

Abstract
The gut microbiome shape the host immune system and influence the outcomes of various cardiovascular disorders, meanwhile, it is a malleable microbial community that can be remodeled in response to various factors, including medications. Experiment implicated ticagrelor, one of the P2Y12 inhibitors, exerted antibacterial activity against gram-positive bacteria established from clinical evidence that it could reduce the incidence of infection-related disease. Here we performed 16S rRNA and metagenomic analysis from patients with unstable angina pectoris (UAP) treated with ticagrelor plus aspirin or clopidogrel plus aspirin for one month to determine the composition and functions of the gut microbiome difference between the two main medications. Our results suggested that the functional peptidoglycan (PGN) and Staphylococcus aureus (S. aureus) infection biosynthesis pathways were downregulated with ticagrelor-aspirin treatment in the gut. Furthermore, we found these changes were accompanied by increasing peripheral regulatory T cells (Tregs) and ectonucleotidases CD39/CD73 expressions with responding to ameliorating inflammation. To further validate the result, 16S rRNA and metabolomic analysis were carried out upon Western-diet (WD)-fed ApoE-/- mice, we found mice treated with ticagrelor plus aspirin exerted optimal synergistic effects on reducing the plaque burden and ameliorating inflammation through altered composition and functions of the gut microbiome which keeping in line with our clinical findings. The current study, for the first time, demonstrated the athero-protective effect of ticagrelor and displayed its therapeutic value at least partially attributed to its manipulation of gut microbiota.

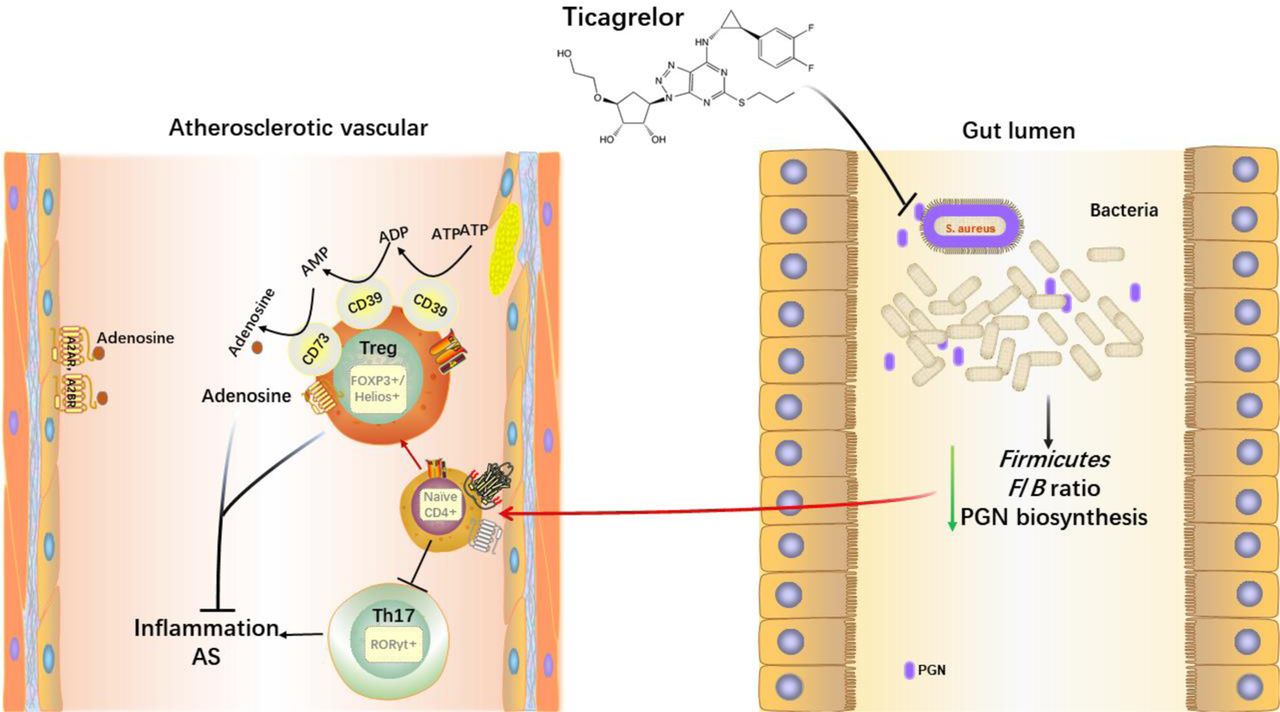
Graphic abstractTicagrelor acts on intestinal bacteria after oral administration, reducing several pathogenic Gram-positive (G+) bacteria at phyla Firmicutes and the Firmicutes/Bacteroidetes (F/B) Ratio, thus decreasing the gut peptidoglycan (PGN) synthesis. The reduction of the above pathogenic factors causes the polarization of peripheral T lymphocytes toward Treg cells. The elevated Treg cells are accompanied by an increased expression of CD39/CD73 on Tregs, which hydrolyze the extracellular ATP generated from damaged endothelial cells into adenosine (ADO) with anti-inflammatory properties and ultimately inhibited the inflammation of atherosclerosis (AS).
Introduction
Based on numerous studies, atherosclerosis (AS) has now been considered chronic inflammation, various stimuli can trigger and sustain the inflammation. As one such stimulus, chronic infection shared the common pathophysiological milieu of chronic inflammation1. A distant or direct infection of vessel wall cells has been thought to be the cause or promoter of atherosclerotic plaques with the supports by finding the bacteria DNA in the plaques 2, 3. Actually, since the middle of the 1990s, bacterial or viral infections have been considered to involve in AS4, it is increasingly believed that gut flora, as comprises a treasure trove of immunomodulatory bacteria, has a cross-link to AS risk5. Numerous studies indicated that bacteria-derived metabolites like short-chain fatty acids (SCFAs)6, 7, bile acids (BAs)8, 9, and the high-profile Trimethylamine N-Oxide (TMAO)10, regulated the risk of CVD. In addition to these metabolites, lipopolysaccharide (LPS) and peptidoglycan (PGN), known as the main components of the bacterial cell wall, can be recognized by the innate and adaptive immune system to reinforce the CVD pathogenesis potentially5. Unlike LPS restrict to gram-negative (G-) bacteria, PGN is present in both G- and gram-positive (G+) bacteria, while is predominant in G+ bacteria. A study enrolled 13 healthy individuals and 12 patients with symptomatic AS using metagenomes revealed that genes encoding PGN synthesis were enriched in patients’ gut metagenomes 11. In fact, two decades ago, PGN was found to gain access to atherosclerotic plaques and associated with the occurrence of a vulnerable plaque phenotype as a proinflammatory bacterial antigen12. Furthermore, a review recently highlighted the role of PGN as a determinative of brain inflammation13. All these facts indicated that gut PGN exerts a pro-inflammation profile in chronic inflammatory diseases, including AS. These collective findings have led to the idea that if the gut PGN biosynthesis pathway has been inhibited, can the atherosclerotic and related inflammation be ameliorated?
Adaptive immunity has a major impact on AS, with pro- and anti-atherosclerotic effects exerted by different subpopulations of T cells14. Regulatory T-cells (Tregs) are a subset of CD4+ T cells with various immunosuppressive and anti-inflammatory functions15. The transcription factors Foxp3 and Helios are the master regulators of Tregs immunosuppressive functions16, 17. It is well documented that the frequency of Tregs was reduced in the peripheral circulation of patients with carotid artery plaques18. Similarly, the number of Tregs was low in all stages of human atherosclerotic lesions, with measurements in surgical or biopsy samples19. It is well accepted that secretion of IL-10 by Tregs is important for the suppression, particularly in regulating inflammation against pathogens or foreign particles20. IL-10 has been found to play a role in prevention of AS and promotion of a stable plaque phenotype in ApoE-/- mice21. In addition of IL-10, the mechanism of T cell on the suppression of inflammation should partially due to ectoenzymes CD39 (ENTPD1, ectonucleoside triphosphate diphosphohydrolase-1) and CD73 (NT5E, ecto-5′-nucleotidases) highly expressed on Tregs that can hydrolyze extracellular ATP generated from damaged vessel walls to adenosine (ADO), subsequently suppress effector T cells by binding to its receptor ADORA2A22. Another subset of CD4+ T cells-T helpers 17 (Th17) mainly produce a strong pro-inflammatory cytokine IL-17A and have been observed in atherosclerotic plaques both in humans and animals23, 24. The expression of transcription factor nuclear receptor RORγt characterizes the Th17 cells. It is controversial of the role of IL-17A in AS, with some study suggesting that IL-17A is pro-atherogenic or athero-protective 24, 25, and a further study suggesting that IL-17A has no effect on AS26.
Dual antiplatelet therapy (DAPT), consisting of aspirin and a P2Y12 inhibitor, ticagrelor or clopidogrel, is essential for the prevention and treatment of recurrent thrombotic events in patients with ACS or MI27. As a new P2Y12 inhibitor, ticagrelor can bind to its receptor P2Y12 reversibly to conduct effect without liver enzyme metabolism. Notably, a handful of clinical trials have established the superiority of ticagrelor against ACS, including the study that ticagrelor could significantly reduce the major adverse cardiovascular events (MACE) as compared to clopidogrel in patients with ACS28 and provided ongoing benefits if continued long term29. Based on a current meta-analysis, ticagrelor could significantly reduce recurrent myocardial infarction30. Furthermore, a post hoc analysis indicated that ticagrelor could lower the mortality following pulmonary adverse events and sepsis compared to clopidogrel in the PLATelet inhibition and patient outcomes (PLATO) trial31. Evidence continued to mount that, ticagrelor exerted the potential of improving lung function in inpatients with pneumonia32. Taken together, these observations indicated the inevitable role of ticagrelor in reducing the incidence of infection-related disease while functioning its antiplatelet profile. Herein, scientists performed a study in vivo and in vitro that uncovered ticagrelor and its major metabolite AR-C124910 had antibacterial activity against all G+ strains tested and was inefficient against G- strains even in a higher concentration33.
Despite the studies above demonstrated the anti-bacteria signature of ticagrelor in vivo and in vitro, the impact of this first-line clinical medication on host gut microbiome has not been experimentally verified, where the constant dialog between gut microbiome and immune system take place. Herein, the driving concept of this study is to explore the athero-protective effect of ticagrelor in improving the immuno-inflammation via modifying the gut dysbiosis both in humans and experimental mice. Considering the privilege of its anti-bacterial characteristic, and the given close relationship of microbiota-immune interactions during the inflammation34, 35, we therefore hypothesized that ticagrelor may regulate the gut microbiota, in particular, reduce the synthesis of PGN in gut, thus potentiate anti-inflammatory property via regulating the immune system of Treg/Th17 axis. To explore the hypothesis, we attempted to comprehensively profile the composition and function of the gut microbiota by 16S rRNA sequencing and metagenomes in unstable angina pectoris (UAP) patients with the intervention of ticagrelor plus aspirin or clopidogrel plus aspirin as antiplatelet therapies, the inflammation and adaptive immunity related regulators Tregs, CD39/CD73 Tregs and Th17 cells have been detected simultaneously. To substantiate the hypothesis, mice experiment was conducted parallelly to exclude the interference factors in humans. Finally, the intervention with ticagrelor after depletion of the gut microbiome by antibiotics in mice was elicited to confirm whether the microbiota plays an indispensable role in immuno-inflammatory regulation by ticagrelor.
Results
Characteristics of the study population
From July 12 to December 25, 2021, a total of 103 participants of UAP patients were screened. 75 patients were enrolled, with 39 assigned to the ticagrelor-aspirin (TA) group and 36 allocated to the clopidogrel-aspirin (CA) group, in parallel with matched ages, gender, and BMI. Finally, 11 patients in the TA group and 10 patients in the CA group donated their fecal and blood samples on admission and one-month follow-up in the study. The clinical characteristics and concomitant medications use of the 21 patients were summarized in Table 1. There was no significant difference of previous and medications history, nor of the laboratory variables. All subjects in two groups underwent percutaneous coronary intervention (PCI) for the treatment of vessel narrowing and occlusion.
Characteristics of study participants.
TA decreased the PGN and S. aureus infection biosynthesis pathways
We performed a detailed comparison of gut microbial profiles using 16S rRNA gene sequencing and metabolomics in the fecal samples. The Shannon index at phylum level in TA group revealed a tendency toward alpha diversity compared with CA group (P=0.054, Figure 1a), in terms of β diversity, there was no distinct features between the two groups at the phylum level (Figure 1b). The relative abundance of phyla Firmicutes was visibly lower in the TA group than CA group (P=0.06, Figure 1c), but there were no differences in phyla Bacteroidetes or Firmicutes / Bacteroidetes (F/B) ratio (Figure 1c). Next, we used gut metagenomics to explore the functional pathways that the two different medications may influence. Surprisingly, we found the PGN biosynthesis pathway as the highest proportion of the whole functional pathways was lower in the TA group (the first red rectangle, Figure 1d). PGN biosynthesis contributed to S. aureus proliferation, which was also limited under TA treatment (The second red rectangle, Figure 1d). The KEGG map of PGN biosynthesis was generated and displayed associated with the S. aureus after two different treatments (Figure 1e). Strikingly, we found two PGN biosynthetic enzymes 3.4.6.14 and 1.3.1.98, especially the most predominant 3.4.6.14, were less enriched in the gut metagenomes under TA treatment (Figure 1f). The baseline of the two pathways and enzymes had no significant differences between the two groups (Figures 2Sa-d). Our data collectively suggested that ticagrelor played a potential role in regulating gut microbial dysbiosis in AS.

Distinct gut microbial composition and function under the one-month treatments of ticagrelor-aspirin and clopidogrel-aspirin in individuals with UAP. Shannon index at the phylum and genus levels by 16S rRNA (a). Principal coordinate analysis (PCA) at the phylum level was performed based on Bray-curtis (b). Relative abundance of Firmicutes and Bacteroidetes at the phylum level, and the Firmicutes to Bacteroidetes (F/B) ratio by 16S rRNA (c). Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways generated from the metagenomic analysis based on the reads number relative (d). Expression of enzymes involved in PGN biosynthesis based on the KEGG Orthology database (e). Proportions of two enzymes that had statistical differences involved in PGN biosynthesis (f). The 2-tailed Student’s t-test was used to detect significant difference between two groups (a-e). Data were expressed as mean±SEM (a, b). PGN biosynthesis pathway map (map00550) is cited from http://www.kegg.jp/kegg/kegg1.html with permission. PGN: peptidoglycan. Post TA: Post ticagrelor-aspirin. Post CA: Post clopidogrel-aspirin.
Alterations of gut microbiota in patients with TA correlated with increased Tregs
Plenty of clinical and animal experiments indicate that AS is an inflammatory disease which is involved in the onset and progression of the disease. Accumulating evidence solidly demonstrate that microbial dysbiosis notably affects the inflammatory statement in AS36 and T cells play a pivotal role in cross-linking of the inflammation and dysbiosis of intestinal flora in AS 34. Therefore, we hypothesized that the alteration of gut microbiota in AS patients with the two different antiplatelet therapy at least partially affected the adaptive immune system through regulating Treg/Th17 balance and subsequent inflammation indicators in peripheral circulation. We assessed the mentioned immune cells by flow cytometry and observed an increased frequency of Foxp3+ Tregs and Foxp3+Helios+ Tregs in the PBMC after one-month TA treatment (Figure 2a, b, e, f). Helios+ Tregs was increased in TA group compared to the baseline, while in the CA group there was nearly significant difference from baseline (P=0.07) or TA group (P=0.06) (Figure 2c, d). There was also a significant reduction of IL17A+CD4+ T helper cells in TA group and a visible reduction in the CA group from the baseline (P=0.054), nevertheless, there was no significant difference between two treated groups (Figure 2g, h). As the nuclear transcription factor of Th17, the expression of RORγt was measured, but no difference was found between pre or after treatment for each medication compared to baseline, as well as between TA and CA (Figure 2i, j). Meanwhile, qPCR was used to investigate the mRNA levels of Foxp3, Helios, RORC, together with the inflammation cytokines IL-17A, IL-10, TNF-α in PBMC, the results indicated that neither treatment significantly modified the Helios or RORC expression from baseline or between treated groups (Figure S4a, b). The expression of Foxp3 was elevated in TA group from baseline but not differed from CA group (Figure S4c). The mRNA levels of IL-17A, IL-10, TNF-α were consistently decreased in two treated groups from baseline, but there was no significant difference between the treated groups except TNF-α. (Figure S4d, e, f). Finally, the plasma levels of IL-10 and IL-17A were detected by ELISA. Unfortunately, the concentration of IL-10 was not affected in two treatments from baseline nor between TA and CA (Figure S4g). Either use of TA or CA could decrease the IL-17A level, but there still showed no significant difference between the parallel treatments (Figure S4h). These findings indicated that TA exerted the anti-inflammatory profile by up-regulating Tregs, but irrelevant to the Th17 cells and the production of IL-10 or IL-17A.

Ticagrelor-aspirin increased the Tregs but not Th17 cells compared to clopidogrel-aspirin. Representative flow cytometry plots and quantifications of regulatory T cells and Th17 cells in the PBMC from individuals of pre-treatment, TA, and CA groups (a-j). Representative flow cytometry plots (a, c, e) and quantifications (b, d, f) of Foxp3+, Helios+, and Foxp3+Helios+ Tregs respectively within the CD4+ population. Representative flow cytometry plots (g, i) and quantifications (h, j) of RORγt+ and IL-17A+ Th17 respectively within the CD4+ population. Each symbol represents an individual in diverse group. Data were shown as mean ± SEM and were analyzed by one-way ANOVA followed by a post hoc Tukey’s test. TA: ticagrelor-aspirin, CA: clopidogrel-aspirin, PBMC: Peripheral blood mononuclear cells, Foxp3: Forkhead box protein P3, a transcription factor of Treg, Helios: a transcription factor of Tregs, RORγt: RAR-related orphan receptor γt, a transcription factor of Th17. IL-17A: interleukin 17A. *P<0.05, **P<0.01, ***P<0.001
CD39-CD73-adenosine signaling is essential for TA mediated anti-inflammation
CD39-CD73 expressed on Tregs played a component role in immunosuppression, that can hydrolyze the extracellular ATP (eATP) released by dying or damaged cells into ADO37, and it functions as anti-inflammation in many impaired vessels including the atherosclerotic vessels. As illustrated in our previous findings, we next tried to detect the expression of CD39-CD73 on Tregs by flow cytometry. Our results revealed that CD39+ Tregs could be elevated both in the TA and CA groups, while TA exerted better effect than CA (Figure 3a, b). A significant elevation of CD73+ Tregs was observed in TA but not in CA from the baseline, but no superiority was seen between TA and CA (P=0.06) (Figure 3c, d). Because of fast degradation of ADO within seconds, we tested the mRNA levels of ADO receptors including ADORA2A, ADORA2B in PBMC. The outcome showed that ADORA2A and ADORA2B gene expression could be increased in TA compared to pretreatment. In contrast, we didn’t see any significant changes of ADORA2A and ADORA2B gene expression in CA treatment compared to the baseline. When it came to compare the two genes between CA and TA, we could visibly see a higher expression in TA group but without statistically significance, the P values were 0.051 and 0.08 respectively (Figure 3e, f). These observations implied that CD39 and CD73 expressed on Tregs could influence peripheral inflammation, further emphasizing the therapeutic potential of ticagrelor targeting these molecules in AS.

Representative flow cytometry plots (a, c) and quantifications (b, d) of CD39+Foxp3+ and CD73+Foxp3+cells within the CD4+ populations. The gene expressions ADORA2A and ADORA2B in PBMC were analyzed by qPCR and normalized to β-ACTIN (e, f). Each symbol represents an individual in diverse group. TA: ticagrelor-aspirin, CA: clopidogrel-aspirin. Data were shown as mean ± SEM and were analyzed by one-way ANOVA followed by a post hoc Tukey’s test. ADORA2A: adenosine receptor A2A. ADORA2B: adenosine receptor A2B. *P<0.05, **P<0.01, ***P<0.001
TA might exert optimal synergistic effects on anti-atherosclerosis profile in ApoE-/- mice
To investigate the effects of different antiplatelet medicine alone or combinations, which mimic the clinical applications, ApoE-/- mice were treated by Ticagrelor (Tica), Ticagrelor+Aspirin (TA), Clopidogrel (Clop), Clopidogrel+Aspirin (CA), Aspirin (Asp) and vehicle as control (Model) respectively. As the basic indicators, body weight, total bile acid, and lipid profiles exerted no statistical differences among the diverse groups (Figure S4a-g). Oil Red O staining of the atherosclerotic areas both in whole aorta and aorta sinus presented that compared to model, atherosclerotic lesion formation was markedly decreased in Tica and TA treated. Notably, TA exhibited better plaque burden attenuation than the other groups (Figure 4b-e, h). Any single use of Tica, Clop, Asp or combinations TA and CA could elevate the collagen deposition (Figure 4f, h), suggesting that the medications promoted the stability of atherosclerotic plaques. Collectively, these pathological detections indicated that ApoE-/- mice received either Tica, Clop, Asp alone or combination TA, CA, they all could ameliorate atherosclerostic plaques, of particular, TA seemed to exert optimal syneristc effects on athero-protective property.

Effects of different antiplatelet medications’ treatments limited plaque progression in atherosclerotic ApoE-/- mice. Schematic diagram of diverse treatments in ApoE-/-mice (a). Representative of Oil Red O in the whole aorta and aorta sinus (b, d). Quantifications of Oil Red O in the whole aorta and aorta sinus (c, e). Representative and quantification of Masson staining in aorta sinus (f, g). Representative histological H&E section of the aorta sinus (h). Scar bar, 200 μm. Tica: ticagrelor. TA: ticagrelor+aspirin. Clop: clopidogrel. CA: clopidogrel+aspirin. Asp: aspirin. Data were shown as mean ± SEM and were analyzed by one-way ANOVA followed by a post hoc Tukey’s test. *P<0.05, **P<0.01, ***P<0.001
TA modulated the gut dysbiosis in ApoE-/- mice to meet an analogous character presented in human, confirmed its mechanism of TA in decreasing the gut PGN biosynthesis and S. aureus infection pathways
PCR-free library was constructed based on Illumina Nova sequencing platform sequencing, and then Paired-End sequencing was performed. By splicing Reads, an average of 97,264 tags were measured per sample, and an average of 90,830 valid data were obtained after quality control. The rarefaction curves of all the samples displayed the sequence number nearly to 50000, indicating the sequencing depth was adequate (Figure S5a). In terms of alpha diversity, we observed significant differences between TA vs Model (P<0.05), TA vs Asp (P<0.05), and TA vs CA (P<0.05) (Figure 5a) by Shannon’s index. To further evaluate the overall gut microbiota community, β diversity of PCA based on the weighted UniFrac distances was conducted (Figure 5b), single Tica or Clop exhibited different compared to Model (P<0.01) and Asp (P<0.01), while the combination TA showed no significant differences from other groups. According to the species annotation results, the maximum value sorting method was used to select the top 5 species with the largest abundance in each group with generating a Sankey diagram. The Sankey diagram can intuitively view the relative abundance and distribution at the phylum level (Figure 5c). We could obviously conclude that in Tica and TA groups, the most predominant taxa were Bacteroidota and Verrucomicrobiota, the proportion of Firmicutes came to the third place of the whole phylum level (Figure 5c). Clop could really upregulate the Firmicutes population, while CA or Asp showed no obvious changes compared to Model by visual inspection (Figure 5c). To further explore the relative proportion of dominant taxa at the phylum and genus levels, we conducted Matastat analyze in groups (Figure 5d, e). As expected, Firmicutes were reduced remarkably in Tica and TA group compared to other groups (Figure 5d), which was consistent with our previous clinical findings (Figure 1b). Notably, the phyla Desulfobacterota in CA treated group exhibited higher proportion compared to Model or to other treatments (P<0.01, Figure 5d), which was reported as opportunistic pathogens in obese mice38. The F/B ratio decreased dramatically in Tica and TA compared to model and Clop related treatments, while Clop and CA changed the F/B ratio very slightly and had no significant difference compared to Model (Figure 5f). It is reported that Firmicutes and F/B ratio were positively related to AS progression39. At genus level, we noted that, Tica and TA could sharply reduce the Bilophila, Blautia, Clostridium_sensu_stricto_1, and [Eubacterium]_fissicatena_group compared to Model, Clop or CA treatments (Figure 5e), all these species were catalog of phylum Firmicutes (Figure S5d) and were considered pathogenic bacteria except Blautia. Instead, Parasutterella, Muribaculum were significantly elevated in TA, while Faecalibaculum and Roseburia both of them are phyla Firmicutes, were incredibly increased in Tica group (Figure S6f, g). In addition, the genera Akkermasia contributed to phyla Verrucomicrobiota increase in Tica group (Figure 5e, S5e). Finally, the functional pathways were predicted by Tax4fun. Encouragingly, the gut PGN biosynthesis and S. aureus infection pathways were obviously limited by TA treatment (Figure 5g, h), which confirmed our previous findings in human study.

Ticagrelor and Ticagrelor-aspirin regulated the gut microbiota to a “healthier” state in atherosclerotic ApoE-/- mice. α diversity of Shannon index (a) and β diversity by weighted unifrac (b). Relative abundance of taxa at the phylum level (c). Matastat analysis of the relative proportion of taxa at the phylum level (d) and genus level (e). Firmicutes to Bacteroidetes (F/B) ratio (f). PGN and S. aureus infection pathways were predicted by Tax4fun of 16S rRNA sequencing data (g, h). PGN: peptidoglycan. S. aureus: staphylococcus aureus. Tica: ticagrelor. TA: ticagrelor+aspirin. Clop: clopidogrel. CA: clopidogrel+aspirin. Asp: aspirin. Data were shown as mean ± SEM and were analyzed by one-way ANOVA followed by a post hoc Tukey’s test. *P<0.05, **P<0.01, ***P<0.001.
Treg/Th17 axis and CD39-CD73 signaling are essential for Tica and TA-mediate anti-inflammation effect in ApoE-/- mice
In view of the effectiveness of Tica and TA on atherosclerotic plaque burden amelioration and the gut microbiota regulation in mice mentioned above, we further investigated the Treg/Th17 axis, due to abundant studies indicated that PGN and S. aureus had close relationship with T cell differentiation and the inflammation13, 40–42. Hence, we investigated the Treg/Th17 cells proportions in blood and inflammation indicators in plasma. Single Foxp3+Treg, or Helios+Treg, as well as both Foxp3+ Helios+Treg cells were obviously elevated in Tica and TA compared to model and other treatments, while we didn’t see such effects in Clop and CA treatments (Figure 6a-f). Unlike our human study, in mice model the Th17 cells were remarkably reduced in all treated groups compared to Model, meanwhile TA exhibited most conspicuous effect (Figure 6g, h). To assess the inflammatory indicators in diverse groups, Cytometric Beads Array (CBA) was used to detect the pro-inflammatory IFN-γ, IL-1β, IL-6, MCP-1, TNF-α, IL-17A and anti-inflammatory IL-10. Both P2Y12 inhibitors had a marked effect on inflammatory indicators response by reducing pro-inflammatory cytokines compared to Model, and there was no significant difference between them (Figure 6i). TA showed distinct feature in lowering the IL-17A level, while in increasing IL-10, Tica did better compared to Model and other groups (Figure 6i). To clarify the relationship of CD39, CD73 and ADO, we detected the co-expressions of CD39, CD73 and ADO receptors with immunofluorescence staining in aorta sinus. Both the areas of CD39+ADORA2A+ and CD73+ADORA2A+ were increased in all treatments, while Tica and TA exhibited excellent effects than other medicine (Figure 7a-d). We previously mentioned that eATP can damage vessel wall, which can be conversed to ADO by CD39 and CD73 step by step. Hence, we tested the plasma ATP concentration to confirm whether higher expression of CD39 and CD73 correlated with lower ATP. Notably, in Tica and TA groups, the concentration of ATP could be reduced obviously compared to other groups (Figure 7e). Surprisingly, CA reduced the ATP concentration as well, the explanation maybe, besides CD73 and CD39, ADO can also be generated by alkaline through phosphatases sequential removal of phosphate groups from ATP43. Correlation analysis indicated that both elevated expressions of CD39+ADORA2A+ and CD73+ADORA2A+ had a negative correlation with plasma ATP concentration (Figure 7f, g), thus revealing that the anti-inflammatory ADO may generate from the hydrolysis of ATP by CD39 and CD73.

Ticagrelor-aspirin exerted optimal synergistic effects among all the antiplatelet medicine in balancing the Treg/Th17 axis and anti-inflammatory profile in atherosclerotic ApoE-/- mice. Representative flow cytometry plots (a, c, e) and quantifications (b, d, f) of Foxp3+, Helios+, and Foxp3+Helios+ Tregs respectively within the CD4+ population in blood. Representative flow cytometry plots (g) and quantifications (h) of IL-17A+ Th17 within the CD4+ population in blood. Inflammatory cytokines (IFN-γ, IL-1β, IL-6, MCP-1, TNF-α, IL-17A, and IL-10) were detected and quantified in plasma (i). Tica: ticagrelor. TA: ticagrelor+aspirin. Clop: clopidogrel. CA: clopidogrel+aspirin. Asp: aspirin. Data were shown as mean ± SEM and were analyzed by one-way ANOVA followed by a post hoc Tukey’s test. *P<0.05, **P<0.01, ***P<0.001, ****P<0.0001.

Expression and relationship among the CD39, CD73, ADORA2A and ATP among diverse groups in atherosclerotic ApoE-/- mice. Representative fluorescence images of CD39 (red) with ADORA2A (green) in aortic root lesions from ApoE-/-mice in diverse groups (a). Nuclei were stained with DAPI (blue). Graph showed the number of CD39+ ADORA2A+ cells in indicated groups (c). Representative fluorescence images of CD73 (red) with ADORA2A (green) in aortic root lesions from ApoE-/-mice in diverse groups (b). Nuclei were stained with DAPI (blue). Graph showed the number of CD73+ ADORA2A+ cells in indicated groups (d). Analysis of the concentration of circulating ATP from ApoE-/-mice (e). Analysis of the correlation between the concentration of circulating ATP and the number of CD39+ ADORA2A+ (f), CD73+ ADORA2A+ (g) cells in the atherosclerotic plaques of aortic sinuses from ApoE-/-mice on different treatments. Scale bar, 40 μm. Tica: ticagrelor. TA: ticagrelor+aspirin. Clop: clopidogrel. CA: clopidogrel+aspirin. Asp: aspirin. ADORA2A: adenosine receptor A2A. DAPI: 4′,6-diamidino-2-phenylindole. Data were shown as mean ± SEM and were analyzed by one-way ANOVA followed by a post hoc Tukey’s test. *P<0.05, **P<0.01, ***P<0.001, ****P<0.0001.
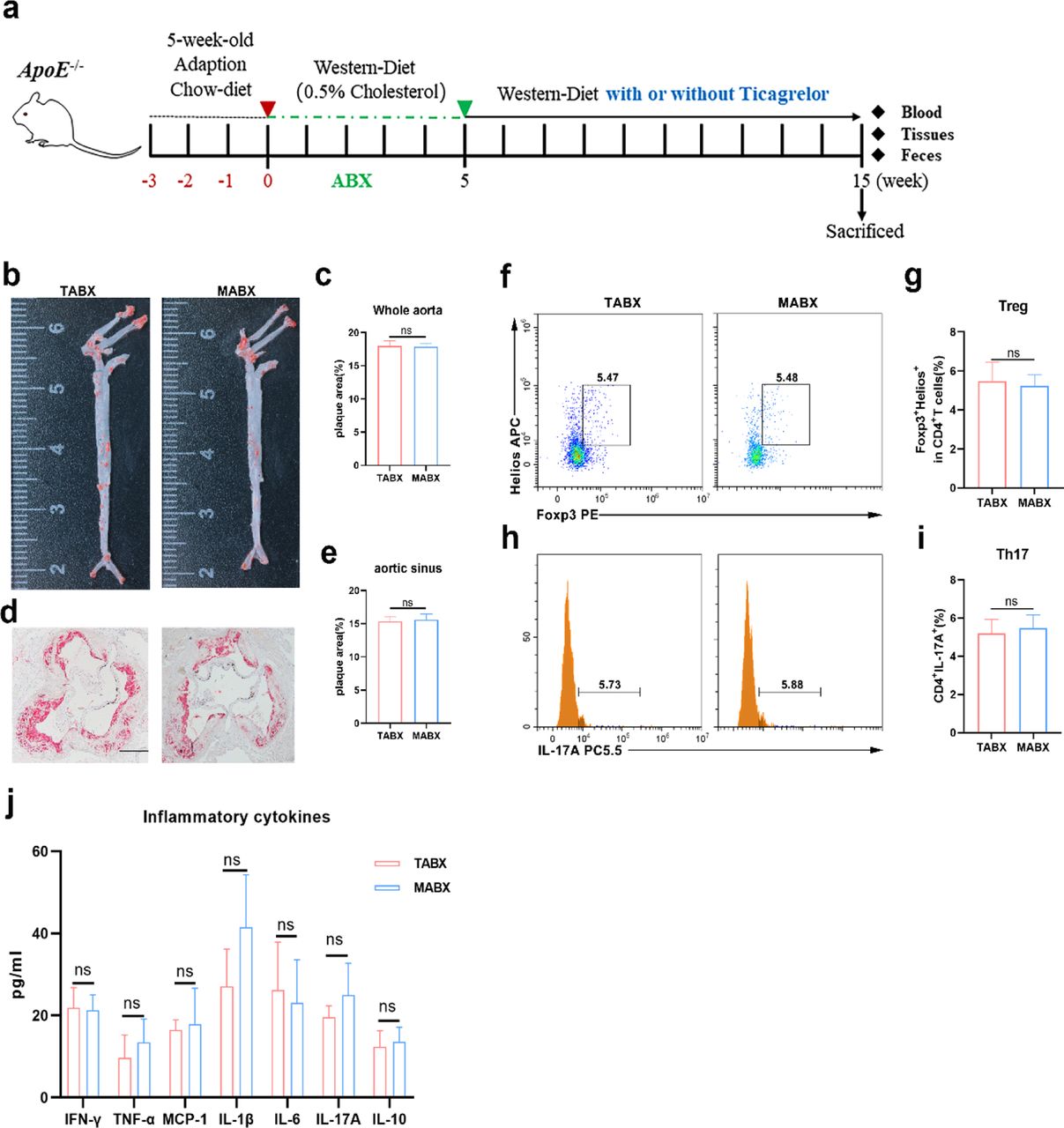
Microbiome depletion terminated the athero-protective profile by ticagrelor
To explore the association between the gut microbiome and athero-protective profile by ticagrelor in ApoE-/- mice, we treated 8-week-old mice with an antibiotic cocktail (ABX) for 5 weeks to establish an antibiotic-induced microbiome depletion condition. Depletion of microbiota with ABX was confirmed by qPCR using universal 16S rRNA primers. After 5 weeks of ABX, mice were treated with ticagrelor (TABX) or vehicle (MABX) as control respectively. The plaque burden of the whole aorta and aorta sinus in TABX did not differ from those of control mice (Figure 8b-e). Consistent with the atherosclerotic plaques, significant difference of the frequencies of Foxp3+Helios+ Tregs, IL17A+CD4+ T helper cells and inflammatory cytokines were not observed in ABX induced mice in absence of ticagrelor or not (Figure 8f-j). These findings strengthened our hypothesis that ticagrelor exerted as an immuno-inflammatory modulator in the context of AS in a microbiome-dependent manner.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Effects of Tica on plaque area, circulatory Treg/Th17 cells and inflammatory cytokines of ApoE-/- mice after removed microbiota by ABX. Schematic diagram of ABX experiment (a). Representative of Oil Red O in the whole aorta and aorta sinus (b, d). quantifications of Oil Red O in the whole aorta and aorta sinus (c, e). Representative flow cytometry plots (f, h) and quantifications (g, i) of Foxp3+Helios+ Tregs and IL-17A+ Th17 respectively within the CD4+ population in blood. Concentration of inflammatory cytokine in plasma between the two groups (j). n=5 per each group. Scar bar, 200 μm. TABX: ticagrelor-ABX, MABX: Model-ABX. Data were shown as mean ± SEM and were analyzed by the Student’ t-test.
Discussion
The immune system is a complex network composed of innate and adaptive components with an excellent capacity to adapt and respond to highly diverse challenges. Collectively, this network acts as a rigid regulator of host homeostasis in the context of microbial and environmental encounters. Studies have uncovered the critical role of the microbiota in promoting and balancing all aspects of the immune system44, 45. Growing evidence indicate that microbial alterations have been shown to regulate the inflammatory response and adaptive immune system, which is required for host defense against intestinal infections46–48. Among the adaptive immune system, Tregs and Th17 cells play a pivotal role in sustaining and restoring homeostasis in diseases49, 50, including cardiovascular disease51.
In this work, we have shown both clinical and murine measures to explore the gut microbiome changes regulated by the first-line clinical platelet-modifying medicine, the P2Y12 inhibitors, ticagrelor and clopidogrel. Our results suggested that ticagrelor or ticagrelor combined aspirin function as bacterial growth modulators in the gut and exerted their effectiveness in reducing the phylum Firmicutes and F/B ratio, which was considered to be harmful to atherosclerosis39, thus decreasing the gut PGN and S. aureus infection biosynthesis to prevent the development of AS. Our data indicated that the observed reduction of gut PGN biosynthesis resulted from altered enzymes that contributed to the pathway of PGN biosynthesis by the remodeled gut microbiota, which subsequently, led to the downregulation of S. aureus infection biosynthesis. In addition, ticagrelor and ticagrelor-aspirin in regulating gut flora suppressed the production of pro-inflammatory cytokines, increased the relative population of the circulatory of Tregs, including Foxp3+Treg, Helios+Treg, Foxp3+Helios+Treg immune cells, and the elevated production of CD39/CD73 expressed on Treg cells which can hydrolyze the extracellular ATP (eATP) generated from damaged endothelial cell to ADO contributed to the anti-inflammatory effect as well. However, there was no statistical difference in Th17 cells under the two different treatments, which is in line with the findings of the controversial role of Th17 in AS9, 24, 52, 53
Indeed, since the post hoc analysis of the PLATO trial indicated that ticagrelor was superior to clopidogrel in infectious-related diseases, studies focused on ticagrelor in reducing the S. aureus infection were popped out54–57. Our findings for the first time provided evidence for gut microbiome-dependent anti-inflammation properties of ticagrelor, which indicated that ticagrelor could affect the immuno-inflammation system in addition to its well-known antiplatelet effects. Phyla Firmicutes and Bacteroides are the most predominant taxa in the human and murine gut, and it is widely accepted that decrease of Bacteroides, increase of Firmicutes and the ratio of Firmicute to Bacteroides (F/B) contributed to promoting atherosclerosis39. Most of the G+ bacteria contribute to the Firmicutes. Surprisingly, we found Firmicutes were decreased after ticagrelor-aspirin intervention compared to clopidogrel-aspirin. In the mice study, we found the genera Bilophyla, Clostridium_sensu_stricto_1 and [Eubacterium]_fissicatena_group reduced remarkably after the treatment of ticagrelor or ticagrelor-aspirin, these species generated from Phyla Firmicutes, were considered pathogen-related bacteria as well58–61. Blautia, derived from phyla Firmicutes, with potential probiotic property62, was sharply reduced with the treatment of ticagrelor or ticagrelor-aspirin, while enriched in clopidogrel-aspirin or aspirin. These intricate results led us to determine the production of SCFAs, for one of the SCFAs, the butyric acid generated from most of the phyla Firmicutes. Surprisingly, all of the SCFAs increased slightly in ticagrelor, or ticagrelor-aspirin treated groups (Data available), together with the genera Faecalibaculum and Roseburia contributed to phyla Firmicutes incredibly enriched in ticagrelor treated group, these data reinforced the concept that ticagrelor functions as modifying the microbiome even though it targets the phylum Firmicutes. Meanwhile, the metagenome analysis uncovered that the bacterial PGN biosynthesis was limited by ticagrelor. PGN, a well-known component of the bacterial cell wall, is especially prominent in G+ bacteria (up to 70 layers), can exert robust inflammatory effects63, including T cell polarizing cytokines, which are required for T helper cells activation64. In our study, we found a decreased biosynthesis of PGN under ticagrelor-aspirin treated accompanied with increased production of peripheric Tregs, while mice were under antibiotic-induced microbiome depletion, the proportion of Tregs kept at the same level in presence or absence of ticagrelor, which validated the hypothesis of microbiota-mediated anti-inflammation and immune regulation by ticagrelor.
Tregs mediated suppression functions as negative regulation of immune-mediated inflammation and features prominently in acute and chronic infections, cancer, and metabolic inflammation65. Foxp3, the transcription factor that specifies the Treg cell lineage, essential for Treg cell differentiation and suppressor function and defines the Treg cell lineage65, 66. Helios (IKZF2) belongs to the Ikaros transcription factor family, is found critically required to maintain a stable Treg cell phenotype in the inflammatory diseases in recent years67. To be noted, studies indicated Helios enhances regulatory T cell function in cooperation with Foxp3 in patients with rheumatoid arthritis68. Our results demonstrated that the treatment with ticagrelor could obviously up-regulate the expression of Foxp3+ Tregs, Helios+ Tregs, and Foxp3+Helios+ Tregs, which displayed suppressor function of adaptive immune system in AS. By contrast, a previous study which enrolled volunteers to determine the two main P2Y12 inhibitors response to systemic inflammation by injection of Escherichia coli endotoxin, the result of the study indicated that ticagrelor could obviously increase the anti-inflammatory cytokine IL-10, however, we didn’t see any perturbation of IL-10 level under established groups. Discrepancies between our findings and this previous report may be explained as a cytokine related to Tregs, IL-10 is produced by other type of cells as well, such as macrophage, mast cell, eosinophils, neutrophils, B cells, natural killer cells and dendritic cells, resulting the dual function of anti- and pro-inflammation69. Additional explanation of stable level of IL-10 after using the P2Y12 inhibitors may on account of little is known about the interplay of IL-10 and other pro-/anti-inflammatory cytokines in the pathogenesis of AS. In recent years, Th17 cells, a subset of CD4+ T cells, were found to be pro- and anti-atherogenic effects70. In our study, single use of P2Y12 inhibitors or they combined with aspirin respectively could reduce the production IL-17A compared to baseline, which in line with the study that ticagrelor and clopidogrel could obviously reduce the development of Th17 cells in EAE, an experimental multiple sclerosis model71. But we found there was no superiority of ticagrelor to clopidogrel. IL-17A is the cytokine that produced by Th17 cells, but γδ T cells represent another source of IL-17 in intestinal. Our unshown data of mice exhibited no difference population of γδ T cells in the diverse groups. Furthermore, IL-17 family members contain IL-17A, IL-17B, IL-17C. IL-17D, IL-17E, IL-17F, the crosslink of these IL-17 family members in AS has not yet been studied. It seemed like that in our study, Tregs and Th17, partners of trading off and taking turns, left elevated Tregs to display the suppressor function generated by ticagrelor via a microbiota-dependent manner.
eATP as a kind of danger-associated molecular patterns (DAMPs), binds to purinergic receptors to trigger signaling cascades to induce an inflammatory response72. A mice model clarified the fact eATP increased released in AS and enhanced AS via binding to its receptor P2Y2 through leukocyte and monocyte recruitment73. CD39, together with CD73, both ectoenzymes that hydrolyze eATP to pericellular ADO step by step. Evidence elucidated the absence of CD39 and CD73 exacerbate the progression of AS37, 74. Extracellular ADO exerts its biological effects through the engagement of 4 distinct subtypes of ADO receptors, namely, ADORA1, ADORA2A, ADORA2B and ADORA3 receptors. Immunosuppressive activities of ADO are mainly mediated by Gs-coupled ADORA2A and ADORA2B receptors75. Researchers concluded that CD39 and CD73 are surface markers of Tregs that impart a specific biochemical signature characterized by ADO generation that has functional relevance for cellular immunoregulation22. In the current study, we detected the ADO receptors, ADORA2A and ADORA2B as indirect markers of ADO due to the fast degradation of ADO within seconds by no effective means. Our data suggested that altered microbiota induced by ticagrelor or ticagrelor-aspirin activated the Tregs and their increased production of CD39 and CD73 might limit the systemic inflammation in AS.
In conclusion, ticagrelor or ticagrelor-aspirin mediated multiple alterations of composition and function in gut microbiota, ameliorated inflammation status, and uncovered a potential mechanism of CD39-CD73-Treg associated with better AS outcomes beyond the function as P2Y12 inhibitors. Our findings shed new light on the implication of the intestinal microbiota–immune system–cardiovascular axis in atherosclerotic development by the intervention of ticagrelor or tiagrelor-aspirin, specifying the indication of ticagrelor in infection related disease via modifying the gut microbiota.
Materials and Methods
Study population
A total of 103 patients with UAP were screened in, 11 of whom took Ticagrelor+Aspirin (TA) and 10 with Clopidogrel+Aspirin (CA). All patients should meet the inclusion criteria as follows: 1. Coronary atherosclerotic plaques confirmed by coronary computed tomography angiography (CCTA) or coronary angiography (CAG); 2. Age between 40-70 years old; 3. No previous history of antiplatelet therapy. Patients with: 1. Infectious, genetic, autoimmunity-mediated, metabolic diseases; 2. Digestive disorders (inflammatory bowel disease, acute/chronic diarrhea, chronic constipation); 3. Severe hepatic and renal insufficiency; 4. Chronic respiratory diseases (COPD, asthma, etc.); 5. Antibiotics treatment within the past 1 month with the course of treatment exceeding 3 days were excluded. The enrollment strategy was displayed in Figure S1. Peripheral venous blood and fecal samples were collected on admission and one-month follow-up for every enrolled patient. Plasma and PBMCs were isolated from each fresh blood sample, and PBMCs were prepared for the subsequent experiments of flow cytometry and mRNA extraction. The stool samples were collected and immediately stored at −80℃ for further study. The study was conducted in accordance with the declaration of Helsinki. All subjects provided written, informed consents for the participation of the study. All blood and fecal samples were collected with ethics approval from the General Hospital of Ningxia Medical University (No. KYLL-2021-432). The study was registered at Chinese Clinical Trial Registry (Registration number ChiCTR2100051564).
Animals and Experimental design
Seventy-two male ApoE-/- mice (C57BL/6J) and 10 male C57BL/6J mice were purchased at 5 weeks of age and were housed in under SFP conditions at 22℃ with a 12h light/dark cycle and free access to water and food. Up to 5 mice were allocated per cage and had three weeks of adaptation time with a chow diet before the experiment. Mice were fed with a Western diet containing 0.5% cholesterol together with different treatments of administration of Tica (Ticagrelor; Brilinta, 120mg/kg/day), Clop (Clopidogrel; Plavix, 48mg/kg/day), Asp (Aspirin; Bayer S.p.A, 67mg/kg/day) and the combination of TA (ticagrelor 120mg/kg/day+Aspirin 67mg/kg/day), CA (Clopidogrel 48mg/kg/day+Aspirin 67mg/kg/day) and vehicle as control (Model) respectively for 15 weeks. Dosage of ticagrelor was known to provide comparable platelet inhibitory effect in mice76, and dosages of clopidogrel and aspirin were 0.42 and 0.56-fold to ticagrelor corresponding to the dosages used in humans77. For antibiotic-induced microbiome depletion, the mice were supplied antibiotics cocktail (ABX, consisting of ampicillin 1 g/L, vancomycin 500 mg/L, neomycin 1 g/L, and metronidazole 1 g/L, all of them were bought from Sigma) to deplete gut commensal bacteria in their drinking water for 5 weeks, and fresh ABX was replaced every other day. At the end of the study, animals were euthanized with 4% sodium pentobarbital and sacrificed for the subsequent study. This animal experiment has been approved by Laboratory Animal Ethical and Welfare Committee of Ningxia Medical University (No. 2020-078).
16S rRNA sequencing, processing, and analyzing of the gut microbiota
Total microbial genomic DNA was extracted from fecal samples of humans and mice using the E.Z.N.A.® DNA Kit (Omega Bio-Tek, Norcross, GA, U.S.) according to the manufacturer’s instructions. The quality and concentration of DNA were determined by 1.0% agarose gel electrophoresis and a Nano Drop® ND-2000 spectrophotometer (Thermo Scientific Inc., USA) and kept at −80 ℃ prior to further use. The hypervariable region V3-V4 of the bacterial 16S rRNA gene was amplified with primer pairs 338F (5’-ACTCCTACGGGAGGCAGCAG-3’) and 806R (5’-GGACTACHVGGGTWTCTAAT-3’) by an ABI Gene Amp® 9700 PCR thermocycler (ABI, CA, USA). The PCR reaction mixture included 4 μL 5× Fast Pfu buffer, 2 μL 2.5 mM dNTPs, 0.8 μL each primer (5 μM), 0.4 μL Fast Pfu polymerase, 10 ng of template DNA, and ddH2O to a final volume of 20 µL. All samples were amplified in triplicate. The PCR product was extracted from 2% agarose gel and purified using the AxyPrep DNA Gel Extraction Kit (Axygen Biosciences, Union City, CA, USA) according to manufacturer’s instructions and quantified by Quantus™ Fluorometer (Promega, USA). Purified amplicons were pooled in equimolar amounts and paired-end sequenced on an Illumina MiSeq PE300 platform (Illumina, San Diego, USA) according to the standard protocols. After demultiplexing, the resulting sequences were filtered with fastp for quality control (0.19.6)78 and merged with FLASH ( v1.2.11)79. Then the high-quality sequences were denoised using DADA280 plugin in the Qiime281(version 2020.2) pipeline with recommended parameters, which obtained single-nucleotide resolution based on error profiles within samples. DADA2 denoised sequences are usually called amplicon sequence variants (ASVs). To minimize the effects of sequencing depth on alpha and beta diversity measurement, the number of sequences from each sample was rarefied to more than 40,000, which still yielded an average good’s coverage of 97.90%. Taxonomic assignment of ASVs was performed using the Naive Bayes consensus taxonomy classifier implemented in Qiime2 and the SILVA 16S rRNA database (v138). The metagenomic function was predicted by Tax4Fun 82 in mice experiment.
Bioinformatic analysis of the gut microbiota was carried out using the Majorbio Cloud platform (https://cloud.majorbio.com). Based on the ASVs information, rarefaction curves and alpha diversity indices including observed ASVs, Shannon index was calculated with Mothur v1.30.1. The similarity among the microbial communities in different samples was determined by PCA (Principal Component Analysis) based on Bray-curtis dissimilarity using Vegan v2.5-3 package in human data or by weighted unifrac in mice. The MetaStat analyzing was used to perform with the hypothesis testing on the species abundance data between groups in different levels and q-values were obtained by the correcting p-values to assess the significant differences between groups. P values of <0.05 were considered statistically significant.
Metagenomic analysis
Total genomic DNA was extracted from human fecal samples using the E.Z.N.A.® DNA Kit (Omega Bio-tek, Norcross, GA, U.S.) as manufacturer’s instructions. Concentration and purity of extracted DNA were determined with TBS-380 and NanoDrop2000, respectively. DNA extract quality was checked on 1% agarose gel. DNA extract was fragmented to an average size of about 400 bp using Covaris M220 (Gene Company Limited) for paired-end library construction, which was constructed using NEXTFLEX® Rapid DNA-Seq (Bioo Scientific, Austin, TX, USA). Paired-end sequencing was performed on Illumina NovaSeq (Illumina Inc., San Diego, CA, USA) at Majorbio Bio-Pharm Technology Co., Ltd. using NovaSeq reagent kits according to the manufacturer’s instructions (www.illumina.com).
The data were analyzed on the free online platform of Majorbio Cloud Platform (www.majorbio.com). Briefly, the paired-end Illumina reads were trimmed of adaptors, and length<50 bp or with a quality value <20 or having N bases were considered low-quality reads and were removed by fastp78 (https://github.com/OpenGene/fastp, version 0.20.0). Reads were aligned to the human genome by BWA83 (http://bio-bwa.sourceforge.net, version 0.7.9a) and any hit associated with the reads and their mated reads were removed. Metagenomics data were assembled using MEGAHIT84(https://github.com/voutcn/megahit, version 1.1.2), which makes use of succinct de Bruijn graphs. Contigs with a length ≥ 300 bp were selected as the final assembling results used for further annotation. The KEGG annotation was conducted using Diamond85 (http://www.diamondsearch.org/index.php, version 0.8.35) and transformed to the Kyoto Encyclopedia of Genes and Genomes database (http://www.genome.jp/keeg/) with an e-value cutoff of 1e-5. Reads number relative was used to downstream analysis. Two-side Wilcoson rank sum test was used to determine gene abundance significant difference between groups. P values of <0.05 were considered statistically significant.
PBMC isolated from venous blood
Peripheral blood mononuclear cells (PBMCs) were isolated from both human and mice. After 5 ml of whole blood with EDTA-coated tube were collected and centrifugated at 600×g for 10 min, plasma was isolated and stored at −80℃ for further use. Then the PBS was added to the rest of the blood and gently mixed. Next, the mixture was softly underlayed on the Lymphocyte Separation Medium and centrifugated at 600×g for 25 min at room temperature. The ratio of blood, PBS, and Lymphocyte Separation Medium was 1:1:1. PBMCs were harvested from the buffy coat layer and resuspended in media contained 10% DMSO and 90% fetal bovine serum (FBS) and stored in liquid nitrogen for downstream experiments.
Quantitative determination of mRNA expression
Total RNA were extracted from the fresh isolated PBMC, protocol was mention above, and E.Z.N.A.® Blood RNA Kit (OMEGA, #R6834) was used according to the manufacturer’s instructions. cDNA was synthesized to a total of 500ng RNA in a 20 μL system using PrimeScript™ RT Reagent kit (TAKARA, #RR036A). q-PCR were performed on anlytik jena (qTOWER 2.0) PCR machine using Perfect Start Green qPCR Super Mix (TransGen, #AQ601). The program was performed as follows: 94℃ for 30 s, 94℃ for 5 s and 60℃ for 15 s, repeated for 40 cycles; 72℃ for 15 s (melt curve). Gene expression was analyzed using SYBR Green Master mix and selected primers used in this study were listed in Table S1. The expression of target genes was normalized to the expression of β-ACTIN, and shown as fold change relative to the pre-treatment group based on the 2−△△Ct method.
Flow cytometric analysis (FACS)
PBMCs were quickly thawed at 37℃ and cell suspensions were adjusted to a density of 1×106 cells in 100 μL of medium (RPMI 1640 with 10% FBS) and stimulated with Cell Stimulation Cocktail (plus protein transport inhibitors) (500×) (eBioscienceTM, #00-4975) for 4 h at 37°C for Th17 staining whilst Foxp3/Transcription Factor Staining Buffer Set (eBioscience, #00-5523) was used for transcription factor staining as manufacturer’s instructions. Cells were stained with antibodies listed below for surface and intracellular markers (4℃, 30 min). CD4-APC-eFluor 780 (eBioscienceTM, #47-0048), CD39-PE-Cy7 (eBioscienceTM, #25-0399), CD73-FITC (eBioscience, #11-0739), Foxp3-PE (eBioscienceTM, #12-4776), Helios-eFluor 450 (eBioscienceTM, #48-9983), RORγt-APC (eBioscienceTM, #17-6988), IL-17A-PE (eBioscienceTM, #25-7179), all these above were used for human study. In mice experiment, we used the antibodies as follow: CD4-FITC (eBioscienceTM, #11-0042), Foxp3-PE (eBioscienceTM, #12-5773), Helios-APC (eBioscienceTM, #17-9883), IL-17A-PC5.5 (eBioscienceTM, #11-7177). The fluorescent staining was performed after blocking the Fc receptor with the anti-CD16/CD32 antibody. FACS acquisition was performed with the cytometer Cytomics FC500 (Beckman Coulter). Finally, the Tregs and Th17 cells were analyzed by Beckman Cyto FLEX flow cytometer (Beckman Bioscience, United States).
Measurement of cytokines in plasma
LEGENDplexTM Mouse Inflammation Standard Cocktail (Biolegend, #740371) was used to measure the cytokines in plasma of the mice according to the manufacturer’s instruction. Samples were diluted as 1:2, a total of 7 standards were used per cytokine to generate standard curve. Data were analyzed by LEGENDplex (version 8.0). For human IL-10 and IL-17A cytokines’ measurement, we used commercially available ELISA kit, IL-10 (Boster, #EK0416) and IL-17A (Boster, #EK0430) following the manufacturer’s instructions.
Histological analysis and immunofluorescence staining
Frozen aortic roots were cut in 5 μm-thick serial cryosections and stained with Oil-Red O, hematoxylin and eosin and Masson’s trichrome respectively for the quantification of lesion size and collage area. The whole aorta was isolated from carotid artery to iliac artery and was stained with Oil-Red O to access the lesion area in general. Atherosclerotic lesion of the whole aorta or aorta sinus staining above was observed using the microscope (Olympus, Japan). For immunofluorescence staining, 5 μm-thick sections of the frozen aortic roots were used to examine the expression of detected antibodies. Briefly, sections were covered 10% goat serum to block the nonspecific antigens at room temperature for 30 min. Then the sections were incubated with the primary antibodies of anti-rabbit CD39, anti-rabbit CD73, anti-mouse ADORA2A FITC (1:100, Santa Cruz, United States) overnight at 4℃. The next day, the slides were incubated with a suitable secondary antibody conjugated goat anti-rabbit IgG-HRP for 1 h at room temperature. Finally, cell nuclei were mounted with DAPI for 10 minutes at room temperature. Images were captured with blinded manner of Leica DMI3000+ DFC310FX fluorescence microscope (Leica, Germany). The positive areas in plaque were quantified by Image-Pro Plus 6.0.
Statistics
Graphpad Prism (Version 8) and SPSS 22.0 were used for all the data analysis and the value were expressed as the mean±SEM. The significant differences between groups were calculated by unpaired Student’s t test, or one-way ANOVA (Tukey’s multiple comparison test). Normality and lognormality tests were carried out before the parametric analysis of Student’s t test and one-way ANOVA. Welch’s corrections were used when variances between the groups were unequal. Pearson correlation coefficient assay was used to analyze the expression correlation. Both significant differences (P < 0.05) and trends (P < 0.1) were reported where appropriate for all tests performed.
Competing Interests
The authors have declared that no competing interest exists.
Data availability
Sequence data of 16S rRNA and metagenome that support the findings of this study have been deposited in National Center for Biotechnology Information (NCBI) with the primary accession code: SRP371254. Raw data of SCFA will be made available from the corresponding author upon reasonable request.
Abbreviations
- AS
- atherosclerosis
- CVD
- cardiovascular Disease
- ACS
- acute coronary syndrome
- ASVD
- atherosclerotic vascular disease
- MI
- myocardial infarction
- UAP
- unstable angina pectoris
- MACE
- major adverse cardiovascular events
- AE
- Adverse event
- CCTA
- coronary computed tomography angiography
- CAG
- coronary angiography
- PCI
- percutaneous coronary intervention
- DAPT
- dual antiplatelet therapy
- TMAO
- trimethylamine N-Oxide
- SCFA
- short-chain fatty acid
- LPS
- lipopolysaccharide
- PGN
- peptidoglycan
- 16S rRNA
- 16S ribosomal RNA
- Treg
- regulatory T cell
- eATP
- extracellular ATP
- ADP
- adenosine diphosphate
- AMP
- adenosine monophosphate
- ADO
- adenosine
- ADORA2A
- adenosine receptor A2A
- ADORA2B
- adenosine receptor A2B
- KEGG
- Kyoto Encyclopedia of Genes and Genomes
References
- 1.↵
- 2.↵
- 3.↵
- 4.↵
- 5.↵
- 6.↵
- 7.↵
- 8.↵
- 9.↵
- 10.↵
- 11.↵
- 12.↵
- 13.↵
- 14.↵
- 15.↵
- 16.↵
- 17.↵
- 18.↵
- 19.↵
- 20.↵
- 21.↵
- 22.↵
- 23.↵
- 24.↵
- 25.↵
- 26.↵
- 27.↵
- 28.↵
- 29.↵
- 30.↵
- 31.↵
- 32.↵
- 33.↵
- 34.↵
- 35.↵
- 36.↵
- 37.↵
- 38.↵
- 39.↵
- 40.↵
- 41.
- 42.↵
- 43.↵
- 44.↵
- 45.↵
- 46.↵
- 47.
- 48.↵
- 49.↵
- 50.↵
- 51.↵
- 52.↵
- 53.↵
- 54.↵
- 55.
- 56.
- 57.↵
- 58.↵
- 59.
- 60.
- 61.↵
- 62.↵
- 63.↵
- 64.↵
- 65.↵
- 66.↵
- 67.↵
- 68.↵
- 69.↵
- 70.↵
- 71.↵
- 72.↵
- 73.↵
- 74.↵
- 75.↵
- 76.↵
- 77.↵
- 78.↵
- 79.↵
- 80.↵
- 81.↵
- 82.↵
- 83.↵
- 84.↵
- 85.↵




