

Abstract
Clam gardens are habitat modifications established by coastal Indigenous Peoples of northwest North America to enhance intertidal clam habitat productivity to provide secure and reliable local food resources. These gardens were established long ago and are mostly unmaintained in present times. To determine whether unmaintained clam gardens still provide a more productive and beneficial habitat than unmodified clam beaches, Pacific littleneck clams (Leukoma staminea) were transplanted to unmaintained clam gardens or unmodified reference beaches and then evaluated for growth, survival, and transcriptomic signatures of the gill and digestive gland after 16 weeks. All beaches in the study were characterized for sediment grain-size distribution, carbonate concentration, and organic content to identify sediment characteristics that may differ between clam garden and reference sites, as well as potentially contribute to differences in clam productivity. Clam growth and survival, sediment grain-size distribution, and carbonate and organic content were not significantly affected by unmaintained clam gardens. Wide variations in survival, growth, and sediment characteristics were observed among beaches. Examining across beach location resulted in the identification of significant negative effects of small rocks (2.00 – 4.75 mm), and silt (< 63 µm) on both growth and survival. Sand (250 – 500 µm) had a significant positive influence on both growth and survival while fine sand (125 – 250 µm) had a significant positive effect only on growth. Coarse sand (0.50 – 1.00 mm) had a significant positive effect on survival, while very fine sand (63 – 125 µm), carbonate, and organic content all had significant negative effects. To evaluate transcriptomic effects, a de novo transcriptome for L. staminea was assembled. The final assembly contained 52,000 putative transcripts and those specific to each of the two tissues were identified. This revealed that similar functional categories were enriched in tissue-specific genes, but each tissue had its own transcripts comprising the categories. Transcriptomic analysis revealed differential expression in individuals from clam gardens, and although this effect was small in terms of the numbers of genes, specific response genes were identified consistently in both tissues. In summary, while the unmaintained clam gardens did not impact clam growth and survival over the 16 weeks of the study, it did have an effect at the level of the transcriptome. Furthermore, correlations of transcripts associated with either high or low survival provide new insights into ecological associations of these genes in this non-model organism. In summary, localized environmental factors are likely to have a greater influence on Pacific littleneck clam physiology, growth, and survival than the presence/absence of unmaintained clam gardens.
1 INTRODUCTION
Coastal shorelines have been altered to increase productivity and facilitate the harvesting of natural resources for centuries (Erlandson et al. 2008). Ancient clam gardens and fishing weirs were shoreline constructions built to increase clam habitat and trap fish, respectively, providing productive and predictable local food resources for coastal Indigenous Peoples (Neudorf et al. 2017). These coastal shoreline modifications provide important insights into cultural food practices, traditional technologies, economies, values, and ancestral practices of coastal communities (Deur et al. 2015, Jackley et al. 2016, Lepofsky et al. 2020, Smith et al. 2019).
Construction of clam gardens in North America is estimated to have begun approximately 3,500 years ago (Smith et al. 2019). These gardens were created by rolling rocks and boulders to the low-tide line and building a wall parallel to the shoreline (Deur et al. 2015), which subsequently allowed for the deposition of sediment between the wall and the high-tide mark. This created a wider intertidal shellfish habitat area with a reduced slope (Neudorf et al. 2017) at a tidal height of 1–2 m above chart datum, where shellfish such as Pacific littleneck clams (Leukoma staminea), a traditional food item for coastal Indigenous Peoples, typically occur (Deur et al. 2015, Groesbeck et al. 2014). This reduced beach slope also allowed a thin layer of seawater to be retained on the accumulated sediment, reducing desiccation risk, facilitating clams to remain in shallow accessible portions of the intertidal zone, and maximizing submersion time for feeding and therefore growth (Deur et al. 2015). The exterior (ocean) side of the clam garden wall creates rocky reef habitat for other marine invertebrates including octopuses, sea cucumbers, whelks, chitons, red turban snails, Dungeness crabs, and red rock crabs, many of which are also consumed by humans (Caldwell et al. 2012, Deur et al. 2015, Lepofsky et al. 2017).
Clam gardens are found in the intertidal zone from Alaska (USA), throughout British Columbia (BC, Canada), and into Washington State (USA) (Groesbeck et al. 2014). Northern Quadra Island, off the east coast of Vancouver Island, has perhaps the highest density of clam gardens on the northwest coast of North America (Neudorf et al. 2017). On Quadra Island there are 45 clam gardens on the northwest side of the island in Kanish Bay and 49 on the northeast side in Waiatt Bay (Groesbeck et al. 2014). In these two bays, clam gardens cover approximately 35% of the shoreline, increasing clam habitat by 26–35% (Lepofsky et al. 2020).
Traditionally, clam gardens were maintained and tended with practices that included predator exclusion, selective harvesting, and the addition of gravel and crushed shell (i.e., shell hash) (Deur et al. 2015). Shell hash may provide larval shellfish settlement cues and protect the young recruits from water acidity and predators (Green et al. 2012, Sponaugle and Lawton 1990). Clam gardens were tended into the 20th century, but most of them, including all the beaches in the present study, were not maintained at the time of the present study (Williams 2006).
Research on untended clam gardens in Kanish Bay and Waiatt Bay found significantly higher densities of Pacific littleneck clams and butter clams (Saxidomus giganteus), increased survival of clam recruits, and higher Pacific littleneck clam growth rates compared to unmodified beaches (Groesbeck et al. 2014). Studies on butter clams on the central coast of BC found that densities were 2.44 times greater on average in clam gardens compared to densities on unmodified beaches (Jackley et al. 2016). The higher density was thought to be due to the creation of a wider intertidal area in the clam gardens.
Community composition may also be influenced by clam gardens (Jackley et al. 2016). Greater abundances of many taxa occur on clam garden beaches relative to unmodified beaches, including Nematoda, Harpacticoida, Chironomidae, and Bivalvia (e.g., butter clams and Pacific littleneck clams). The abundance of these taxa has been shown to be positively associated with the amount of surface shell and gravel present (Cox et al. 2019). A study comparing grain sizes and carbonate content in beach sediment on Quadra Island found 2.8 – 12.9 times more carbonate, and on average a smaller percentage of silt, in sediment from clam gardens compared to that from unmodified beaches (Salter 2018). Similarly, greater amounts of gravel and shell hash have been observed in clam gardens relative to unmodified beaches (Groesbeck et al. 2014). Unmodified beaches nearby clam gardens are, by contrast, primarily composed of silt, sand, and mud (Jackley et al. 2016). Such differences in sediment composition are attributed to the tending practices of Indigenous Peoples, which included the addition of gravel and crushed whole clam shells to clam gardens (Groesbeck et al. 2014, Williams 2006).
The effect of sediment shell hash on clam productivity is not definitive. Sediment carbonate saturation state is a significant seawater variable that can affect the settlement and recruitment of larval and juvenile infaunal bivalves (Green et al. 2012). Seawater calcium carbonate (CaCO3) saturation state is important in the formation and maintenance of shell thickness and integrity (Chadwick et al. 2019). Ocean acidification results in a decrease in seawater pH and an alteration in the bioavailability of CaCO3. A decrease in CaCO3 saturation state can negatively affect the formation and maintenance of shells of marine molluscs and other calcifiers (Evans et al. 2014, Green et al. 2012, Waldbusser and Salisbury 2014). Some studies have shown that beaches with increased levels of sediment shell hash have increased clam settlement and productivity (Green et al. 2012, Groesbeck et al. 2014, Salter 2018), while others have reported negative (Munroe 2016) or neutral (Greiner et al. 2018) correlations with early post-settlement bivalve growth and recruitment. A study on Quadra Island found that the addition of crushed shell had a positive effect on transplanted littleneck clam growth and final tissue biomass in both unmaintained clam gardens and unmodified beaches (Salter 2018). Most of these studies looked at variables such as growth and survival over a relatively short period of time (i.e., 30 days to six months, the exception being Munroe (2016) where the trial ran for two years), whereas the effects of these variables may take a longer time to manifest as physiological changes in clams.
Genomic tools such as transcriptomics offer the ability to capture early signals of environmental stressors and are increasingly used to evaluate their biological effects on aquatic life (Milan et al. 2011, Sutherland et al. 2012). Gene-expression data can then be linked with survival, growth, and environmental information and used to predict possible physiological effects (Evans et al. 2011, Miller et al. 2017). Transcriptomics can not only yield faster results but may also be used for taxonomic identifications (Bourlat et al. 2013, Sutherland et al. 2022). The present study aims to determine whether Pacific littleneck clam health and productivity, as assessed by growth and survival as well as transcriptomic profiling, are affected by the presence of unmaintained clam gardens and various sediment characteristics (e.g., grain-size distribution, organics, and carbonates). To this end, we assembled a de novo reference transcriptome and characterized Pacific littleneck clam transcriptome responses in replicated unmaintained clam garden beaches and unmodified reference beaches in the same area. Measured environmental parameters and clam phenotypes (e.g., growth, survival) were considered while characterizing the transcriptomic responses. The combination of abiotic measurements and organismal and molecular phenotypes with the characterized transcriptome provides valuable context for both the effects of different environmental variables on Pacific littleneck clams and on the functions of response genes in this non-model ecologically and culturally important species.
2 MATERIALS AND METHODS
2.1 Study sites
The intertidal coastlines of Kanish Bay and adjoining Small Inlet on Quadra Island were chosen for this study due to an abundance of historic clam gardens. Quadra Island is located off the northeast coast of Vancouver Island, in southwest BC, and is in the traditional territories of the Kwakwaka’wakw (Laich-kwil-tach) and northern Coast Salish Peoples (Smith et al. 2019). Three unmaintained clam garden beaches (CG, designated as A, C, and E) and three unmodified reference sites (Ref, designated as B, D, and F) were selected in Kanish Bay and Small Inlet (Figure 1). Beach B was later discovered to be a damaged clam garden with ∼ 75% of the wall destroyed (possibly from logs being dragged down the beach). In this study beach B is still considered a reference beach as it does not appear to be an outlier from the other reference beaches in any of the sediment characteristics (Table 2). These clam gardens have likely not been tended in accordance with Indigenous practices since the early-to-mid 1900s (Deur et al. 2015).

Locations of experimental clam gardens (A, C, and E, designated by triangles), unmodified (reference) beaches (B, D, and E, designated by circles), and clam collection site (designated by a star) on Quadra Island, British Columbia, Canada.
2.2 Clam transplantation experiment
Juvenile Pacific littleneck clams (N = 400), between 1 and 2 cm in shell height (hinge to shell margin), were hand collected from a site in Kanish Bay (Figure 1) between 1 and 2 m Mean Lower Low Water level on May 7, 2016, with all field work occurring that year. Each was individually wet weighed, measured for shell height and length (i.e., the widest part of the shell at 90° to the shell height), and then haphazardly placed into 18 groups of 20 clams each. Clams were held in a pearl net that was deployed below the low intertidal zone in Heriot Bay (Quadra Island) until deployment. Clams on beach A were deployed on May 10; those on beaches B, C, and D on May 11; and those on beaches E and F on May 12. The different deployment days were required due to tidal heights and weather conditions. At low tide at each of the six beaches, the 1.5–1.8 m intertidal zone was demarcated and three plastic mesh cubes (length x diameter x height: 20 × 20 × 4 cm; made from high-density polyethylene, Vexar™), each containing 20 clams, were buried in the top 20 cm of the sediment, 5 m apart and parallel to the shore in the middle of the demarcated zone. Each mesh cube was defined as one replicate plot (P). Each plot was numbered so that Beach A contained P1–P3; Beach B: P10-P12; Beach C: P4-P6; Beach D: P7-P9; Beach E: P13–P15; Beach F: P16– P18.
Transplanted clams were left in situ for 16 weeks and collected on August 30–31, 2016. Upon retrieval, clams were quickly transported in bags in coolers to the Hakai Institute laboratory on Quadra Island. The wet weight, shell height, shell length, and dead/alive status were recorded for each individual.
Growth and survival per plot were calculated as:
 In addition, some visual observations were made on animal condition, following the protocols developed for the Canadian National Animal Aquatic Health Program (NAAHP), including observations of animal state (i.e., health/vitality, condition) and digestive gland colouration.
In addition, some visual observations were made on animal condition, following the protocols developed for the Canadian National Animal Aquatic Health Program (NAAHP), including observations of animal state (i.e., health/vitality, condition) and digestive gland colouration.
2.3 Sediment collection and analysis
Sediment-core samples (length x diameter: 15 × 5 cm) were collected adjacent to each cube (n = 3 cores per beach) to assess grain size of core. Additional sediment-core samples (10 × 3.5 cm) were also collected adjacent to each cube to assess percent carbonate and percent organic matter content. Each core was placed individually in labeled sample bags and frozen at -20°C until analysis.
To determine grain-size distribution, the sediment samples were dried at 100°C for 24 hr, weighed, transferred to a series of nested sieves (mesh size: 4.75 mm, 2 mm, 1 mm, 500 µm, 250 µm, 125 µm, and 63 µm), and shaken in a sediment shaker for 15 min. Each size fraction was then weighed and expressed as a percentage of the total sediment weight. A scale very similar to the Wentworth scale of sediment size classes was used to classify the grain sizes: pebble and larger gravel called “rocks” in the present study (> 4.75 mm), granule gravel called “small rocks” in the present study (2.00 – 4.75 mm), very coarse sand (1.00 – 2.00 mm), coarse sand (0.50 – 1.00 mm), sand (250 – 500 µm), fine sand (125 – 250 µm), very fine sand (63 – 125 µm), and silt (< 63 µm) (Wentworth 1922).
The loss on ignition method (Heiri et al. 2001) was used to determine sediment percent organic matter and carbonate content. Briefly, the sediment samples were dried at 100°C for 48 hr in previously-ashed (450°C for 8 hr) crucibles, and the dry weights measured. Dried sediment samples were then ashed in a muffle furnace at 435°C for 8 hr. After cooling the samples in a desiccator for 1 hr, the weights of the ashed samples were recorded. The percent organic matter was then calculated using the following equation:
 Sediment percent carbonate content was determined following 2 hr at 950°C and 2 hr of cooling in a desiccator, and calculated using the equation:
Sediment percent carbonate content was determined following 2 hr at 950°C and 2 hr of cooling in a desiccator, and calculated using the equation:

2.4 Data analysis: clam size, growth and survival, and sediment characteristics
Data were analyzed in R v.4.0.2 (R Core Team 2022). Model assumptions of normality were assessed using histograms, quantile-quantile plots of the data, and the Shapiro-Wilk test. Homogeneity of the residuals was assessed using plots of model residuals versus fitted values and Bartlett’s test for homogeneity of variance. Nested ANOVAs (plots nested within beach location), using the linear mixed-effects function in the nlme package, were used to test for differences between clam garden and unmodified reference beaches (i.e., beach type, two levels) in initial and final clam shell heights, wet weights, percent growth, and percent survival as well as individual sediment characteristics (i.e., rocks, small rocks, very coarse sand, coarse sand, sand, fine sand, very fine sand, silt, percent organic matter, and percent carbonate concentration), with beach location (six levels) included as a random factor. Significance was considered when p ≤ 0.05. ANOVAs, linear mixed-effects models, and post-hoc least squares means omitting beach type were used to test for significant differences among beaches in percent growth, percent survival, sediment carbonates, organics, and individual grain sizes. Principal component analysis (PCA) and biplot figures were generated in R using the prcomp function in the default stats package and the ggbiplot function/package. Pearson correlation between the variables was evaluated with the cor function of the stats package of R and inspected with corrplot v.0.92 (Wei and Simko 2021). All code is available (see Data Availability).
2.5 RNA sampling and extraction
For gene-expression analysis, small (∼2 × 2 mm) sections of gill and digestive gland tissues were excised from each surviving clam using sterile techniques and stored in RNAlater as per manufacturer protocols (Ambion). Bivalve gills are a target tissue in studies of rapid environmental responses, whereas digestive glands are accumulatory organs used in toxicological and immunology investigations of longer-term effects of environmental change and exposure (Gosling 2008, Milan et al. 2011). RNA from tissue sections of 25– 30 mg was individually extracted from both tissues using RNeasy kits (Qiagen) after homogenization in 2-mL tubes filled with Lysing Matrix D (MP Biomedicals) in a Tissuelyser II (Qiagen) at 25 Hz for 2 min. To eliminate DNA contamination, a DNase protocol was applied (Turbo DNA-free Kits, Ambion) that inactivates and removes the DNase enzyme after DNA digestion. RNA concentrations were quantified on a NanoDrop 1000 Spectrophotometer (Thermo Fisher Scientific) after DNase treatment.
2.6 Library preparation for RNA sequencing
RNA sequencing pools were created for each of the six locations (beaches) using five haphazardly selected individuals from each plot within the beach. Plots were only used if they had at least five surviving clams. This resulted in three plots per beach being used for all locations except for beaches E and F, which only had five or more survivors in a single plot. This produced 14 RNA pools per tissue (N = 28 pools total) to be used for library synthesis and sequencing. RNA quantities were normalized, and equimolar pools of RNA were created for both tissues of the five selected individuals from each plot. RNA quality and quantity of each pool were determined using the RNA 6000 Nano chip on a 2100 Bioanalyzer (Agilent).
Following RNA quality control checks (NanoDrop and Bioanalyzer), mRNA libraries were generated from 250 ng of total RNA using an mRNA enrichment with the NEB Next Poly(A) Magnetic Isolation Module (New England Biolabs) and cDNA synthesis was conducted using NEBNext RNA First Strand Synthesis and NEBNext Ultra Directional RNA Second Strand Synthesis Modules. Remaining library preparation used NEBNext Ultra II DNA Library Prep Kit for Illumina, with adapters and PCR primers (NEB). The remaining steps of library preparation were taken using the Quant-iT™ PicoGreen® dsDNA Assay Kit (Life Technologies) and the Kapa Illumina GA with Revised Primers-SYBR Fast Universal Kit (Kapa Biosystems (Pty) Ltd.). Average fragment size was determined using a LabChip GX instrument (PerkinElmer). Libraries were run across four lanes of an Illumina HiSeq4000 PE 100 platform (Illumina) and the mean (± SD) read pairs per library (both tissues combined) was 54.4 ± 9.5 million. All library preparation and sequencing steps were undertaken at the Genome Québec Innovation Centre (Montreal, Canada).
2.7 Transcriptome assembly and annotation
For the de novo clam transcriptome assembly, Trinity 2.5.1 (--min_kmer_cov 2) (Grabherr et al. 2011) was used to assemble the 28 libraries. Before running the assembly, Trimmomatic 0.36 (ILLUMINACLIP:TruSeq3-PE-2.fa:2:30:10 LEADING:3 TRAILING:3 SLIDINGWINDOW: 4:15 MINLEN:36) was used to clean the read data to obtain a more accurate assembly (Bolger et al. 2014). With the resulting Trinity assembly (1,695,678 sequences), the TransDecoder 5.0.1 pipeline (Haas et al. 2013) was used to predict likely coding sequences (1,277,478 sequences), which uses homology (pfam-a, UniProt) and open reading frame (ORF) information. The best representative transcript was chosen for each gene, as predicted by TransDecoder, that had an ORF type of ‘complete’, which required protein homology. In cases where there were multiple transcripts with complete ORFs for a gene, the transcript with the largest ORF was chosen. Further filtering was carried out to choose only transcripts ≥ 100 bp (to remove small transcripts that could cause analysis error), followed by scanning of pfam-a and Uniprot annotations to remove repetitive element-related keywords to finally produce a transcriptome of 52,000 transcripts, each putatively representing a single gene of which 42,708 had Uniprot IDs.
2.8 Data analysis: RNA sequencing data
Raw reads were imported to the repository Simple_reads_to_counts (see Data Availability) for quantitation. Raw reads were gently trimmed for quality and adapters removed using Trimmomatic (Bolger et al. 2014) with the following flags: ILLUMINACLIP:$VECTORS:2:30:10; SLIDINGWINDOW:20:2; LEADING:2; TRAILING:2; MINLEN:80. Both the raw and the trimmed data were inspected using FastQC (Andrews 2010) and multiQC (Ewels et al. 2016). The reference transcriptome was indexed using bowtie2 (Langmead and Salzberg 2012) and reads were aligned against it in local mode, allowing 40 alignments to be retained per read (-k 40). For comparison with the results from the reference transcriptome, the Manila clam (Ruditapes philippinarum) genome (GCA_009026015.1_ASM902601v1) (Yan et al. 2019) was also tested as a potential reference by indexing the genome using hisat2 (Kim et al. 2015) and aligning the Pacific littleneck clam reads against the Manila clam genome (also retaining a maximum of 40 alignments per read; -k 40). The littleneck clam alignments against the littleneck reference transcriptome were quantified using eXpress using default parameters (Roberts and Pachter 2013). The alignments against the Manila clam genome in the test were also quantified using eXpress default parameters but with the flag --rf-stranded. Resultant effective read counts quantified against the de novo reference transcriptome were used in downstream analyses.
Transcriptome analyses were all conducted as described in the repository ms_clam_garden (see Data Availability). Transcript annotation, effective counts, and phenotypic data were all imported into R to be analyzed with edgeR v.3.36.0 (Robinson et al. 2010). Datasets were constructed for all samples or only gill or only digestive gland (DG) samples (N = 3 datasets). Each dataset was input into a DGEList, then filtered for low expression by requiring at least 10 reads to be aligned to the transcript in at least five individuals (i.e., counts per million (cpm) > 0.82–1.03, depending on the dataset). Samples were normalized by TMM normalization as applied by calcNormFactors() and multidimensional scaling (MDS) plots were produced for each dataset (i.e., both tissues combined, gill tissue only, DG tissue only).
Tissue-specific expression was determined by identifying transcripts expressed in one tissue and not the other. The normalized counts for these transcripts were then obtained from the all-sample dataset and 200 of the highest expressed tissue-specific transcripts from both tissues were plotted using heatmap() of the stats package in R. Gene Ontology (GO) enrichment was conducted using the tissue-specific transcripts from both tissues compared to all expressed transcripts for that tissue. Enrichment analysis was conducted in DAVID Bioinformatics (Huang et al. 2007).
Differential expression analysis was conducted using limma v.3.50.1 (Ritchie et al. 2015). Both tissues were analyzed separately and differentially expressed genes were identified using beach type (i.e., CG vs. Ref) as an explanatory variable using a gene-wise negative binomial generalized linear model with quasi-likelihood tests (glmQLFit) and retaining all transcripts with p ≤ 0.001. Differentially expressed genes were also identified based on survivorship, where the percent survival of each plot was used to bin the plot into one of high, medium, or low survival based on the fourth quartile, second and third quartiles, or the first quartile of percent survival, respectively. Statistical significance was conducted as above but using contrasts between each survival group. All other analyses and plots were conducted in R and are available in the repository listed above.
3 RESULTS
3.1 Clam growth and survival
There were no significant differences between CG and Ref beaches in any measured growth or survival variable (ANOVAs, p ≥ 0.168, Table 1). The mean (± SD) initial clam wet weight and shell height for CG and Ref beaches (N = 180) was 1.5 ± 0.2 g and 1.8 ± 0.3 g and 1.5 ± 0.1 cm and 1.6 ± 0.1 cm, respectively (Table 1). After 16 weeks in situ, these increased to 3.9 ± 1.3 g and 3.8 ± 2.9 g and 1.8 ± 0.3 cm and 1.9 ± 0.4 cm, which in terms of height, was an increase in growth of 20.5 ± 15.3% and 18.0 ± 22.6%, respectively. Calculating percent growth using height includes both the alive and dead clams and is therefore considered to be a more accurate measurement of growth compared to using wet weights. Wet weights only include surviving clams, and therefore does not capture the reduced growth that may have occurred in clams that eventually died. Clam survival was 63.3 ± 28.3% in CG and 58.3 ± 37.0% in Ref beaches, respectively.
Average (± SD) clam size/survival and sediment characteristic variables for clam garden (CG) and reference (Ref) beaches and associated linear model p-values (with beach location as a nested effect). No variables were significantly affected by the CG/Ref factor or showed a significant deviation from normality as measured by Shapiro Wilk test.
Beach location (independent of the CG/Ref factor), however, significantly affected all growth and survival variables (Table 2, SI Figure S1). An unplanned difference occurred in the initial size of clams among the six beaches (initial wet weight and shell height ANOVAs p = 0.031 and p = 0.044, respectively, Table 2, SI Figure S2). This difference was specifically due to significantly larger clams inadvertently planted on beach D (Table 2, SI Figure S2). Percent growth was highest on beaches A (31.2 ± 7.2%), C (29.7 ± 2.8%), and D (47.0 ± 8.2%) and lowest in beaches B (6.6 ± 6.8%), E (0.8 ± 0.9%), and F (0.3 ± 0.6%) (Table 2), while percent survival was highest in beaches A (88.3 ± 12.6%), B (71.7 ± 27.5%), C (68.3 ± 12.6%), and D (86.7 ± 5.8%) and lowest in beaches E (33.3 ± 23.6%) and F (16.7 ± 24.7%) (Table 2).
Average (± SD) clam size/survival and sediment characteristic variables for each beach and associated linear model p-values.
Gross observations showed that most of the survivors on beaches A (CG), B (Ref), C (CG), and D (Ref) were considered healthy (Table 3). These beaches had 68–88% survival, with the percentage of clams considered weak (0–5%), emaciated (0–5%), or having very pale digestive glands (0–22%) being generally low, with the exception of beach D, where 22% of clams had pale digestive glands (Table 3). In contrast, survivors from beaches E (CG) and F (Ref) had only 17–33% survival, and had more clams considered weak (13–15%), emaciated (15–22%), and with digestive glands being very pale (13–22%) than the other beaches. Beaches E and F are therefore considered poorer performing sites.
Gross observations (health, condition and digestive gland coloration) of surviving clams in each beach (A-F).
3.2 Beach sediment characteristics
There were no significant differences between CG and Ref beaches for any of the sediment characteristics (ANOVAs, p ≥ 0.392, Table 1). Beach location (independent of the CG/Ref factor), however, significantly affected all sediment characteristics except percentage of small rocks (Table 2). A PCA of these sediment characteristics separates the beach locations into three groups (Figure 2), where pairs of beaches are clustered based on geographical locations (Figure 1). Beach A (CG), on a west-facing bay, was exposed to wave action from within and outside Kanish Bay. Beach A grouped with beach B (Ref), an east-facing bay that was also exposed to wave action within Kanish Bay (Figure 1). These beaches are the two sites that are closest to, but on opposite sides of, the entrance of Kanish Bay and show correlation with coarse sand and sand (Figure 2). Beach C (CG) is a south-facing sheltered beach within Small Inlet and groups with beach D (Ref), a north-facing beach on the opposite side of Small Inlet. Beaches C and D correlated with rocks, fine sand, and final weight/shell height (Figure 2). Beach E (CG) and F (Ref), two proximate beaches on the south side of Kanish Bay, face north and are exposed to wave action. These sites correlated with carbonates, organics, and small rocks, and are opposite to percent survival (Figure 2). Notably, these beach groupings remain constant when including only abiotic characteristics in the PCA or when including only survival and growth variable (data not shown). The PCA was also run on the data but excluding beach D (due to the aforementioned initial size difference of this beach), and this showed similar associations and groupings, although beach C grouping on its own (SI Figure S3).

Principal component analysis biplot showing beach groupings including beach D and correlations in terms of percent survival and growth of Pacific littleneck clams and various beach sediment characteristics. Each beach is shown as A–F (as per Figure 1 locations) and plots within beach location as 1–3. Colours of plots and ellipses are for visualization purposes only.
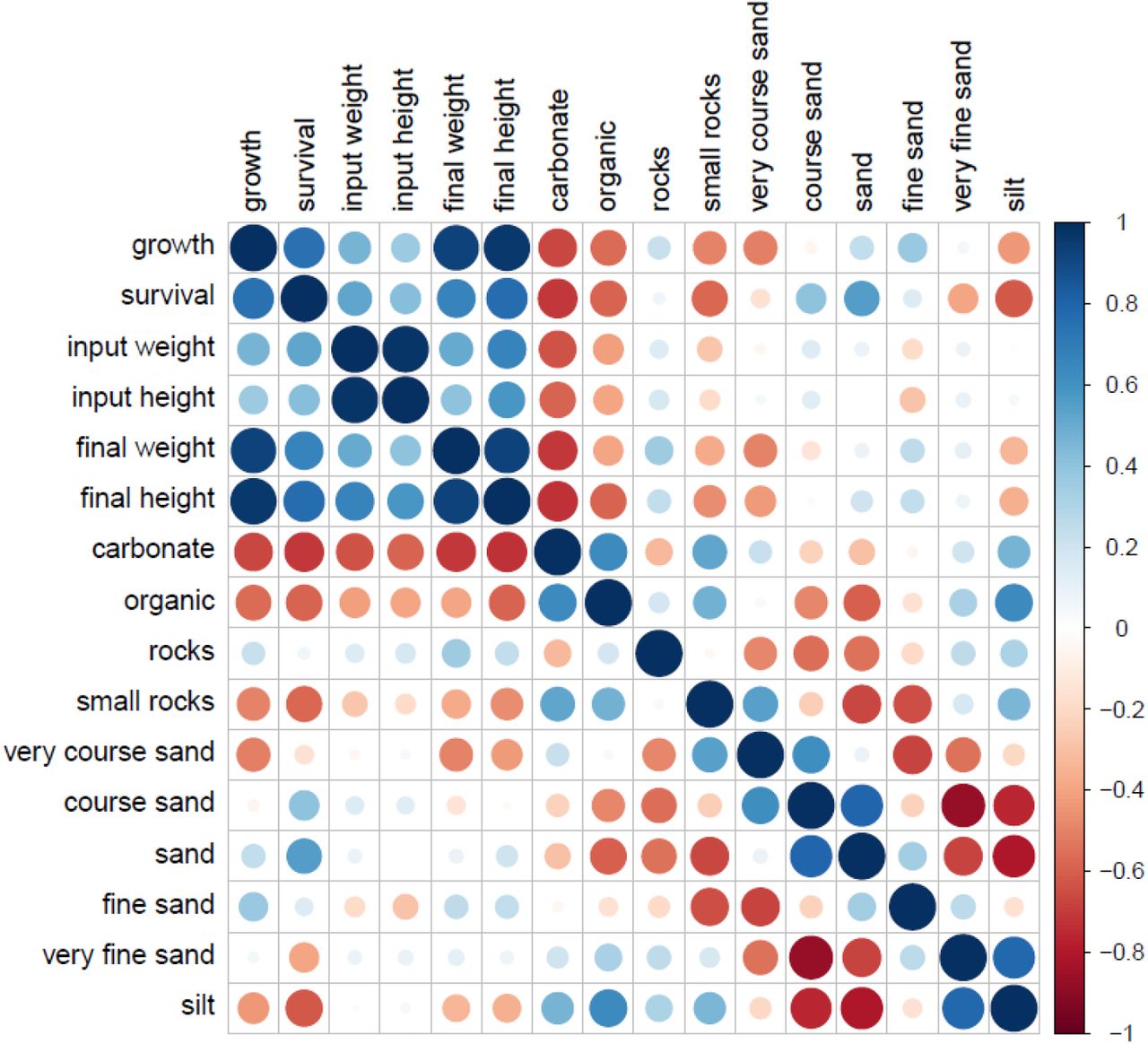
Most sand types, silt, and rocks were either positively or negatively correlated with growth and survival variables (SI Figure S4). To investigate correlations between sediment characteristics and clam growth and survival further, beach D was again excluded as described above (SI Figure S5). Abiotic variables with significant positive associations with growth included sand (p = 0.018, R2 = 0.31), and fine sand (p = 0.022, R2 = 0.29). Negative associations with growth were silt (p = 0.002, R2 = 0.49), and small rocks (p = 0.014, R2 = 0.34). Variables with significant positive associations with survival were coarse sand (p = 0.014, R2 = 0.34), and sand (p = 0.002, R2 = 0.48). Negative associations with survival included carbonates (p = 0.008, R2 = 0.39), organics (p = 0.035, R2 = 0.24), as well as small rocks (p = 0.021, R2 = 0.30), very fine sand (p = 0.002, R2 = 0.49), and silt (p = 0.002, R2 =0.51) (SD Additional File S1, without Beach D)..
3.3 Transcriptomic overview and de novo reference transcriptome
The gill and DG libraries produced on average (± SD) 53.8 ± 10.7 and 55.0 ± 8.3 M read pairs per library respectively. All 28 libraries, comprising 305 GB of high-quality sequence data, were used to create a de novo transcriptome assembly (Table 4; see Data Availability). This assembly was comprised of 50,000 contigs (i.e., putative transcripts), of which 42,708 were given annotation by sequence similarity to the SwissProt database.
Overview of RNA-seq reads and quality scores. Totals that include both reads in the pair are indicated by (R1+R2). Values are shown in billion (B) or million (M).
Alignments of the quality-controlled reads resulted in a mean (± SD) overall alignment rate of 27.87 ± 2.48% against the Pacific littleneck clam de novo transcriptome. For comparison, the alignment rate against the Manila clam (i.e., the closest relative with a reference genome) resulted in very low alignments of 1.10 ± 0.17% (gill), 2.59 ± 1.00% (DG). All downstream analyses used the de novo transcriptome quantified transcripts. After filtering the dataset containing all 28 samples, 33,825 transcripts (67.7%) were expressed in at least five samples above the applied threshold (see Materials and Methods).
An MDS plot of samples from both tissue types showed that gill samples were distinctly separate from DG samples across the first dimension, explaining 65% of the variation in the dataset (Figure 3). The second dimension, which explained only 3% of the variation separated the DG samples, but not the gill samples. One outlier sample was observed in the gill (P12).

All sample MDS plot showing groupings of gill (G) and digestive gland (DG) tissues of Pacific littleneck clams sampled from plots P1–P18 (i.e., Beach A: P1–P3; Beach B: P10–P12; Beach C: P4–P6; Beach D: P7–P9; Beach E: P15; Beach F: P18).
3.4 Tissue-specific expression
Samples from each tissue were then analyzed separately after conducting tissue-specific filtration and normalization. The gill and DG samples on average (± SD) had 12.9 ± 3.0 M and 15.5 ± 2.9 M reads aligning per library and the datasets were found to have 28,391 and 24,699 transcripts expressed, respectively, after the low expression filter. Inspecting transcripts that were present in only one of the tissue types identified 8,795 (31.0%) gill-specific transcripts and 5,103 (20.7%) DG-specific transcripts (Table 5; SD Additional File S2). The highest expressed tissue-specific transcripts are shown in SI Figure S6 (n = 200 shown for each tissue).
Differentially expressed gene overview comparing beach type (clam garden (CG) vs. reference beach (Ref)), beach survival (high vs. medium vs. low), tissue type (gill vs. digestive gland), or expressed in background. Note that here differential expression is considered when p ≤ 0.001 and log2 fold-change > 0.58 (1.5-fold overexpressed) or < -0.58 (1.5-fold underexpressed).
Tissue-specific transcripts were compared to all expressed genes in that tissue to identify enriched GO categories in the tissue-specific list. Gill-specific transcripts were enriched for biological adhesion (N = 496), ion transport (N = 494), signal transduction (N = 1,184), response to stimulus (N = 1,839), G-protein coupled receptor signalling pathways (N = 210), and defense response (N = 402, p < 0.0001), among others (SD Additional File S3). DG-specific transcripts were enriched for biological adhesion (N = 308), defense response (N = 263), immune response (N = 223), innate immune response (N = 149), humoral immune response (N = 58), organonitrogen compound catabolic process (N = 130), defense response to bacteria (N= 82), and ion transport (N = 201, p < 0.0001), among others (SD Additional File S3).
3.5 Effect of clam gardens on gene expression
The gill dataset analyzed by multidimensional scaling (MDS) has 13% and 11% of the total variation explained by dimensions 1 and 2, respectively (Figure 4A). In general, in the gill samples, no clear clustering was observed by beach type (i.e., CG vs. Ref) nor any other variables (e.g., survival, sand, silt, or carbonates). There were two outlier samples (P12 from beach B and P7 from beach D) in the MDS plot, both of which were Ref beaches and had 45% and 80% survival, respectively.

MDS plots for (A) gill and (B) digestive gland tissues of Pacific littleneck clams where each datapoint is plotted as clam garden (CG) or reference beach (Ref) with the following information: % survival, % carbonates, beach location identifier (A—E), and plot identifier (P1–P18) (i.e., Beach A: P1– P3; Beach B: P10–P12; Beach C: P4–P6; Beach D: P7–P9; Beach E: P15; Beach F: P18).
The DG dataset analyzed by MDS resulted in 14% and 9% of the total variation being explained by dimensions 1 and 2, respectively (Figure 4B). Some grouping was observed along dimension 2 with CG samples clustering together, except for sample P15 from beach E. Ref samples were more spread out along both axes. A gradient was observed across dimension 1 along survival, with high survival being positioned in negative dimension 1, medium survival near 0-1, and low survival in positive dimension 1, although the trend was not universal for all samples. Percent carbonate, which was correlated with survival (see above), also trended along dimension 1 in the opposite direction to survival (i.e., high survival was associated with low percent carbonate, SI Figure S7).
Differential expression between CG and Ref was evaluated for each tissue separately (Table 5). The gill had 56 transcripts differentially expressed as a result of CG (FC > 1.5, p ≤ 0.001, no multiple test correction (MTC)) (see Discussion; SD Additional File S2). Of these transcripts, 35 had identifiers recognized by DAVID Bioinformatics, but no Gene Ontology (GO) enrichment was identified (p > 0.01). Most of these transcripts were overexpressed in the CG samples (N = 42), including several transcripts associated with immune/apoptotic or stress-related functions—two transcripts annotated as tumour necrosis factor (TNF)-related (FC > 1.5), one as death effector domain (DED)-related (i.e., top overexpressed transcript, FC > 71.5), and universal stress protein A-like protein (FC > 4.7). In total, 14 transcripts were underexpressed in the CG, including a heat shock 70 kDa protein 12a (FC = 17.9). A full list of differentially expressed genes is available in SD Additional File S2.
The DG had 37 transcripts differentially expressed between CG and Ref samples. This included 11 overexpressed and 26 underexpressed transcripts in CG (SD Additional File S2). Using the full, bidirectional list of differentially expressed transcripts, several enriched GO categories were observed (SD Additional File S3), including biological process categories iron ion homeostasis (N = 3, p = 0.008) and negative regulation of hydrolase activity (N = 5, p < 0.001). The highest overexpressed transcript in the CG samples was replicase polyprotein annotated from the cricket paralysis virus (FC > 500), which suggests that viral proteins were also captured in the study and that they were differentially abundant between beach types. Multiple complement-, iron-, and protein-folding-related transcripts were observed in the DG, including overexpression of a transcript annotated as c1q domain (2.3-fold), thrombospondin-1 (2-fold), soma ferritin, and cytochrome P450 (FC > 1.5). Transcripts under-expressed in the DG included interferon-induced very large GTPase 1 (FC > 8.5), apoptosis inhibitor IAP (4.3-fold), low affinity immunoglobulin epsilon Fc receptor (FC > 2), c-type mannose receptor 2, and platelet endothelial aggregation receptor 1 (FC > 1.5), among others (SD Additional File S2).
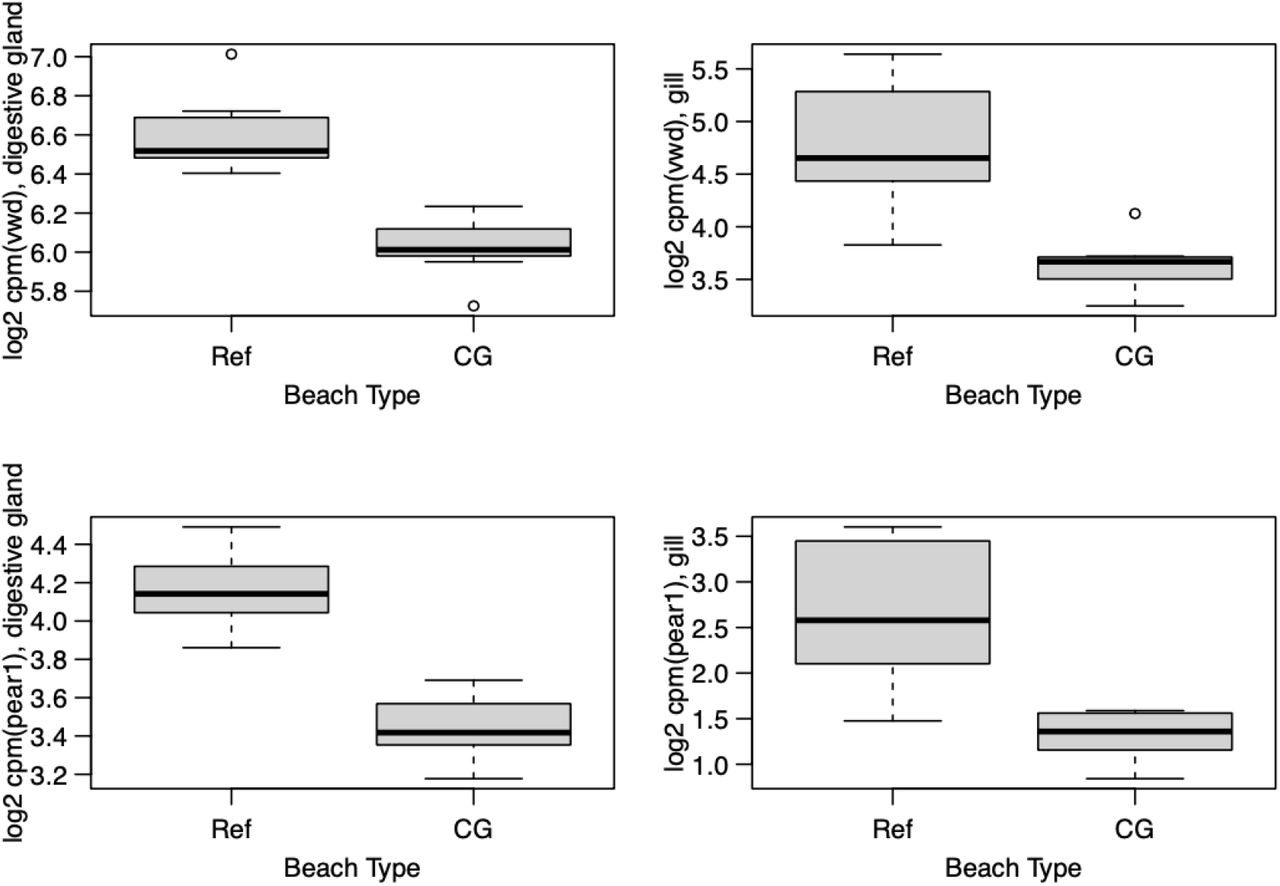
Consistency was observed between the tissues in the CG response, where two of the 91 unique differentially expressed transcripts were differentially expressed in both tissues. This is notable given that the datasets were normalized, filtered, and analyzed separately. This included the under expression in the CG samples of a transcript annotated as von Willebrand factor type D domain (vwd) and of a transcript annotated as platelet endothelial aggregation receptor 1 (pear1; Figure 5) in both tissues.

Box plots showing expression of genes of interest identified in both digestive glands and gills of Pacific littleneck clams, which were underexpressed in clam gardens (CG) relative to reference beaches (Ref). Vwd = von Willebrand factor type D domain; pear1 = platelet endothelial aggregation receptor 1.
3.6 Gene expression responses and differential survival
Given the differential survival observed among the beaches and plots, genes correlating with survival were investigated to determine what might be driving these differences and to potentially identify genes that are related to survival or mortality in this species. Genes were of particular interest that were differentially expressed between low and medium survival, between medium and high survival, or incrementally differentially expressed at each step between high, medium, and low survival (see Materials and Methods for beach survivorship classification).
The majority of differentially expressed genes by survival in the gill were overexpressed between the low and medium survival groups, rather than between medium and high survival (Table 5). Transcripts overexpressed in the low survival group relative to the medium survival group (N = 26) were involved in functions such as heat-shock, interferon response, and ligase activity (SD Additional Files S2, S3). For example, heat-shock protein (hsp) 90 (FC > 60) and three different transcripts annotated as subunits of hsp70 (FC > 2) were overexpressed specifically in the low survival group. Transcripts overexpressed in the medium survival group relative to the high survival group included six transcripts of various functions (SD Additional File S2). Fewer transcripts were overexpressed in the higher survival group (i.e., high vs. medium, N = 11; medium vs. low, N = 14) and these generally were involved in immune functions, but also various other functions. Overexpression of toll-like receptor 1 (FC > 2) and complement C1q-like protein 4 (FC > 3.5) was observed in medium survivors relative to low survivors.
Similar to the gill, in the DG the majority of the differentially expressed transcripts by survival were overexpressed in low relative to medium surviving clams (N = 34) and were generally involved in immune system or heat-shock or transport. Transcripts annotated as hsp70 12a and 12b, as well as several aminopeptidases, were overexpressed in low relative to medium and in medium relative to high survival (Figure 6). All the overexpressed genes in medium relative to high surviving clams (N = 18) were also overexpressed in the low relative to medium survival group comparison. Very few genes were overexpressed in the high survival clams relative to medium (N = 4) or the medium survival clams relative to low survival (N = 8), and these transcripts had various functions, including a transcript annotated as mucin-2 protein (Figure 6).

Linear regression of differential expression of selected genes of interest versus percent survival in Pacific littleneck clams. Transcript expression and their correlation to survival include (top left to bottom right) heat shock protein family A member 12A (hsp70-12a), aminopeptidase, and mucin-2 in the digestive gland (DG) and hsp70-12a, hsp90, and toll-like receptor 1 in the gill.
4. DISCUSSION
4.1 Effects of clam gardens on clam and beach characteristics
Results from the present study showed that there was no significantly-enhanced growth or survival on unmaintained clam garden (CG) versus reference (Ref) non-walled beaches. Using similarly-sized clams that were collected on the same days and at the same location as the present work Salter (2018) also reported no significantly enhanced growth or survival on unmaintained CG compared to non-walled beaches (controls) in Kanish and Waiatt Bays. It should be noted that the clam gardens in the present study were not being maintained according to traditional tending practices and this maintenance may influence clam survival, growth, and transcriptomic profile, as well as various abiotic characteristics (i.e., sediment grain size, carbonate content, organic content). Salter (2018), however, also included some CG traditional tending practices (i.e., adding a 1 cm-thick layer of crushed shell mixture three times throughout the summer) on replicate treatment plots on both beach types and reported a positive effect of the shell hash treatment on growth and survival in both CG and non-walled beaches. Therefore, both the present study and Salter (2018) did not observe a positive effect of CG on growth and survival in the absence of tending but Salter (2018) reported a positive effect with the addition of crushed shell hash in both beach types.
Some trends were observed in the transcriptomic data that suggest that the CG had a transcriptomic effect on the clams, albeit not a strong effect. Most notably, consistent genes were identified in two tissues even though very few genes were found to be differentially expressed, and the CG and Ref samples showed a slight separation in the PCA of DG tissue. The two genes that were identified as underexpressed in the CG in both tissues were annotated as von Willebrand factor type D domain (vwd) and platelet endothelial aggregation receptor 1 (pear1). The former is present in many proteins, including mucins and other extracellular glycoproteins, with one in particular being reported to be up-regulated in response to heat shock in the Pacific oyster, Crassostrea gigas (Zhang et al. 2015). The function of the latter in shellfish remains unknown; in vertebrates it is a cell membrane protein involved in platelet aggregation. Homologous genes of platelet activation are present in the Hong Kong oyster (Crassostrea hongkongensis) and are regulated in response to hyposalinity (Xiao et al. 2018). In terms of the other genes differentially expressed by clams in CG, the general functions were not clear. Immune genes and stress-response genes were found both over- and under-expressed in CG versus Ref (e.g., a heat-shock protein transcript was found to be underexpressed in the CG and a universal stress protein-annotated transcript was overexpressed in the CG). In general, the functions of these transcripts in the Pacific littleneck clam responding to the CG factor will be useful to continue to characterize as the genomic information develops for this species and additional transcriptomic studies are conducted in relation to environmental stress and maintained or unmaintained clam gardens. These molecular phenotypes may indicate a response to the CG from the 16-week in situ study period that is below the level of growth and survival phenotypic responses. Further work with longer study periods would be required to determine whether these transcriptome signatures precede macro-phenotypic responses.
Sediment grain sizes, percent carbonate content, and percent organic matter did not differ between CG and Ref in the present study, although there was a significant level of variation with these factors within both beach types and among locations. For example, carbonate content varied from 4 to 10% in CG and from 2 to 15% in Ref beaches, while silt varied from 0 to 4% and from 1 to 5%, respectively. This is somewhat contrary to Salter’s (2018) study that in untreated plots found that CG contained 2.8 – 12.9 times more carbonate and a smaller percentage of silt than non-walled beaches (Salter 2018). The reason for this discrepancy is not known and would be valuable to address with future studies. The lack of differences observed in the present study in growth and survival, as well as only a minor effect being observed in the clam molecular phenotypes, from being grown in CG relative to Ref beaches, may in part be due to the lack of tending of the clam gardens, as is traditionally conducted. Future studies would benefit from fully examining the impacts of tending practices on sediment characteristics alongside clam growth and survival, which would involve clam garden beach restoration and active maintenance for long periods, with further analysis of impacts on shellfish molecular phenotypes and productivity.
4.2 Abiotic variation among beach location and effects on clam growth and survival
Across the six sampling locations, beaches differed in most of the variables examined (i.e., biotic response variables: growth, survival; abiotic characteristics: organic level, carbonate concentration, grain sizes). Beaches clustered based on these sediment characteristics and differential growth and survival were noted among the locations. Given these trends, abiotic factors potentially involved in survival as well as molecular responses to survival were characterized.
The sedimentary environment is very important for clam growth and survival (Joo et al. 2021). Littleneck clams typically live as infaunal burrowers between 5 and 15 cm depth, in soft sand or sandy mud in the intertidal zone (Lazo 2004). In the present study, growth was affected by small rocks, sand, fine sand and silt. Survival was affected by carbonates, organics, small rocks, coarse sand, sand, very fine sand, and silt. Joo et al. (2021) also found that grain size and sorting of the sediment determined the amount of pore-water dissolved oxygen and organic matter, both affecting growth and survival of juvenile Manila clams, with optimal survival and growth linked to poorly-sorted sediment with an average grain size of medium sand (180 – 335 µm). Similarly, the grain size of the sand in the present study, which had a significant positive effect on survival, was 250 – 500 µm. Salter (2018) found increased water residency time positively affected clam biomass and growth and reported a weak negative effect of silt on clam density and biomass in bivalve surveys and contrary to the present study also found a positive effect of increased sediment silt content on clam biomass, in clam transplant experiments. An early multi-year study looking at environmental factors affecting growth in the hard clam Venus mercenaria (now Mercenaria mercenaria) found a consistent decrease in growth with increasing sediment silt-clay content and (Pratt and Campbell 1956). All their results showed a negative relationship between growth and fineness of the sediment (Pratt and Campbell 1956). Permeability of the sediment is an important factor for burrowing filter feeders’ pumping activity, which is essential for respiration, excretion, and nutrition and Pratt et al. (1956) found permeability to be an inverse function of the sediment silt-clay content. In addition, commercial Manila clam beds studied in Spain had higher survival of clams planted in sand-gravel than in those placed in mud (Cigarria and Fernandez 2000).
Sediment carbonate content had a significant negative effect on clam survival in the present study, where sediment with more than 8% carbonates had markedly decreased growth and survival. Similarly, Munroe (2016) found that the average daily growth of early post-settled Manila clams over two years was negatively correlated with sediment carbonate content (Munroe 2016). This is not a universally observed trend in the literature (Groesbeck et al. 2014, Salter 2018) and counters expectations given the expected importance of carbonate for clam shell growth and early recruitment. The addition of crushed shell hash alters sediment calcium-carbonate saturation states, as it increases surface sediment porewater aragonite content and pH (Green et al. 2009). For instance, there was a three-fold increase in hard clam burrowing and recruitment by adding crushed shell hash to a final concentration of 8% of wet sediment weight (Green et al. 2012). Salter (2018) increased carbonate content significantly (1.9–7.2 times) by addition of shell hash, which increased clam growth and tissue biomass in both clam garden and control beaches after six months (Salter 2018).
Crushed shell hash also changes the physical structure of the sediment as, like gravel, it can create interstitial spaces in the sediment, thereby increasing pore-water dissolved oxygen, organic-matter content, and substrate stability, as well as providing protection from predators for juvenile clams (Joo et al. 2021, Sponaugle and Lawton 1990, Thompson 1995). Thus, there may be potentially multiple benefits of adding shell material for bivalve growth/survival that are not linked to carbonate concentration. The effect of shell hash or crushed shell on bivalve growth and survival is likely dependent on a wide variety of factors including amount of shell, size and compactness of shell particles, species generating the shell hash, species/size of bivalve grown, stocking densities, tidal height, temperature, and types/densities of predators. For example, the application of very finely crushed shell hash could have a negative effect by decreasing interstitial spaces in the sediment and thus, as seen with silt, decreasing sediment permeability. Recent studies investigating the efficacy of shell hash to mitigate acidification of intertidal sediments found it unsuccessful at raising pore-water pH of acidic sediments (Beal et al. 2020, Doyle and Bendell 2022). Doyle et al. (2022) found the addition of shell hash to be site dependent as it reduced variation in pH at an intertidal site with pH 8.03 but did not mitigate the pH of mud flats with pH 7.59. At this point it appears that more research is required to comprehensively understand the effects of shell hash on bivalve growth and survival.
4.3 Pacific littleneck clam transcriptomics
In non-model organisms, such as the Pacific littleneck clam, advances in molecular ecological and physiological understanding of the species can be advanced through transcriptomic tools and the results these tools generate. Here it was clear that the reference transcriptome generated de novo was necessary for the study, given that the closest relative genome (i.e., Manila clam) resulted in a very low alignment rate from the Pacific littleneck clam reads. Further, the use of different tissues in the present study revealed high levels of tissue-specific expression (i.e., 21-31% of the transcripts expressed in the tissue). As multiple individuals were included in each sample (i.e., pools), the true number of individuals sampled for the tissue-specific expression analysis is greater than the number of libraries, increasing the reliability of the tissue-specific genes. The tissue-specific expression analysis indicated that for both tissues, tissue-specific transcripts were involved in signal transduction, defense and immune response, and ion transport, among other processes. Therefore, although the specific transcripts are different, similar functions are enriched in the tissue-specific gene lists for both the gill and the DG, although exceptions exist.
The presence of differential survival among the locations provided a valuable opportunity for analyzing transcripts that are associated with mortality/survival. These characterizations, over time through multiple transcriptome studies generating more evidence, can begin to characterize specific transcripts that are involved with specific stress types (Evans et al. 2011, Miller et al. 2017, Sutherland et al. 2012). This can enable the development of Ecological Association Ontology data collection for non-model species (Pavey et al. 2012) and in the future may be used to characterize underlying factors associated with observed mortality or morbidity (e.g., abiotic stress compared to bacterial or viral infection). Although the Pacific littleneck clam remains largely uncharacterized at the molecular level, some of the genes identified in the present study correlated with survival, including differential expression in the gill and DG of heat-shock proteins, immune response genes, and other functional categories. Additional transcriptomic studies on the species in different environmental conditions over different temporal and spatial scales should help to improve these categorizations and identify stressor-specific or generalized response genes.
Differential expression was observed in transcripts involved in immune defense. For example, transcripts annotated as complement C1q protein complex or involved in cell adhesion and oxidation-reduction processes had high expression in high surviving beaches and may indicate increased activity of immune defense responses. Complement C1q proteins are an important component of the innate immune system (Zhang et al. 2014) and are upregulated in response to bacterial challenges (McDowell et al. 2014) and salinity stress (Zhao et al. 2012). Cell adhesions are critical for the development, maintenance, and function of multicellular organisms and are essential for invertebrate immunity (Johansson 1999).
In the present study, potentially due to the relatively low sample size from pooling individuals and due to having fewer plots with a sufficient number of survivors at some beaches and the high number of expressed transcripts, it was not possible to apply multiple test corrections. Therefore, a more stringent p-value was applied (p < 0.001), but the associations of genes to CG or differential survival should be taken with caution. Further studies confirming these functions will be beneficial to understand their role in clam transcriptomics.
4.4 Viral presence and clam health
The use of RNA-sequencing for the present analysis identified a differentially expressed virus transcript in the CG versus Ref samples, which could have unexpected effects on either gene expression activity or phenotypic responses, as could any other uncharacterized factor influencing clam growth/physiology (e.g., temperature, beach exposure). The top overexpressed transcript in the DG from CG was a replicase polyprotein from the cricket paralysis virus (CPV), which is a positive-sense, single-stranded RNA virus from the family Dicistroviridae in the order Picornavirales. Members of the Dicistroviridae family are widely distributed in nature with the broadest host range of all small RNA viruses in invertebrates (Bonning 2009). As the Dicistroviridae viral sequences isolated in the present study ranged in size from 1479 to 7371 nucleotides, one can deduce that they represent viral mRNA, instead of viral genomes. Some dicistroviruses can be non-pathogenic or cause subtle impacts (reduced longevity and fecundity), while others result in rapid paralysis (Bonning 2009).
Even though there was transcriptomic evidence of viral presence in all the beaches, most of the Pacific littleneck clams from the medium- and high-survival groups did not exhibit any signs of pathophysiology. Other clams (mostly from the low-survival group), however, were emaciated or watery and had slightly to extremely pale-coloured digestive glands, an indicator of reduced feeding and potentially compromised function and health. Pale digestive glands could also be the result of paramyxean parasites responsible for disease in marine mollusks, with infection being linked to digestive tropism interfering with food adsorption and causing pale-yellowish digestive glands with thin, watery flesh in both mussels and oysters (Alfjorden 2017, Carella et al. 2011). In the present study, however, there was no significant parasite gene expression at 16 weeks and histopathology would be needed to confirm any paramyxean parasite infections. It is also worth noting that the single time point of sample collection in the present study and the analysis of only surviving clams may have reduced the likelihood of viewing viral or parasitic gene expression in these samples.
Although very pale digestive glands and emaciation was observed in clams from beaches with lower growth and survival, histopathology or genetic identification would again be required to confirm any viral gene expression linkages to observed impacts on organismal health. Health status is most likely linked to multiple stressors comprised of abiotic (e.g., temperature, sediment composition, water residency) and biotic factors (e.g., bacteria, viruses, harmful algae). RNA sequencing combined with de novo transcriptome assembly revolutionizes virus detection and allows for the screening of a broad range of symptomatic and asymptomatic virus species in a non-discriminatory manner, thereby displaying a broader picture of the environmental factors present (Nagano et al. 2015). The current dataset has yet to be fully characterized for viral sequences and could be mined further at a later date, particularly as more resources are developed for this species.
CONCLUSIONS
The de novo assembly of a Pacific littleneck clam reference transcriptome enabled the detection of a small but significant differentially expression response in clams from CG and Ref beaches. This was observed even though no significant phenotypic differences in growth or survival were observed, nor differences in sediment characteristics between the two unmaintained clam gardens or reference sites. However, significant phenotypic differences in growth and survival and in sediment characteristics between beaches allowed for the discovery of several sediment characteristics significantly correlated with growth and survival. The unexpected result of carbonates having a negative effect on survival may partially be due to the lack of clam garden tending practices and future work would benefit from examining the effects these practices and shell-hash have on bivalve health. Transcripts related to stress response were identified by conducting differential expression analysis among beaches of largely differing survival. The characterized transcriptome provides a valuable resource for future transcriptomic studies investigating genetic and phenotypic responses of clams in different habitats and environmental conditions.
DATA AVAILABILITY
Supplemental materials, including all abiotic and phenotypic site metadata needed for data analysis, transcript annotation, quantified gene expression counts, sample phenotypes:
Reference transcriptome: https://doi.org/10.6084/m9.figshare.19735753
RNA-seq bioinformatics pipeline: https://github.com/bensutherland/Simple_reads_to_counts
RNA-seq analysis and physiological/abiotic analysis pipeline: https://github.com/raapm/ms_clam_garden
SUPPLEMENTAL DATA
Additional File S1. Abiotic variables with survival and growth linear model p and R2 values, including and excluding beach D.
Additional File S2. Differentially expressed genes and background gene lists for both tissues in Pacific littleneck clams. Gene lists include clam garden vs. reference, survival groups, and tissue-specific expression.
Additional File S3. Gene Ontology enrichment results for the clam garden analysis, the survival analysis, and the tissue-specific expression.
SUPPORTING INFORMATION
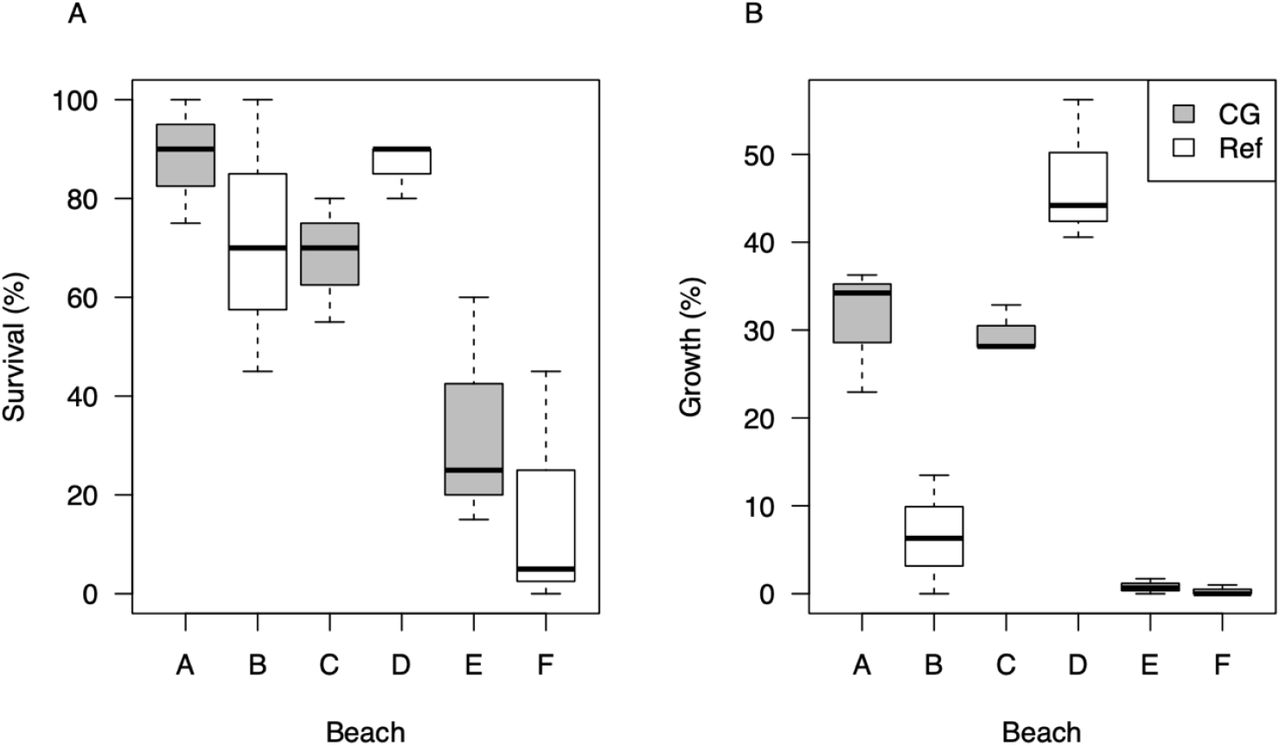
Boxplots of (A) percent survival and (B) percent growth of Pacific littleneck clams by beach (A–F).

Boxplots of (A) input weight and (B) input height of Pacific littleneck clams by beach (A– F).

Principal component analysis biplot showing beach groupings and correlations in terms of survival and growth of Pacific littleneck clams and various beach sediment characteristics excluding beach D. Each beach plot is shown as A-F (as per Figure 1 locations) and 1-3 depending on the plot within the beach location. Colours of plots and ellipses are for visualization purposes only.

Pearson correlations between biotic and abiotic variables including growth and survival including beach D.

Pearson correlations between biotic and abiotic variables including growth and survival of Pacific littleneck clams excluding beach D.

Highest expressed tissue-specific transcripts in the gill (G) and digestive gland (DG) tissues of Pacific littleneck clams within each plot (P1–P18) (i.e., Beach A: P1–P3; Beach B: P10–P12; Beach C: P4–P6; Beach D: P7–P9; Beach E: P15; Beach F: P18). Samples were clustered based on expression of these transcripts, with similarly expressing sample grouping. The higher the expression, the darker the colour of the cell, with each row as an individual transcript.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Percent survival of Pacific littleneck clams versus percent carbonate in sediments. Beach D is included in this figure, and when it is not included, the adjusted R2 = 0.39 (p = 0.008), see SD Additional File S1.
ACKNOWLEDGEMENTS
Funding for this study was provided by the Hakai Institute. We acknowledge the Heiltsuk Nation and the Wuikinuxv Nation, which have stewarded and protected the territory we work within since time immemorial. We thank the Hakai Institute, the Tula Foundation, and the Clam Garden Network for facilitating the research and for their continued support. We also thank Caitlin Smith, Morgan Black, Kayla Balmer, Kayla Long, and Kieran Cox for their invaluable help in the field, Dr. Eric Rondeau for his assistance in the laboratory, and Allie Byrne for creating Figure 1, showing the location of the experimental sites.
Footnotes
Data Deposition: Raw RNA-sequencing data have been uploaded to SRA under BioProject PRJNA818991, BioSamples SAMN26893882–SAMN26893909.
REFERENCES




