

Abstract
A common problem experts experience is the challenge of overcoming the ceiling effects of highly trained sensorimotor skills. Recent studies have demonstrated that the amount of practice fails to explain interindividual variation in experts’ motor expertise, implicating hidden bottlenecks that prevent experts from overcoming this ceiling. Here, we show that somatosensory function in motion is one that defines further improvement of the sensorimotor skills of expert pianists. We found that even in expert pianists, specialized training that was designed to enhance somatosensory perception in motion improved sensorimotor skills in which feedback, but not feedforward, control plays a predominant role. Furthermore, training increased reliance on afferent information in motion relative to predictive information that could originate from internal models of movement. Together, these findings suggest that enhancement of somatosensory processing in motion improves the somatosensory-motor feedback loop and thereby enabling experts to surpass the ceiling that initially limited their motor expertise.
Introduction
A ceiling effect, which is defined as a lack of further improvement of a skill following repetitions of practicing in the late stage of training, is a well-known phenomenon in sensorimotor learning. Highly trained individuals, such as athletes or musicians, have suffered from this problem in many situations and even tend to increase the amount of practice in an effort to excel beyond this ceiling. In contrast, a series of recent studies demonstrated that the amount of deliberate practice explains only approximately 20-40% of the interindividual variation in experts’ motor expertise1,2, which begs the question of possible hidden bottlenecks that prevents experts from further improving their expertise. One possible candidate for the bottleneck is insufficient somatosensory function because somatosensory information plays an important role in the sophisticated fine control of movements in terms of correcting and updating motor outputs3–6. We recently found a positive relationship between somatosensory-motor integration functions and the precise force control ability of expert pianists7. Previous studies also demonstrated that individuals who have practiced dexterous movements since childhood (e.g., expert musicians) have superior passive somatosensory functions compared to that of untrained individuals 8,9, and loss of sensory functions degrades fine motor skills 10,11. These findings emphasize the role of somatosensory information in the fine control of movements, which in turn suggests that somatosensory function can be a bottleneck for breaking through the ceiling of fine motor control unless it improves through conventional practice.
We recently developed a specialized training approach, termed “active haptic training (AHT)”, which successfully allowed experts to surpass the ceiling that initially limited their sensorimotor skills12. This training aimed to enhance somatosensory function in motion (i.e., active somatosensory function) and demonstrated enhancement of both active somatosensory function and fine motor control specifically in expert pianists. However, it remains unclear whether the training improved the participants’ active somatosensory function and thereby somatosensory-based feedback-driven control of movements or whether the training improved the feedforward control of participants’ movements and thereby the reduced movement variability collaterally enhanced the active somatosensory perception in motion. The former predicts that the training effect varies depending on the feedback-related gain of movements, which represents the extent that motor execution utilizes sensory feedback information obtained from previous motor actions13,14. Thus, the training effect is likely to emerge only in the sensorimotor skills in which feedback control plays a predominant role. Alternatively, the latter predicts that the training effect on a person’s sensorimotor skills is independent of the feedback-related gain of movements.
An active somatosensory function related to both active perception and feedback motor control is the sensorimotor integration function that involves integrating afferent signals from sensory receptors and signals used to predict sensory consequences of movements based on the internal forward model 15–17. The Bayesian integration model for sensorimotor integration posits that the method of integrating predictive and afferent information depends on the reliability of the information so that the effects of neural noise that disturbs the signals for sensory and sensorimotor processing can be minimized 18. This theory suggests that the modulation of either or both types of sensory information are a putative mechanism underlying the enhancement of motor skills and active somatosensory perception by AHT. However, integrating afferent information with predictive information not only enhances active somatosensory perception but also results in attenuation of sensorimotor processes (i.e., sensorimotor attenuation), by which the sensory consequence of the voluntary action is perceived to be weaker than the same sensory event that was externally generated19,20. Interestingly, aging reduces acuity of afferent information through structural and functional decline in sensory receptors, which increases the relative reliance on predictive information in the integration process and thereby augments sensorimotor attenuation18. In contrast, sensorimotor attenuation is reduced when the incoming sensory signals generated by one’s actions are unpredictable21. Thus, sensorimotor attenuation can be used as an index for assessing changes in the reliability of these two forms of sensory information on integration through AHT. Specifically, attenuation can be reduced or enhanced if AHT increases the reliance on the afferent or predictive information, respectively.
The present study aims to clarify the mechanisms mediating the enhancement of sensorimotor skills and active somatosensory perception in expert pianists through specialized active somatosensory training (i.e., AHT). A series of two experiments demonstrated first that the effects of AHT on sensorimotor skills depended on the feedback-related gain in motor execution, second that AHT increases the reliance on afferent information in somatosensory processing in motion, and third that AHT did not improve passive somatosensory perception. These findings provide the first evidence that optimizing somatosensory processing in motion can improve the somatosensory-motor feedback loop that remains being sophisticated through conventional training even in expert practitioners.
Results
Two experiments were performed to examine the mechanisms underlying the effects of AHT on fine motor skill and active somatosensory perception in well-trained individuals. In total, 102 pianists and 10 musically untrained healthy individuals participated in this study. All pianists majored in piano performance in a musical conservatory and/or had extensive and continuous private piano training under the supervision of a professional pianist/piano professor. To eliminate the possible effects of auditory and visual feedback on the performance of all experimental tasks, we used a muted piano and instructed each participant to perform each behavioral task while listening to white noise presented through headphones worn on the ears and while keeping their eyes closed.
Experiment 1
The purpose of Experiment 1 was to clarify whether AHT improved motor skill via enhancement of feedback-driven control of movements. The impact on motor skill was tested by assessing whether AHT improves motor skill in a manner dependent on or independent of the feedback-related gain of the task performance. To examine this, we designed a series of two experiments. The first experiment identified that the tempo of repetitive keystrokes performed affects the feedback-related gain of that movement. The second experiment tested whether the positive effect of AHT on the target keystroke skill, for which we previously confirmed the ceiling effect in conventional training12, depends on the tempo that varies in relation to the feedback-related gain.
Experiment 1-1
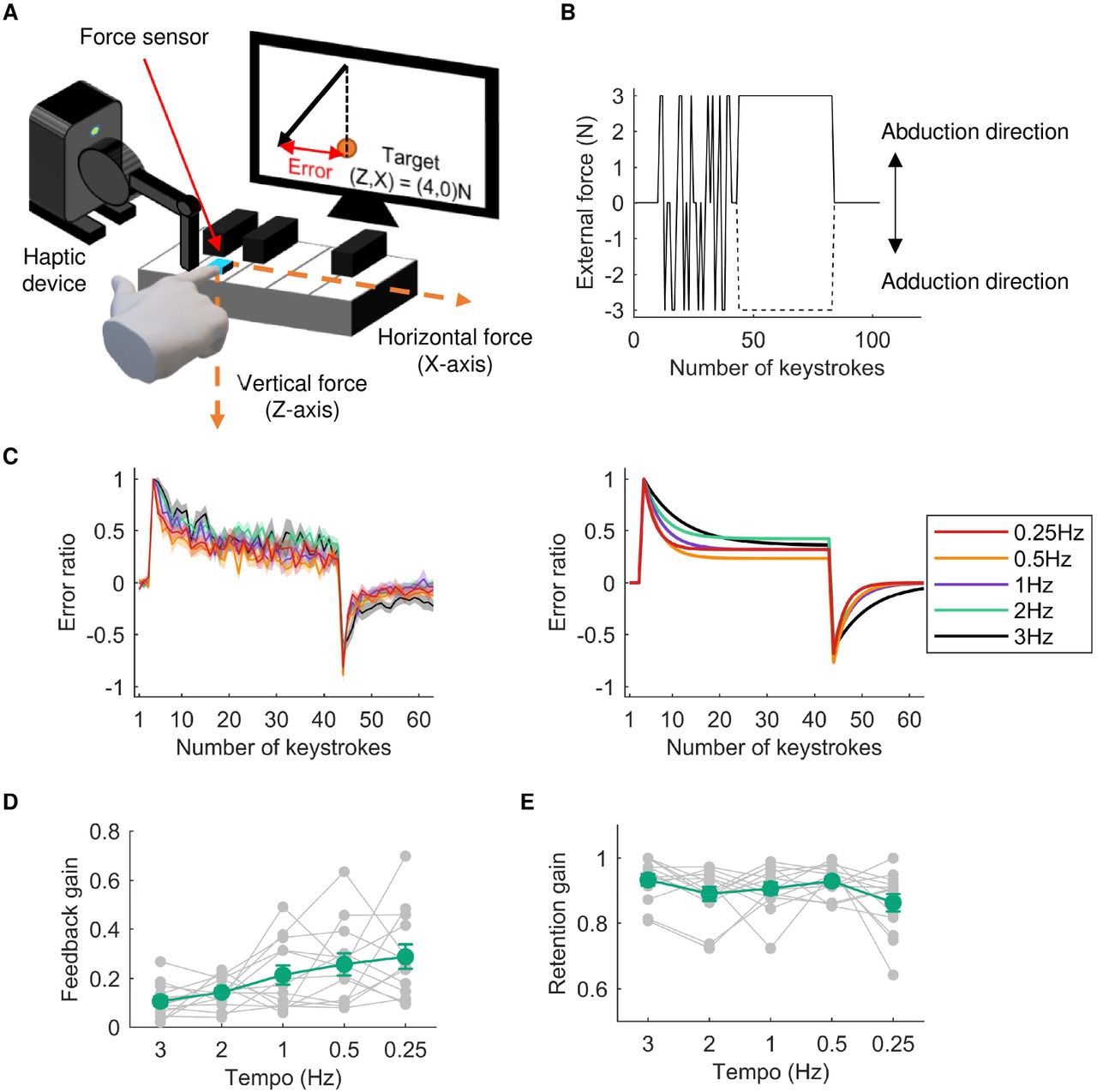
Experiment 1-1 tested whether the feedback-related gain of repetitive keystroke movements depends on the keystroke tempo. We hypothesized that at a faster tempo, the sensory feedback information obtained from a keystroke is more difficult to utilize in the force control of the subsequent keystrokes (i.e., lower feedback-related gain) because the inter-keystroke interval and finger-key contact duration are too short to obtain task-relevant feedback information and to integrate it into motor outputs. To test this theory, 13 expert pianists performed a force-field adaptation task (Figure 1A) using repetitive keystrokes with a single finger at a variety of tempi (3, 2, 1, 0.5, and 0.25 Hz). Each pianist participant performed repetitive piano keystrokes at a certain tempo using the right index finger to which a haptic device was attached. In this task, participants were instructed to strike the piano key so that the fingertip force applied to the key could match a predetermined target force vector. We recorded the 2-dimensional fingertip force applied to the key during each keystroke using a force sensor (USL06-H5-100N. Tech Gihan, Japan) attached to the key surface. We calculated the two-dimensional fingertip force vector (i.e., the X and Z axes, corresponding to the horizontal and vertical directions, respectively) at the moment when the vertical force reached its maximum value during each key depression. The pianists can see the force vector and a target force vector in every keystroke, which are visually displayed on a monitor placed in front of them. We instructed pianists to match the fingertip force vector to a target force vector ((X, Z) = (0N, 4N)) for each keystroke as accurately as possible. While performing this task, the haptic device applied an external force to the finger in the direction of the X axis according to the perturbation schedule shown in Figure 1B (see details in the Methods section). The participants were thus required to compensate for this perturbing force that was applied perpendicular to the keystroke direction.

A: An illustration that explains the force-field adaptation task. The subjects were instructed to strike a piano key with predetermined vertical and horizontal forces at the fingertip of their right index fingers. A haptic device was attached to these fingers, and an external force was applied to these fingers along the horizontal direction during the random and adaptation phases. B: An example of the perturbation schedule during the force-field adaptation task. After the first 10 keystrokes, whether the external force applied to the finger and the direction of external force were determined randomly (i.e., random phase). After that, the external force was removed at 3-5 keystrokes (the number of trials was randomly determined in each trial). During the subsequent 40 keystrokes, the external force was applied to the finger in either the abduction or adduction direction (i.e., adaptation phase). Finally, the external force was removed during the last 20 keystrokes (i.e., washout phase). C: The left panel shows average adaptation curves across subjects in each tempo condition. The right panel shows the average adaptation curves fitted by the single state-space model. Each color line represents each condition, and the shadow areas represent the standard error. D, E: Average feedback-related gain (D) and retention gain (E) across subjects in each tempo condition obtained by model fitting. The green and gray lines represent the mean and individual plots, respectively. Error bars represent the standard error.
The averaged learning curves across the participants in each of the 5 tempo conditions are shown in Figure 1C. We calculated the deviation of the fingertip force from the target force on the X axis as an error index. In the beginning of the adaptation phase, the error was suddenly increased due to the perturbing force produced artificially by the haptic device. The error gradually decreased, although the slope of the decrement of the error appeared to differ among tempi values. To confirm this, we fitted the data by a single state-space model:

where X represents motor outputs at the nth keystroke, e represents error between the target and motor outputs at the nth keystroke, A is the retention gain that represents how much motor outputs at a certain keystroke are utilized in motor outputs at its subsequent keystroke, and B is the feedback-related gain that represents how much motor outputs at a certain keystroke are influenced by the error obtained from the previous keystroke. We found that the feedback-related gain increased as the tempo decreased (Figure 1D; one-way repeated-measures analysis of variance (ANOVA): F2.75,33.05=5.43, p<0.01). In contrast, the retention gain did not differ across tempi (Figure 1E; one-way repeated-measures ANOVA: F4,48=2.23, p=0.08).
Experiment 1-2
In Experiment 1-1, we demonstrated that the feedback-related gain of the target keystroke task depends on the keystroke tempo. The purpose of Experiment 1-2 was to test the effect of AHT on the force variability of repetitive keystrokes performed at different tempi. Thirty-six pianists were categorized into two groups according to whether explicit feedback on performance success (i.e., reinforcement signals) was provided during AHT (i.e., feedback (FB) and no-FB groups). Before and after AHT, the participants performed the active somatosensory test (i.e., weight discrimination threshold (WDT)) and the motor test.
Active haptic training
AHT was designed based on our previous study 12. Participants were instructed to strike a piano key, to which the haptic device was attached, twice in succession at the peak key-descending velocity amounting to 50% of the maximum peak velocity of the key-descending movements (MVK) by the right index finger. During AHT, the haptic device increased the key weight during either the first or second keystroke by pulling up on the key. After performing two successive keystrokes, participants were instructed to answer which keystroke was perceived as heavier. Following each trial, we provided auditory feedback on whether the answer was correct or incorrect (i.e., performance success). The AHT consisted of 20 blocks, each of which comprised 20 trials.
Active somatosensory test
The active somatosensory test required each participant to perform a weight discrimination task that assessed the discrimination threshold in response to changes in the key weight when striking the piano key with the right index finger. Participants were instructed to strike a key at a peak key-descending velocity amounting to 50% of the MVK twice in succession at each trial. During only one of the two strikes, the haptic device increased the key weight to an amount determined by a Bayesian adaptive staircase method (details are included in the Methods). Subsequently, participants indicated which keystroke they perceived as being more difficult. The active somatosensory test consisted of 40 trials, and the WDT was calculated as the mean value of the posterior function obtained through the Bayesian procedure22 across the 40 trials (details are in the Methods section).
Figure 2A and 2B shows the changes in the WDT through AHT in participants in both groups. Two-way mixed measures ANOVA yielded a significant interactive effect between the group and time factors on the WDT (F1,34=4.19, p<0.05). A post hoc test revealed a significant reduction in the WDT through AHT in participants in the FB group (F1,17=15.39, p<0.01) but not in participants in the no-FB group (F1,17=0.32, p=0.58). The WDT obtained during the presession did not differ between participants in the groups (F1,34<0.01, p=0.97), but the WDT in the postsession for participants in the FB group was lower than that of participants in the no-FB group (F1,34=5.89, p=0.02). The results confirmed a specific effect of AHT with the reinforcement signals on the WDT.

A, B: The average weight discrimination threshold across subjects obtained before and after AHT in the FB (A) and noFB (B) groups. Green and brown lines represent the mean values across participants and the gray line represents the individual plots. C, D: The average error index across subjects obtained before and after AHT for participants in the FB group (C) and no-FB (D) groups. E: An example of a time-course of key-movement velocity during the target keystroke task. The upper panel shows the whole time-course of the key-movement velocity in a single trial of the 2 Hz condition. The lower panel shows an enlarged view of the key-movement velocity in a single keystroke. The data show that the duration of the key-descending phase of a single keystroke was less than 100 ms. We calculated the coefficient of variation in the peak key-descending velocity across each keystroke as an error index. **: pre vs. post for participants in the FB group, p<0.01.
Target keystroke task
The participants performed the target keystroke task at each of various tempi before and after AHT. In this task, the participants repetitively struck a piano key 20 times at a rate of 3, 2, 1, 0.5, 0.25 Hz using their right index fingers (Figure 2E). They were instructed to produce 50% of the MVK. During the first 10 keystrokes, the participants received visual feedback information on the peak key-descending velocity value of each keystroke; this feedback was not provided during the subsequent 10 keystrokes. We calculated a coefficient of variation for the peak velocity values across the last 10 keystrokes as an index of the force production consistency. We repeated this task over 5 trials and calculated an average value of this index across the trials.
Figures 2C and 2D show changes in the error index through AHT in participants in both groups at each tempo. Three-way mixed measures ANOVA yielded significant interactive effects between the group and time factors (F1,34=4.69, p=0.04) and between the time and tempo factors (F4,136=2.78, p=0.03) on the error index. This indicated that the difference in the error index between the pre- and postsessions depended on the group and the tempo. Then, we performed two-way mixed ANOVA on the error index obtained from each of the groups. In participants in the FB group, there was a significant interactive effect between the time and tempo factors on the error index (F4,68=2.63, p=0.04). Post hoc tests revealed a significant difference in the error index between the pre- and post-sessions only for the tempi of 0.25 Hz (F1,17=14.85, p<0.01) and 0.5 Hz (F1,17=14.90, p<0.01), showing a smaller value at the postsession. In contrast, in participants in the no-FB group, although we found a significant main effect of tempo (F2.54,43.25=9.74, p<0.01), neither the main effect of the time (F1,17=1.91, p=0.18) nor the interactive effect between time and tempo factors on the error index (F4,68=1.13, p=0.35) was significant. In summary, AHT with the provision of FB information improved the target keystroke skill specifically at slow tempi with high feedback-related gain of movements.
Experiment 2
The purpose of Experiment 2 was to reveal the mechanisms underlying the change in active somatosensory perception through AHT. AHT is designed to assess weight discrimination perception, which is classified as active perception. Such active perception is formed by the integration between afferent information derived from the sensory organs and predictive information generated by internal models represented in the central nervous system. To examine whether and in what manner the way of integrating afferent and/or predictive information was modulated through AHT, we focused on sensorimotor attenuation. The amount of sensorimotor attenuation has been considered to reflect relative reliance on afferent and predictive information at the integration of the two types of information 19. Thus, assessing the amount of sensorimotor attenuation before and after AHT can provide an opportunity to evaluate changes in the integration of the two types of information. First, we developed and tested a novel keystroke force-matching task that aimed to assess the amount of sensorimotor attenuation. Second, we examined changes in sensorimotor attenuation through AHT. Finally, we examined changes in passive somatosensory perception through AHT to confirm that the enhancement of active somatosensory perception is mediated by modulation of somatosensory functions related only to active perception, rather than one related to overall somatosensory perception.
Keystroke force-matching task
A robust quantitative measure of sensorimotor attenuation can be obtained by the force matching task. In that task, a constant force was applied to a subjects’ finger by a torque motor; subjects were then instructed to reproduce the force they had just felt by pressing with a finger on the other hand23. We customized this task so that it was suitable for piano keystroke movements. In this task, participants were asked to produce target keystroke movements that were passively generated by a haptic device prior to voluntarily performing the keystrokes as accurately as possible (active condition). The haptic device applied downward force to the right index finger of the participants with a predetermined force level at a tempo of 2 Hz. The force was applied to each participant’s finger 10 times successively in each trial, and each applied force lasted 200 ms. After the passive movements (i.e., the target keystroke), the haptic device was automatically detached from the participant’s finger. Then, each participant was instructed to reproduce the target keystroke generated by the haptic device as accurately as possible (i.e., the reproduction keystroke). In a control condition that examines whether the sensory attenuation effect is not simply due to a failure of memory, participants reproduced target keystroke movements indirectly by controlling the force output of the haptic device attached to their fingers. The participants can control the force level of the keystrokes generated by the haptic device attached to their right index fingers by pressing several predetermined piano keys using their left hands. Each key corresponds to each of the following functions: ‘increase force function’, ‘decrease force function’, ‘reproduction function’, and ‘target function’. ‘Target function’ triggers passive keystrokes (i.e., target keystroke) with one of the predetermined force levels (1.5 N, 2.4 N, and 2.8 N). Participants can manipulate the force level in steps of 0.1 N by selecting the ‘increase/decrease force function’ for the passive keystrokes triggered by the ‘reproduce function’ (i.e., keystroke reproduction). We instructed each participant to produce keystrokes with the velocity that matched the peak key-descending velocity of the passively generated keystrokes triggered by pressing the ‘target function’ and pressing the ‘reproduction function’ as accurately as possible.
We calculated the ratio of a difference in the peak key-descending velocity between the target keystroke and reproduced keystroke (i.e., (reproduction–target)/target) as an index of the amount of sensory attenuation. The positive and negative values indicated stronger and weaker keystrokes for the reproduced keystroke than target keystroke, respectively.
Experiment 2-1
The purpose of this experiment was to clarify whether the keystroke force-matching task can successfully assess sensorimotor attenuation and whether the amount of sensorimotor attenuation differs between expert pianists and musically untrained individuals (nonmusicians). We hypothesized that pianists would show greater sensorimotor attenuation than nonmusicians because the reliance on predictive information derived from an internal model of keystroke movements in active somatosensory processing can be larger in pianists than in nonmusicians. Ten pianists and 10 nonmusicians participated in this study. They performed the keystroke-matching task under both active and control conditions. In both conditions, the haptic device passively moved the participants’ fingers by applying 4 different force levels (1.5 N, 2.1 N, 2.4 N, and 2.8 N) during the target keystrokes.
Figure 3A shows the results of the keystroke force-matching task for participants in both groups. In the active condition, the pianists showed higher key-descending velocity of the reproduction strokes than that of the target keystrokes specifically in the low force levels (1.5 N and 2.1 N). This overcompensation is considered to result from the sensorimotor attenuation and reflect the integration of the afferent and predictive information19. In contrast, such overcompensation of the key-descending velocity in the reproduced keystrokes was not evident in nonmusicians. Two-way mixed ANOVA yielded significant main effects of group (F1,18=6.20, p=0.02) and force level (F2.33,41.98=20.96, p<0.01) on sensorimotor attenuation (Figure 3B). In contrast, in the control condition, the ratio was near 0 at all force levels, and two-way mixed ANOVA detected no significant difference in the change ratio between participants in the groups and between the force levels (main effect of group: F1,18=1.75, p=0.22; main effect of force level: F1.73,31.12=0.91, p=0.40; interaction effect: F1.73,31.12=0.84, p=0.42).
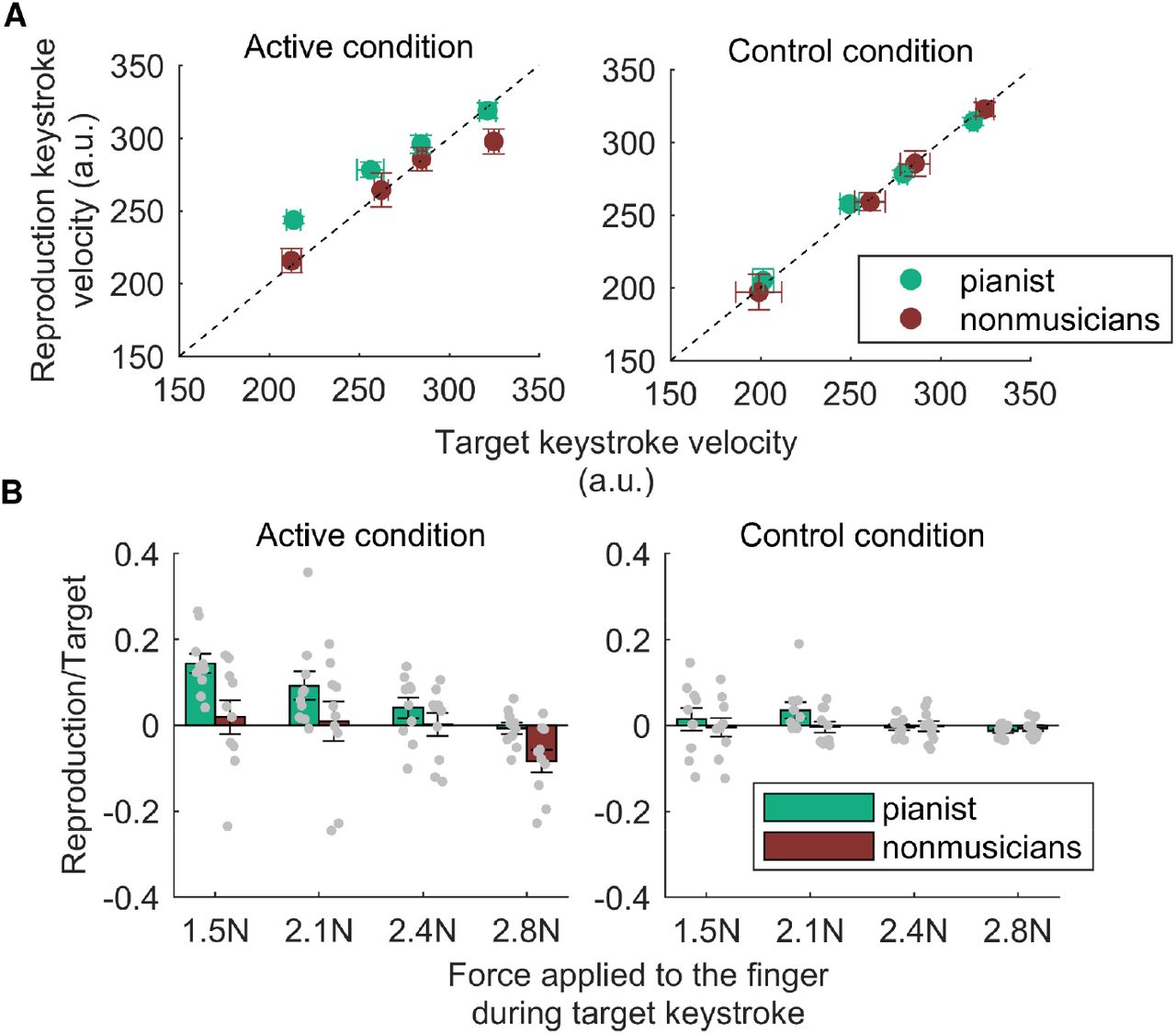
A: Key-descending velocity generated by either directly striking a piano key (i.e., active condition, left panel) or indirectly striking a piano key using the haptic device (i.e., control condition, right panel) as a function of the externally generated target key-descending velocity. Green and brown plots represent the results obtained from the pianists and nonmusicians, respectively. B: The average values of velocity overcompensation across subjects in the active (left panel) and control (right panel) conditions. A positive value indicates sensorimotor attenuation. Gray plots represent the individual data. Error bars represent the standard error.
Experiment 2-2
The result of Experiment 2-1 demonstrated that the keystroke force-matching task can successfully identify the sensorimotor attenuation in expert pianists when the haptic device moved the participant’s finger with a low level of force during the target keystrokes. Experiments 1-2 were designed to examine the effect of AHT on sensorimotor attenuation. Thirty pianists performed the keystroke force-matching task with a low level of force (1.5 N) and an active somatosensory test before and after performing AHT.
Experimental group
The pianists were categorized into two groups according to whether explicit feedback on performance success (i.e., reinforcement signals) was provided during AHT (FB and no-FB groups). Because our previous study demonstrated no enhancement of active somatosensory perception without providing reinforcement signals during AHT 12, a comparison of participants in these two groups can assess changes in dependent variables through improvement in active somatosensory perception.
Active somatosensory test
Figure 4A shows the changes in the WDT due to AHT in the pianists and nonmusicians. Two-way mixed ANOVA detected a significant interactive effect between the group and time factors on the WDT (F1,28=5.18, p=0.03). Post hoc tests revealed a significant reduction in the WDT following AHT for participants in the FB group (F1,14=19.66, p<0.01) but not for participants in the no-FB group (F1,14=2.32, p=0.15). Furthermore, the WDT at the postsession differed significantly between participants in the two groups (F1,28=7.86, p<0.01), unlike the WDT at the presession (F1,28=0.30, p=0.59).
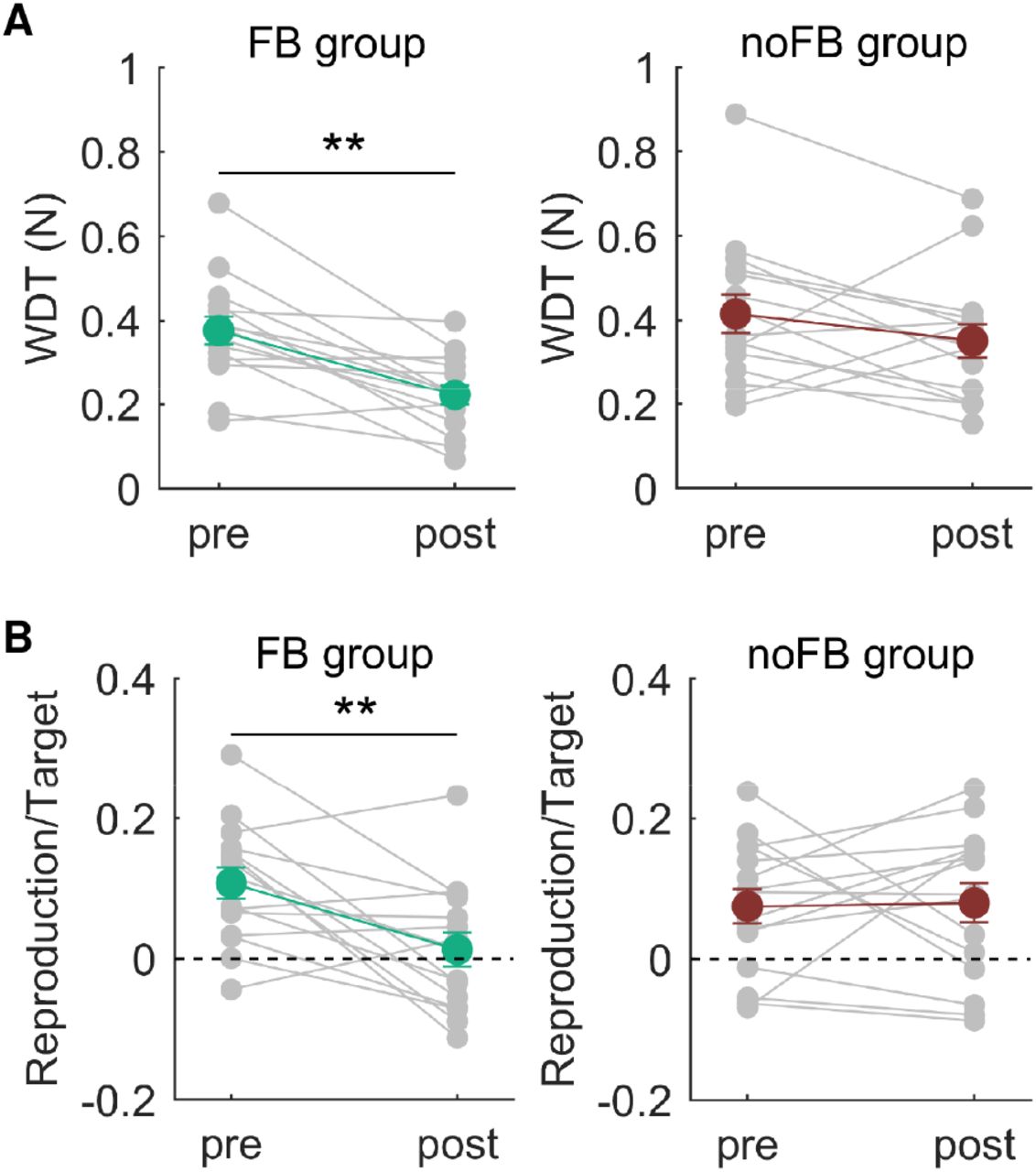
A: The average weight discrimination threshold across subjects obtained before and after AHT. B: The average values of velocity overcompensation across subjects in the active condition obtained before and after AHT. Error bars represent the standard error. **: pre vs. post, p<0.01.
Keystroke force-matching task
Figure 4B shows the change in the amount of sensorimotor attenuation through AHT in participants in both groups. Two-way mixed ANOVA yielded a significant interactive effect between the group and time factors on the amount of sensorimotor attenuation (F1,28=5.55, p=0.03). Post hoc tests revealed a significant reduction in the amount of sensorimotor attenuation through AHT in participants in the FB group (F1,14=10.92, p<0.01) but not in participants in the no-FB group (F1,14=0.02, p=0.88). Because the amount of sensorimotor attenuation at the presession did not differ between the groups (F1,28=0.97, p=0.33), these results suggest an increase in the relative reliance on the sensory afferent information to the predictive information through AHT.
Experiment 2-3
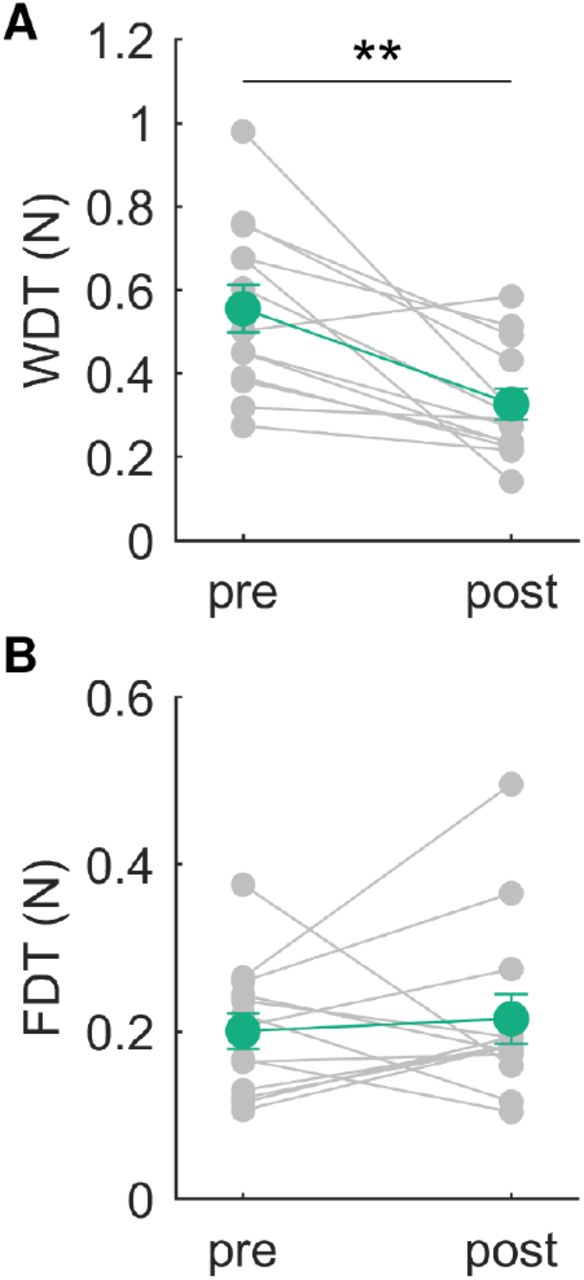
The results of experiments 2-1 and 2-2 demonstrated that the amount of sensorimotor attenuation was decreased through AHT along with the reduction in the WDT. This possibly resulted from changes in somatosensory functions related to active perception, rather than one responsible for the overall somatosensory perception. To confirm this, we assessed passive somatosensory perception before and after AHT. Thirteen expert pianists participated in this experiment and performed AHT with the provision of feedback information during the training session (i.e., the same participants in the experimental group as in the FB group in Experiment 2-2). Before and after AHT, participants performed both active and passive somatosensory tests. Specifically, for the active somatosensory test, we found a reduction in the WDT through AHT (Figure 5A, presession: 0.56±0.21 N; postsession: 0.33±0.14 N (mean±SD); paired t test: t=4.46, p<0.01).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
A B: The average weight (A) and force (B) discrimination threshold across subjects obtained before and after AHT. Error bars represent the standard error. **: pre vs. post, p<0.01.
Passive somatosensory test
The passive somatosensory test required each participant to perform a keystroke-force discrimination task that assessed the discrimination threshold with respect to changes in the force of the keystroke generated by the haptic device attached to the participant’s right index finger. The haptic device applied a downward force on the finger to strike a key twice in succession in each trial. The force differed between the two passive keystrokes. Subsequently, participants indicated which passive keystroke they perceived to be stronger. The passive somatosensory test consisted of 40 trials, and the force discrimination threshold (FDT) was calculated by a mean value of the posterior function obtained by the Bayesian adaptive procedure across the 40 trials (see details in the Methods section).
We found no significant difference in the FDT between the pre- and postsession (Figure 5B, presession: 0.20±0.08 N; postsession: 0.22±0.11 N (mean±SD); paired t test: t=-0.36, p=0.73), which confirmed a lack of any change in passive force perception through AHT.
Discussion
The present study examined the mechanisms mediating the enhancement of sensorimotor skills and active somatosensory perception in expert pianists through specialized active somatosensory training (i.e., AHT). In Experiment 1, we tested the hypothesis that AHT improves motor skills through the sophistication of sensorimotor feedback control. First, we revealed that the feedback-related gain of the repetitive keystrokes depended on movement tempo. Then, we found that the effect of AHT on the target keystroke skill was evident only when the task was performed at a slow tempo, in which the feedback-related gain of movements was high, which indicated that AHT improves feedback control of the keystroke movements in expert pianists. In Experiment 2, we focused on sensorimotor attenuation as an index of the relative reliance between afferent and predictive information on active somatosensory perception. First, we found stronger sensorimotor attenuation in the pianists than in musically untrained individuals, which indicated a larger integration of the predictive information into somatosensory perception during keystrokes in participants in the former group. Second, we found that the amount of sensorimotor attenuation decreased through AHT only in the pianists, which suggested an increase in the relative reliance on the afferent information relative to the predictive information through AHT. Third, in the pianists, passive and active somatosensory perception were unchanged and improved through AHT, respectively, which means that AHT influences somatosensory functions responsible for active perception but not for passive perception. Taken together, AHT improves feedback control of the keystroke movements in expert pianists via the increased reliance on afferent information from the somatosensory organs. The improvement of active somatosensory functions can thus play a key role in overcoming the ceiling effects of experts’ motor expertise.
Sensorimotor skill
A common problem that experts face is how to overcome the ceiling effects on the sensorimotor skills for which they are highly trained. A possible reason why this problem has not yet been solved is a lack of understanding of the bottlenecks that hinder experts’ ability to improve their motor expertise, despite extensive arguments on the roles of genetic predisposition and deliberate practicing in the proficiency of these individuals 24. A previous study demonstrated a relationship between somatosensory-motor integration functions and keystroke skills in expert pianists7. In addition, the somatosensory-motor integration functions in healthy expert pianists differ from those of nonmusicians and patients with focal hand dystonia 25–27. These findings suggest that somatosensory-motor integration functions play a critical role in sensorimotor control and that those functions can be the bottleneck hindering further enhancement of fine motor control in experts if those functions require further sophistication. In agreement with this idea, we recently demonstrated that AHT targeting the somatosensory-motor integration function improved the target keystroke skill of expert pianists beyond the limit of performance improvement that can be achieved by conventional training (i.e., a mere repetition of task performance)12. AHT involves repeatedly discriminating unexpected components (i.e., increment of the weight of the piano key) based on the somatosensory information derived from one’s own movements. Because movement control and learning rely heavily on corrective mechanisms based on errors between perceived and expected motor actions 28,29, it is possible that AHT enhanced the somatosensory-based error-detection function and thereby improved the feedback control component of the target keystroke skill. This predicts that AHT improved the skill only when feedback control plays a dominant role in movement control. The present keystroke skill is characterized as repetitions of a single rapid keystroke movement, which suggests that both feedforward and feedback control processes are likely to contribute to the performance of this skill. Specifically, each single key depression is mainly controlled by the feedforward process because the duration of the movement is shorter than the sensory feedback delay (i.e., less than 100 ms) (see Figure 4A). In contrast, the stroke-by-stroke feedback control process may operate by integrating sensory information obtained from a keystroke to produce motor commands for the subsequent keystrokes. The present study tested which process improved through AHT. First, we found that the feedback-related gain was larger at slower keystroke tempi with longer duration to integrate the feedback somatosensory information obtained from a keystroke into motor outputs of the subsequent keystroke (i.e., sensorimotor feedback loop). Then, we demonstrated that AHT improved the target keystroke skill performed only at slow tempi. These results indicated that AHT improves the sensorimotor feedback control of repetitive keystroke movements and proposed that active somatosensory function, specifically for the error-detection function in motion, is one candidate bottleneck for expert pianists in overcoming the ceiling effect of fine motor control.
Active somatosensory perception
We replicated the previous finding that the WDT in expert pianists decreased through AHT12. The weight discrimination task used in this study requires perceiving the change in the weight of the key through the keypress according to the somatosensory information derived from the keystroke movement. While a number of studies reported that passive somatosensory perception was enhanced through perceptual and motor learning and involved neuroplastic changes in the somatosensory cortex30–34, it remained almost unknown whether and in what manner active somatosensory perception is improved through training. Active perception is formed by integrating the sensory information encoded in the afferent signals originating from the peripheral sensory receptors and the predictive sensory information based on the internal forward models represented in the central nervous system15–17. This predicts that changes in at least one of these two pieces of information underlie the improvement of the perceptual discrimination threshold in motion through AHT. We examined this prediction by probing sensorimotor attenuation, in which the sensory consequences of the voluntary action are perceived to be weaker than the same sensory event when it is externally generated19,20. The integration of predictive information with afferent information thus induces the attenuation of perceived sensory events. In Experiment 1-1, we found larger sensorimotor attenuation during keystrokes in expert pianists than in nonmusicians, which means a larger reliance on sensory prediction for active somatosensory perception in the former individuals. Previous studies have demonstrated that sensorimotor attenuation is reduced when incoming afferent signals are unpredictable or temporally modulated 21,35. Thus, in nonmusicians, a lack of accurate representation of novel mechanical characteristics of the piano may prevent sensory consequences of keystroke movements from being predicted. In addition, previous studies demonstrated modulation of sensorimotor attenuation in elderly individuals18 and patients with schizophrenia36. The present study extends these findings by providing the first evidence of expertise-dependent changes in sensorimotor attenuation. Experiments 1-2 further demonstrated a reduction in sensorimotor attenuation through AHT in pianists. This indicates an increase in the relative reliance on afferent sensory information for active somatosensory perception through AHT. The training effect can be mediated by functional changes in the peripheral system, such as enhancement of the sensitivity of fingertip somatosensory receptors. However, this is unlikely because passive somatosensory perception was unchanged through AHT, in contrast to the enhancement of active somatosensory perception. Another possible reason for this outcome is a change in the motor strategy that influences incoming sensory afferent signals. For example, pianists might learn to hit the key with the most sensitive skin area in the fingertip and/or the finger posture with the highest proprioceptive sensitivity through AHT. This enables pianists to obtain augmented afferent signals during keystrokes. Thus, we postulate that the pianists learned an optimal movement strategy that provided fine-tuned somatosensory information on the key weight during the keystrokes through AHT.
We also replicated our previous result that both WDT and target keystroke skill improved by AHT only when the pianists received reinforcement signals during the training12. A theory of reinforcement learning posits that learners perform tasks by trial and error with a learning policy to maximize the amount of reward obtained. According to this theory, pianists might learn to perform the weight discrimination task by exploring an optimal movement strategy that provides augmented task-relevant somatosensory afferent signals during AHT. This enhances the somatosensory afferent processes in motion, which in turn improves active somatosensory perception and feedback-driven control of movements.
Limitation
There are at least two limitations in this study. One is that the same subject performed the force-field adaptation task multiple times (i.e., 5 tempo conditions × 2 times = 10 times), which may have elicited a saving effect. Saving is the ability of past learning to enhance subsequent learning14,37–39. Thus, it is possible that the adaptation rate might differ between data measured at the beginning and at the end of the experiment. To minimize this possibility, we randomized the order of tempo conditions across participants. We also included a random phase in which the external force was applied randomly in the direction of abduction or adduction. Previous studies demonstrated that exposure to two opposite perturbations during movements inhibits adaptation learning of both types of perturbations, which interferes with the formation of motor memories40,41. The random phase was always followed by the adaptation phase to prevent the saving effect. Furthermore, to prevent prediction of the direction of external force during the adaptation phase, external force was applied once in the in the abduction direction and once in the adduction direction at each tempo in a random order. These schemes might minimize the suppression of the potential effects of saving on the results of the adaptation task.
Another limitation is that while AHT improved the target keystroke skill performed at a tempo of 2 Hz in the previous study12, the same training did not improve the skills of participants in the present study. A contrasting difference is that participants performed the target keystroke task using the right ring and index finger in the previous and present study, respectively. The ring finger has innately lower finger independence and controllability of movements compared with those of the index finger42,43, which is confirmed by a superior performance index of the target keystroke skill in the present study over those observed in the previous study. This suggests a potential for improvement of feedforward motor control specifically in the ring finger. Thus, it is possible that, in the previous study, AHT improved both the feedforward and feedback motor control of the right ring finger in expert pianists, but AHT only improved the feedback motor control of the right index finger of participants in the present study.
Conclusion
The present study tested the hypothesis that somatosensory function is a bottleneck for overcoming the ceiling of experts’ motor proficiency by investigating the mechanisms underlying the effects of AHT on the active somatosensory perception and fine motor control of piano keystrokes in expert pianists. A series of two experiments provided evidence that AHT augmented reliance on the somatosensory afferent information derived from one’s own movements in the somatosensory process in motion, which improves the active somatosensory perception and feedback sensorimotor control in expert pianists. These results indicate that active somatosensory functions remain unoptimized through conventional training even in experts who underwent years of extensive piano training and are therefore a possible bottleneck for overcoming the ceiling effect of expert motor proficiency. We thus conclude that enhancing active somatosensory functions by specialized sensorimotor training can improve the functions of the somatosensory-motor feedback loop, which further enhances experts’ motor expertise.
Materials and methods
Participants
In total, 102 healthy pianists and 10 nonmusicians participated in this study. All of the pianists majored in piano performance at a musical conservatory and/or had extensive and continuous private piano training under the supervision of professional pianists and/or piano professors. All participants gave their written informed consent before engaging in the experiments. All experimental procedures were carried out in accordance with the Declaration of Helsinki and were approved by the ethics committees of the Sony Corporation. To remove the effects of auditory feedback on the performance of all experimental tasks, we muted a piano and instructed each participant to perform each behavioral task while listening to white noise emanating from headphones worn on their ears.
Recording of piano key movements
The vertical motion of the piano key was measured using an optical distance sensor placed under the key at a sampling rate of 500 Hz. The sensor values were low-pass filtered (fifth-order zero-lag Butterworth filter, cutoff frequency: 24 Hz) and differentiated to obtain the velocity values.
Active haptic training
AHT was designed based on our previous study 12. Participants were instructed to strike a piano key, to which a haptic device (Geomagic Touch X, 3D Systems) was attached, twice in succession at a peak key-descending velocity amounting to 50% of the MVK by the right index finger. A warning message of “too weak” or “too strong” was displayed on a monitor placed in front of the participants when the peak key-descending velocity was outside 35-65% of the MVK. During AHT, the haptic device increased the key weight during either the first or second keystroke by pulling up on the key. After performing the two keystrokes, participants were instructed to indicate which keystroke was perceived as heavier. Following each trial, we provided visual feedback on whether the answer was correct or incorrect (i.e., performance success). The AHT consisted of 20 blocks, each of which had 20 trials. In the first block, the haptic device loaded the key to the amount predetermined by the somatosensory test performed prior to the training. To maintain task difficulty and facilitate perceptual learning during the training session, the load for the next block was reduced 10% from the load for the previous block when participants correctly answered more than 80% trials within a block.
Active somatosensory test
The active somatosensory test involved a weight discrimination task that assessed the WDT as to changes in the key weight when striking the piano key with the right index finger. Participants were instructed to strike a key at a peak key-descending velocity amounting to 50% of the MVK twice in succession at each trial. During only one of the two strikes, the haptic device increased the key weight. Subsequently, participants answered which keystroke they perceived as being more difficult. The active somatosensory test consisted of 40 trials, and the WDT was calculated by a Bayesian procedure (i.e., ZEST)22 through the 40 trials. Briefly, ZEST is an efficient method of measuring thresholds of discrimination perception. We first specified prior knowledge, namely, a guess (=0.4N, -0.92 in a logarithmic (log) scale) and an associated standard deviation (=7.39N, 2 in a log scale) for the probability of threshold values. These values were used to calculate the initial probability density function (pdf) of the threshold by assuming it to be a Gaussian function. The pdf was updated every trial based on the result of the discrimination test. The stimulus intensity and the final estimate of the threshold were chosen to correspond to the mean of the latest pdf. Because the ZEST worked based on a log scale, log-scale values were used for statistical testing. By contrast, for visualization, we adopted linear scale values to increase the understandability.
Experiment 1
Experiment 1 was designed to clarify the mechanisms underlying the effect of AHT on the participants’ target keystroke skills. We conducted two experiments to examine the hypothesis that AHT improves feedback motor control.
Experiment 1-1
The purpose of this experiment was to examine whether the feedback-related gain of repetitive keystroke movements depended on the keystroke tempo. Thirteen pianists performed the force-field adaptation task.
Force-field adaptation task
The participants performed repetitive strikes of a piano key at one of the five tempi (3, 2, 1, 0.5, and 0.25 Hz) using the right index finger to which the haptic device was attached. In this task, participants were instructed to strike the piano key so that a fingertip force applied to the key could match a predetermined target force vector. We recorded the 2-dimensional fingertip force applied to the key during each keystroke using a force sensor (USL06-H5-100N. Tech Gihan, Japan) attached to the key. We calculated the two-dimensional fingertip force vector (i.e., the X and Z axes, each of which corresponds to the finger abduction/adduction and downward directions, respectively) at the moment when the Z axis force reached a maximum value during each keystroke. The pianists can see the force vector and a target force vector for every keystroke, which were displayed on a PC monitor located in front of them. We instructed pianists to match the fingertip force vector to a target force vector ((X, Z) = (0N, 4N)) as accurately as possible for each keystroke. During the first 10 keystrokes, the haptic device did not apply any force to the participants’ fingers. After these 10 keystrokes, the haptic device applied an artificial force to the participants’ right index fingers in the direction of abduction or adduction (i.e., X axis). The artificial force direction and whether the artificial force was applied were randomly determined across the keystrokes (random phase), and the random phase was continued over 40 keystrokes. The random phase was used to prevent savings that are defined as faster relearning under a known environment. At 3 to 5 keystrokes after the random phase (the number of keystrokes was determined randomly), the artificial force was removed. Then, during the subsequent 40 keystrokes, the haptic device applied the artificial force to the finger in the direction of the X axis (adaptation phase). The artificial force direction was randomly chosen but was consistent in the adaptation phase. Because each pianist performed this task twice per tempo condition, the abduction and adduction directions of the artificial force during the adaptation phase were chosen once each per tempo condition. Then, the artificial force was removed (washout phase), and the washout phase continued over 20 keystrokes.
Experiment 1-2
The purpose of this experiment was to test whether the effect of AHT on the target keystroke skill depended on the feedback-related gain. Thirty pianists participated in this experiment.
Target keystroke task
The participants performed the target keystroke task with different tempi before and after AHT. In this task, the participants repetitively struck a piano key 20 times at a rate of 3, 2, 1, 0.5, 0.25 Hz using their right index fingers. They were instructed to produce 50% of the MVK. During the first 10 keystrokes, the participants received feedback information on the peak key-descending velocity value of each keystroke; this information was not provided during the subsequent 10 keystrokes. We calculated a coefficient of variation for the peak velocity values across the last 10 keystrokes as an index of the force production consistency. We repeated this task over 5 trials and calculated an average value of this index across the trials.
Protocol
Participants were categorized into either the FB group or the no-FB group. Before and after performing AHT, participants performed the target keystroke task at 5 different tempi (0.25, 0.5, 1, 2, and 3 Hz).
Experiment 2-1
The purpose of this experiment was to examine sensorimotor attenuation during keystroke movements between pianists and nonmusicians. Ten pianists and 10 nonmusicians performed the keystroke-matching task.
Keystroke force-matching task
As a robust quantitative measure of sensorimotor attenuation, the force-matching task has been used in previous studies, in which participants were asked to produce the force that matched the target force produced by a torque motor 23. We customized this task for piano keystrokes. In this task, participants were asked to reproduce a target keystroke movement that was passively generated by the haptic device. The haptic device applied downward force to the participants’ right index fingers with a predetermined force level at a tempo of 2 Hz. The force was applied to the finger 10 times in each trial, and each applied force lasted 200 ms. Following passive movements, the haptic device was automatically detached from the finger, and the participants were instructed to reproduce the target keystroke generated by the haptic device as accurately as possible (active condition). In a control condition that evaluates haptic pressure perception, participants controlled keystroke movements indirectly through the haptic device at the reproduction session. The participants can manipulate the force level of the haptic device via piano keys using their left hands. Each of the 5 keys corresponded to each of the following functions: ‘increase force function’, ‘decrease force function’, ‘reproduction function’, ‘target function’, and ‘finish function’. Participants can manipulate the force level in step of 0.1 N by selecting the ‘increase/decrease force function’ for the passive keystrokes triggered by the ‘reproduce function’. In each trial, participants can confirm the target keystrokes and reproduction keystrokes any number of times by pressing the keys corresponding to ‘target function’ and ‘reproduction function’, respectively. During the control condition, participants can see the PC monitor where the correspondence table between the keys and functions is displayed. We instructed each participant to close his or her eyes when pressing the keys corresponding to the target and reproduction functions and to match the peak key-descending velocities of the target and reproduction keystrokes. Once they perceived that the two types of keystrokes were the same, they were asked to press the ‘finish function’ to proceed with the task.
We calculated the ratio of a difference in the peak key-descending velocity between the target keystroke and reproduced keystroke ((reproduction–target)/target) as an index of the amount of sensory attenuation. The positive and negative values indicate stronger and weaker keystrokes in the reproduced keystroke than in the target keystroke, respectively.
Experiment 2-2
The purpose of this experiment was to examine the effect of AHT on sensorimotor attenuation. Thirty pianists participated in this study.
Protocol
The pianists were categorized into two groups. One group of participants can receive feedback information about whether their answers were correct during AHT (FB group). The pianists in the other group (no-FB group) cannot receive such feedback information during AHT. Before and after AHT, the participants performed the active somatosensory test and sensorimotor attenuation.
Experiment 2-3
This experiment examined whether AHT modulates passive somatosensory functions. Thirteen pianists performed a passive somatosensory test before and after AHT.
Passive somatosensory test
The passive somatosensory test involved a keystroke strength discrimination task that assessed the discrimination threshold regarding changes in the strength of keystrokes generated by the haptic device attached to the participants’ right index fingers. The haptic device applied a downward force to the finger to strike a key twice in succession at each trial. The strength differs between the two passive keystrokes. Subsequently, participants indicated which passive keystroke they perceived as being more difficult. The passive somatosensory test consists of 40 trials, and the FDT was calculated by the ZEST.
Acknowledgments
The present study was supported by JSPS Grant-in-Aid for Transformative Research Areas B (20H04093) for M.H, JST CREST (JPMJCR17A3) and JSPS Grant-in-Aid for Transformative Research Areas B (20H05713) for S.F.
References




