

Abstract
Tendons transmit mechanical forces between muscle and bone, and their biomechanical function requires high tensile strength, which is provided by highly organized collagen fibers. Tenocytes mainly drive tendon growth via extracellular matrix (ECM) production and organization. The biological mechanisms regulating tenocyte differentiation and morphological maturation have not been well established, partly due to the lack of a reliable in vitro system. Recent scaffold-free, cell-based tissue engineering approaches developed several unique in vitro tendon-like constructs. However, the application of these constructs is limited to the study of embryonic tendon development due to high cell density and immature matrix organization. In this study, we developed a scaffold-free, three-dimensional (3D) tendon culture system using mouse tendon cells and a differentially adherent growth channel. The 3D tendon constructs exhibited tissue maturation similar to the postnatal mouse tendon, including decreased cell density, increased thickness, and elongated cells between highly aligned extracellular matrix. The 3D tendon culture system is also feasible for genetic manipulation using adenovirus. Overall, the results suggest that the 3D tendon culture system using mouse tendon cells is a reliable in vitro system to study underlying biological mechanisms that regulate cellular and matrix maturation in postnatal tendon development.
Introduction
Tendons play a critical role in the musculoskeletal system by transmitting mechanical forces between muscle and bone. Its function requires strong tensile strength provided by a highly organized collagen fiber structure (1). Tendon collagen fibers consist of multiple collagen fibrils primarily composed of type I collagen (2). The tensile strength of each tendon is unique, depending on the type (positional, energy-storing; flexor/extensor) and location. Despite their notable tensile strength, tendons are prone to injury. Tendon injuries present a challenging clinical problem because of the slow healing process and the inability to restore original structural stability and mechanical integrity (3–6). Therefore, tendons are attractive targets for tissue engineering and regenerative medicine. However, the biological and mechanical mechanisms regulating cellular and matrix maturation remain unclear, which stands as an obstacle to advancing tendon tissue engineering and regenerative medicine.
Tendon maturation acquires dramatic cellular and matrix changes (7, 8). Tenocytes are primary tendon fibroblasts that drive tendon growth via extracellular matrix (ECM) production and organization (9–11). Tenocyte differentiation is a multistep process that requires specific gene expression and unique morphological changes. Tendon progenitors are marked by the expression of Scleraxis (Scx), a basic helix-loop-helix (bHLH) transcription factor that is critical for tenocyte differentiation (12, 13). Differentiated, active tenocytes highly express type I collagen and tenomodulin (14–16). Besides molecular changes, tenocytes undergo unique morphological maturation. The active tenocytes are relatively big, with a rounder spindle-shape (8). However, fully matured tenocytes become very flat and elongated, which enables tenocytes to align longitudinally between dense and highly organized collagen fibers (8). Mature tendon cells cease proliferation and produce extracellular matrix, which results in matrix maturation and lower cell density in the postnatal tendon (7). Despite the critical role of tenocytes in tendon maturation, the biological mechanisms regulating molecular and morphological changes in tenogenic maturation are not clear. One of the major obstacles is the lack of a reliable in vitro system to investigate the molecular and cellular changes in tenocytes.
Standard monolayer cell cultures have been well-established and successfully used to understand the regulatory mechanisms for cellular differentiation and homeostasis in many musculoskeletal tissues such as bone, cartilage, and muscle (17–20). However, there are no established tendon cell lines and standard monolayer culture systems for studying tendon cells. Therefore, the tendon cell culture relies on the primary cells isolated from various tendons. However, the tenocyte phenotype is not maintained in monolayer culture, and there is variability in cells by isolation method, mouse age, and types of tendons (21, 22). It is also challenging to study ECM organization and morphological maturation of tenocytes without a 3-dimensional (3D) environment.
Much of our current understanding of tendon development and postnatal tendon maturation has come from the use of murine genetic models (23). Single-cell transcriptomic studies have revealed the complex and varied cellular landscape of tendons, consisting of both intrinsic and extrinsic cell populations such as tendon fibroblasts, macrophages, endothelial cells, and pericytes (24–26). Tendon maturation is also regulated by multiple factors such as biological signaling pathways and various mechanical forces (27). Due to this complexity, in vivo mouse models are limited in their ability to mechanistically investigate the independent biological or biomechanical factors that regulate tenocyte and matrix maturation. Therefore, the development of a reliable 3D in vitro tendon culture system will be beneficial to overcome the limitations of the monolayer cell culture systems and animal models.
Recent scaffold-free and cell-based tissue engineering approaches have developed several unique in vitro tendon-like constructs. Tendon constructs have been generated from contracted or rolled-up monolayer cell culture, and implanted to treat tendon injuries, in both small and large animal models (28–30). Another approach utilized 3D dialysis-tube-based roller culture to produce fiber-like cell aggregates presenting tendon-like organization of cells and collagen fibers (31). At the single-fiber scale, a micromold-based technique was used to generate single cellular fibers, wherein cellular self-assembly and fiber formation was directed by differentially adherent growth channels coated with fibronectin (32, 33). These scaffold-free approaches have the advantage of investigating the biological mechanism underlying tendon development, compared to scaffold-based techniques, because the inherited ability of cells determines the self-assembly and the extracellular matrix formation of the constructs, rather than the remodeling of a pre-existing scaffold structure. However, the application of many of these scaffold-free approaches has been limited to studies of embryonic tendon development, due to high cell density and immature matrix organization.
In the current study, we developed a tissue-scale scaffold-free 3D tendon culture system by modifying a previously published method for engineering single fibers via differentially-adherent growth channels (32, 33). We generated tendon-like constructs that display a tissue maturation process with key similarities to postnatal mouse tendon development, including decreased cell density, increased thickness, and elongated cells between highly aligned extracellular matrix. Second Harmonic Generation (SHG) microscopy confirmed the maturation of collagen fibers, and molecular analysis verified the tenogenic differentiation in the constructs. We also tested the feasibility of genetic manipulation in our tendon culture system using adenovirus and tendon cells from Ai9 mice. Our results suggest that the 3D tendon culture system using mouse tendon cells provides a reliable in vitro system to study underlying biological and biomechanical mechanisms regulating postnatal tendon maturation.
Materials and Method
Animals
All studies were approved by the Institutional Animal Care and Use Committee (IACUC) and University Laboratory Animal Resources (ULAR) at the University of Pennsylvania (Philadelphia, Pennsylvania, USA). The study was carried out in compliance with the ARRIVE guidelines (34). The mice from mixed background lines were used to generate the 3D tendon structure. The Ai9 mouse line was purchased from Jackson laboratory (007909) and used to test the feasibility of genetic manipulation by the adenovirus system in the 3D tendon structure (35).
Growth channel assembly
We modified the previously published growth channel self-assembly (32, 33). Growth channels were made by 3D printed mold with liquid 2% agarose in αMEM on a 6-well plate (Figure 1A). The length of channel is one centimeter, and the thickness of the channel is one millimeter. The mold was removed after gelling the agarose for 30 minutes. The agarose channels were sterilized by UV for 30 minutes. Anchors, 3D-printed cylinder wrapped with the hydrophilized electrospun polycaprolactone (PCL), were inserted into both ends of the growth channel area (Figure 1B), and UV sterilization was performed again for 30 minutes. The growth channel was coated with Human plasma-derived fibronectin (0.375mg/ml in 1X PBS, Corning) and dried for 20 minutes.

(A) The base of the 3D tendon cell culture consisted of growth channel area molded into 2% agarose, using 3D-printed-mold, and cylindrical anchors wrapped by hydrophilized PCL. (B) Primary cells from mouse tail tendon were seeded (2.5 × 106 cells per construct) into the fibronectin-coated growth area to generate a 3D tendon construct. (C)The length of the resulting 3D tendon constructs ranged from 7 to 8 mm.
Monolayer culture for primary tail tendon cell
Tail tendons were isolated from 25-28 days old mice. The isolated tail tendons were digested with 5ml of type I collagenase solution (2mg/mL in 1X PBS, Sigma C0130) for one hour in a 37 °C incubator by gently inverting every 10 minutes. The digested tail tendons were cultured in six-well plates with tendon growth medium (αMEM, Gibco) supplemented with 20% Fetal Bovine Serum (FBS, New Zealand origin, Sigma F8317), 1% Penicillin-Streptomycin, and 2mM L-Glutamine) for four days until 80-90% confluency. Then, tendon cells were passaged from a six-well plate to a 100mm tissue culture plate and grown in tendon growth medium for two days till 80-90% of confluency. Finally, the tendon cells were split into five 100mm tissue culture plates (1:5 split ratio). Cells were grown for two more days untill 80-90% confluent, then harvested for 3D tendon cultures. The timeline of monolayer culture is summarized in figure 2 (Figure 2A and 2B).

(A) Three major steps of 3D tendon culture, cell extraction, cell growth in monolayer, and 3D culture. (B) The timeline of monolayer tendon cell culture after cell extraction (P1, passage1; P2, passage 2). (C) The timeline of 3D tendon culture following monolayer culture (D, days after seeding of cells for 3D tendon culture; T, days after TGFβ treatment).
Three-Dimensional (3D) tendon cell culture
Cells were seeded (2.5 × 10^6 cells/well) into the fibronectin-coated agarose growth channel area in each well. Growth media (a-MEM supplemented with 20% fetal bovine serum, 1% penicillin-streptomycin, 2mM L-Glutamine, and 50ug/ml L-Ascorbic acid) was added to the culture plate ten minutes after seeding (Figure 1B), and the seeded culture plates were placed into a standard cell culture incubator (37°C, 5% CO2, and 95% relative humidity). Two days after seeding, growth media was replaced with differentiation media (a-MEM, supplemented with 10% fetal bovine serum, 1% penicillin-streptomycin, Tgfβ (5ng/ml), 2mM L-Glutamine, and 50ug/ml L-Ascorbic acid), and differentiation media was replaced every other day for the duration of time in culture. The timeline of 3D tendon culture is summarized in Figure 2 (Figure 2A and 2C).
Generation of anchors
One-centimeter-wide electrospun polycaprolactone (PCL) strips were rolled around the anchor and tied by a surgical thread. Subsequently, the anchors were hydrophilized by serial soaking with ethanol (100%, 70%, 50%, and 30%) and distilled water for two hours at each step. Anchors were stored in 1X PBS at 4 °C until use.
Virus infection
Adenoviral vectors (Ad5-CMV-GFP and Ad5-CMV-Cre-eGFP) were purchased from the vector development laboratory at Baylor College of Medicine. The 3D tendon constructs were infected with adenoviral vectors one day after seeding. The 3D tendon constructs were washed with PBS twice to remove the existing media, and fresh differentiation media with adenoviral vectors was added the 3D tendon constructs. The concentration of adenovirus was calculated to be 300 MOI. The next day, the virus was removed by replacing the whole media with fresh differentiation media.
RT-PCR and quantitative real-time PCR (qRT-PCR)
RNA was extracted with Trizol and Qiagen RNeasy kit (full name and cat number) from 3D tendon constructs. cDNA was synthesized from 500 ng of total RNA using iScript Reverse Transcription (Bio-Rad, 1708841). The qRT-PCR was performed using SYBR green master mix (Pec, 4385612) on Quantstudio 6. The sequences of primers are listed in Table 1.
Primer sequences for qRT-PCR
Histological analyses
3D tendon constructs were harvested at various stages and fixed in 4% paraformaldehyde solution for one hour at 4 °C. The fixed samples were paraffin-embedded and sectioned with 6µm thickness using microtome for Hematoxylin and Eosin (H&E) and DAPI staining. Nucleus shape and cell density in the outer layer of the 3D tendon structure were analyzed using DAPI-stained samples. The shape of the nucleus was characterized by the ratio of the nuclear transverse length to its length in the direction of the channel length. The fixed samples were embedded in optimal cutting temperature (OCT) compound and sectioned at 10 μm using cryostat for fluorescence imaging.
Second Harmonic Generation
The paraffin-embedded sections were mounted with an anti-fade DAPI solution (Invitrogen, P36935) for imaging. Second Harmonic Generation (SHG) Images were generated by a Leica TCS SP8 MP multiphoton microscope (Leica Microsystems, Buffalo Grove, IL) equipped with a Coherent Chameleon Vision II Ti-Sapphire laser (Coherent, Santa Clara, CA). The images were acquired using excitation at 880 nm. DAPI images were acquired using 405 nm excitation and detection from 414-471 nm. The objective lens was a 40x/1.3 NA HC PL APO CS2 oil immersion lens. Z-stacks were acquired using a zoom of 2, scan speed of 600 Hz, bidirectional scanning, and 6-line averaging. Lateral (x-y) pixel size was 0.0865 micron; z-step size was 0.50 micron.
Statistical analysis
Results are expressed as mean ± SD. At least three 3D tendons per group were analyzed. Differences between values were analyzed by Student’s t-test. P<0.05 is considered significant.
Results
TGFβ is essential for 3D tendon culture
The self-assembly of seeded cells in the fibronectin-coated channel formed the tendon-like construct within 24 hours after the seeding. However, the thickness of the 3D tendon constructs gradually decreased throughout the culture time (Figure 3A). Previous studies suggested that growth factors could be essential for maintaining the engineered tendon constructs (29, 31, 32). Transforming growth factor beta (Tgfβ) is one of the well-known growth factors and is critical for tendon development and tenogenic differentiation in vivo and in vitro (16, 31, 36–43). These previous studies prompted us to use Tgfβ in the 3D tendon culture. Interestingly, the thickness of the construct was well maintained with Tgfβ when compared to the vehicle-treated construct at T14 (Figure 3A). Further analysis with higher magnification microscopy confirmed the reduced thickness at earlier stages such as T4 and T7. Cells were budding out from the surface of the 3D tendon construct without Tgfβ treatment, while Tgfβ treated construct maintained a smooth surface without budding cells (Figure 3B). These results suggest that Tgfβ is essential to promote proper aggregation and self-assembly for the 3D tendon culture.

(A) Dissecting microscope images of 3D tendon constructs at 14 days after TGFβ or vehicle treatment (T14). (B) Higher magnification images of 3D tendon constructs at T4 and T7 after TGFβ or vehicle treatment. Scale bars in lower magnification images indicate 100μm and scale bars in higher magnification images indicate 50μm.
The outer layer of the 3D tendon construct exhibits tissue maturation similar to those in postnatal tendon development
To analyze the structure of the 3D tendon construct, we collected the 3D tendon constructs at different time points and performed histological analysis using H&E-stained longitudinal sections (Figure 4). Interestingly, the outer layer of the construct displays tendon-like histology (Figure 4A, blue box), and presented a similar tissue maturation process that is observed during postnatal mouse tendon development, including flat cells between a highly aligned extracellular matrix (Figure 4A, blue box), increased thickness (Figure 4B), and decreased cell density (Figure 4C). Second Harmonic Generation (SHG) microscopy was performed with 3D tendon constructs to assess the collagen fiber formation (Figure 5). SHG signals were weak until T7, but strong linear SHG signals were detected at T14 and T21, suggesting well-organized collagen fiber formation at later stages. Our histological analysis indicates that the outer layer of the 3D tendon construct undergoes cellular and structural maturation like the postnatal mouse tendon development.
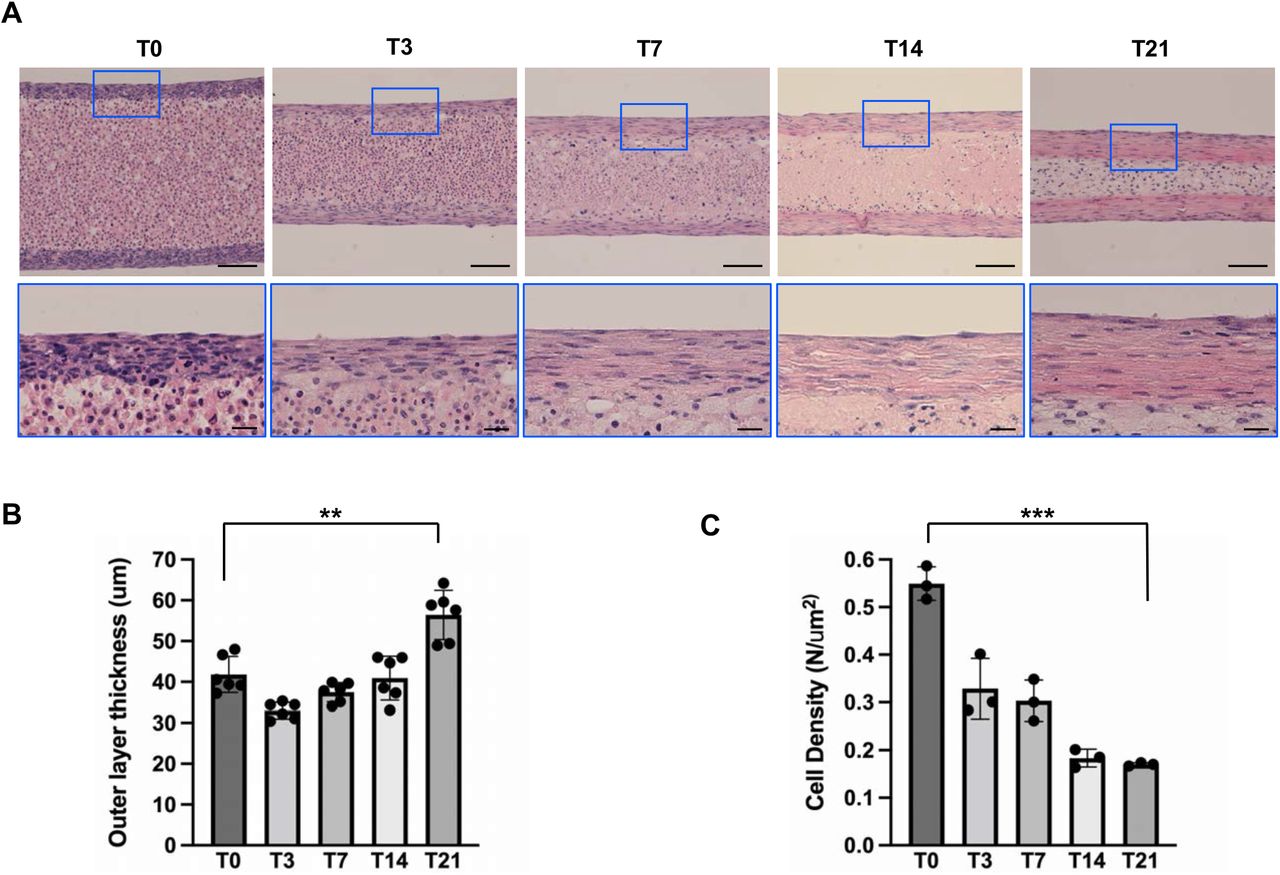
(A) H&E-stained longitudinal sections from the 3D tendons at each stage. Bottom panels are higher magnification of regions of interest (color-corresponding boxed area) of upper panels. (B) Quantification results of outer layer thickness (** indicates P<0.001, n=6), illustrating a significant thickening over 21 days. (C) Quantification results of cell density in outer layer of 3D tendon constructs (*** indicates P<0.005, n=3) revealed a decrease in cell density with construct maturation over 21 days. Scale bars in lower magnification images indicate 100μm and scale bars in higher magnification images indicate 20μm.

Top panels show the Second Harmonic Generation (SHG) microscopy images (collagen fiber formation) of outer layer of 3D tendon constructs at each stage. Middle panels show the DAPI stained images (nucleus) at each stage. Bottom panels are merged images of SHG and DAPI. All scale bars indicate 10μm.
Tenogenic maturation in the 3D tendon constructs
To further investigate cellular maturation, we first examined the morphological changes of cells in the outer layer of the 3D tendon construct (Figure 6). The DAPI-stained image showed that cell density decreased, and cells became more elongated throughout the stages (Figure 6A). To quantify the morphological changes in cells, we measured the ratio of the nuclear transverse length to its length in the direction of the channel length. (Figure 6B). In this measurement, values closer to 1.0 indicates a rounder cell shape, and values closer to 0 indicate a more elongated cell shape. The cells in the 3D tendon constructs are relatively rounder at T0, which is evident with the values closer to 1 (Figure 6B). The cells became relatively elongated at T7 and extremely elongated at T14 and T21, which is evident with the values closer to 0 (Figure 6B). To access the molecular changes in the 3D tendon construct, we examined the expression of tenogenic gene markers. Consistent with histological changes, the expression of tenogenic genes is increased at the later stages (Figure 6C). Specifically, the expression of Scleraxis (Scx), Tenomodulin (Tnmd), and Collagen type 1 (Col1) are increased. The histological and molecular analyses suggest that cells in the outer layer of 3D tendon constructs undergo tenogenic maturation.

To further investigate cellular maturation, we first examined the morphological changes of cells in the outer layer of the 3D tendon construct. (A) The DAPI-stained image showed that cell density decreased, and cells became more elongated throughout the stages. (B) To quantify the imaging results, we measured the ratio of the nuclear transverse length to its length in the direction of the channel length. In this measurement, values closer to 1.0 indicates a rounder cell shape, and values closer to 0 indicate a more elongated cell shape. (C) Quantitative real-time PCR (qRT-PCR) was performed to analyze the expression of tendon-related genes in 3D tendon constructs including Scleraxis (Scx), Tenomodulin (Tnmd), and Collagen type I (Col1). *** indicates P<0.005, n=3 used for analysis.
Genetic manipulation in 3D tendon culture
To test the feasibility of genetic manipulation in 3D tendon culture, we generated a 3D tendon constructs using tendon cells from Ai9 mice that can express the tdTomato gene upon CRE recombinase expression (35). We compared the 3D tendon constructs infected with the adenovirus expressing eGFP only (Ad-CMV-eGFP) or infected by the adenovirus expressing eGFP and CRE recombinase (Ad-CMV-Cre-eGFP). Virus infection was confirmed by the expression of eGFP in the outer layer of the 3D tendon constructs infected by both viruses (Figure 7A, 7E, 7I, and 7M, Green color). CRE recombinase activity was only detected in the 3D tendon constructs infected by Ad-CMV-Cre-eGFP, which is evidenced by the expression of tdTomato (Figure 7F and 7N, Red color). These data suggest that we can manipulate gene expression using adenovirus-mediated infection in the 3D tendon culture.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Upper two rows (A-H) are generated by longitudinal section, and bottom two panels (I-P) are generated by cross section of 3D tendon constructs generated by tendon cells isolated from Ai9 mice. Successful virus infection was indicated by eGFP expression (green fluorescence (A, E, I, and M)). CRE recombinase activity was indicated by tdTomato expression (red fluorescence (F and N)). Blue fluorescence indicates DAPI staining for nucleus (C, G, K, and O). Control virus indicates Ad-CMV-eGFP virus and CRE expressing virus indicates Ad-CMV-Cre-eGFP. All scale bars indicate 100μm.
Discussion
In this study, we successfully generated tendon-like constructs using mouse tail tendon cells. The outer layer of constructs showed tissue maturation similar to the postnatal mouse tendon development regarding molecular, cellular, and matrix changes. We tested the potential application of our tendon culture system for gene manipulation studies. These results suggest that our scaffold-free 3D tendon culture can be a reliable in vitro system to investigate the underlying mechanisms regulating cellular and matrix maturation in tendon. Previous studies suggested the potential requirement of growth factors for the formation of engineered tendon constructs (29, 31, 32). Our result showed that Tgfβ is essential for successful 3D tendon culture. Tgfβ treatment maintains the 3D tendon construct by preventing cells from budding out of the constructs (Figure 1). Tgfβ signaling is critical for tendon development and maturation by inducing tenogenic differentiation and extracellular matrix production (16, 39, 41). Besides Tgfβ, the functions of several growth factors, such as BMPs, CTGF, and FGFs, have been investigated in physiological and pathological tendon conditions using in vivo and in vitro models (16, 44–47). Upon the existence of recombinant proteins of these growth factors, we can use our 3D tendon constructs to test the function of these growth factors in tenogenic cell differentiation and matrix maturation. In addition, the screening for tenogenic factors using this construct will be an interesting future direction.
Several groups developed the scaffold-free tendon constructs by taking advantage of cellular self-assembly (27–32). However, most of the previous constructs exhibited cellular and matrix maturation similar to the embryonic tendon. Our constructs exhibited the characteristics of postnatal tendon maturation. Cells in our 3D tendon constructs undergo morphological maturation from round to elongated cell shape (Figure 4 and Figure 6). The cell density is gradually decreased as the thickness of the tendon-like structure is increased (Figure 4). Second Harmonic Generation (SHG) microscopy showed apparent collagen fiber maturation (Figure 5). This well-characterized construct will be a valuable tool to study cellular and molecular mechanisms regulating postnatal tendon maturation in vitro and give insight into bioengineered graft models. In addition, genetic manipulation is feasible for our 3D tendon culture using an adenovirus system, which suggests the potential usage of our constructs for in vitro genetic studies for specific gene functions in tendon maturation.
The success of the initial monolayer culture is critical for the generation of 3D tendon constructs. In our preliminary study, we found that the tail tendon cells from mice older than one-month grew more slowly in monolayer culture, and we never obtained enough cells for 3D tendon culture in the proper timeline. However, tail tendons from younger mice, e.g., 25-28 days old, were able to produce sufficient cell numbers for the studies herein. We assume that this could be attributed to tendons from young mice have more progenitor cells which can proliferate in vitro monolayer cell culture. Interestingly, the origin of FBS was also critical for the monolayer tendon cell cultures. Our preliminary studies revealed that the primary tendon cells in the monolayer culture with US-originated FBS grew very slow, so these cells could not be used for 3D tendon culture. But the cells with New Zealand-originated and Australian-originated FBS grew well so that we could follow the cell culture timeline. Therefore, we only isolated tail tendon cells from younger than one-month-old mice and used New Zealand-originated FBS for the current study.
We only used mouse tail tendon cells for the current study because we could easily isolate enough cells for 3D tendon culture We can generate six tendon-like constructs from one mouse tail by strictly following the timelines of the monolayer cell culture demonstrated in the Method section. In our experience, neither the Achilles nor Patellar tendons from one mouse could provide enough cells, even at the monolayer cell culture stage. However, we believe that combining multiple tendons from several mice would provide sufficient cells for the 3D tendon culture using Achilles or Patellar tendons. One of the future research directions using this platform is to compare the 3D tendon constructs generated using cells from different tendons, such as energy-storing and positional tendons, to identify whether the origin of cells influences the development, structure, and function of engineered 3D tendon constructs.
In our 3D tendon culture systems, the outer layer of the construct showed a tendon-like structure. The central regions of the construct exhibited degeneration, and eventually, the inner core tissues reduced their thickness (Figure 4A). We speculate that this is due to the nutritional deficiency in the center region. This phenomenon was also described in the previous culture study (31). The potential solution for core cell degeneration can be combined treatment of biomechanical force and growth factors. We are developing a bioreactor in which we can apply cyclic loading to the 3D tendon construct. Investigating the combinatory effects of the cyclic loading and growth factors on the maturation of 3D tendon constructs will be an interesting future direction.
Data Availability Statement
The data that support the findings of this study are available in the methods of this article.
Conflict of Interest Statement
The authors have declared that no conflict of interest exists.
Author Contributions
Study conception and design: KSJ, NAD, DTC, YL, SJH, RLM; Acquisition of data: YL, NRP; Analysis and interpretation of data: YL, KSJ, DTC, NAD, RLM; Drafting of the manuscript: YL, DTC, KSJ.
Acknowledgement
We would like to thank Penn Center for Musculoskeletal Disorders (PCMD) Histology and Biomechanics Core for technical assistance with the histology and 3D printing, respectively. We also thank Cell and Developmental Biology (CDB) Microscopy Core for technical assistance with confocal microscopy. This work was supported in part by a grant from the U.S. National Institutes of Health (NIH), National Institute of Arthritis and Musculoskeletal and Skin Diseases (KSJ: R01 AR079486). The PCMD Cores were supported by a grant from NIH/NIAMS P30AR069619.
Abbreviations
- 3D
- three dimensional
- Scx
- Scleraxis
- Col1
- Collagen type I
- Tnmd
- Tenomodulin
- DAPI
- 4′,6-diamidino-2-phenylindole
- FBS
- Fetal Bovine Serum
- ECM
- Extracellular matrix
- SHG
- Second Harmonic Generation (SHG)
- PBS
- Phosphate-buffered saline
- PCL
- Polycaprolactone
References




