, one possible explanation for increased tolerance in duox mutants is that ROS-derived damage is lower. However, flies also require intracellular ROS (oxidative burst) such as H2O2 (hydrogen peroxide) for the activation of macrophages during wounding and injury, in addition to Toll and Jak/Stat activation (Chakrabarti and Visweswariah 2020). It is possible that these effects are more pronounced in males which would explain our observation that female duox mutants have comparatively higher tolerance. In other work, wild type (wDahomey) males showed higher levels of duox expression and ROS following Ecc (Erwinia catovora) infection (Regan et al. 2016), which may suggest that loss of function of duox might impact males more than females.
Summary of ANCOVA. To assess differences in infection tolerance (fly survival with increasing bacterial burden) following systemic P. entomophila infection with OD600=0.05 infection dose, around 24 hours following infection. We analysed ANCOVA and fitted ‘sex’ as categorical fixed effects, ‘average bacterial load (ln10)’ as a continuous covariate and their interactions as fixed effects for the Jak/Stat mutants.
Summary of F-test pairwise comparisons of estimates of linear slopes (from the linear model) Jak/Stat-mutants compared to the yw wildtype.

(A). The relationship between fly survival (measured as median lifespan) and bacterial load (as mean CFUs - Colony Forming Units) analysed using linear models for female and male flies (yw & Jak/Stat mutants). Each point shows data for median lifespan and mean CFUs of 15 vials (with each vial containing ∼25 flies/sex/fly line combination after 24 hours post systemic bacterial exposure. The data shown here are for the infection doses (OD600=0.05). (B). Represents estimates of negative slope of the linear reaction norm extracted from the linear models. [Asterix ‘*’ on the lower side of the panel B indicates that Jak/Stat mutants are significantly different from wild type yw, analysed using the F-test pairwise comparisons of estimates of the linear reaction norm for both males and females separately (see Table-3)]. Asterix ‘*’ on the upper side of the panel B indicates sex differences within the fly line i.e., males and females significantly differ in tolerance to systemic bacterial P. entomophila infection.
Regarding the sex difference in disease tolerance in flies lacking Socs36E, it could be due to sex differences in regulation of Jak/Stat, and the resulting differences in immunopathology. For example, female flies lacking the negative epigenetic regulator G9a, were found to be more tolerant than males during viral infections (Gupta and Vale 2017). Similar examples also exist from other innate immune pathways where, disrupting the negative regulator of IMD, PGRP-LB (peptidoglycan receptor-LB), affected survival to a greater extent in females following E. coli infection, suggesting the sex-specific role of some of these regulators (Vincent and Dionne 2021). The combination of these observations might explain why flies lacking Socs36E showed sex differences in disease tolerance, particularly in light of the crosstalk between Jak/Stat and IMD pathways (Kemp et al. 2013; Bang 2019; Dostert et al. 2005). An unexpected result is that we found that female flies lacking domeless showed increased survival (and tolerance) relative to the yw wildtype (Fig. 3 - survival and Fig. 4 - tolerance), suggesting that Jak/Stat activation may in fact be costly to flies. Immune deployment and regulation is highly energy demanding (McKean et al. 2008; Schwenke, Lazzaro, and Wolfner 2016; Nystrand and Dowling 2020). The physiological cost of specific individual immune components and pathways remains an open question for future research.
4. Loss of G9a does not affect bacterial disease tolerance
The epigenetic negative regulator of Jak/Stat G9a was previously identified as being important for tolerating Drosophila C Virus (DCV) infections (Merkling et al. 2015). Subsequent work exploring sex differences in this response found that G9a+/+ (control) females had higher tolerance than G9a−/− females, when measured across a range of viral DCV doses (Gupta and Vale 2017). We wanted to test whether the loss of function of G9a also affects fly survival and disease tolerance in response to bacterial infections. Overall, we found that loss of G9a makes both males and females more susceptible to P. entomophila infections, (Fig. 5A for survival and Fig. 5B for hazard ratio, Table 4 and Table S4). To test if this increased mortality in G9a-/- flies was associated with higher bacterial replication we measured bacterial load following ∼24 hours P. entomophila systemic infection. We found that G9a-/- females exhibited higher bacterial load than G9a+/+ (control) flies, while males showed similar bacterial load as G9a+/+ flies (Fig. 5C, Table S5). However, the overall ability to tolerate P. entomophila bacterial infections (i.e., measured as G9a fly’s survival relative to its bacterial load) remained similar across both males and females G9a flies i.e., both G9a-/- and G9a+/+ controls (see Fig. 5D, Table 5, and Table 6 for comparison between estimates of tolerance slope). Thus, despite the previously identified role of this negative regulator of Jak/Stat in tolerating viral infections by reducing immunopathology (Gupta and Vale 2017; Merkling et al. 2015), G9a does not appear to affect bacterial disease tolerance in either sex.
Summary of estimated hazard ratio from the cox proportional model. A greater hazard ratio (>1) indicates that G9a-/- mutant flies are more susceptible to P. entomophila infection than control (G9a+/+) flies.
Summary of ANCOVA. To assess differences in infection tolerance (fly survival with increasing bacterial burden) following systemic P. entomophila infection with OD600=0.05 infection dose, around ∼24 hours following infection. We analysed ANCOVA and fitted ‘sex’ as categorical fixed effects, ‘average bacterial load (ln10)’ as a continuous covariate and their interactions as fixed effects for each of the fly lines (G9a).
Summary of F-test pairwise comparisons of estimates of the linear slopes (linear reaction norm) for G9a -/-relative to G9a +/+ wildtype fly lines.
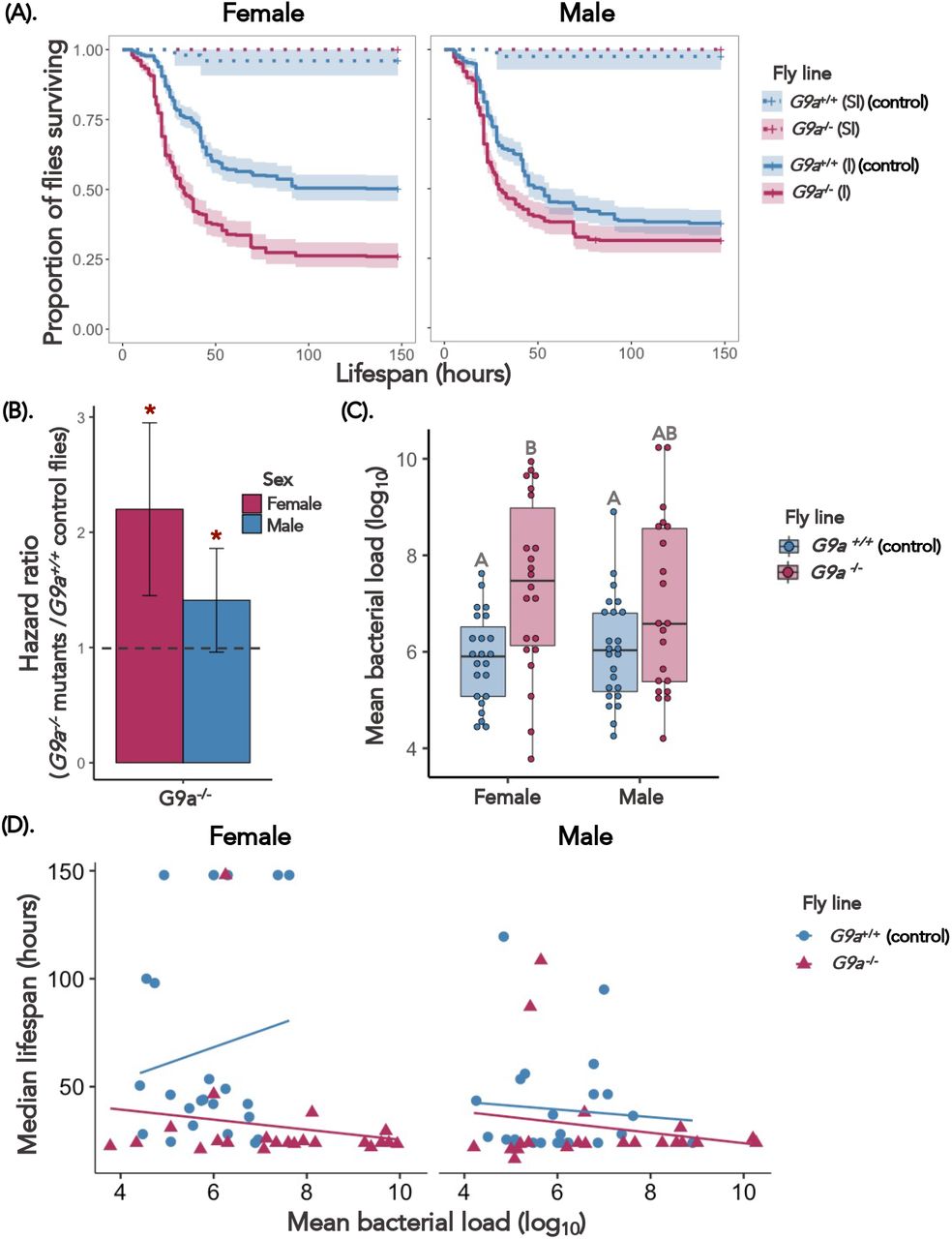
(A) Survival curves for wildtype yw flies and flies lacking G9a the epigenetic regulator of Jak/Stat for female and male flies exposed to systemic P. entomophila of infection dose OD600=0.05 (n=20-30 flies/vial (∼15 vials)/fly line/treatment/sex/infection dose). (B) Estimated hazard ratios calculated from the survival curves for males and female flies (yw and flies without G9a). A greater hazard ratio (>1) indicates higher susceptibility of G9a-/- mutants than control flies to bacterial infection. [‘*’ indicates that the G9a-/- flies are significantly different from G9a+/+ flies]. (C) Bacterial load (mean ln10) measured around 24 hours following infection. [Significantly different fly lines are connected with different alphabets using Tukey’s HSD as a post hoc analysis of pairwise comparisons]. (D) Linear tolerance to P. entomophila infection – the relationship between G9a fly survival (measured as median lifespan) and bacterial load (as mean CFUs - Colony Forming Units) analysed using linear models for female and male flies (both G9a-/- and G9a+/+).
Concluding remarks
Tissue damage signalling and repair mechanisms such as Jak/Stat are important from a therapeutic perspective because they have the potential to boost host tolerance by minimising disease severity (Soares, Gozzelino, and Weis 2014; Vale et al. 2016). Our data show that loss of Jak/Stat pathway components following bacterial infection disrupts disease tolerance and overall survival. These observations have parallels in human infection. For instance, dysregulation of cytokines and interferons (JAK signalling - Tyrosinekinase2) result in immunodeficiency while defective STAT increases the risk of autoimmunity (O’Shea, Holland, and Staudt 2013; O’Shea et al. 2014). Drugs that inhibit JAK have been shown to be effective in treating several autoimmune diseases by targeting cytokine-dependent pathways, while STAT inhibitors have been promising candidates in the context of cancer (Salas et al. 2020; Pérez-Jeldres et al. 2019; Miklossy, Hilliard, and Turkson 2013). It may therefore be possible to repurpose these existing drugs to improve host tolerance of infection. In summary, our work highlights that Jak/Stat signalling mediates disease tolerance during systemic bacterial infection and that this response differs between males and females. Therefore, we suggest that differences in Jak/Stat mediated disease tolerance may be a potential source of sexually dimorphic response to infection in Drosophila.
Authors contribution
A.P and P.F.V conceived the study and designed the experiments; A.P carried out the experimental work with assistance from M.B. and K.M.M; A.P and M.B. acquired the data; A.P analysed the data and drafted the manuscript; A.P & P.F.V wrote the manuscript.
Funding
We acknowledge funding support to P.F.V from the Branco Weiss fellowship and Chancellor’s Fellowship; A.P was supported by the Darwin Trust PhD studentship. We thank School of Biological Sciences, University of Edinburgh. United Kingdom.
Competing interests
None.
Supplementary Information

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
(A) Survival curves for males and females of wildtype yw and Outcross population exposed to systemic P. entomophila of infection dose OD600=0.05 (n=20-30 flies/vial (∼15 vials)/fly line/treatment/sex/infection dose). (B) Estimated hazard ratios calculated from the survival curves for males and female flies (Outcross and yw flies). A greater hazard ratio (>1) indicates higher susceptibility to bacterial infection. [‘n.s’ in the panel indicates that Outcross flies are not significantly different from yw flies]. (C) Internal bacterial load (as mean CFUs-colony forming units) measured around 24 hours post systemic P. entomophila infection. [significantly different fly lines are connected with different alphabets using Tukey’s HSD as a post hoc analysis of pairwise comparisons].
Both wildtype (yw) & Outcrossed flies show similar response to systemic P. entomophila infection
All our Jak/Stat-pathway mutants were in yw genetic background. Previous work has shown that yw flies have lower basal levels of nitric oxide and are more susceptible to infection compared to other lab wild types such as w1118, Oregon R, Canton-S (Eleftherianos et al., 2003). Before addressing whether the Jak/Stat pathway is involved in disease tolerance of bacterial infection, we wanted to better understand how wild type (yw) flies survive P. entomophila infections relative to other wild-type strains. We used an Outcrossed fly population as another control for comparison, which was originally created using 100 pairwise crosses of 113 DRGP lines and maintained as an outcrossed population for over 19 generations [see (Savola et al., 2021) for generation of DGRP Outcrossed population]. We found that males and females of both wild type yw and the Outcrossed flies showed comparable survival following P. entomophila systemic infection and exhibited similar levels of internal bacterial loads when measured ∼24h post-infection (Fig. S1A & S1B, Table S1 for survival; Fig. S1C, for bacterial load). Since both the wildtype (yw) and Outcross fly lines showed similar infection responses to P. entomophila, all remaining experiments only used yw as the wildtype control line.
Summary of mixed effects Cox model, for wildtype yw & flies without Jak/Stat pathway components during systemic P. entomophila infection in. We used data from the individuals of 3-day adult males and females infected with OD600 = 0.05 infection dose of P. entomophila for each fly lines (Jak/Stat -pathway mutants) and specified the model as: survival ∼ treatment x sex x fly line (1|vial), with ‘treatment’, ‘sex’ & ‘fly line’ as fixed effects, and ‘vials’ as a random effect. The table shows model output (ANOVA) for survival post-infection for flies with fully function immune system and lacking Jak/Stat -pathway.
Summary of mixed effects Cox model, for each fly line, wildtype yw & flies without Jak/Stat pathway mutants. We used data from the individuals of 3-day adult males and females infected with OD600 = 0.05 infection dose of P. entomophila for each fly lines (Jak/Stat -pathway mutants) and specified the model as: survival ∼ treatment x sex x fly line (1|vial), with ‘treatment’, ‘sex’ & ‘fly line’ as fixed effects, and ‘vials’ as a random effect. The table shows model output (ANOVA) for survival post-infection for flies with fully function immune system (yw) and flies lacking Jak/Stat – pathway components.
Summary of ln10 transformed bacterial load data around 24 hours following OD600=0.05 P. entomophila systemic infection for wildtype (yw) flies & Jak/Stat pathway mutants, analysed using non-parametric one-way ANOVA (K-W test) by fitting ‘fly line’ (yw and Jak/Stat mutants) as categorical fixed-effects for males and females separately.
Summary of mixed effects Cox model, for G9a Jak/Stat pathway components during systemic P. entomophila infection in. We used data from the individuals of 3-day adult males and females infected with OD600 = 0.05 infection dose of P. entomophila for each fly lines (G9a) and specified the model as: survival ∼ treatment x sex x fly line (1|vial), with ‘treatment’, ‘sex’ & ‘fly line’ as fixed effects, and ‘vials’ as a random effect. The table shows model output (ANOVA) for survival post-infection for flies with fully function immune system and lacking G9a.
Summary of log10 transformed bacterial load data around 24 hours following OD600=0.05 P. entomophila systemic infection for G9a mutants, analysed using non-parametric one-way ANOVA (K-W test) by fitting ‘Fly line’ as categorical fixed-effects for males and females separately.
Acknowledgements
We thank Ronald P. van Rij and David Finnegan for sharing the fly lines, Vincent Doublet and Emily Robertshaw for laboratory help. We are grateful to Darren J. Obbard, Jennifer C. Regan, Luke Holman, Amy B. Pedersen, Daniel H. Nussey, Tiina S. Salminen and the Ashworth fly group members for helpful discussion. Finally, we thank Angela Reid, Alison Fulton, Lucinda Rowe and James King for help with media preparation. Figure-1 was created using BioRender.com.
References




